Abstract
Traditional rice varieties maintained and cultivated by farmers are likely sources of germplasm for breeding new rice varieties. They possess traits potentially adaptable to a wide range of abiotic and biotic stresses. Characterization of these germplasms is essential in rice breeding and provides valued information on developing new rice cultivars. In this study, 307 traditional rice varieties newly conserved at the PhilRice genebank were characterized to assess their phenotypic diversity using 57 morphological traits. Using the standardized Shannon-Weaver diversity index, phenotypic diversity indices averaged at 0.73 and 0.45 for quantitative and qualitative traits, respectively. Correlation analyses among agro-morphological traits showed a high positive correlation in some traits such as culm number and panicle number, flag leaf width and leaf blade width, grain width and caryopsis width. Cluster analysis separated the different varieties into various groups. Principal component analysis (PCA) showed that seven independent principal components accounted for 74.95% of the total variation. Component loadings for each principal component showed morphological characters, such as culm number, panicle number and caryopsis ratio that were among the phenotypic traits contributing positive projections in three principal components that explained 48% of variation. Analyses of results showed high diversity in major traits assessed in farmers’ rice varieties. Based on plant height and maturity, 11 accessions could be potential donor parents in a rice breeding program. Future collection trips and characterization studies would further enrich diversity, in particular traits low in diversity, such as anthocyanin coloration, awn presence, awn color, culm habit, panicle type and panicle branching.
1. Introduction
Rice is a major crop in the Philippines with a production of 16.7 million MT ranking the Philippines eighth in the world [1]. It is a major calorie source for most Filipinos and a major income source for 12 million farmers and their families [2]. Rice production contributes 2.2% to the Gross Domestic Product (GDP) of the country [3]. It is also a culturally important crop to Filipinos as rice is featured in many festivals and rituals [4,5]. The Executive Order No. 1061 in 1985 acknowledged the importance of rice to the country’s economy and livelihood by creating the Philippine Rice Research Institute (PhilRice) mandated to lead the country’s rice research and development programs [6]. In the Philippines, the growth of the rice sector is highly dependent on yield improvements, which can be achieved through breeding new varieties and developing and promoting yield-enhancing technologies [2].
Rice breeders are constantly engaged in developing new rice varieties with higher yield potential to enhance the actual yield obtained by farmers in the field. One approach in plant breeding, proposed as early as 1968 is new ideotypes development [7]. This “plant-type concept of breeding” resulted from pioneering studies showing close associations between yield and certain morphological characters in response to nitrogen application [8,9]. In 1966, the selection for the semi-dwarf rice plant type led to the release of the first modern high yielding variety, IR8, which commenced the “green revolution” in Asia [8]. After 28 years of successful release of the IR8, a yield plateau was observed and prompted rice breeders to propose a new plant type (NPT) during the International Rice Research Institute (IRRI) strategic planning workshop in 1993 [8,10]. As such, morphological characters rather than physiological traits were considered for NPT rice because they were easy to distinguish in a breeding program [11].
Rice genetic resources are key components to breeding programs, and farmers have played important roles in contributing to rice diversity by developing and nurturing thousands of rice varieties for several years [12]. This vast wealth of rice germplasm including landraces and traditional varieties is a good source of important alleles to develop new rice varieties. These germplasms serve as the foundation of any rice breeding program because they are the source of important traits necessary for improving and developing new breeds of rice varieties [13]. Several reports have shown the utilization of rice landraces in developing new varieties. IR8, dubbed a miracle rice [14], was the product from crosses between two landraces: a semi dwarf rice Dee-geo-woo-gen and tall, vigorous rice Peta [15]. The submergence tolerance SUB1 QTL was identified from submergence tolerant rice landrace FR13A. Its identification and characterization led to successful introgression of the QTL to rice mega-varieties [16]. Recently, the NAL1 allele that was identified from tropical Japonica rice landrace Daringan significantly increased the yield of modern rice cultivars [17].
Characterization of rice germplasms increases its utility in any breeding program. The use of agro-morphological traits is the most common approach utilized to estimate relationships between genotypes [18]. This approach was employed to assess diversity on ancestral lines of improved rice varieties in the Philippines [13], the indigenous rice in Yunnan, China [19] and the rice landraces in Nepal [18]. The conservation and characterization of these genetic resources is a necessity not only for posterity, but also for utilization in different improvement programs such as breeding for improved yield and tolerance to various stresses. It is important to assess the diversity of these germplasms materials to provide insights in the diversity of these germplasms.
Thus, this study assessed the phenotypic diversity of new rice germplasm farmers’ varieties conserved at the PhilRice genebank. Information generated from phenotyping these germplasms can be used as basis for future collection trips to augment diversity in the genebank collections as well as baseline information for utilization in rice breeding programs.
2. Results and Discussion
2.1. Germplasm Characterization
2.1.1. Diversity in Qualitative Traits
Phenotyping is an important activity to evaluate the utilization of the germplasm collection in a genebank. In this study, 307 traditional rice varieties recently conserved at the PhilRice genebank were scored and measured using 39 qualitative and 18 quantitative morphological characters. These germplasms were comprised of 215 Indica, 89 Javanica and three Japonica varieties (appendix Table A1). Among the qualitative characters scored, ligule shape and culm kneeling ability were observed invariants. All the germplasm characterized had a two-cleft ligule shape and the culms had no kneeling ability (Table 1). Twenty of the qualitative traits scored were dominated by one character in each trait with a distribution ranging between 76%–95%. As a result, these twenty agronomic traits had low diversity indices ranging between 0.12–0.45. These were mainly awn-related characters such as presence, color, distribution, and type. Awn color and panicle were the lowest calculated indices (H′ = 0.12) because 95% of the varieties scored had no awn and 92% had a medium length panicle type. Most of the varieties had thick culms and an erect culm habit. Moderately diverse traits were observed for 15 descriptors with indices ranging between 0.46–0.74. Most of these traits were inflorescence-related traits such as panicle and spikelet characters. Diversity in caryopsis pericarp color (seed coat color) (Figure 1) was also evident with all states being represented in the rice varieties evaluated. White seed coat color was the predominant state (50.5%), followed by red seed coat color (30.0%), variable purple (5.9%), light brown (4.6%) and purple (4.2%).
Four of the 39 traits scored had a high diversity with an average index of 0.87. Two of these traits were culm-related which assessed rice sturdiness during maturity and harvest. Although the predominant character was intermediate lodging resistance, 37% of the rice varieties had strong to very strong lodging resistance at the mature stage. The endosperm type trait had the highest calculated diversity index of 0.99. The reason for this is that all the endosperm type descriptors (1 = non-glutinous, 2 = Intermediate, 3 = glutinous) were identified in the characterized germplasm.

Table 1.
Qualitative traits showing the predominant state observed, distribution (%) and the calculated Shannon diversity indices (H′) for each descriptor scored.
