


Phenotypic Variation and Relationships between Grain Yield, Protein Content and Unmanned Aerial Vehicle-Derived Normalized Difference Vegetation Index in Spring Wheat in Nordic–Baltic Environments

 , , ,
, , ,
Abstract
:1. Introduction
- Assess the variation in the GY, GPC, and UAV-derived NDVI across different years and nitrogen fertilization rates in the Nordic–Baltic region;
- Evaluate the performance and stability of spring wheat genotypes concerning the GY and GPC under varying N fertilization rates and diverse agrometeorological conditions;
- Explore the quantitative relationships between GY, GPC, and UAV-derived NDVI variables across multi-environmental trials during different plant growth stages.
2. Materials and Methods
2.1. Plant Materials
2.2. Field Trials
2.3. Phenotyping of Field Traits
2.4. UAV Data Acquisition and Processing
2.5. Data Statistical Analysis
3. Results
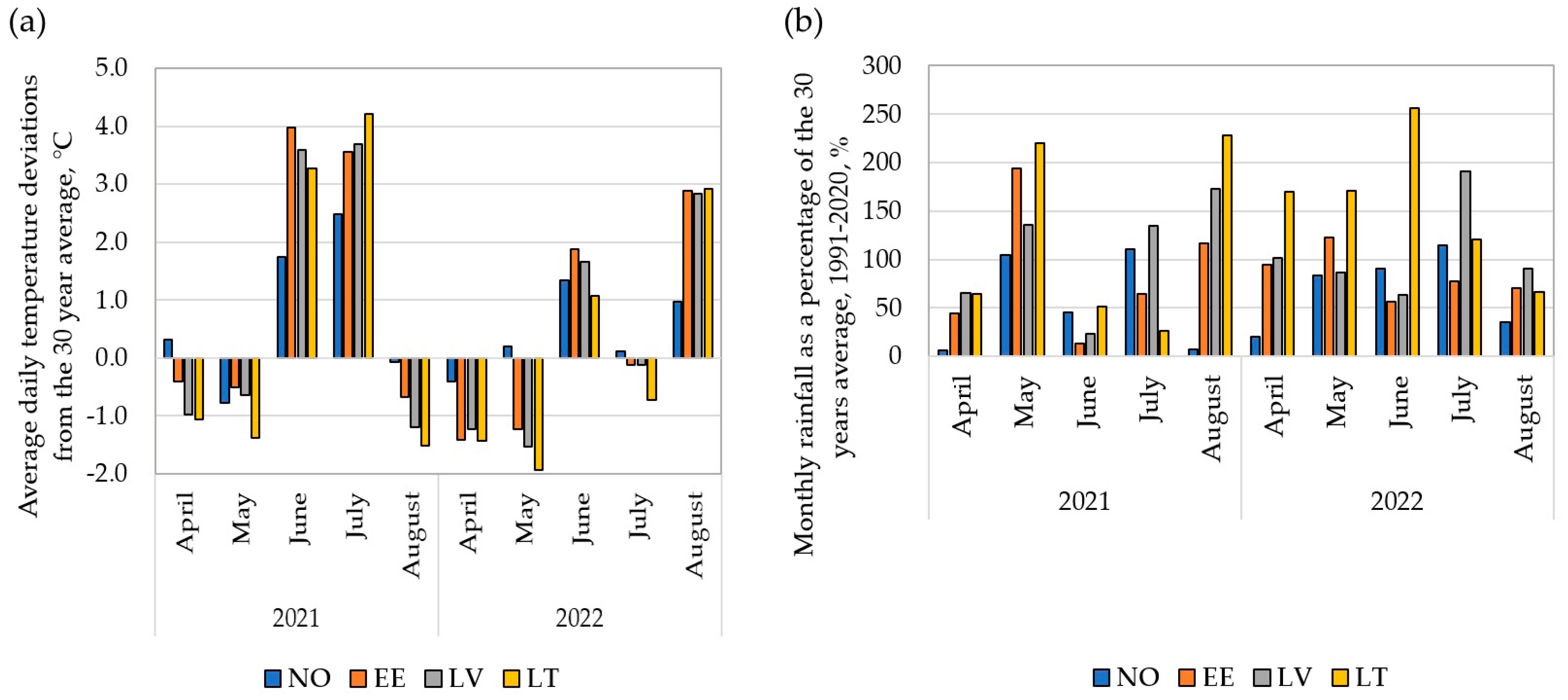
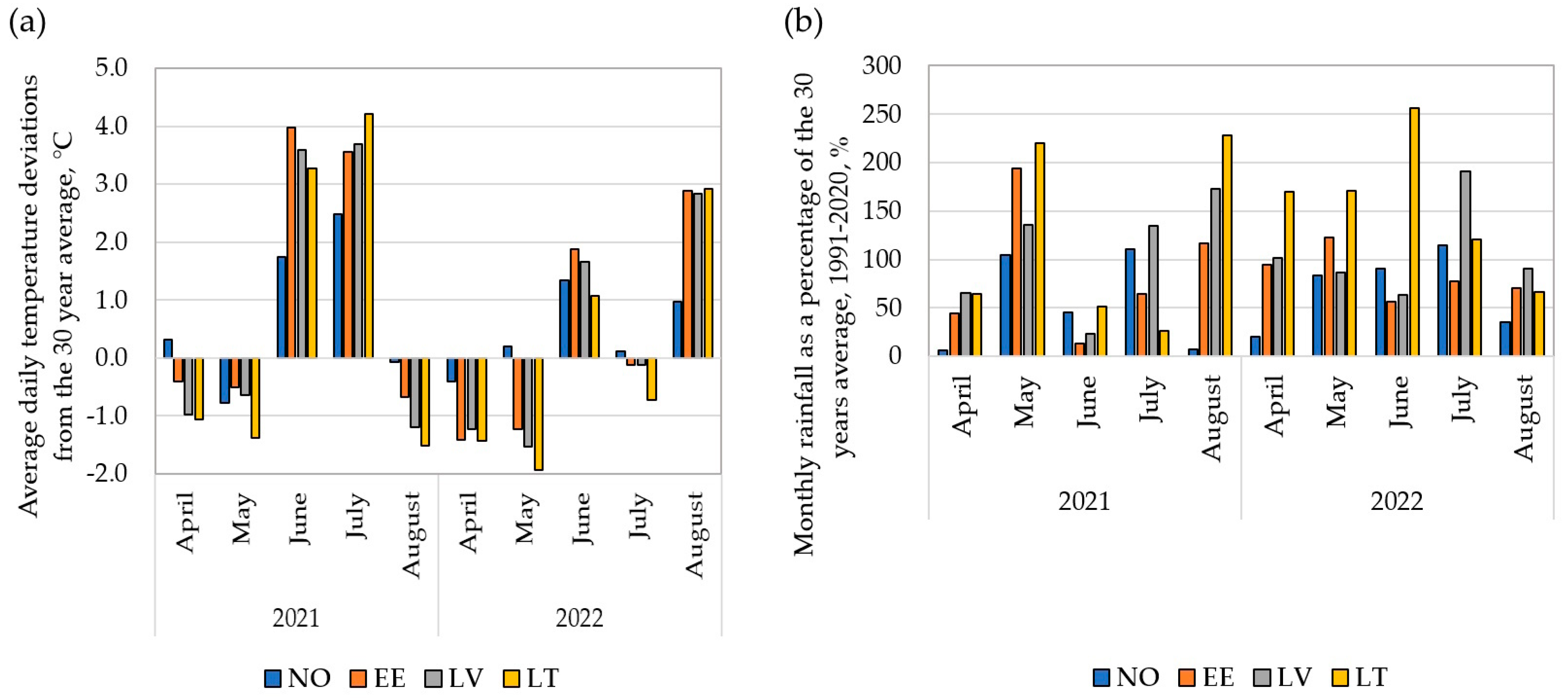
3.1. Variation in Meteorological Data
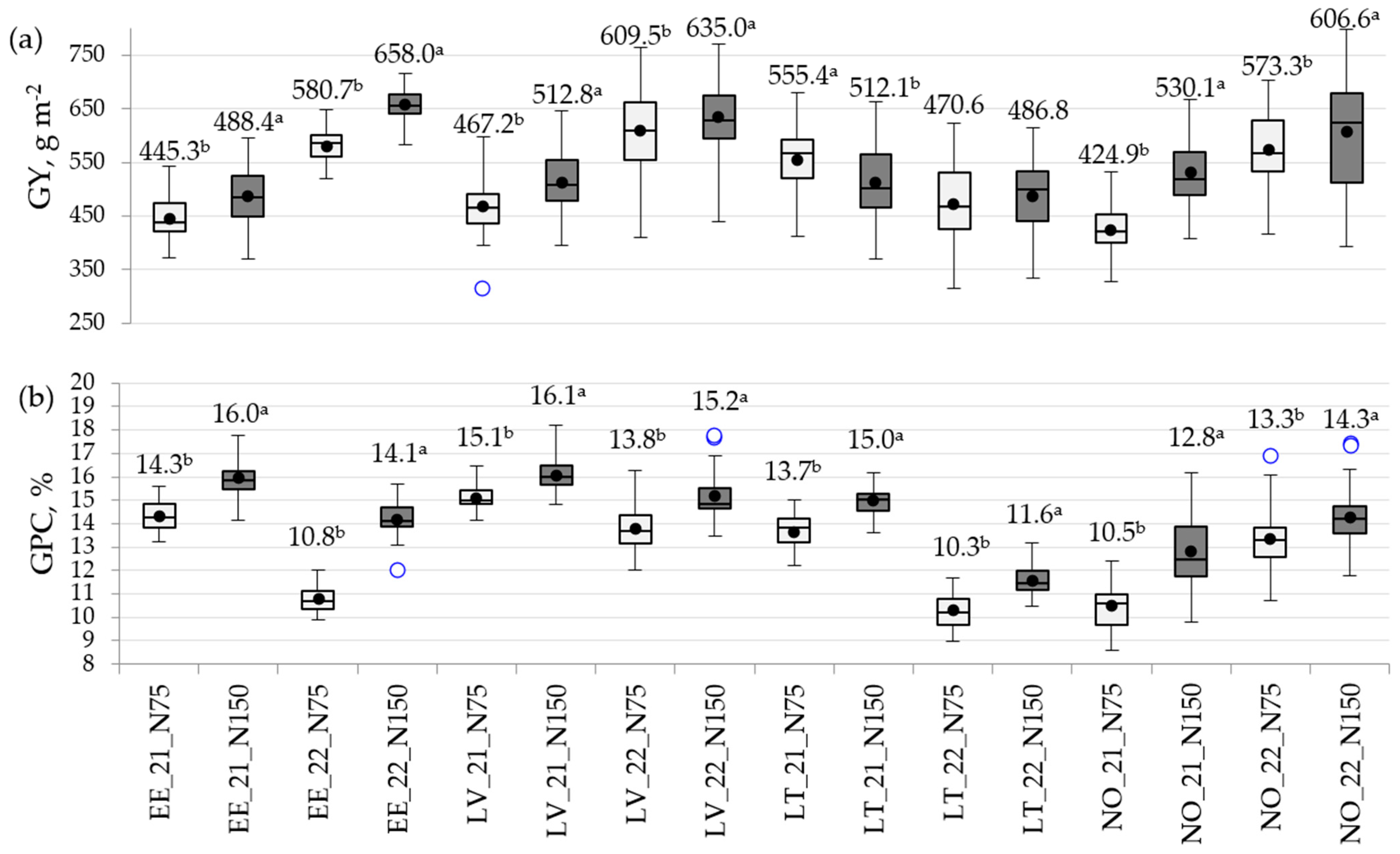
