Genotype-by-Environment Interaction and Yield Stability of Maize Single Cross Hybrids Developed from Tropical Inbred Lines

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Germplasm
2.2. Experimental Sites
2.3. Site Preparation and Soil Analysis
2.4. Experimental Design
2.5. Data Collection
- MC = moisture content in grains at harvest (%)
- 0.8 = Shelling coefficient
- 10,000 m2 corresponds to 1 hectare
- 15% = moisture content required in maize grain at storage
2.6. Statistical Analysis
3. Results
3.1. Analysis of Variance for Grain Yield across Environments
3.2. Yield Performance of the 20 Best Performing Hybrids and Four Checks across Eleven Environments
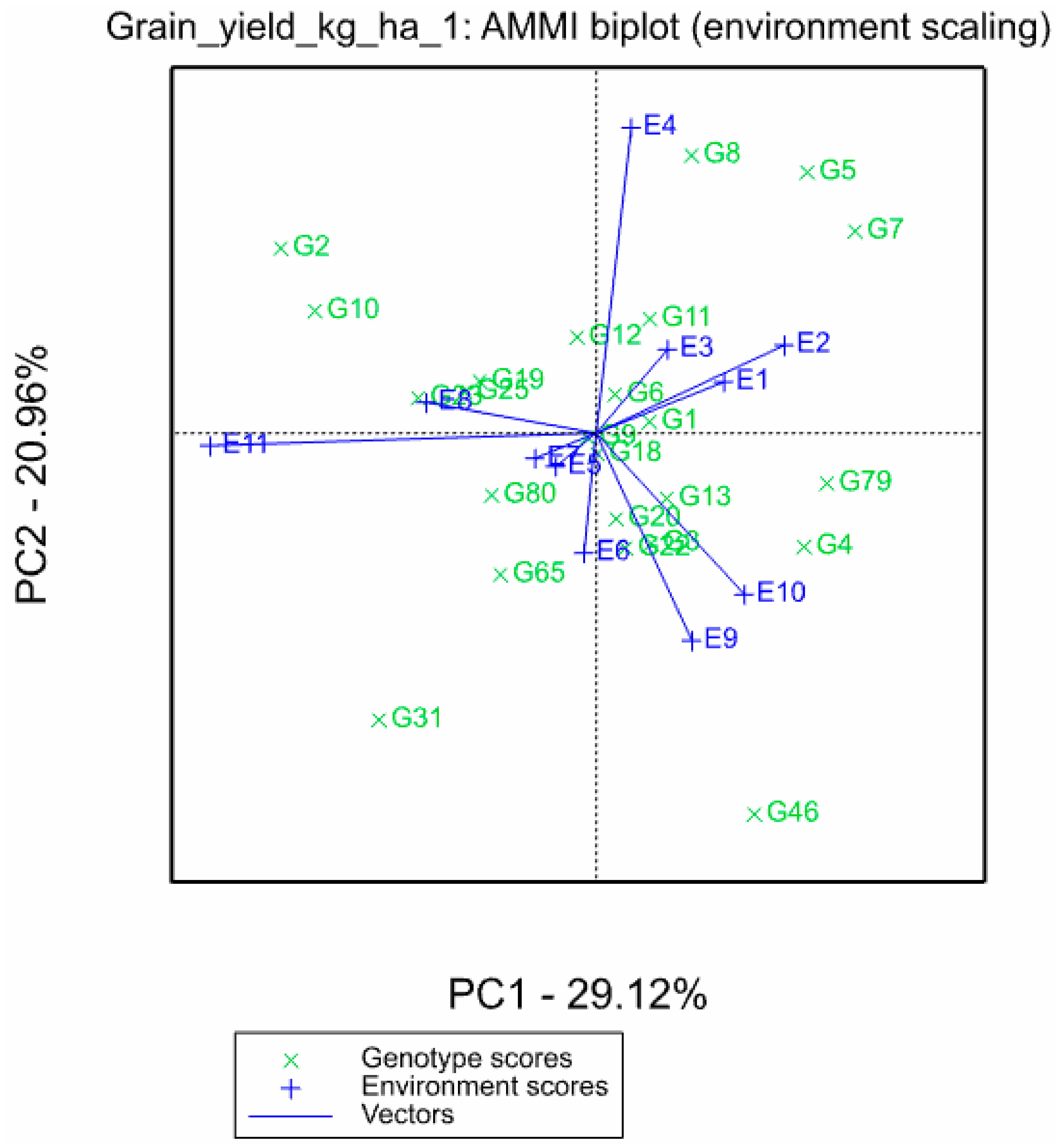
3.3. Additive Main Effect and Multiplicative Interaction (AMMI) Analysis of 24 Hybrids for Grain Yield
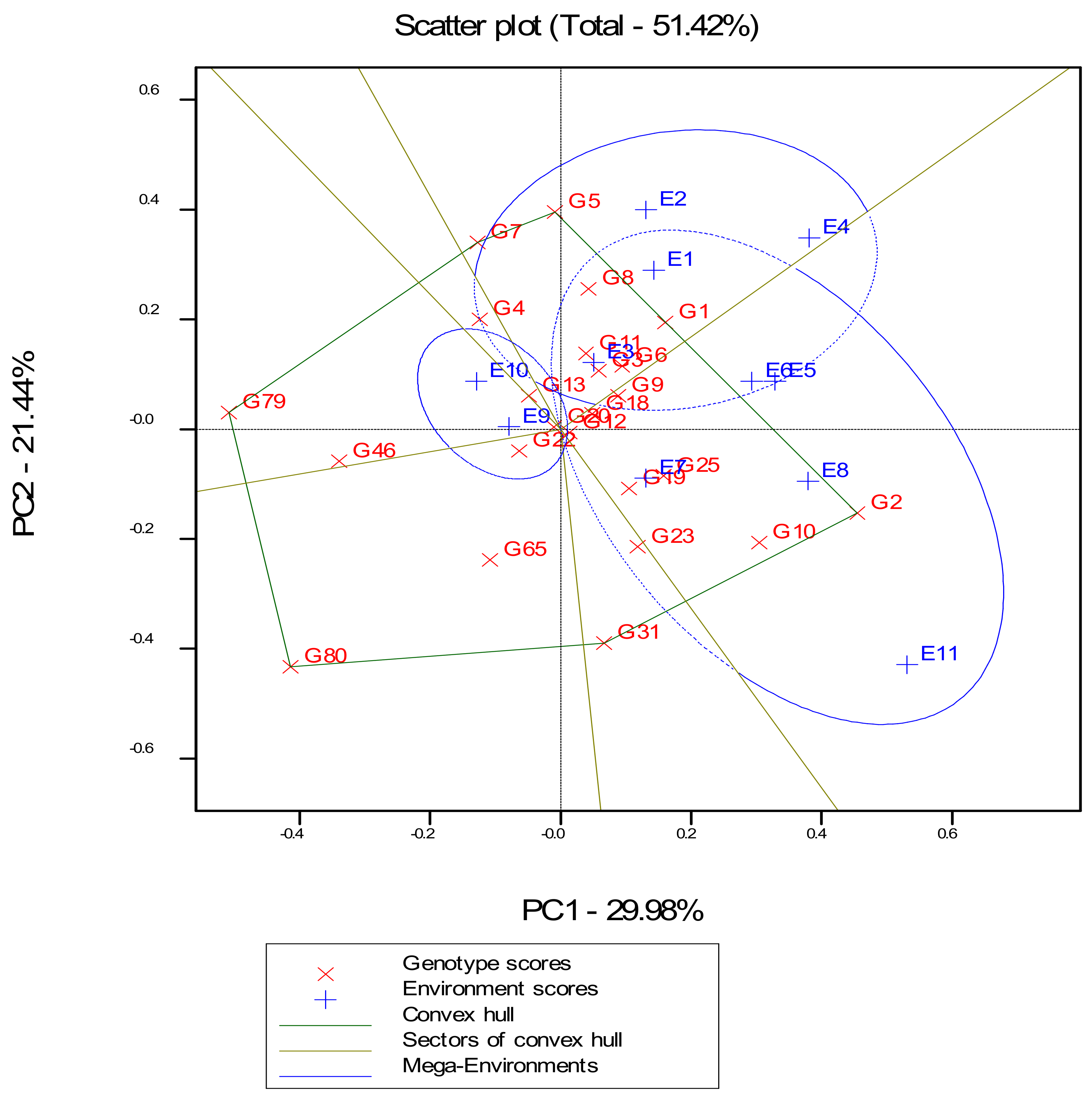
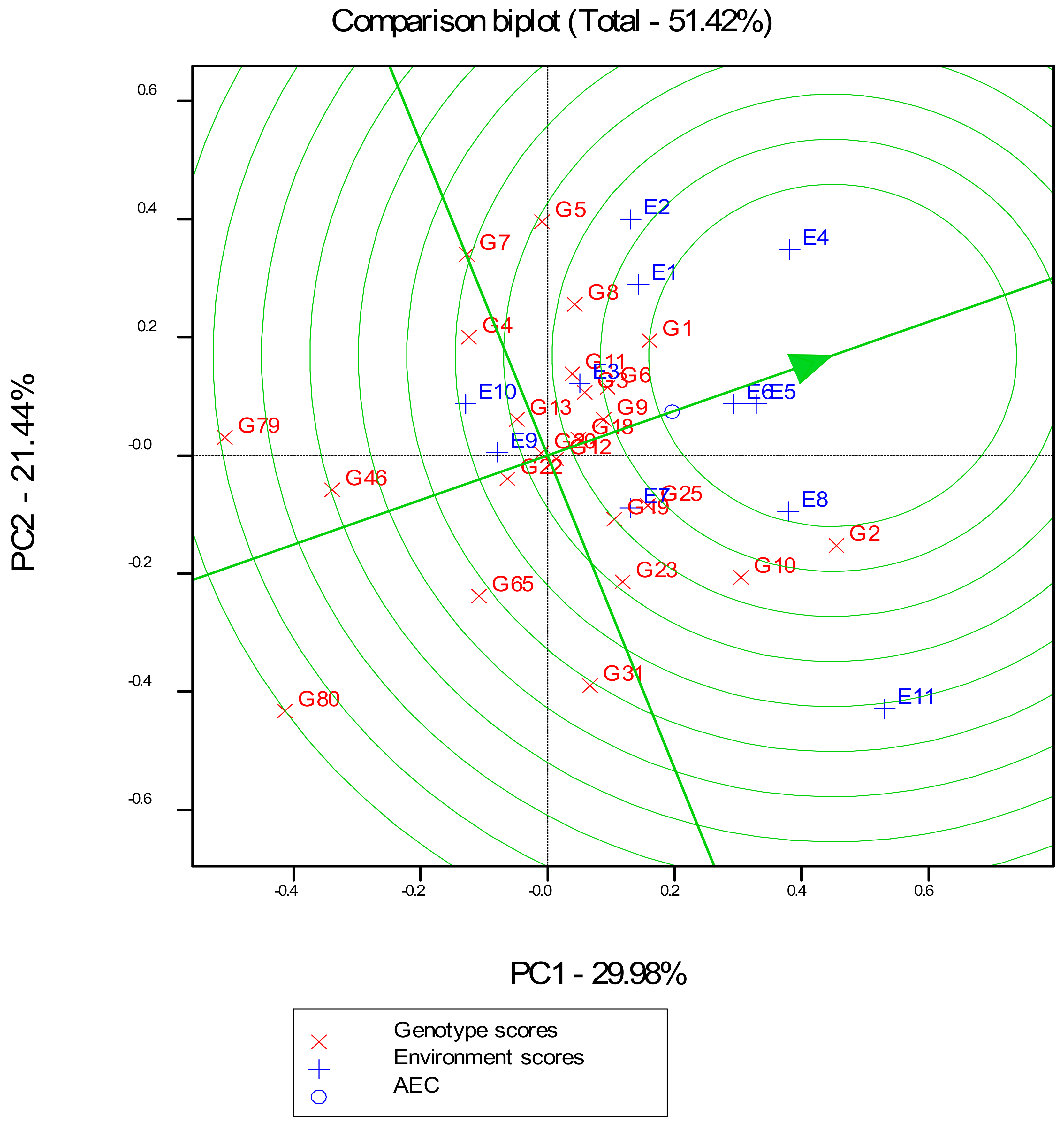
3.4. GGE Biplot Analysis of Best 20 Hybrids and Four Checks
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Badu-Apraku, B.; Akinwale, R. Identification of early-maturing maize inbred lines based on multiple traits under drought and low N environments for hybrid development and population improvement. Can. J. Plant Sci. 2011, 91, 931–942. [Google Scholar] [CrossRef][Green Version]
- Ngo Nonga, F. Durabilité des Activités Agricoles des Exploitations Familiales Agricoles à Base de Maïs du Grand Sud Cameroun, 2èmes Journées de Recherches en Sciences Sociales; INRA SFER CIRAD: Lille, France, 2008; p. 20. [Google Scholar]
- Derera, J. Genetic Effects and Associations between Grain Yield Potential, Stress Tolerance and Yield Stability in Southern African Maize (Zea mays L.) Base Germplasm; School of Biochemistry, Genetics, Microbiology and Plant Pathology, Faculty of Science and Agriculture, University of KwaZulu-Natal: Durban, South Africa, 2005; p. 175. [Google Scholar]