| Descriptor | Predominant State | % | States Observed | H′ Index |
|---|---|---|---|---|
| Invariant | ||||
| Ligule Shape | 2-ccenter | 100.00 | 1 | 0.00 |
| Culm Kneeing Ability | Absent | 100.00 | 1 | 0.00 |
| Low diversity | ||||
| Awn Color (Late) | Awnless | 95.11 | 4 | 0.12 |
| Panicle Type | Medium (~25 cm) | 91.86 | 3 | 0.12 |
| Awn Color (Early) | Awnless | 91.53 | 6 | 0.18 |
| Awn Distribution | Awnless | 91.53 | 5 | 0.20 |
| Culm Diameter Type | Thick | 96.42 | 2 | 0.22 |
| Awn Type | Awnless | 91.21 | 5 | 0.22 |
| Panicle Secondary Branching | Sparse | 89.58 | 3 | 0.25 |
| Culm Habit/Angle | Erect (<15°) | 85.67 | 4 | 0.26 |
| Sterile Lemma Type | Medium | 62.54 | 3 | 0.27 |
| Culm Anthocyanin Coloration on Nodes | Absent | 90.55 | 4 | 0.28 |
| Awn Presence | Absent | 91.53 | 3 | 0.29 |
| Leaf Blade Pubescence | Intermediate | 91.86 | 3 | 0.29 |
| Flag Leaf Attitude/Angle | Erect | 83.71 | 4 | 0.30 |
| Leaf Blade Attitude | Erect | 89.90 | 3 | 0.31 |
| Sterile Lemma Color | Straw | 88.27 | 4 | 0.32 |
| Panicle Attitude | Drooping | 84.36 | 5 | 0.37 |
| Stigma Color | White | 77.85 | 4 | 0.41 |
| Auricle Color | Whitish | 75.90 | 6 | 0.45 |
| Moderate diversity | ||||
| Lemma & Palea Color (Late Observation) | Straw | 66.45 | 8 | 0.46 |
| Panicle Attitude of Main Axis | Slightly drooping | 70.03 | 3 | 0.49 |
| Leaf Blade Length Type | Intermediate (~50 cm) | 60.59 | 3 | 0.54 |
| Culm Underlying Node Color | Green | 73.29 | 3 | 0.56 |
| Leaf Blade Width Type | Intermediate | 65.15 | 2 | 0.59 |
| Flag Leaf Attitude (Late Measurement) | Descending | 71.01 | 4 | 0.63 |
| Culm Length Type | Intermediate to long | 36.81 | 6 | 0.63 |
| Culm Number Type | Intermediate (~15 culms) | 75.24 | 3 | 0.65 |
| Late Lemma Apiculus Color | Straw | 47.56 | 9 | 0.65 |
| Lemma & Palea Pubescence | Short hairs | 60.91 | 5 | 0.66 |
| Panicle Exsertion | Well exerted | 55.37 | 5 | 0.67 |
| Caryopsis Pericarp Color (Seed Coat Color) | White | 50.49 | 7 | 0.68 |
| Caryopsis Shape | Long spindle-shaped | 43.97 | 5 | 0.71 |
| Productivity | Intermediate | 70.03 | 3 | 0.71 |
| Apiculus Shape | Curved | 78.83 | 2 | 0.74 |
| High diversity | ||||
| Early Lemma Apiculus Color | Straw | 26.71 | 8 | 0.79 |
| Culm Lodging Resistance | Intermediate | 42.02 | 5 | 0.84 |
| Culm Strength | Intermediate | 41.04 | 5 | 0.84 |
| Endosperm Type | Non-glutinous | 41.37 | 3 | 0.99 |
| Average diversity | 0.45 | |||
Overall, the diversity in qualitative traits was low with an average index of 0.45. Several traits that were classified as low diversity might be prioritized in future collection trips to enhance their diversity in our genebank. Although not all these traits are linked to yield, some traits such as panicle and culm diameter types are useful parameters for improving yield. Wu et al., have shown that large culm rice varieties have a higher number of grains per panicle and a longer spike length [20].

Figure 1.
Diversity in grain color and caryopsis pericarp color of the different traditional rice varieties screened. Numbers in parenthesis are the collection numbers of the rice germplasm.
2.1.2. Diversity in Quantitative Traits
The 307 varieties showed diverse phenotypes in terms of plant height, leaf blade, flag leaf culm number and panicle number, among others. Table 2 sums up the quantitative morphological characters showing the highest and lowest values measured for each character. Most of the traits had moderate (5) to high (12) diversity indices. Awn length was the only trait that showed low diversity (H′ = 0.02). This could be attributed to only 14 varieties exhibiting awns in their grains. Traits with moderate diversity included caryopsis (length, width and ratio) and culm descriptors (diameter and number). Culm number could be associated with yield if all tillers produced inflorescence. Culm strength and culm lodging resistance had diversity indices of 0.84. Maturity of characterized germplasm ranged from 71 to 154 days. Rice variety Inuway (CollNo. 10869) had the shortest maturity. This variety was collected in an upland ecosystem in the Aurora province. Plant height varied from 68 cm to 161 cm, observed in Kinakaw (CollNo. 10857) and Dinorado (CollNo. 11049), respectively. Dinorado is an upland variety popular in the Arakan Valley of North Cotabato known for its sweet aroma, pinkish grain and good eating quality [4]. Additionally, the majority of the farmers’ varieties (82%) were >100 cm tall with an average height of 116 cm and a median of 117 cm contradicting breeders’ preference of 90–100 cm tall rice varieties among other characteristics to serve as potential donor for NPT breeding program [6].
Table 2.
Quantitative descriptors and calculated Shannon-Weaver index (H′) of evaluated rice varieties, and calculated numbers enclosed in parenthesis are the collection numbers of the rice germplasm.
| Descriptors | H′ | Min Trait Value | Variety | Max Trait Value | Variety | Mean Trait Value (± Standard Deviation) |
|---|---|---|---|---|---|---|
| Low Diversity | ||||||
| Awn Length (mm) | 0.02 | 1.98 | Puchagwan (11241) | 69.64 | Burdagol (11083) | 0.80 (6.01) |
| Moderate Diversity | ||||||
| Caryopsis Length (mm) | 0.64 | 4.37 | Milagrosa (11102) | 13.72 | Binaka (10838) | 6.49 (1.04) |
| Caryopsis Width (mm) | 0.70 | 4.15 | Kaimpas (11315) | 1.28 | Doriat Pula (10934) | 2.23 (0.37) |
| Culm Diameter at Basal Internode (mm) | 0.72 | 0.58 | Fancy-1 (11105) | 12.40 | Speaker (11080) | 8.06 (1.92) |
| Culm Number | 0.73 | 5.20 | Lubang (11341) | 34.80 | Rc 18 (Pula) (11312) | 16.30 (4.48) |
| Caryopsis Length/Width Ratio Score | 0.74 | 1.42 | Kaimpas (11315) | 5.12 | Binaka (10838) | 2.98 (0.60) |
| High Diversity | ||||||
| Panicle Length (cm) | 0.77 | 20.90 | Pinarompong (10932) | 38.80 | Unoy (Umangan) (11245) | 27.27 (2.67) |
| Maturity (day) | 0.77 | 71.00 | Inuway (10869) | 154.00 | 18-Pula (11061); Galo (11205) | 106.84 (35.69) |
| Grain Width (mm) | 0.78 | 1.51 | Duriat (10943) | 6.56 | Kipil (10837) | 2.58 (0.53) |
| Panicle Number Per Plant | 0.78 | 5.20 | Lubang (11341) | 34.80 | Rc 18 (Pula) (11312) | 16.26 (4.46) |
| Flag Leaf Length (cm) | 0.79 | 23.80 | Bolinao (11256) | 72.80 | Kinakaw (10857) | 38.57 (7.56) |
| Leaf Blade Length (cm) | 0.80 | 34.80 | C-4 Dinorado (11317) | 88.40 | Lubang (11356) | 58.36 (9.13) |
| Sterile Lemma Length (mm) | 0.81 | 1.56 | M10-2 (11086) | 3.86 | Minindoro (Pula) (10844) | 2.47 (0.39) |
| 100 Grain Weight (g) | 0.81 | 1.10 | Kinakaw (10857) | 5.50 | Binaka (10810) | 2.45 (0.54) |
| Culm Length (cm) | 0.82 | 67.80 | Kinakaw (10857) | 160.60 | Dinorado (11049) | 116.03 (15.67) |
| Grain Length (mm) | 0.84 | 5.29 | Dinorado (10860) | 10.76 | Binaka (10810) | 8.48 (1.05) |
| Flag Leaf Width (cm) | 0.85 | 1.10 | Elon-Peta (11346) | 2.60 | Binisaya (10826) | 1.95 (0.32) |
| Leaf Blade Width (cm) | 0.85 | 0.96 | Rc 18 (Pula) (11312) | 2.48 | Binisaya (10826) | 1.64 (0.30) |
| Average Diversity | 0.73 | |||||
Grain trait diversity was also observed in the germplasm (Figure 1). Of the 307 accessions, only 14 rice varieties had awns present on the grains with their lengths ranging from 2 mm to 70 mm with the variety Burdagol (CollNo. 11083) having the longest awn. The presence of awns is considered an important trait in rice domestication. Grains of wild rice have long awns that protect the grains from animal pilfering. Some reports suggest that the presence of awns in grains aids bird resistance in agricultural crops [21,22]. Early studies in sorghum breeding have shown that varieties with long awns or that are strongly awned are more resistant to bird attacks than varieties with no awns [23]. Awned grains along those with few tillers and long panicles were found to be the characteristics of the bulu or Javanica group within the tropical Japonica varieties [24]. On the other hand, cultivated rice varieties have short awns allowing for easier harvesting than varieties with long awns [25]. Low tillering capability (three to four tillers) was one of the criteria used by IRRI rice breeders in selecting donor parents to be used in developing NPTs of rice [8]. Tillering ability among the 307 farmers’ rice varieties ranged from five to 35 tillers. Rc 18-Pula (CollNo. 11312) showed the highest tillering ability among the farmers’ varieties. It is a red-coated rice variety collected in the Bohol province and considered one of the popular varieties in the area based on Bertuso’s survey [26]. This variety is a rice farmer’s selection in 1997 [26] from his field planted with PSB Rc 18 (has a white seed coat), a modern rice variety released in 1994 for irrigated rice ecosystem [2]. Sturdy culm was another criterion used for donor parent selection for NPTs. The majority of the germplasms characterized (96%) had thick culms with a ≥5 mm culm diameter. However, when rice varieties were assessed for lodging resistance, only 37% showed strong or very strong lodging resistance. This could be attributed to the plant height, a major factor in lodging resistance in rice [27]. Having a short plant structure is currently the preferred trait for improving lodging resistance in rice [28].
