
Sir4 Deficiency Reverses Cell Senescence by Sub-Telomere Recombination



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Strains, Oligonucleotides, and Genetic Manipulations
2.2. Sporulation and Micromanipulation
2.3. Telomere Length and Y′ Element Analyses by Southern Blot
2.4. Cell Senescence Analysis
2.5. Quantitative Analysis of Genomic Y′ Element Copy Numbers
2.6. Statistical Significance Calculation
3. Results
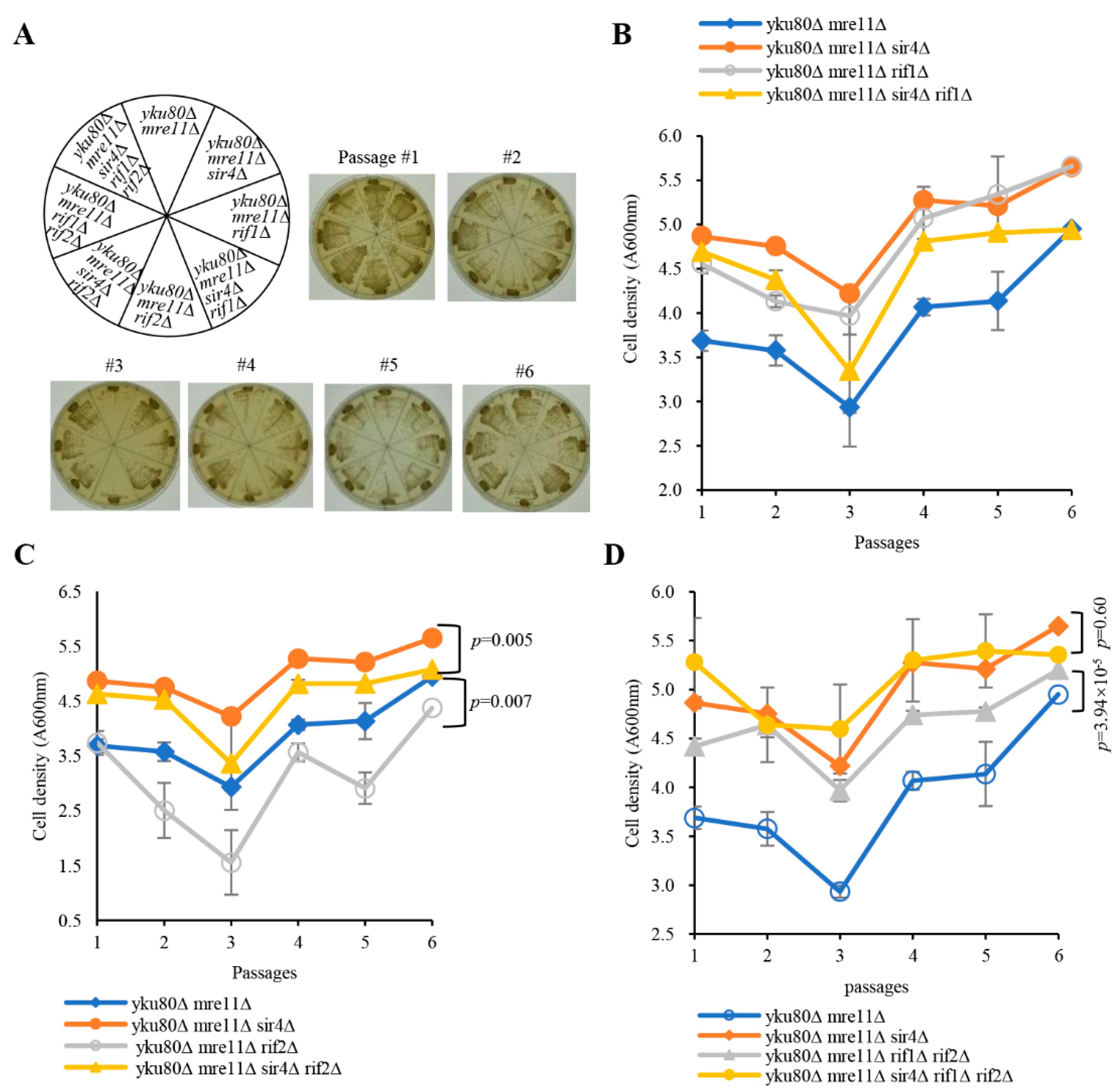
3.1. Sir3/4, But Not Sir2, Deficiency Rescues Senescence
3.2. Rad51 Is Required for Senescence Evasion Induced by Sir4 Deficiency
3.3. Rif1, Rif1/2, But Not Rif2 Deficiency Suppresses Senescence
3.4. Rif1 and Rif1/2 Deficiencies Lengthen Telomeres of yku80∆ mre11∆ and yku80∆ mre11∆ sir4∆ Cells
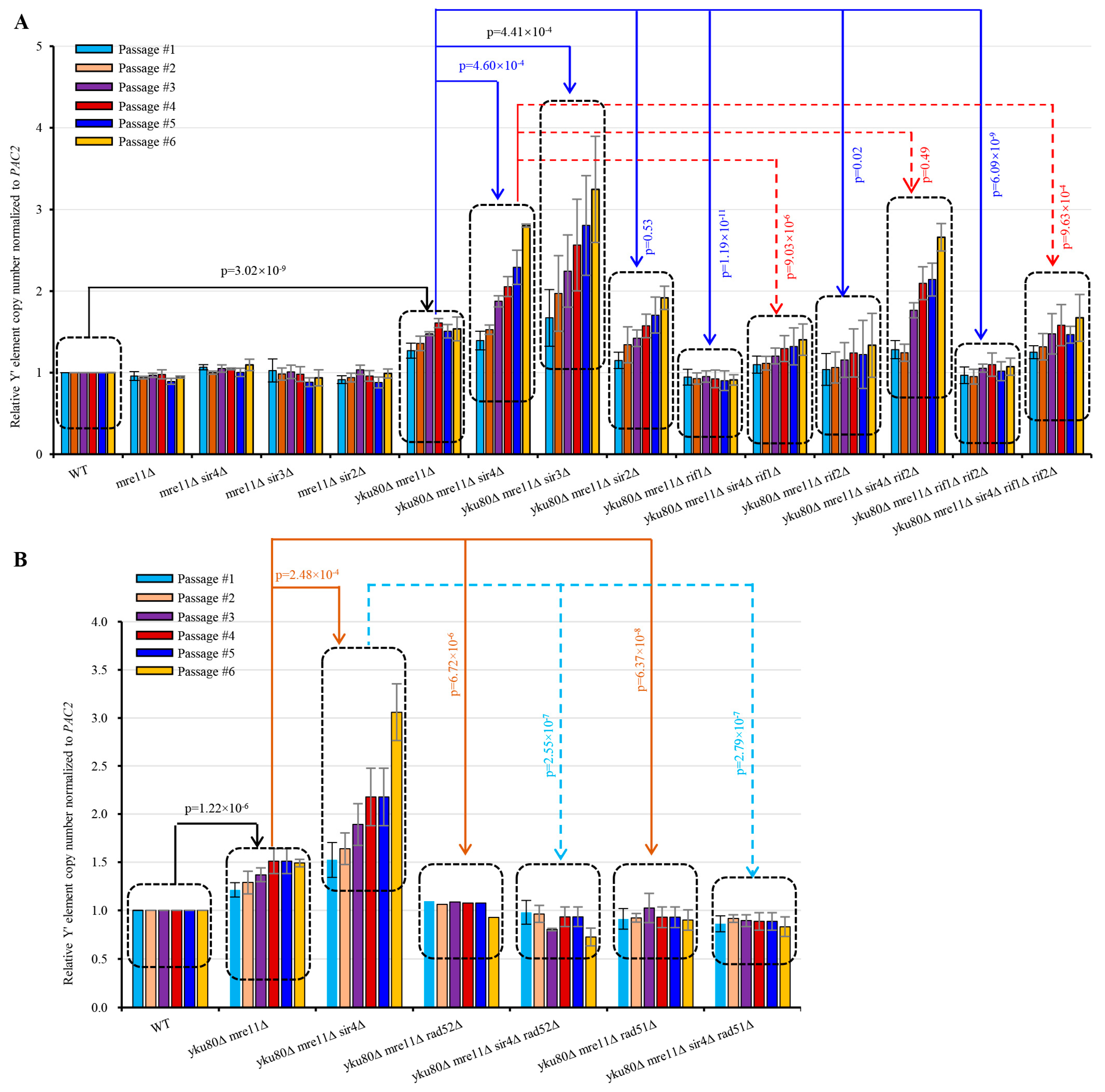
3.5. Sir4 Deficiency Enhances, whereas Rif1 or Rif1/2 Deficiency Suppresses, Y′ Element Amplification
3.6. Rif1 or Rif1/2 Deficiency Compromises sir4Δ-induced Y′ Element Recombination
3.7. Rad51 Deficiency Eliminates Y′ Element Amplification in yku80∆ mre11∆ and yku80∆ mre11∆ sir4∆ Cells
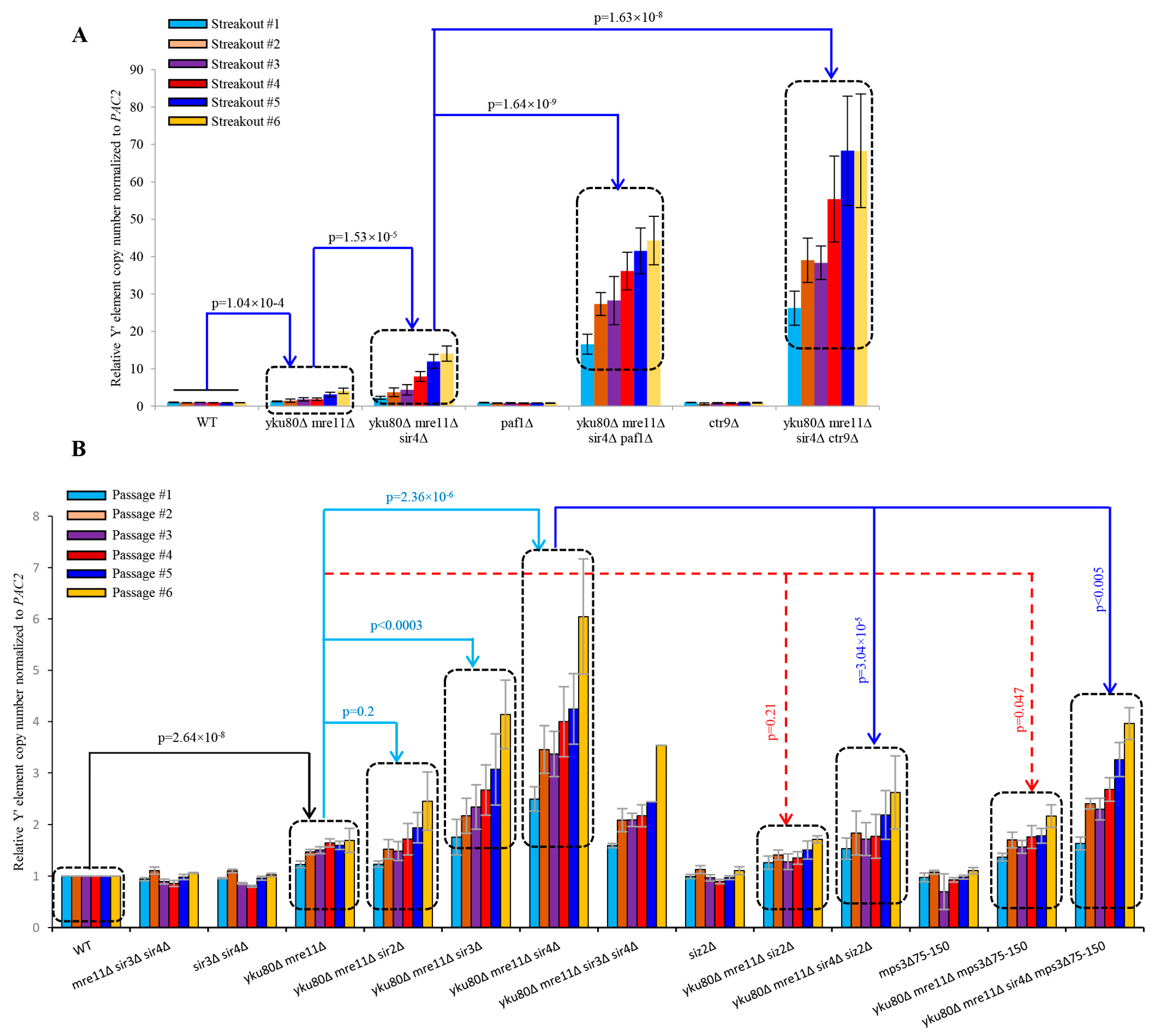
3.8. Increased TERRA and Loss of Telomere Perinuclear Localization Contributes to the Y′ Element Amplification Induced by Sir4 Deficiency
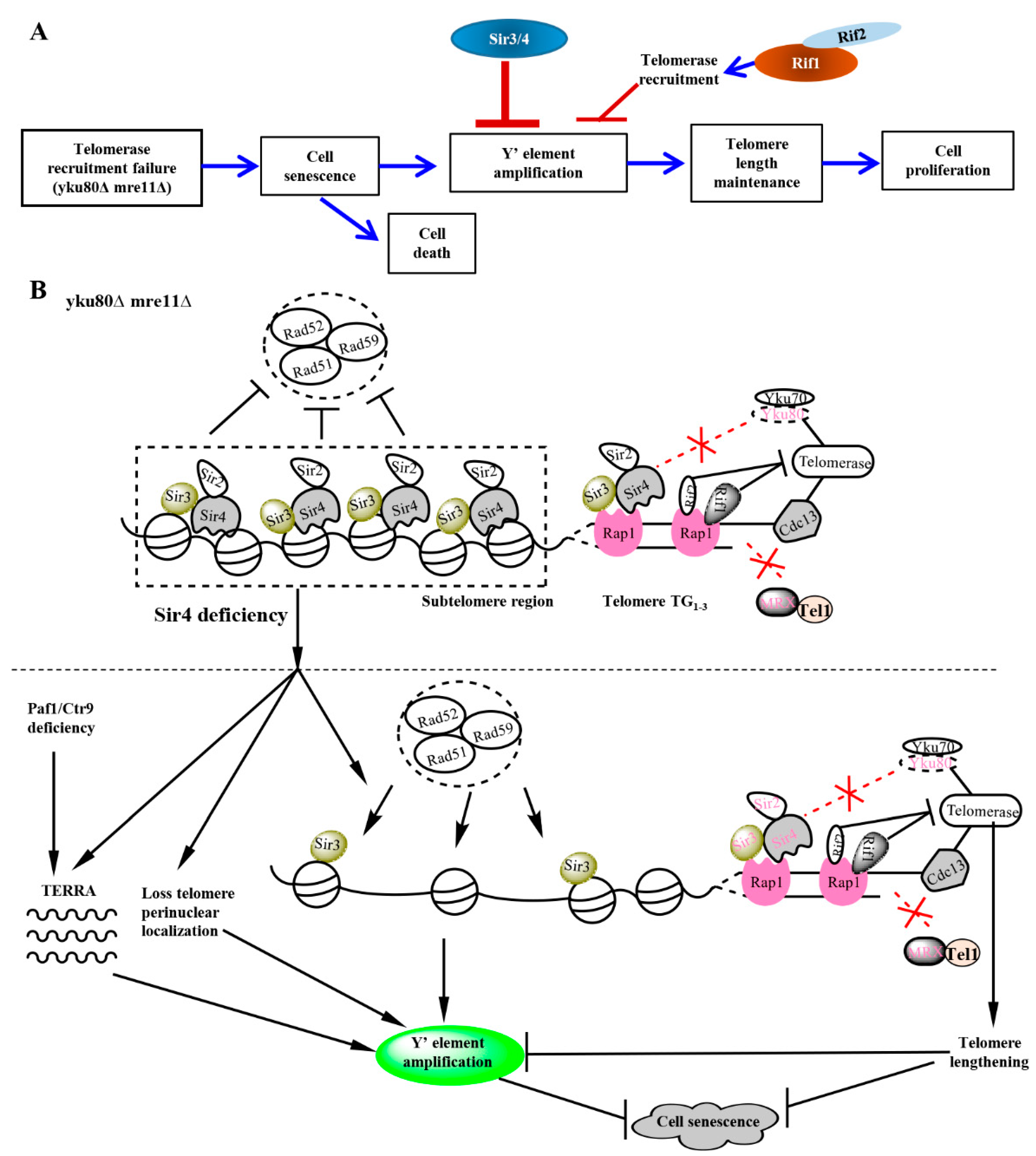
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shay, J.W. Role of Telomeres and Telomerase in Aging and Cancer. Cancer Discov. 2016, 6, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Opresko, P.L.; Shay, J.W. Telomere-associated aging disorders. Ageing Res. Rev. 2017, 33, 52–66. [Google Scholar] [CrossRef] [PubMed]