- Mafouasson, A.H.N.; Kenga, R.; Gracen, V.; Yeboah, A.M.; Mahamane, N.L.; Tandzi, N.L.; Ntsomboh-Ntsefong, G. Combining Ability and Gene Action of Tropical Maize (Zea mays L.) Inbred Lines under Low and High Nitrogen Conditions. J. Agric. Sci. 2017, 9, 222–235. [Google Scholar]
- Ajala, S.O.; Olaniyan, A.B.; Olayiwola, M.O.; Job, A.O. Yield improvement in maize for tolerance to low soil nitrogen. Plant Breed. 2018, 137, 118–126. [Google Scholar] [CrossRef]
- Bänziger, M.; Edmeades, G.O.; Beck, D.; Bellon, M. Breeding for Drought and Nitrogen Stress Tolerance in Maize: From Theory to Practice; CIMMYT: Mexico, Mexico, 2000. [Google Scholar]
- Abakemal, D.; Shimelis, H.; Derera, J. Genotype-by environment interaction and yield stability of quality protein maize hybrids developed from tropical-highland adapted inbred lines. Euphytica 2016, 209, 757–769. [Google Scholar] [CrossRef]
- Citizens Association for the Defense of Collective Interests (ACDIC). The Maize Crisis and the Misfortunes of Cameroon’s Agriculture; ACDIC: Yaounde, Cameroon, 2010; p. 46. [Google Scholar]
- Bänziger, M.; Setimela, P.S.; Hodson, D.; Vivek, B. Breeding for improved drought tolerance in maize adapted to Southern Africa. “New directions for a diverse planet”. In Proceedings of the 4th International Crop Science Congress, Brisbane, Australia, 26 September–1 October 2004; [CDROM]: International Crop Science Congress: Brisbane, Australia, 2004. [Google Scholar]
- Sibiya, J.; Tongoona, P.; Derera, J.; Rij, N. Genetic analysis and genotype by environment (G X E) for grey leaf spot disease resistance in elite African maize (Zea mays L.) germplasm. Euphytica 2012, 185, 349–362. [Google Scholar] [CrossRef]
- Falconer, D.S.; Mackay, T.F.C. Introduction to Quantitative Genetics; Longman: New York, NY, USA, 1996. [Google Scholar]
- Miti, F. Breeding Investigations of Maize (Zea mays L.) Genotypes for Tolerance Tolow Nitrogen and Drought in Zambia. Ph.D. Thesis, University of Kwa-Zulu Natal, Pietermaritzburg, South Africa, 2007; 196p. [Google Scholar]