In general, diversity in quantitative traits was moderate with an average index of 0.73. Nearly all the traits measured showed moderate to high diversity.
2.2. Correlation among Traits
Using Pearson’s product-moment correlation, an analysis was done to assess the relationship among the morphological traits. It is useful to determine the relationship among the morphological traits since this information will be useful in the utilization of the germplasm as well in the collection of the germplasm based on the target traits. Several traits showed significant correlations (r = 0.195; p < 0.05) among each other. A heat map (Figure 2) was constructed to visualize the traits that had weak (r ≤ 0.35), moderate (r = 0.36–0.67) and strong (r = 0.68–1.00) correlations [29]. An analysis showed that 89% of the trait combinations had weak correlations while 10% had moderate correlations. Only the correlation between the panicle number per plant and the culm number (r = 0.998) was strong. This showed that all tillers were productive tillers and able to bear inflorescence. The panicle number per plant ranged from five to 35. Sterile lemma length and 100-grain weight showed a moderate correlation (r = 0.50). This was expected since any increase in sterile lemma length would positively affect the grain weight. A high correlation was also observed between the flag leaf width and leaf blade width (r = 0.59) indicating that an increase in leaf blade width might also result in an increase of flag leaf width. A positive correlation was also observed between flag leaf width and grain width (r = 0.40). Flag leaves are important in grain filling, as 80% of the total carbohydrate stored in the grains is produced by the top two leaves in rice [30].
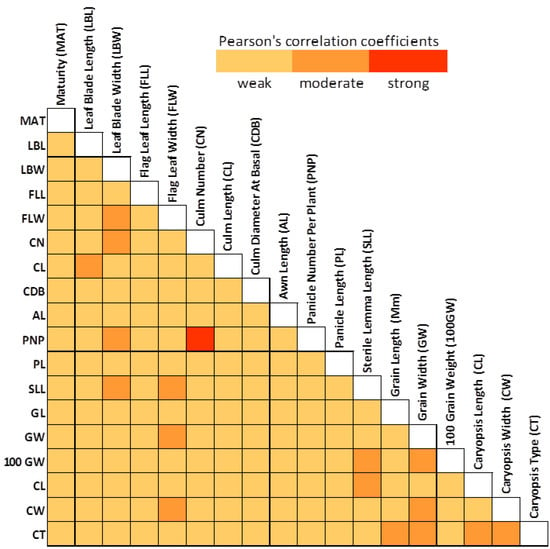
Figure 2.
Heat map showing calculated Pearson’s product-moment correlation coefficients among morphological traits measured in all germplasms screened. Correlation coefficients were classified as weak (r ≤ 0.35), moderate (r ≥ 0.36) and strong (r > 0.68) [29].
These characteristics are essential to rice breeders as it has been demonstrated that the flag leaf area increased grain yield by increasing the number of spikelets per panicle [31]. Flag leaves were reported to be the major source of phloem-delivered photoassimilates during the grain-filling stage in rice [32]. Previous studies have shown that cutting of flag leaves could result to up to 45% grain yield loss [33]. A recent review by Biswal and Kohli outlined the importance of flag leaf traits in cereal breeding for drought tolerance [34]. Flag leaf sheath is one of the main sources of carbohydrate for rice grain filling under drought condition [35].
Although the correlation analyses showed only one combination trait that had a strong correlation, there were 16 combination traits that had moderate correlations. These traits that had moderate to high correlations could be used as a basis for the utilization of these sets of germplasm for breeding purposes as well as for planning future collection trips targeting specific traits. Trait correlations can be used by breeders either to simultaneously improve correlated traits or reduce undesirable side effects when trying to improve only one of the correlated traits [36].
2.3. Cluster and Principal Component Analyses of Rice Germplasm
The relationship among the 307 farmers’ rice varieties as revealed by Unweighted Pair Group Method with Arithmetic Mean (UPGMA) cluster analysis is shown in Figure 3. Truncating the tree at the Euclidean distance of 1.13 resulted in 24 clusters. In the truncated tree, 10 clusters had single accession, another 10 clusters had two to ten accessions; three clusters had 23–50 accessions and one big cluster had 137 accessions. The fact that 83% of the clusters formed contained one to a few accessions implied a diversity in the collection. Among the single-accession clusters, most accessions were collected from the Palawan and Kalinga provinces. The majority of these germplasms were of the Indica type while two accessions belonged to Javanica. These single-accession clusters were considered distinct from each other and the rest of the clusters. Cluster 3 (variety C-4 Dinorado; CollNo. 11317), for example, had similar traits to the accessions in Cluster 2 for most traits, such as leaf blade width type, flag leaf width, type, and erect flag leaf attitude at early stage, but differed by having very short leaf blades and horizontal flag leaf attitude at a late stage. Similarly, the variety Kinakaw (CollNo. 10857, Cluster 24) was distinct from the rest of the clusters because it had a short plant stature, the longest flag leaf length and the lightest 100-grain weight. Other single-accession clusters such as Cluster 21 (Burdagol CollNo. 11083), Cluster 22 (variety Chay-ot; CollNo. 11246) and Cluster 23 (variety Ifo; CollNo. 11255) were peculiar because of their long awns (30–70 mm). Most of the accessions (90%) had no awns while the rest had short awns (2–16 mm). Variety Benangkar (CollNo. 10923) was another single-accession cluster, which was characterized by the presence of purple lines in its culm nodes, which is a trait not very common among the rest of the accessions screened. Cluster 1, one of the four big clusters, was characterized as having a short plant stature (average height of 94.8 cm) compared to Clusters 2 (110.53 cm) and 4 (121.03 cm). Most of the accessions in Cluster 2 belonged to the Indica type (90%).
Overall, cluster analysis provided an insight into the diversity of the collections as shown by the number of clusters formed with one to 10 members when the dendrogram was truncated at 1.13 distance. A distinct variety was separated from the rest of the germplasm pool as exemplified by the single-accession clusters.