- Martínez, P.; Blasco, M.A. Telomere-driven diseases and telomere-targeting therapies. J. Cell Biol. 2017, 216, 875–887. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Falandry, C.; Bonnefoy, M.; Freyer, G.; Gilson, E. Biology of Cancer and Aging: A Complex Association With Cellular Senescence. J. Clin. Oncol. 2014, 32, 2604–2610. [Google Scholar] [CrossRef] [PubMed]
- Hass, E.P.; Zappulla, D.C. The Ku subunit of telomerase binds Sir4 to recruit telomerase to lengthen telomeres in S. cerevisiae. eLife 2015, 4, e07750. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xue, J.; Churikov, D.; Hass, E.P.; Shi, S.; Lemon, L.D.; Luciano, P.; Bertuch, A.A.; Zappulla, D.C.; Géli, V.; et al. Structural Insights into Yeast Telomerase Recruitment to Telomeres. Cell 2018, 172, 331. [Google Scholar] [CrossRef] [PubMed]
- Kupiec, M. Biology of telomeres: Lessons from budding yeast. FEMS Microbiol. Rev. 2014, 38, 144–171. [Google Scholar] [CrossRef] [PubMed]
- Wellinger, R.J.; Zakian, V.A. Everything You Ever Wanted to Know About Saccharomyces cerevisiae Telomeres: Beginning to End. Genetics 2012, 191, 1073–1105. [Google Scholar] [CrossRef]
- Lundblad, V.; Szostak, J.W. A mutant with a defect in telomere elongation leads to senescence in yeast. Cell 1989, 57, 633–643. [Google Scholar] [CrossRef]
- Lendvay, T.S.; Morris, D.K.; Sah, J.; Balasubramanian, B.; Lundblad, V. Senescence Mutants of Saccharomyces cerevisiae With a Defect in Telomere Replication Identify Three Additional EST Genes. Genetics 1996, 144, 1399–1412. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.S.; Gottschling, D.E. TLC1: Template RNA component of Saccharomyces cerevisiae telomerase. Science 1994, 266, 404–409. [Google Scholar] [CrossRef]