- Yan, W.; Tinker, N.A. Biplot analysis of multi-environment trial data: Principles and applications. Can. J. Plant Sci. 2006, 86, 623–645. [Google Scholar] [CrossRef]
- Kenga, R. Combining Ability Estimates and Heterosis in Selected Tropical Sorghum (Sorghum bicolor (L.) Moench); Department of Plant Science, Faculty of Agriculture, Ahmadu Bello University Zaria: Zaria, Nigeria, 2001; p. 186. [Google Scholar]
- Khalil, I.A.; Rahman, H.U.; Rehman, N.U.; Arif, M.; Khalil, I.H.; Iqbal, M.; Hidayatullah; Afridi, K.; Sajjad, M.; Ishaq, M. Evaluation of maize hybrids for grain yield stability in north-west of Pakistan. Sarhad J. Agric. 2011, 27, 213–218. [Google Scholar]
- IITA. Maize 2014. Available online: http://www.iita.org/maize (accessed on 16 June 2014).
- Etoundi, S.M.N.; Dia, K.B. Determinants of the adoption of improved varieties of Maize in Cameroon: Case of CMS 8704. Globalisation, Institutions and African Economic Development. In Proceedings of the African Economic Conference, Tunis, Tunisia, 12–14 November 2008; pp. 397–413. [Google Scholar]
- Badu-Apraku, B.; Oyekunle, M.; Obeng-Antwi, K.; Osuman, A.S.; Ado, S.G.; Coulibay, N.; Yallou, C.G.; Abdulai, M.; Boakyewaa, G.A.; Didjeira, A. Performance of extra-early maize cultivars based on GGE biplot and AMMI analysis. J. Agric. Sci. 2012, 150, 473–483. [Google Scholar] [CrossRef]
- Ndhlela, T. Improvement Strategies for Yield Potential, Disease Resistance and Drought Tolerance of Zimbabwean Maize Inbred Lines; Department of Plant Sciences (Plant Breeding), University of the Free State: Bloemfontein, South Africa, 2012; 295p. [Google Scholar]
- Adu, G.B.; Akromah, R.; Abdulai, M.S.; Obeng-Antwi, K.; Kena, A.W.; Tengan, K.M.L.; Alidu, H. Assessment of Genotype by Environment interactions and Grain Yield Performance of Extra-Early Maize (Zea mays L.) Hybrids. J. Biol. Agric. Healthc. 2013, 3, 7–15. [Google Scholar]
- Badu-Apraku, B.; Akinwale, R.O.; Menkir, A.; Obeng-Antwi, K.; Osuman, A.S.; Coulibaly, N.; Onyibe, J.E.; Yallou, G.C.; Abdullai, M.S.; Didjera, A. Use of GGE Biplot for Targeting Early Maturing Maize Cultivars to Mega-environments in West Africa. Afr. Crop Sci. J. 2011, 19, 79–96. [Google Scholar] [CrossRef][Green Version]