Principal component analysis (PCA) was employed to reduce the complexity of the data set while retaining the variation within the data set as far as possible [37]. The PCA resulted in 18 independent principal components that had a cumulative explained variance of 100% (Table 3). Following the Proportion of Variance Criterion [38], seven principal components (PCs) were retained that had a cumulative variance of 75%. The first component accounted for 22.5% of the total variation in the data set while the second and third principal components contributed 14.5% and 10.6%, respectively. Together, these three components could explain 47.6% of the total variation in the characterized rice germplasm. Analysis of the factor loadings of the characters in the retained PCs showed that phenotypic traits that contributed to yield showed high positive loadings in PC 1 (Table 4). These traits were culm number, panicle number per plant and caryopsis ratio score with factor loadings of 0.649, 0.651 and 0.529, respectively. These three morphological characters could have contributed to the maximum variability in PC 1 which explained 22.5% of the total variation in the data set. Among these three traits, only panicle number per plant was classified as high diversity while the other traits had moderate diversity indices. In PC 2, leaf blade length presented the highest factor loading of 0.482. This showed that leaf blade length was the major morphological character that contributed to the variation in PC 2 which explained 14.5% of the variation. In PC 3, plant height (culm length) showed a high loading of 0.454.

Figure 3.
Dendrogram generated by cluster analysis of morphological characters using Unweighted Pair Group Method with Arithmetic Mean (UPGMA).
Table 3.
Computed eigenvalues of the different principal components with corresponding proportion and cumulative explained variance.
| Components | Eigenvalue | Explained Variance | |
|---|---|---|---|
| Percent | Cumulative | ||
| 1 | 4.04 | 22.46 | 22.46 |
| 2 | 2.61 | 14.52 | 36.98 |
| 3 | 1.91 | 10.61 | 47.59 |
| 4 | 1.61 | 8.97 | 56.56 |
| 5 | 1.26 | 7.01 | 63.58 |
| 6 | 1.05 | 5.84 | 69.41 |
| 7 | 1.00 | 5.53 | 74.95 |
| 8 | 0.82 | 4.57 | 79.52 |
| 9 | 0.71 | 3.96 | 83.48 |
| 10 | 0.64 | 3.53 | 87.01 |
| 11 | 0.57 | 3.19 | 90.21 |
| 12 | 0.49 | 2.75 | 92.95 |
| 13 | 0.41 | 2.30 | 95.26 |
| 14 | 0.33 | 1.85 | 97.11 |
| 15 | 0.29 | 1.61 | 98.72 |
| 16 | 0.21 | 1.16 | 99.87 |
| 17 | 0.02 | 0.12 | 99.99 |
| 18 | 0.00 | 0.01 | >100% |
Table 4.
Factor loadings (eigenvectors) for the different morphological characters for the principal components retained.
| Descriptors | Principal Components | ||||||
|---|---|---|---|---|---|---|---|
| PC 1 | PC 2 | PC 3 | PC 4 | PC 5 | PC 6 | PC 7 | |
| Maturity (day) | 0.33 | 0.29 | 0.34 | 0.10 | 0.40 | 0.23 | 0.23 |
| Leaf Blade Length (mm) | −0.11 | 0.48 | 0.53 | 0.14 | 0.12 | −0.08 | −0.31 |
| Leaf Blade Width (mm) | −0.67 | −0.21 | −0.04 | −0.26 | −0.25 | 0.25 | 0.04 |
| Flag Leaf Length (mm) | −0.17 | 0.21 | 0.43 | 0.46 | −0.11 | 0.13 | −0.26 |
| Flag Leaf Width (mm) | −0.73 | −0.12 | −0.01 | 0.00 | −0.14 | 0.21 | 0.21 |
| Culm Number | 0.65 | −0.28 | −0.36 | 0.52 | −0.08 | 0.17 | −0.09 |
| Culm Length (cm) | −0.34 | 0.37 | 0.45 | 0.11 | −0.14 | 0.33 | 0.02 |
| Basal Culm Diameter (mm) | −0.33 | −0.33 | −0.01 | 0.15 | −0.32 | 0.54 | 0.17 |
| Awn Length (mm) | −0.11 | −0.01 | 0.06 | 0.22 | −0.54 | −0.57 | 0.41 |
| Panicle Number/plant | 0.65 | −0.28 | −0.36 | 0.51 | −0.08 | 0.18 | −0.10 |
| Panicle Length (cm) | −0.27 | 0.01 | 0.33 | 0.59 | −0.27 | −0.20 | 0.01 |
| Sterile Lemma Length (mm) | −0.51 | −0.67 | 0.10 | −0.12 | 0.18 | −0.03 | −0.08 |
| Grain Length (mm) | 0.06 | −0.47 | 0.37 | 0.26 | 0.46 | −0.06 | 0.46 |
| Grain Width (mm) | −0.66 | 0.13 | −0.29 | 0.34 | 0.39 | −0.09 | 0.10 |
| 100-Grain Weight (g) | −0.51 | −0.46 | −0.03 | 0.28 | 0.29 | −0.06 | −0.02 |
| Caryopsis Length (mm) | −0.10 | −0.73 | 0.31 | −0.08 | −0.05 | −0.14 | −0.43 |
| Caryopsis Width (mm) | −0.72 | 0.08 | −0.38 | 0.18 | 0.10 | −0.13 | −0.33 |
| Caryopsis Length/Width Ratio Score | 0.53 | −0.60 | 0.52 | −0.18 | −0.10 | −0.02 | −0.05 |
3. Experimental Section
3.1. Germplasm Characterization
A total of 307 rice germplasms conserved at the Philippine Rice Research Institute (PhilRice) genebank which were collected from different parts of the country (Figure 4) were used to evaluate the genetic diversity of farmers’ traditional rice varieties and landraces. Characterization of the collected rice germplasms was done at the at the PhilRice Central Experimental Station, Maligaya, Science City of Munoz, Nueva Ecija (15°40′ N, 120°53′ E, 57.6 masl) during the wet cropping season (WS) in 2009. Prior to sowing, seeds were incubated in the oven set at 37 °C for 12 h to break inherent seed dormancy. Seeds were sown in a raised seedbed in the greenhouse and covered with coconut coir dust. Seven-day-old seedlings were transplanted into the field following a planting distance of 25 cm × 25 cm. A total of 20 plants per accession in three replicates were planted in the field for characterization. Agronomic characters were measured and scored following the Rice Descriptors [39]. A total of 18 quantitative and 39 qualitative characters were selected to score the germplasm collections.
Figure 4.
Philippine map showing the provinces (areas in green) where the rice germplasm were collected. The map was generated using DIVA-GIS ver.7.5.0 [40].
3.2. Data Analyses
Descriptive statistics was done using PROC UNIVARIATE procedure in SAS ver. 9.3 (SAS Institute, Cary, NC, USA [41]). Correlations among morphological traits measured were analyzed using PROC CORR procedure in SAS. Shannon-Weaver diversity index (H′) was used to calculate the phenotypic diversity of the characterized farmers’ varieties following the protocol used by Sotto and Rabara [42]. An arbitrary scale was adapted from Jamago and Cortes [43] to categorize the computed indices into maximum (H′ = 1.00), high (H′ = 0.76–0.99), moderate (H′ = 0.46–0.75) and low diversity (0.01–0.45). Diversity indices of collected germplasm were calculated based on phenotypic frequency using standardized Shannon-Weaver Diversity index formula:
wherein
H′ = −Ʃpi(log2pi)/log2N
pi = frequency proportion of the descriptor state
N = number of states
The standardized Shannon-Weaver provided a constrained index between zero and one with the highest value indicating maximum abundance [44,45]. Multivariate statistical analyses of characterization data were conducted using principal component (PCA) and cluster (CA) analyses. PCA was employed to identify the different morphological characters that contributed to the most variance in the measured variables. In PCA, the raw data were standardized and the distance matrix using the variance-covariance coefficients was computed. The Proportion of Variance criterion was used to identify the different principal components that contributed to the total variance in the dataset [23]. PCA and CA were done using NTSYSpc version 2.1 software [46]. The distance matrix was generated using the Euclidean Distance Coefficients and was used as input for clustering using the unweighted pair group of arithmetic means (UPGMA) method.