- Lemieux, B.; Laterreur, N.; Perederina, A.; Noël, J.F.; Dubois, M.L.; Krasilnikov, A.S.; Wellinger, R.J. Active Yeast Telomerase Shares Subunits with Ribonucleoproteins RNase P and RNase MRP. Cell 2016, 165, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Stellwagen, A.E.; Haimberger, Z.W.; Veatch, J.R.; Gottschling, D.E. Ku interacts with telomerase RNA to promote telomere addition at native and broken chromosome ends. Genes Dev. 2003, 17, 2384–2395. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.E.; Stellwagen, A.E.; Diede, S.J.; Singer, M.S.; Haimberger, Z.W.; Johnson, C.O.; Tzoneva, M.; Gottschling, D.E. The function of a stem-loop in telomerase RNA is linked to the DNA repair protein Ku. Nat. Genet. 2001, 27, 64–67. [Google Scholar] [CrossRef] [PubMed]
- Bertuch, A.A.; Lundblad, V. EXO1 Contributes to Telomere Maintenance in Both Telomerase-Proficient and Telomerase-Deficient Saccharomyces cerevisiae. Genetics 2004, 166, 1651–1659. [Google Scholar] [CrossRef]
- Mimitou, E.P.; Symington, L.S. Ku prevents Exo1 and Sgs1-dependent resection of DNA ends in the absence of a functional MRX complex or Sae2. EMBO J. 2010, 29, 3358–3369. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Wang, Z.; Liu, J.-P. Roles of Telomere Biology in Cell Senescence, Replicative and Chronological Ageing. Cells 2019, 8, 54. [Google Scholar] [CrossRef]