- Eberhart, S.A.; Russell, W.A. Stability parameters for comparing varieties. Crop Sci. 1966, 6, 36–40. [Google Scholar] [CrossRef]
- Gauch, H.G. Model selection and validation for yield trials with interaction. Biometrics 1988, 44, 705–715. [Google Scholar] [CrossRef]
- Gauch, H.G.; Zobel, R.W. Identifying mega-environments and targeting genotypes. Crop Sci. 1997, 37, 311–326. [Google Scholar] [CrossRef]
- Zobel, R.W.; Wright, M.J.; Gauch, H.G. Statistical analysis of a yield trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Yan, W.; Hunt, L.A.; Sheng, Q.; Szlavnics, Z. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci. 2000, 40, 597–605. [Google Scholar] [CrossRef]
- Gauch, H.G.; Zobel, R.W. AMMI analyses of yield trials. In Genotype by Environment Interaction; Kang, M.S., Gauch, H.G., Eds.; CRC Press: Boca Raton, FL, USA, 1996; pp. 85–122. [Google Scholar]
- Ebdon, J.S.; Gauch, H.G. Additive main effects and multiplicative interaction analysis of national turfgrass performance trials. Crop Sci. 2002, 42, 497–506. [Google Scholar] [CrossRef]
- Gauch, H.G. Statistical analysis of yield trials by AMMI and GGE. Crop Sci. 2006, 46, 1488–1500. [Google Scholar] [CrossRef]
- Cooper, M.; Stucker, R.E.; DeLacy, I.H.; Harch, B.D. Wheat breeding nurseries, target environments, and indirect selection for grain yield. Crop Sci. 1997, 37, 1168–1176. [Google Scholar] [CrossRef]
- Yan, W. GGEbiplot-a windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agron. J. 2001, 93, 1111–1118. [Google Scholar] [CrossRef]
- The, C.; Ngonkeu, M.L.; Zonkeng, C.; Apala, H.M. Evaluation and Selection of Maize (Zea mays L.) Genotypes Tolerant to Low N Soil. Optimizing Productivity of Food Crop Genotypes in Low Nutrient Soils; IAEA-TECDOC-1721: Vienna, Austria, 2013; pp. 251–264. [Google Scholar]