4. Conclusions
Phenotyping of germplasm materials is an important undertaking in genetic resource conservation to ensure efficient conservation management as well as its effective utilization especially in breeding programs. In this study, 307 rice varieties were characterized to assess their phenotypic diversity. Diversity analyses showed that 46% of the qualitative traits scored had low diversity indices compared to only 5% in quantitative traits. Overall, the rice germplasm showed moderate diversity based on quantitative characters (average index of 0.73). In contrast, the qualitative characters had a low diversity (average index of 0.45). In comparison to other landraces collections, the diversity in qualitative traits of our germplasm was higher than what Bajracharya et al., observed in the landraces from Nepal [18]. The complete qualitative data set of our collection is available in appendix Table A1.
In order to enrich the diversity of qualitative traits in our collection at the PhilRice genebank exploration trips may be conducted. Collection gaps should be identified in the genebank’s germplasm collection and should be prioritized in future collection trips. An emphasis on farmers’ varieties that have been planted in a community for several generations should be considered when planning for collection trips. It should also considered to look for diversity in qualitative characters that have shown low diversity indices such as presence of awns, panicle type, and culm habit type among other characters. Other factors that can be considered during collection of these rice germplasms are the characters that have high correlations with other characters. Collecting diversity for certain traits or characters could also lead to a high diversity if the characters are highly correlated.
The success of any conservation program could also be measured by the amount of which these genetic resources are being utilized. Data generated from characterization of these 307 rice varieties can be utilized as baseline information for the utilization of these germplasms for any rice breeding program such as breeding for ideoypes. For example, 37 accessions meets the plant height criterion (90–100 cm) for NPT among which 11 accessions also meets the maturity requirement (110–120 days). Short stature variety was one of the criteria to select a variety for use in a breeding program to address lodging. The optimal growth duration to achieve maximum yield is about 120 days [8]. A linear increase in total biomass had been observed when the growth duration was increased from 95 days to 135 days [47]. Efforts should be invested in promoting various stakeholders in rice production to utilize these germplasms. Effective utilization of these germplasms can be enhanced if these materials are fully characterized and evaluated for their potential use in breeding programs. Evaluation of these germplasms should be conducted to assess their potential as donor parents for the breeding of new varieties with improved responses to various abiotic and biotic stresses. Broadening of the genetic base through utilization of diverse germplasms in breeding for new rice varieties may be able to break the yield barrier that rice breeders are currently trying to address.
Acknowledgments
We would like to acknowledge the assistance of Rodel Valdez, Edgar Vallejo, Daisy Villanueva, Jackie Reyes and Robert Punzal in field preparation, characterization and maintenance of the germplasm in the field. We also appreciate the assistance of Jennifer Jara-Rabara for editing the manuscript. This study was supported by PhilRice grant No. FEP02-030 to RCR.
Author Contributions
Roel C. Rabara designed the experiment, analyzed the data and written the manuscript. Marilyn C. Ferrer conducted the phenotyping experiments, assisted in data analysis and contributed in writing the manuscript. Celia L. Diaz contributed in phenotyping the germplasm. Ma. Cristina V. Newingham contributed in data encoding and database management. Gabriel O. Romero contributed in writing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAOSTAT, Classic Version; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. Available online: http://faostat.fao.org/site/339/default.aspx (accessed on 5 August 2013).
- Sebastian, L.S.; Alviola, P.A.; Francisco, S.R. Bridging the rice yield gap in the Philippines. In Bridging the Rice Yield Gap in the Asia-Pacific Region; Papademetriou, M.K., Dent, F.J., Herath, E.M., Eds.; FAO Regional Office for Asia and the Pacific: Bangkok, Thailand, 2000; p. 13. [Google Scholar]
- Cororaton, C.B. Philippine Rice and Rural Poverty: An Impact Analysis of Market Reform Using CGE; International Food Policy Research Institute (IFPRI): Washington, DC, USA, 2006. [Google Scholar]
- Reyes, L.C. Banking seeds. Rice Today 2012, 11, 16–19. [Google Scholar]
- Aguilar, F.V. Rice in the Filipino Diet and Culture; Philippine Institute for Development Studies: Makati, Philippines, 2005. [Google Scholar]
- Establishing the Philippine Rice Research Institute (PRRI). Executive Order 1061; PRRI: Manila, Philippines, 1985. [Google Scholar]
- Donald, C.M. The breeding of crop ideotypes. Euphytica 1968, 17, 385–403. [Google Scholar] [CrossRef]
- Peng, S.; Khush, G.; Cassman, K. Evolution of the new plant ideotype for increased yield potential. In Breaking the Yield Barrier Proceedings of a Workshop on Rice Yield Potential in Favourable Environments; International Rice Research Institute: Manila, Philippines, 1994. [Google Scholar]
- Yoshida, S. Physiological aspects of grain yield. Annu. Rev. Plant Physiol. 1972, 23, 437–464. [Google Scholar] [CrossRef]
- Tonini, A.; Cabrera, E. Opportunities for Global Rice Research in a Changing World; International Rice Research Institute: Los Baños, Philippines, 2011. [Google Scholar]
- Peng, S.; Khush, G.S.; Virk, P.; Tang, Q.; Zou, Y. Progress in ideotype breeding to increase rice yield potential. Field Crops Res. 2008, 108, 32–38. [Google Scholar] [CrossRef]
- Bellon, M.; Pham, J.; Jackson, M. Genetic conservation: A role for rice farmers. In Plant Genetic Conservation: The in situ Approach; Maxted, N., Ford-Lloyd, B., Hawkes, J., Eds.; Chapman and Hall: London, UK, 1997; pp. 263–289. [Google Scholar]
- Caldo, R.; Sebastian, L.; Hernandez, J. Morphology-based genetic diversity analysis of ancestral lines of Philippine rice cultivars. Philipp. J. Crop Sci. 1996, 21, 86–92. [Google Scholar]