- Nugent, C.I.; Hughes, T.R.; Lue, N.F.; Lundblad, V. Cdc13p: A Single-Strand Telomeric DNA-Binding Protein with a Dual Role in Yeast Telomere Maintenance. Science 1996, 274, 249. [Google Scholar] [CrossRef]
- Lundblad, V.; Blackburn, E.H. An alternative pathway for yeast telomere maintenance rescues est1− senescence. Cell 1993, 73, 347–360. [Google Scholar] [CrossRef]
- Nugent, C.I.; Bosco, G.; Ross, L.O.; Evans, S.K.; Salinger, A.P.; Moore, J.K.; Haber, J.E.; Lundblad, V. Telomere maintenance is dependent on activities required for end repair of double-strand breaks. Curr. Biol. 1998, 8, 657–662. [Google Scholar] [CrossRef]
- Maringele, L.; Lydall, D. EXO1 Plays a Role in Generating Type I and Type II Survivors in Budding Yeast. Genetics 2004, 166, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Le, S.; Moore, J.K.; Haber, J.E.; Greider, C.W. RAD50 and RAD51 Define Two Pathways That Collaborate to Maintain Telomeres in the Absence of Telomerase. Genetics 1999, 152, 143–152. [Google Scholar] [PubMed]
- Signon, L.; Malkova, A.; Naylor, M.L.; Klein, H.; Haber, J.E. Genetic Requirements for RAD51- and RAD54-Independent Break-Induced Replication Repair of a Chromosomal Double-Strand Break. Mol. Cell. Biol. 2001, 21, 2048. [Google Scholar] [CrossRef] [PubMed]
- Lydeard, J.R.; Jain, S.; Yamaguchi, M.; Haber, J.E. Break-induced replication and telomerase-independent telomere maintenance require Pol32. Nature 2007, 448, 820–823. [Google Scholar] [CrossRef] [PubMed]
- Churikov, D.; Charifi, F.; Simon, M.-N.; Géli, V. Rad59-Facilitated Acquisition of Y′ Elements by Short Telomeres Delays the Onset of Senescence. PLoS Genet. 2014, 10, e1004736. [Google Scholar] [CrossRef] [PubMed]