- Tchienkoua, M. Soil and Land-Use Survey of the Northern Section of the Mbalmayo Forest Reserve (Southern Cameroon). A Study for the Selection of the IITA Humid Forest Station Site; Ressource and Crop Management Research Monography N° 24; International Institute of Tropical Agriculture: Ibadan, Nigeria, 1996; p. 65. [Google Scholar]
- Heanes, D.L. Determination of organic C in soils by an improved chromic acid digestion and spectrophotometric procedure. Commun. Soil Sci. Plant Anal. 1984, 15, 1191–1213. [Google Scholar] [CrossRef]
- Buondonno, A.; Rashad, A.A.; Coppola, E. Comparing tests for soil fertility. II. The hydrogen peroxide/sulfuric acid treatment as an alternative to the copper/selenium catalyzed digestion process for routine determination of soil nitrogen-Kjeldahl. Commun. Soil Sci. Plant Anal. 1995, 26, 1607–1619. [Google Scholar] [CrossRef]
- Anderson, J.M.; Ingram, J.S.I. Tropical Soil Biology and Fertility: A Handbook of Methods, 2nd ed.; CAB International, The Cambrian News: Aberstwyth, UK, 1993; p. 221. [Google Scholar]
- Mehlich, M. Mehlich 3 soil text extractant: A modification of the Mehlich 2 extractant. Commun. Soil Sci. Plant Anal. 1984, 15, 1409–1416. [Google Scholar] [CrossRef]
- Barnhisel, R.; Bertsch, P. Aluminum. In Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties; Page, A., Miller, R., Keeney, D., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 275–300. [Google Scholar]
- Mosquera, A.; Mombiela, F. Comparison of three methods for determination of soil Al in an unbuffered salt-extract. Commun. Soil Sci. Plant Anal. 1986, 17, 30–35. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Statistical Analysis System (SAS), version SAS/STAT 9.2; SAS Institute Inc.: Cary, NC, USA, 2008.
- Cochran, W.G.; Cox, G.M. Experimental Designs; John Wiley & Sons: Hoboken, NJ, USA, 1960. [Google Scholar]
- The IBP Breeding Management System Version 2.1. The Intergated Breeding Platform. May 2014. Available online: https://www.integratedbreeding.net/breeding-management-system (accessed on 13 May 2014).