- Ronald, P. A case study of rice from traditional breeding to genomics. In The Role of Biotechnology in a Sustainable Food Supply; Popp, J.S., Jahn, M.M., Matlock, M.D., Kemper, N.P., Eds.; Cambridge University Press: New York, NY, USA, 2012; p. 10. [Google Scholar]
- Hargrove, T.; Coffman, W.R. Breeding history. Rice Today 2006, 5, 34–38. [Google Scholar]
- Bailey-Serres, J.; Fukao, T.; Ronald, P.; Ismail, A.; Heuer, S.; Mackill, D. Submergence tolerant rice: SUB1’s journey from landrace to modern cultivar. Rice 2010, 3, 138–147. [Google Scholar] [CrossRef]
- Fujita, D.; Trijatmiko, K.R.; Tagle, A.G.; Sapasap, M.V.; Koide, Y.; Sasaki, K.; Tsakirpaloglou, N.; Gannaban, R.B.; Nishimura, T.; Yanagihara, S.; et al. NAL1 allele from a rice landrace greatly increases yield in modern Indica cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 20431–20436. [Google Scholar] [CrossRef]
- Bajracharya, J.; Steele, K.A.; Jarvis, D.I.; Sthapit, B.R.; Witcombe, J.R. Rice landrace diversity in Nepal: Variability of agro-morphological traits and SSR markers in landraces from a high-altitude site. Field Crops Res. 2006, 95, 327–335. [Google Scholar] [CrossRef]
- Yawen, Z.; Shiquan, S.; Zichao, L.; Zhongyi, Y.; Xiangkun, W.; Hongliang, Z.; Guosong, W. Ecogeographic and genetic diversity based on morphological characters of indigenous rice (Oryza sativa L.) in Yunnan, China. Genet. Resour. Crop Evol. 2003, 50, 567–577. [Google Scholar] [CrossRef]
- Wu, L.-L.; Liu, Z.-L.; Wang, J.-M.; Zhou, C.-Y.; Chen, K.-M. Morphological, anatomical, and physiological characteristics involved in development of the large culm trait in rice. Aust. J. Crop Sci. 2011, 5, 1356–1363. [Google Scholar]
- Bullard, R.; York, J. Breeding for bird resistance in sorghum and maize. In Progress in Plant Breeding-1; Russell, G., Ed.; Butterworths: London, UK, 1985; pp. 193–222. [Google Scholar]
- Bullard, R.W. Characteristics of Bird-resistance in Agricultural Crops; Denver Wildlife Research Center: Denver, CO, USA, 1988. [Google Scholar]
- Jowett, D. Breeding bird-resistant sorghum in East Africa. Plant Breed. Abstr. 1967, 37, 85. [Google Scholar]
- Vaughan, D.A.; Lu, B.-R.; Tomooka, N. The evolving story of rice evolution. Plant Sci. 2008, 174, 394–408. [Google Scholar] [CrossRef]
- Hu, G.; Zhang, D.; Pan, H.; Li, B.; Wu, J.; Zhou, X.; Zhang, Q.; Zhou, L.; Yao, G.; Li, J.; et al. Fine mapping of the awn gene on chromosome 4 in rice by association and linkage analyses. Chin. Sci. Bull. 2011, 56, 835–839. [Google Scholar] [CrossRef]
- Bertuso, A. Farmers’ Management of Rice Genetic Diversity: A Study on Enhancing Red Rices in Bohol, Philippines, in Department of Crop Science; Wageningen University: Wageningen, The Netherlands, 2000; p. 144. [Google Scholar]
- Hitaka, H. Studies on the lodging of rice plants. Jpn. Agric. Res. Q. 1969, 4, 1–6. [Google Scholar]
- Ookawa, T.; Hobo, T.; Yano, M.; Murata, K.; Ando, T.; Miura, H.; Asano, K.; Ochiai, Y.; Ikeda, M.; Nishitani, R.; et al. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 2010, 1, 1–11. [Google Scholar]
- Taylor, R. Interpretation of the correlation coefficient: A basic review. J. Diagn. Med. Sonogr. 1990, 6, 35–39. [Google Scholar] [CrossRef]
- Gladun, I.; Karpov, E. Production and partitioning of assimilates between the panicle and vegetative organs of rice after flowering. Russ. J. Plant Physiol. 1993, 40, 629–633. [Google Scholar]
- Yue, B.; Xue, W.Y.; Luo, L.J.; Xing, Y.Z. QTL analysis for flag leaf characteristics and their relationships with yield and yield traits in rice. Acta Genet. Sin. 2006, 33, 824–832. [Google Scholar] [CrossRef]
- Narayanan, N.N.; Vasconcelos, M.W.; Grusak, M.A. Expression profiling of Oryza sativa metal homeostasis genes in different rice cultivars using a cDNA macroarray. Plant Physiol. Biochem. 2007, 45, 277–286. [Google Scholar] [CrossRef]
- Abou-khalifa, A.A.B.; Misra, A.; Salem, A.E.-A.K. Effect of leaf cutting on physiological traits and yield of two rice cultivars. Afr. J. Plant Sci. 2008, 2, 147–150. [Google Scholar]
- Biswal, A.; Kohli, A. Cereal flag leaf adaptations for grain yield under drought: Knowledge status and gaps. Mol. Breed. 2013, 31, 749–766. [Google Scholar]
- Garcia, A.; Dorado, M.; Perez, I.; Montilla, E. Effect of water deficit on the distribution of photoassimilates in rice plants (Oryza sativa L.). Interciencia 2010, 35, 46–54. [Google Scholar]
- Chen, Y.; Lubberstedt, T. Molecular basis of trait correlations. Trends Plant Sci. 2010, 15, 454–461. [Google Scholar] [CrossRef]
- Ringnér, M. What is principal component analysis? Nat. Biotechnol. 2008, 26, 303–304. [Google Scholar]
- O’Rourke, N.; Hatcher, L. A Step-by-Step Approach to Using SAS for Factor Analysis and Structural Equation Modeling, 2nd ed.; SAS Institute: Cary, NC, USA, 2013. [Google Scholar]
- Bioversity International, International Rice Research Institute and WARDA Africa Rice Center. Descriptors for Wild and Cultivated Rice (Oryza spp.); Bioversity International; International Rice Research Institute: Rome, Italy, 2007. [Google Scholar]
- Hijmans, R.; Guarino, L.; Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Genet. Resour. Newsl. 2001, 127, 15–19. [Google Scholar]
- SAS Institute. I. What’s New in SAS 9.3; SAS Institute: Cary, NC, USA, 2012. [Google Scholar]
- Sotto, R.; Rabara, R.C. Morphological diversity of Musa balbisiana in the Philippines. J. Nat. Stud. 2007, 6, 37–46. [Google Scholar]
- Jamago, J.M.; Cortes, R.V. Seed diversity and utilization of the upland rice landraces and traditional varieties from selected areas in Bukidnon, Philippines. IAMURE Int. J. Ecol. Conserv. 2012, 4, 112–130. [Google Scholar]
- Pielou, E.C. An Introduction to Mathematical Ecology; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1969. [Google Scholar]
- Lexerød, N.L.; Eid, T. An evaluation of different diameter diversity indices based on criteria related to forest management planning. For. Ecol. Manag. 2006, 222, 17–28. [Google Scholar] [CrossRef]
- Rohlf, F. NTSYSpc:Numerical Taxonomy System; Version 2.1; Exeter Publishing, Ltd.: Setauket, New York, NY, USA, 2002. [Google Scholar]
- Akita, S. Improving yield potential in tropical rice. In Progressin Irrigated Rice Research; International Rice Research Institute: Manila, Philippines, 1989; pp. 41–73. [Google Scholar]
Appendix
Table A1.
Ecogeographic race of the different traditional rice varieties.
| Collection Number | Cultivar Name | Province | Eco-geographic Race |
|---|---|---|---|
| 10809 | Galo | Aurora | Indica |
| 10810 | Binaka (Malagkit) | Aurora | Indica |
| 10811 | Palawan | Aurora | Indica |
| 10813 | Galo (Malagkit) | Aurora | Indica |
| 10815 | Galo | Aurora | Indica |