- Laud, P.R.; Multani, A.S.; Bailey, S.M.; Wu, L.; Ma, J.; Kingsley, C.; Lebel, M.; Pathak, S.; DePinho, R.A.; Chang, S. Elevated telomere-telomere recombination in WRN-deficient, telomere dysfunctional cells promotes escape from senescence and engagement of the ALT pathway. Genes Dev. 2005, 19, 2560–2570. [Google Scholar] [CrossRef]
- McEachern, M.J.; Iyer, S. Short Telomeres in Yeast Are Highly Recombinogenic. Mol. Cell 2001, 7, 695–704. [Google Scholar] [CrossRef]
- Morrish, T.A.; Greider, C.W. Short Telomeres Initiate Telomere Recombination in Primary and Tumor Cells. PLoS Genet. 2009, 5, e1000357. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Vega-Palas, M.A.; Grunstein, M. Rap1-Sir4 binding independent of other Sir, yKu, or histone interactions initiates the assembly of telomeric heterochromatin in yeast. Genes Dev. 2002, 16, 1528–1539. [Google Scholar] [CrossRef]
- Aparicio, O.M.; Billington, B.L.; Gottschling, D.E. Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell 1991, 66, 1279–1287. [Google Scholar] [CrossRef]
- Kueng, S.; Oppikofer, M.; Gasser, S.M. SIR Proteins and the Assembly of Silent Chromatin in Budding Yeast. Annu. Rev. Genet. 2013, 47, 275–306. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, F.; Laterreur, N.; Cusanelli, E.; Ouenzar, F.; Querido, E.; Wellinger, R.J.; Chartrand, P. Live Cell Imaging of Telomerase RNA Dynamics Reveals Cell Cycle-Dependent Clustering of Telomerase at Elongating Telomeres. Mol. Cell 2011, 44, 819–827. [Google Scholar] [CrossRef]