- Acquaah, G. Principles of Plant Genetics and Breeding; Wiley-Blackwell Publishers: Hoboken, NJ, USA, 2007. [Google Scholar]
- Yan, W.; Tinker, N.A. An intergrated biplot analysis system for displaying interpreting, exploring genotyped by environments interactions. Crop Sci. 2005, 45, 1004–1016. [Google Scholar] [CrossRef]
- Yan, W.; Rajcan, I. Biplot evaluation of test sites and trait relations of soybean in Ontario. Crop Sci. 2002, 42, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Aroga, R.; Ambassa-kiki, R.; The, C.; Enyong, L.; Ajala, S.O. On farm evaluation of performance of selecetd improved maize varieties in the forest zone of Central Cameroon. In Proceedings of the Seventh Eastern and Southern Africa Regional Maize Conference, Nairobi, Kenya, 5–11 February 2002; pp. 432–437. [Google Scholar]
- Aroga, R.; Coderre, D. Abondance et diversité des foreurs de tiges et grains dans une biculture maïs-arachide au centre du Cameroun. Afr. Crop Sci. J. 2000, 8, 365–374. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Code | Genotypes | Code |
|---|---|---|---|
| CLYN246 × 87036 | G1 | 88069 × 87036 | G39 |
| TL-11-A-1642-5 × 87036 | G2 | ATP S8 30 Y-3 × 87036 | G40 |
| CLWN201 × Exp1 24 | G3 | CML 254 × Exp1 24 | G41 |
| J16-1 × Exp1 24 | G4 | CLYN246 × 9071 | G42 |
| 1368 × 87036 | G5 | CLWN201 × 9071 | G43 |
| CLQRCWQ26 × Exp1 24 | G6 | CML343 × Exp1 24 | G44 |
| TL-11-A-1642-5 × Exp1 24 | G7 | CLQRCWQ26 × 9071 | G45 |
| TZ-STR-133 × 87036 | G8 | ATP S9 30 Y-1 × Exp1 24 | G47 |
| CLWN201 × 87036 | G9 | ATP S6-20-Y-1 × 9071 | G48 |
| ATP S6-20-Y-1 × Exp1 24 | G10 | J16-1 × 9071 | G49 |
| CLA 18 × Exp1 24 | G11 | CML 358 × Exp1 24 | G50 |
| ATP S6-20-Y-1 × 87036 | G12 | Entrada 3 × 87036 | G51 |
| Cam inb gp1 17 × 87036 | G13 | CML494 × 87036 | G52 |
| J16-1 × 87036 | G14 | J18-1 × 87036 | G53 |
| 4001STR × 87036 | G15 | CML 444 × 87036 | G54 |
| CML343 × 87036 | G16 | Cam inb gp1 17 × 9071 | G55 |
| CLA 18 × 87036 | G17 | ATP S5 31 Y-2 × 9071 | G56 |
| CML395 × Exp1 24 | G18 | V-481-73 × Exp1 24 | G57 |
| CML451 × 87036 | G19 | Cla 17 × Exp1 24 | G58 |
| CML343 × 9071 | G20 | 5057 × Exp1 24 | G59 |
| 88069 × 9071 | G21 | ATP S8 30 Y-3 × Exp1 24 | G60 |
| CLQRCWQ26 × 87036 | G22 | KU1414 × Exp1 24 | G61 |
| ATP S6 20 Y-2 × Exp1 24 | G23 | TZ-STR-133 × Exp1 24 | G62 |
| 4001STR × 9071 | G24 | ATP S5 31 Y-2 × Exp1 24 | G63 |
| ATP S5 31 Y-2 × 87036 | G25 | ATP S6 20 Y-2 × 9071 | G64 |
| ATP S9 30 Y-1 × 87036 | G26 | Cla 17 × 87036 | G66 |
| 1368 × Exp1 24 | G27 | ATP S8 30 Y-3 × 9071 | G67 |
| CML165 × 87036 | G28 | CML 254 × 87036 | G68 |
| CML 358 × 87036 | G29 | 88094 × 87036 | G69 |
| KU1414 × 87036 | G30 | TZ-STR-133 × 9071 | G70 |
| Entrada 29 × Exp1 24 | G31 | CML451 × 9071 | G71 |
| CML494 × 9071 | G32 | CML 254 × 9071 | G72 |
| CML 444 × Exp1 24 | G33 | TZMI 102 × 87036 | G73 |
| 88069 × Exp1 24 | G34 | TZMI 102 × Exp1 24 | G74 |
| Cam inb gp1 17 × Exp1 24 | G35 | Ku1409 × 9071 | G75 |
| CLYN246 × Exp1 24 | G36 | Entrada 3 × 9071 | G76 |
| 1368 × 9071 | G37 | 5012 × 87036 | G77 |
| Ku1409 × 87036 | G38 | Ku1409 × Exp1 24 | G78 |
| Checks | |||
| 87036 × Exp1 24 | G46 | 87036 × 9071 | G79 |
| Exp1 24 × 9071 | G65 | 88069 × Cam inb gp1 17 | G80 |
| Site | Latitude, Longitude and Altitude | Environments Code | Year | Season | Soil Management | Average Rain Fall |