| 10816 | Galo (Malagkit) | Aurora | Indica |
| 10817 | Brilyante | Aurora | Indica |
| 10818 | Galo | Aurora | Indica |
| 10819 | Malagkit | Aurora | Indica |
| 10821 | Inuway | Aurora | Indica |
| 10822 | Lubag/Galo | Aurora | Indica |
| 10823 | Palawan | Aurora | Javanica |
| 10824 | Lubag (With Awn) | Aurora | Indica |
| 10825 | Palawan | Aurora | Javanica |
| 10826 | Binisaya | Aurora | Indica |
| 10827 | Galo (Malagkit) | Aurora | Indica |
| 10828 | Gayanggang | Aurora | Indica |
| 10829 | Lubag | Aurora | Indica |
| 10830 | Malagkit (Puti) | Aurora | Javanica |
| 10831 | Inuhay | Aurora | Indica |
| 10832 | Palawan | Aurora | Javanica |
| 10833 | Inuhay | Aurora | Indica |
| 10834 | Galo | Aurora | Javanica |
| 10835 | Galo (Malagkit) | Aurora | Javanica |
| 10836 | Galo (Haba) | Aurora | Indica |
| 10837 | Kipil | Aurora | Japonica |
| 10838 | Binaka (Malagkit) | Aurora | Javanica |
| 10839 | Gayanggang | Aurora | Indica |
| 10840 | Palawan | Aurora | Javanica |
| 10841 | Sinagat | Aurora | Javanica |
| 10842 | Minindoro (Puti) | Aurora | Indica |
| 10844 | Minindoro (Pula) | Aurora | Indica |
| 10847 | Malagkit-Itim | Aurora | Javanica |
| 10848 | Palawan | Aurora | Javanica |
| 10850 | Lubag (Malagkit) | Aurora | Javanica |
| 10852 | Binernal | Aurora | Javanica |
| 10854 | Francis Rice | Aurora | Javanica |
| 10855 | Inuway | Aurora | Indica |
| 10856 | Galo | Aurora | Indica |
| 10857 | Kinakaw | Aurora | Indica |
| 10859 | Binisaya | Aurora | Javanica |
| 10860 | Dinorado | Aurora | Javanica |
| 10861 | Sinabado | Aurora | Indica |
| 10862 | Galo | Aurora | Indica |
| 10864 | Binisaya | Aurora | Indica |
| 10865 | Kinamalig | Aurora | Javanica |
| 10866 | Galo | Aurora | Indica |
| 10869 | Inuway | Aurora | Indica |
| 10870 | Galo | Aurora | Indica |
| 10874 | Azucena | Aurora | Indica |
| 10876 | Pirurutong (Malagkit) | Aurora | Javanica |
| 10877 | Kapirit | Aurora | Indica |
| 10880 | Milagrosa (Pilit) | Palawan | Indica |
| 10882 | Minantika | Palawan | Indica |
| 10884 | Pinutyukan | Palawan | Indica |
| 10885 | Milagrosa (Pula) | Palawan | Indica |
| 10886 | Doryat | Palawan | Indica |
| 10887 | Blandi | Palawan | Indica |
| 10888 | Doryat | Palawan | Indica |
| 10889 | Tipak | Palawan | Indica |
| 10890 | Rambo | Palawan | Indica |
| 10891 | Kinadoy | Palawan | Indica |
| 10892 | Kinadoy | Palawan | Indica |
| 10893 | Malagkit | Palawan | Indica |
| 10894 | Unknown | Palawan | Indica |
| 10898 | Tipak | Palawan | Javanica |
| 10899 | Dinorado | Palawan | Indica |
| 10900 | Malagkit (Pula) | Palawan | Javanica |
| 10901 | Dinorado | Palawan | Indica |
| 10902 | Dinorado | Palawan | Indica |
| 10903 | Buringan | Palawan | Indica |
| 10905 | Mating | Palawan | Indica |
| 10906 | Lawang | Palawan | Indica |
| 10908 | Muring-Oring | Palawan | Javanica |
| 10909 | Kinoron | Palawan | Indica |
| 10910 | Pinangpang | Palawan | Indica |
| 10911 | Red Rice | Palawan | Indica |
| 10912 | Binoring-Boring | Palawan | Javanica |
| 10914 | Sagang (Malagkit Puti) | Palawan | Javanica |
| 10915 | Fortuna | Palawan | Indica |
| 10916 | Calubid | Palawan | Indica |
| 10917 | Black Rice | Palawan | Javanica |
| 10918 | Milagrosa | Palawan | Indica |
| 10919 | Benareng | Palawan | Indica |
| 10921 | Minaola | Palawan | Javanica |
| 10923 | Benangkar | Palawan | Javanica |
| 10924 | Mangatas | Palawan | Indica |
| 10925 | Dinayudo | Palawan | Indica |
| 10926 | Dinamit | Palawan | Indica |
| 10927 | Tipak | Palawan | Indica |
| 10928 | Inamoy | Palawan | Indica |
| 10929 | Laok-Laok | Palawan | Indica |
| 10930 | Pinarongpong | Palawan | Javanica |
| 10931 | Kalinayin | Palawan | Indica |
| 10932 | Pinarompong | Palawan | Javanica |
| 10933 | Pinalawan | Palawan | Indica |
| 10934 | Doriat Pula | Palawan | Indica |
| 10939 | Pangpang | Palawan | Indica |
| 10940 | Inantote | Palawan | Indica |
| 10943 | Duriat | Palawan | Indica |
| 11015 | Metao | Negros Occidental | Javanica |
| 11016 | Kutsiam | Negros Occidental | Indica |
| 11017 | Aowot | Negros Occidental | Javanica |
| 11019 | Milagrosa | Negros Occidental | Indica |
| 11020 | Dinorado | Negros Occidental | Indica |
| 11021 | Vietnam Rice | Negros Occidental | Indica |
| 11022 | Pilit | Negros Occidental | Indica |
| 11023 | Azucena | Negros Occidental | Indica |
| 11025 | Unknown 2 | Negros Occidental | Indica |
| 11026 | Kasolid | Negros Occidental | Indica |
| 11027 | Pilit | Negros Occidental | Indica |
| 11032 | M-45 | Negros Occidental | Indica |
| 11035 | Malagaya | Negros Occidental | Javanica |
| 11036 | Dinorado | Negros Occidental | Indica |
| 11037 | Jao Dam Khi Kwai | Negros Occidental | Indica |
| 11038 | Ngacheik | Negros Occidental | Indica |
| 11040 | Tapol | Agusan Del Norte | Indica |
| 11041 | Kapakra | Agusan Del Norte | Indica |
| 11042 | Pilit-Tapol | Agusan Del Norte | Indica |
| 11043 | Señorita | Agusan Del Norte | Indica |
| 11044 | Red Cotabato | Misamis Oriental | Javanica |
| 11045 | Mimis | Misamis Oriental | Indica |
| 11046 | Mal-Os | Misamis Oriental | Javanica |
| 11047 | Kutong | Misamis Oriental | Indica |
| 11049 | Dinorado | Misamis Oriental | Javanica |
| 11050 | Kayatan | Misamis Oriental | Javanica |
| 11051 | Unknown | Misamis Oriental | Indica |
| 11052 | Kabuyog | Misamis Oriental | Javanica |
| 11053 | Gakit | Misamis Oriental | Javanica |
| 11055 | Dinorado | Misamis Oriental | Indica |
| 11056 | Speaker | Misamis Oriental | Javanica |
| 11057 | Dinorado | Misamis Oriental | Indica |
| 11058 | Gakit | Misamis Oriental | Javanica |
| 11059 | Unknown | Misamis Oriental | Indica |
| 11060 | Mabango | Misamis Oriental | Indica |
| 11061 | 18 (Pula) | Misamis Oriental | Indica |
| 11062 | Ir-9 | Misamis Oriental | Indica |
| 11063 | 18 (Puti) | Misamis Oriental | Indica |
| 11064 | Unknown | Misamis Oriental | Javanica |
| 11065 | Dinorado | Misamis Oriental | Indica |
| 11066 | Speaker | Misamis Oriental | Javanica |
| 11067 | Kabuyok | Misamis Oriental | Javanica |
| 11068 | Pilit | Misamis Oriental | Javanica |
| 11069 | Kabuyok | Misamis Oriental | Javanica |
| 11070 | Mixture from Kabuyok | Misamis Oriental | Javanica |
| 11071 | Dinorado | Misamis Oriental | Indica |
| 11072 | Speaker | Misamis Oriental | Javanica |
| 11074 | Kabuyok | Misamis Oriental | Javanica |
| 11075 | Gakit | Misamis Oriental | Javanica |
| 11076 | Dinorado | Misamis Oriental | Indica |
| 11077 | Speaker | Misamis Oriental | Javanica |
| 11078 | Kayatan | Misamis Oriental | Indica |
| 11079 | Pilit-Tapol | Misamis Oriental | Indica |
| 11080 | Speaker | Misamis Oriental | Javanica |