- Wotton, D.; Shore, D. A novel Rap1p-interacting factor, Rif2p, cooperates with Rif1p to regulate telomere length in Saccharomyces cerevisiae. Genes Dev. 1997, 11, 748–760. [Google Scholar] [CrossRef] [PubMed]
- Pfingsten, J.S.; Goodrich, K.J.; Taabazuing, C.; Ouenzar, F.; Chartrand, P.; Cech, T.R. Mutually Exclusive Binding of Telomerase RNA and DNA by Ku Alters Telomerase Recruitment Model. Cell 2012, 148, 922–932. [Google Scholar] [CrossRef]
- Longtine, M.S.; Mckenzie III, A.; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J. RAdditional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; He, M.H.; Peng, J.; Duan, Y.M.; Lu, Y.S.; Wu, Z.; Gong, T.; Li, H.T.; Zhou, J.Q. Tethering telomerase to telomeres increases genome instability and promotes chronological aging in yeast. Aging 2016, 8, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, J.-P. A method for efficient quantitative analysis of genomic subtelomere Y′ element abundance in yeasts. Yeast 2020, 37, 373–388. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.; Lydall, D. Paf1 and Ctr9, core components of the PAF1 complex, maintain low levels of telomeric repeat containing RNA. Nucleic Acids Res. 2017, 46, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Bupp, J.M.; Martin, A.E.; Stensrud, E.S.; Jaspersen, S.L. Telomere anchoring at the nuclear periphery requires the budding yeast Sad1-UNC-84 domain protein Mps3. J. Cell Biol. 2007, 179, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, H.C.; Luke, B.; Schober, H.; Kalck, V.; Lingner, J.; Gasser, S.M. The PIAS homologue Siz2 regulates perinuclear telomere position and telomerase activity in budding yeast. Nat. Cell Biol. 2011, 13, 867. [Google Scholar] [CrossRef] [PubMed]