|---|---|---|---|---|---|---|
| Mbalmayo | 3°31′ N, 11°30′ E, 641 m a.s.l. | E1 | 2012 | Minor | Low N | 488.87 mm |
| E2 | 2012 | Minor | Optimum N | 488.87 mm | ||
| E5 | 2013 | Major | Low N | 583.46 mm | ||
| E6 | 2013 | Major | Optimum N | 583.46 mm | ||
| E9 | 2013 | Minor | Low N | 499.66 mm | ||
| E10 | 2013 | Minor | Optimum N | 499.66 mm | ||
| Nkolbisson | 3° 44 N, 11°36 E, 650 m a.s.l. | E3 | 2012 | Minor | Low N | 281 (October–November) * |
| E4 | 2012 | Minor | Optimum N | 281 (October–November) * | ||
| E7 | 2013 | Major | Low N | 936 mm | ||
| E8 | 2013 | Major | Optimum N | 936 mm | ||
| E11 | 2013 | Minor | Optimun N | 662 mm |
| Chemical Characteristics | Mbalmayo | Nkolbisson | ||
|---|---|---|---|---|
| 0–10 cm | 10–20 cm | 0–10 cm | 10–20 cm | |
| Exchangeable Ca2+ (cmol kg−1) | 5.92 | 2.58 | 1.53 | 0.88 |
| Exchangeable Mg2+ (cmol kg−1) | 1.15 | 0.63 | 0.77 | 0.46 |
| Exchangeable K+ (cmol kg−1) | 0.11 | 0.06 | 0.38 | 0.24 |
| Cation Exchange Capacity (cmol kg−1) | nd | nd | 10.55 | 9.37 |
| Organic Carbon % | 1.30 | 0.58 | 1.87 | 1.51 |
| C/N | 9.90 | 8.03 | 15.90 | 12.94 |
| Bray Phosphorus (mg kg−1) | 2.11 | 0.99 | 13.85 | 3.10 |
| pH 1:1 (H2O) | 5.97 | 5.04 | 4.54 | 4.36 |
| Total Nitrogen % | ||||
| In 2012 | 0.13 | 0.07 | 0.12 | 0.12 |
| In 2013 | 0.11 | 0.15 | 0.06 | 0.11 |
| Source | df | Sum of Squares | % Contribution to Sum of Squares | Mean Square | Pr > F |
|---|---|---|---|---|---|
| Env | 10 | 2,447,399,522 | 60.13 | 244,739,952 | <0.0001 |
| Rep (Env) | 11 | 127,598,961 | 11,599,906 | <0.0001 | |
| Block (Env × Rep) | 220 | 382,845,773 | 1,740,208 | 0.0001 | |
| Genotype | 79 | 277,051,837 | 6.81 | 3,506,985 | <0.0001 |
| Env × Genotype | 790 | 1,345,460,727 | 33.05 | 1,703,115 | <0.0001 |
| Error | 649 | 767,427,967 | 1,182,478 | ||
| Corrected Total | 1759 | 6,096,183,487 | |||
| CV | 26.15 | ||||
| R2 | 0.87 |
| Genotypes | E1 | E2 | E3 | E4 | E5 | E6 | E7 | E8 | E9 | E10 | E11 | Mean Across |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LO | OP | LO | OP | LO | OP | LO | OP | LO | OP | OP | ||
| 1368 × 87036 | 4790 | 6118 | 2547 | 8253 | 3956 | 4953 | 2038 | 5876 | 3247 | 4419 | 3734 | 4546 |
| TZ-STR-133 × 87036 | 4382 | 5665 | 1508 | 8269 | 2225 | 5507 | 3604 | 7397 | 1602 | 5129 | 4166 | 4499 |
| CLQRCWQ26 × Exp1 24 | 4253 | 4817 | 1520 | 6324 | 5404 | 6317 | 2778 | 6816 | 2163 | 5570 | 5110 | 4639 |
| CLWN201 × 87036 | 4093 | 4739 | 2799 | 4987 | 5293 | 7657 | 3318 | 6535 | 2545 | 3223 | 4779 | 4540 |
| CLYN246 × 87036 | 3979 | 4782 | 1327 | 7809 | 5157 | 7437 | 3175 | 7017 | 3922 | 5880 | 4940 | 5040 |
| TL-11-A-1642-5 × Exp1 24 | 3940 | 6427 | 5401 | 7311 | 2807 | 4755 | 2987 | 3773 | 3718 | 4828 | 4449 | 4583 |
| CML343 × 9071 | 3911 | 4774 | 1234 | 5672 | 3810 | 6142 | 2910 | 6011 | 4257 | 5062 | 5672 | 4486 |
| CML395 × Exp1 24 | 3896 | 4396 | 1024 | 6999 | 3749 | 6599 | 2989 | 5127 | 3595 | 5742 | 6342 | 4572 |
| CLQRCWQ26 × 87036 | 3557 | 4088 | 2546 | 5711 | 2144 | 7155 | 3646 | 5188 | 4550 | 5081 | 5783 | 4485 |
| CLA 18 × Exp1 24 | 3426 | 4745 | 2317 | 7627 | 4370 | 5485 | 2358 | 5906 | 2911 | 5785 | 5376 | 4567 |
| J16-1 × Exp1 24 | 3290 | 4707 | 2531 | 6424 | 4315 | 6054 | 2782 | 5789 | 4664 | 7270 | 3724 | 4694 |