| 11081 | Dinorado | Misamis Oriental | Indica |
| 11082 | Unknown | Misamis Oriental | Javanica |
| 11083 | Burdagol | Negros Occidental | Indica |
| 11086 | M10-2 | Negros Occidental | Javanica |
| 11089 | M45 | Negros Occidental | Indica |
| 11091 | M119-Nl | Negros Occidental | Indica |
| 11092 | A-G-17 | Negros Occidental | Indica |
| 11094 | Mb-4 | Negros Occidental | Indica |
| 11096 | Inday | Negros Occidental | Indica |
| 11097 | Pangasinan | Negros Occidental | Indica |
| 11098 | M108 | Negros Occidental | Indica |
| 11100 | M11-6-1 | Negros Occidental | Indica |
| 11101 | Tape | Negros Occidental | Indica |
| 11102 | Milagrosa | Negros Occidental | Indica |
| 11105 | Fancy-1 | Negros Occidental | Indica |
| 11106 | Galicia F-1 | Negros Occidental | Indica |
| 11107 | Elon-Elon | Negros Occidental | Indica |
| 11108 | Manabang | Negros Occidental | Javanica |
| 11109 | Solig | Negros Occidental | Indica |
| 11110 | Kabagte | Negros Occidental | Javanica |
| 11111 | Azucena | Negros Occidental | Indica |
| 11112 | Ebod | Negros Occidental | Javanica |
| 11113 | Kotsiam | Negros Occidental | Indica |
| 11114 | Kalisan | Negros Occidental | Javanica |
| 11115 | Katipak | Negros Occidental | Indica |
| 11116 | Palawan | Negros Occidental | Javanica |
| 11117 | Bonlaw | Negros Occidental | Javanica |
| 11118 | Mangasa | Negros Occidental | Indica |
| 11119 | Pula | Agusan Del Norte | Indica |
| 11120 | Denorado | Agusan Del Norte | Indica |
| 11121 | Unknown | Agusan Del Norte | Indica |
| 11122 | M-23 | Agusan Del Norte | Indica |
| 11123 | Dinorado | Agusan Del Norte | Indica |
| 11124 | Dinorado | Agusan Del Norte | Indica |
| 11125 | Dinurado | Agusan Del Norte | Indica |
| 11127 | Tapul-Pilit | Agusan Del Norte | Indica |
| 11183 | Farasang | Kalinga | Indica |
| 11197 | Palawan | Nueva Ecija | Indica |
| 11198 | Binaka | Nueva Ecija | Indica |
| 11199 | Inuway | Nueva Ecija | Indica |
| 11200 | Galo | Nueva Ecija | Indica |
| 11201 | Palawan | Nueva Ecija | Javanica |
| 11202 | Ginobyerno | Nueva Ecija | Javanica |
| 11203 | Sinumay | Nueva Ecija | Indica |
| 11204 | Dinorado | Nueva Ecija | Indica |
| 11205 | Galo | Nueva Ecija | Indica |
| 11206 | Palawan | Nueva Ecija | Javanica |
| 11207 | Inuway | Nueva Ecija | Indica |
| 11208 | Galo | Nueva Ecija | Indica |
| 11209 | Palawan | Nueva Ecija | Javanica |
| 11210 | Palawan | Nueva Ecija | Javanica |
| 11211 | Dinorado | Nueva Ecija | Indica |
| 11212 | Inuway | Nueva Ecija | Indica |
| 11213 | Palawan | Nueva Ecija | Javanica |
| 11214 | Minindoro | Nueva Vizcaya | Javanica |
| 11215 | Palawan | Nueva Vizcaya | Javanica |
| 11216 | Brilyante | Nueva Vizcaya | Indica |
| 11217 | Galo | Nueva Vizcaya | Indica |
| 11231 | White Intan | Kalinga | Indica |
| 11232 | Intan (Red) | Kalinga | Indica |
| 11233 | Waray | Kalinga | Indica |
| 11234 | Red Intan | Kalinga | Indica |
| 11236 | Ummunoy | Kalinga | Javanica |
| 11237 | Kintuman | Kalinga | Javanica |
| 11238 | Ingtan (Red) | Kalinga | Indica |
| 11239 | Tuhuwan | Kalinga | Javanica |
| 11240 | Gwayay (Chay-Ot) | Kalinga | Javanica |
| 11241 | Puchagwan | Kalinga | Javanica |
| 11242 | Unoy (Ummunoy) | Kalinga | Javanica |
| 11243 | Unoy (Tu-Par) | Kalinga | Javanica |
| 11244 | Chay-Ot (Ojak) | Kalinga | Javanica |
| 11245 | Unoy (Umangan) | Kalinga | Javanica |
| 11246 | Chay-Ot (Ifuwan) | Kalinga | Javanica |
| 11247 | Chomalingan | Kalinga | Javanica |
| 11248 | Chinannay | Kalinga | Indica |
| 11249 | Inasotiyan | Kalinga | Javanica |
| 11250 | Finongod 2 | Kalinga | Indica |
| 11251 | Finongod | Kalinga | Javanica |
| 11252 | Innoway | Kalinga | Javanica |
| 11253 | Innoyan | Kalinga | Indica |
| 11254 | Mingol | Kalinga | Indica |
| 11255 | Ifo | Kalinga | Indica |
| 11256 | Bolinao | Kalinga | Javanica |
| 11282 | 7 Tonner | Bukidnon | Indica |
| 11283 | Red Rice | Bukidnon | Indica |
| 11284 | Red Tonner | Bukidnon | Indica |
| 11285 | Barako | Bukidnon | Indica |
| 11286 | Red Rice | Bukidnon | Indica |
| 11287 | Burdagol (Pilit) | Bukidnon | Indica |
| 11288 | Pilit | Bukidnon | Indica |
| 11289 | Pilit | Bukidnon | Indica |
| 11290 | Red Tonner | Bukidnon | Indica |
| 11291 | Red Rice | Bukidnon | Javanica |
| 11292 | Brown Rice | Bukidnon | Indica |
| 11293 | Dwarf Variety | Bukidnon | Indica |
| 11294 | M-3 | Bukidnon | Indica |
| 11295 | Elon-Elon | Bukidnon | Indica |
| 11296 | Red Tonner | Bukidnon | Indica |
| 11297 | Baknap | Bukidnon | Indica |
| 11298 | Azucena | Bukidnon | Indica |
| 11299 | Bakiki | Bukidnon | Indica |
| 11300 | Lubang | Bohol | Indica |
| 11301 | Lubang | Bohol | Indica |
| 11302 | Unknown | Bohol | Indica |
| 11303 | Muddy Rice | Bohol | Indica |
| 11304 | Kaimpas | Bohol | Indica |
| 11305 | Milagrosa | Bohol | Javanica |
| 11306 | Milagrosa | Bohol | Indica |
| 11307 | Inibi (Red) | Bohol | Indica |
| 11308 | Kaliga | Bohol | Indica |
| 11309 | Muddy Rice | Bohol | Indica |
| 11310 | Malagkit (Puti) | Bohol | Indica |
| 11311 | Kaliga (Red) | Bohol | Indica |
| 11312 | Rc 18 (Pula) | Bohol | Indica |
| 11313 | Kaliso | Bohol | Javanica |
| 11314 | Muddy Rice | Bohol | Indica |
| 11315 | Kaimpas | Bohol | Japonica |
| 11316 | Malagkit | Bohol | Indica |
| 11317 | C-4 Dinorado | Bohol | Indica |
| 11318 | Kabus-Ok | Bohol | Indica |
| 11319 | Rc 10 (Pula) | Bohol | Indica |
| 11320 | Ir 36 | Bohol | Indica |
| 11321 | Ping Rice | Bohol | Indica |
| 11322 | Pilit | Bohol | Indica |
| 11323 | Kabus-Ok | Bohol | Indica |
| 11324 | Kabus-Ok | Bohol | Indica |
| 11325 | 75 (Pula) | Bohol | Indica |
| 11326 | Unknown | Bohol | Indica |
| 11327 | Kabus-Ok | Bohol | Indica |
| 11328 | Unknown | Bohol | Indica |
| 11329 | Japan Red | Bohol | Indica |
| 11330 | Gl-2 | Bohol | Indica |
| 11332 | Rmp/Kaimpas | Bohol | Indica |
| 11333 | Bilar Red | Bohol | Indica |
| 11334 | Kananoy | Bohol | Indica |
| 11337 | Pilit Tapol | Bohol | Indica |
| 11338 | Miracle Pilit | Bohol | Javanica |
| 11339 | 7-7 Red | Bohol | Indica |
| 11340 | Red Rice | Bohol | Indica |
| 11341 | Lubang (M) | Bohol | Indica |
| 11342 | Hubahib | Bohol | Javanica |
| 11343 | Bares | Bohol | Japonica |
| 11345 | Mal-Us | Bohol | Indica |
| 11346 | Elon-Peta | Bohol | Indica |
| 11347 | Pungko | Bohol | Indica |
| 11348 | Melobina | Bohol | Indica |
| 11349 | Inabaka | Bohol | Indica |
| 11350 | Kayupo | Bohol | Indica |
| 11351 | Sulig | Bohol | Indica |
| 11352 | Ceres | Bohol | Javanica |
| 11354 | Torboho Red | Bohol | Javanica |
| 11355 | Pilit Taba | Bohol | Indica |
| 11356 | Lubang | Bohol | Javanica |
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).