- Takata, H.; Tanaka, Y.; Matsuura, A. Late S Phase-Specific Recruitment of Mre11 Complex Triggers Hierarchical Assembly of Telomere Replication Proteins in Saccharomyces cerevisiae. Mol. Cell 2005, 17, 573–583. [Google Scholar] [CrossRef] [PubMed]
- de Bruin, D.; Zaman, Z.; Liberatore, R.A.; Ptashne, M. Telomere looping permits gene activation by a downstream UAS in yeast. Nature 2001, 409, 109. [Google Scholar] [CrossRef] [PubMed]
- Poschke, H.; Dees, M.; Chang, M.; Amberkar, S.; Kaderali, L.; Rothstein, R.; Luke, B. Rif2 Promotes a Telomere Fold-Back Structure through Rpd3L Recruitment in Budding Yeast. PLoS Genet. 2012, 8, e1002960. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Hong, X.; Wang, L.; Liang, C.-Y.; Liu, J.-P. Sir4 Deficiency Reverses Cell Senescence by Sub-Telomere Recombination. Cells 2021, 10, 778. https://doi.org/10.3390/cells10040778
Liu J, Hong X, Wang L, Liang C-Y, Liu J-P. Sir4 Deficiency Reverses Cell Senescence by Sub-Telomere Recombination. Cells. 2021; 10(4):778. https://doi.org/10.3390/cells10040778
Chicago/Turabian StyleLiu, Jun, Xiaojing Hong, Lihui Wang, Chao-Ya Liang, and Jun-Ping Liu. 2021. "Sir4 Deficiency Reverses Cell Senescence by Sub-Telomere Recombination" Cells 10, no. 4: 778. https://doi.org/10.3390/cells10040778
APA StyleLiu, J., Hong, X., Wang, L., Liang, C.-Y., & Liu, J.-P. (2021). Sir4 Deficiency Reverses Cell Senescence by Sub-Telomere Recombination. Cells, 10(4), 778. https://doi.org/10.3390/cells10040778