| CML451 × 87036 | 3284 | 4478 | 1528 | 6569 | 2879 | 5796 | 4916 | 7004 | 2847 | 4799 | 6569 | 4590 |
| ATP S5 31 Y-2 × 87036 | 3198 | 4407 | 1425 | 6711 | 4636 | 5877 | 2554 | 6812 | 3240 | 4604 | 7038 | 4571 |
| ATP S6-20-Y-1 × 87036 | 3076 | 4951 | 2447 | 6167 | 3171 | 4555 | 4717 | 8184 | 2655 | 4973 | 4829 | 4518 |
| Cam inb gp1 17 × 87036 | 3043 | 4332 | 761 | 6957 | 3809 | 6089 | 3923 | 5703 | 4604 | 5624 | 4663 | 4499 |
| ATP S6 20 Y-2 × Exp1 24 | 3036 | 3336 | 3651 | 6138 | 3684 | 5448 | 4337 | 6321 | 3074 | 4527 | 7549 | 4621 |
| Entrada 29 × Exp1 24 | 2989 | 3667 | 1536 | 4077 | 4092 | 5742 | 3085 | 5846 | 4905 | 5981 | 9114 | 4602 |
| CLWN201 × Exp1 24 | 2889 | 4491 | 1209 | 6464 | 5707 | 7568 | 3212 | 6532 | 4059 | 5708 | 4380 | 4750 |
| TL-11-A-1642-5 × 87036 | 2734 | 3303 | 1448 | 9531 | 4985 | 6231 | 3531 | 8874 | 3230 | 5006 | 8619 | 5198 |
| ATP S6-20-Y-1 × Exp1 24 | 2554 | 3220 | 2376 | 7367 | 4641 | 6739 | 4697 | 7719 | 2558 | 3281 | 7458 | 4761 |
| Checks | ||||||||||||
| 87036 × Exp1 24 | 2887 | 3801 | 623 | 2765 | 3241 | 6826 | 3350 | 6045 | 4232 | 6410 | 2710 | 3909 |
| 87036 × 9071 | 2175 | 2511 | 699 | 6253 | 1903 | 3273 | 2375 | 4011 | 3986 | 5968 | 2067 | 3211 |
| Exp1 24 × 9071 | 1791 | 3475 | 984 | 4559 | 2850 | 5635 | 2281 | 7950 | 3778 | 4692 | 5155 | 3912 |
| 88069 × Cam inb gp1 17 | 782 | 1280 | 1799 | 4049 | 1804 | 2754 | 2783 | 4600 | 1952 | 4424 | 5032 | 2823 |
| Means | 3066 | 4132 | 1661 | 5882 | 3719 | 5413 | 3130 | 5832 | 3208 | 4993 | 4738 |
| Source | df | Sum of Squares | Mean Squares | Contribution to Total Variation (%) | F Probability |
|---|---|---|---|---|---|
| Genotypes | 23 | 67,848,890 | 2,949,952 | 7.891443 | <0.001 |
| Environments | 10 | 514,356,619 | 51,435,662 | 59.82435 | <0.001 |
| Interactions (G × E) | 230 | 277,572,490 | 1,206,837 | 32.2842 | |
| IPCA 1 | 32 | 80,827,507 | 2,525,860 | 29.22 | <0.001 |
| IPCA 2 | 30 | 58,189,285 | 1,939,643 | 20.96 | <0.001 |
| Residuals | 168 | 138,555,699 | 824,736 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mafouasson, H.N.A.; Gracen, V.; Yeboah, M.A.; Ntsomboh-Ntsefong, G.; Tandzi, L.N.; Mutengwa, C.S. Genotype-by-Environment Interaction and Yield Stability of Maize Single Cross Hybrids Developed from Tropical Inbred Lines. Agronomy 2018, 8, 62. https://doi.org/10.3390/agronomy8050062
Mafouasson HNA, Gracen V, Yeboah MA, Ntsomboh-Ntsefong G, Tandzi LN, Mutengwa CS. Genotype-by-Environment Interaction and Yield Stability of Maize Single Cross Hybrids Developed from Tropical Inbred Lines. Agronomy. 2018; 8(5):62. https://doi.org/10.3390/agronomy8050062
Chicago/Turabian StyleMafouasson, Hortense Noëlle Apala, Vernon Gracen, Martin Agyei Yeboah, Godswill Ntsomboh-Ntsefong, Liliane Ngoune Tandzi, and Charles Shelton Mutengwa. 2018. "Genotype-by-Environment Interaction and Yield Stability of Maize Single Cross Hybrids Developed from Tropical Inbred Lines" Agronomy 8, no. 5: 62. https://doi.org/10.3390/agronomy8050062
APA StyleMafouasson, H. N. A., Gracen, V., Yeboah, M. A., Ntsomboh-Ntsefong, G., Tandzi, L. N., & Mutengwa, C. S. (2018). Genotype-by-Environment Interaction and Yield Stability of Maize Single Cross Hybrids Developed from Tropical Inbred Lines. Agronomy, 8(5), 62. https://doi.org/10.3390/agronomy8050062