3.2. Variation in Traits by Location and Year
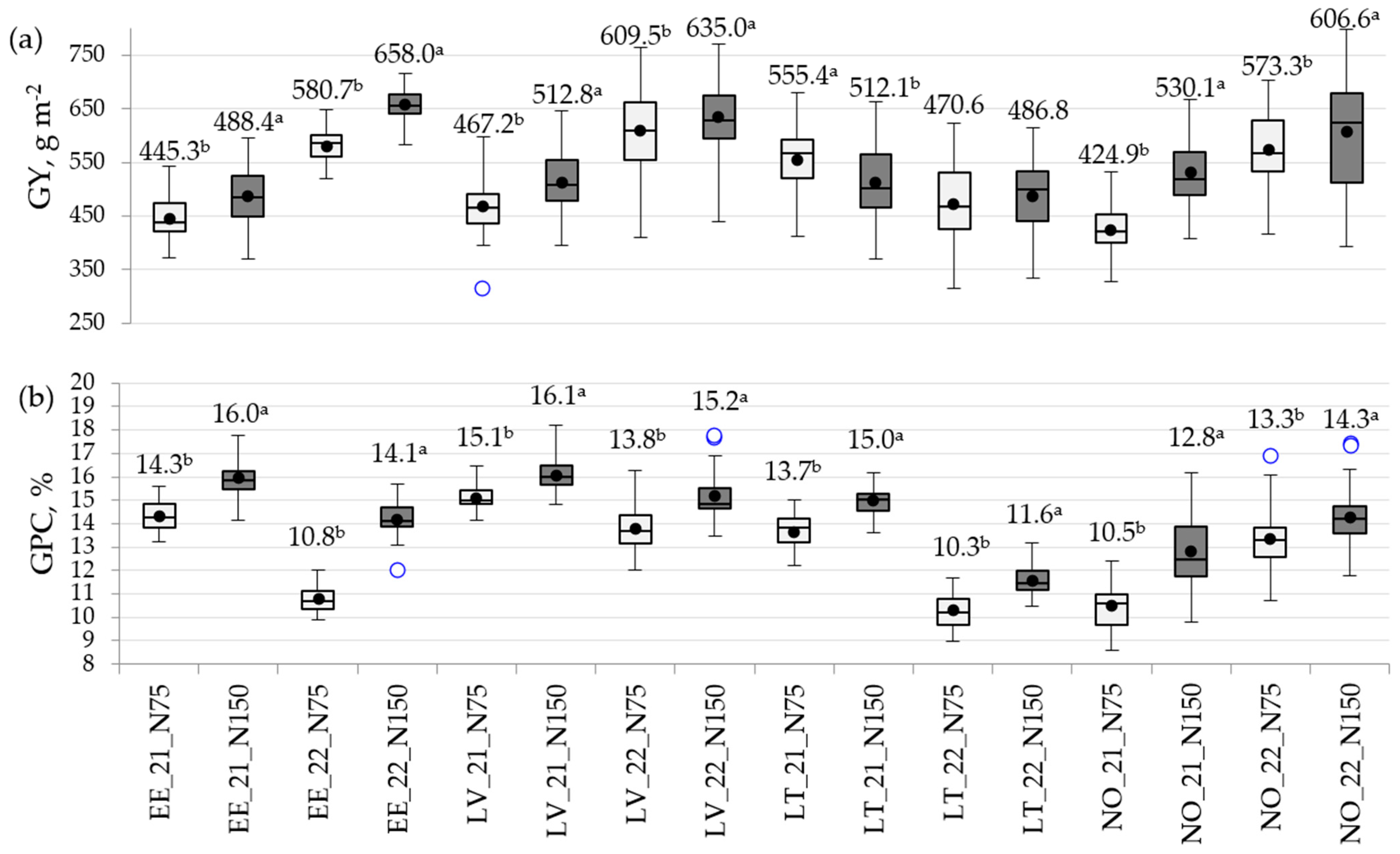
3.3. The Impact of N Rate on Phenotypic Variation in Traits
3.4. Mean Performance and Stability of Wheat Genotypes under Contrasting Environments
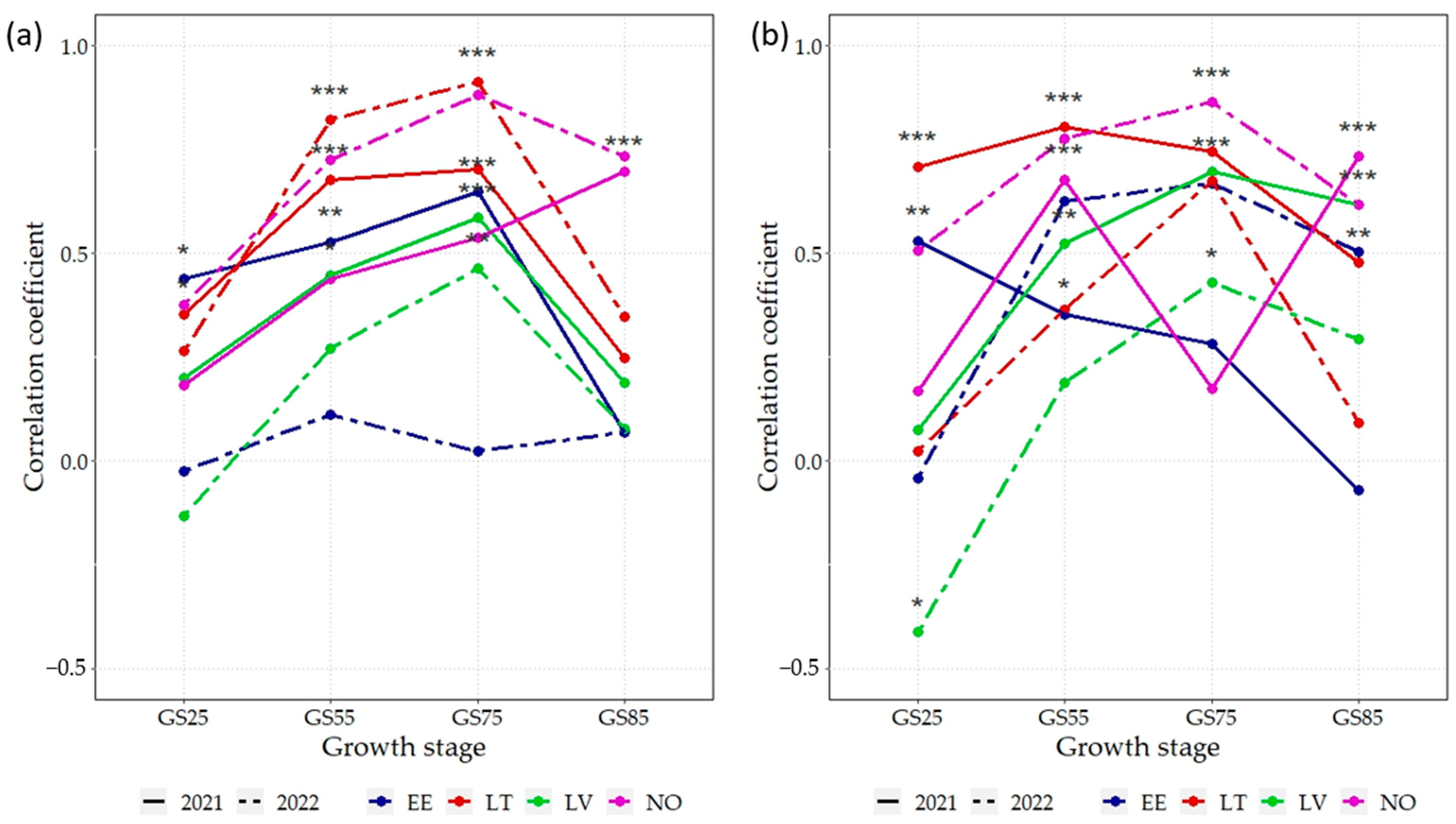
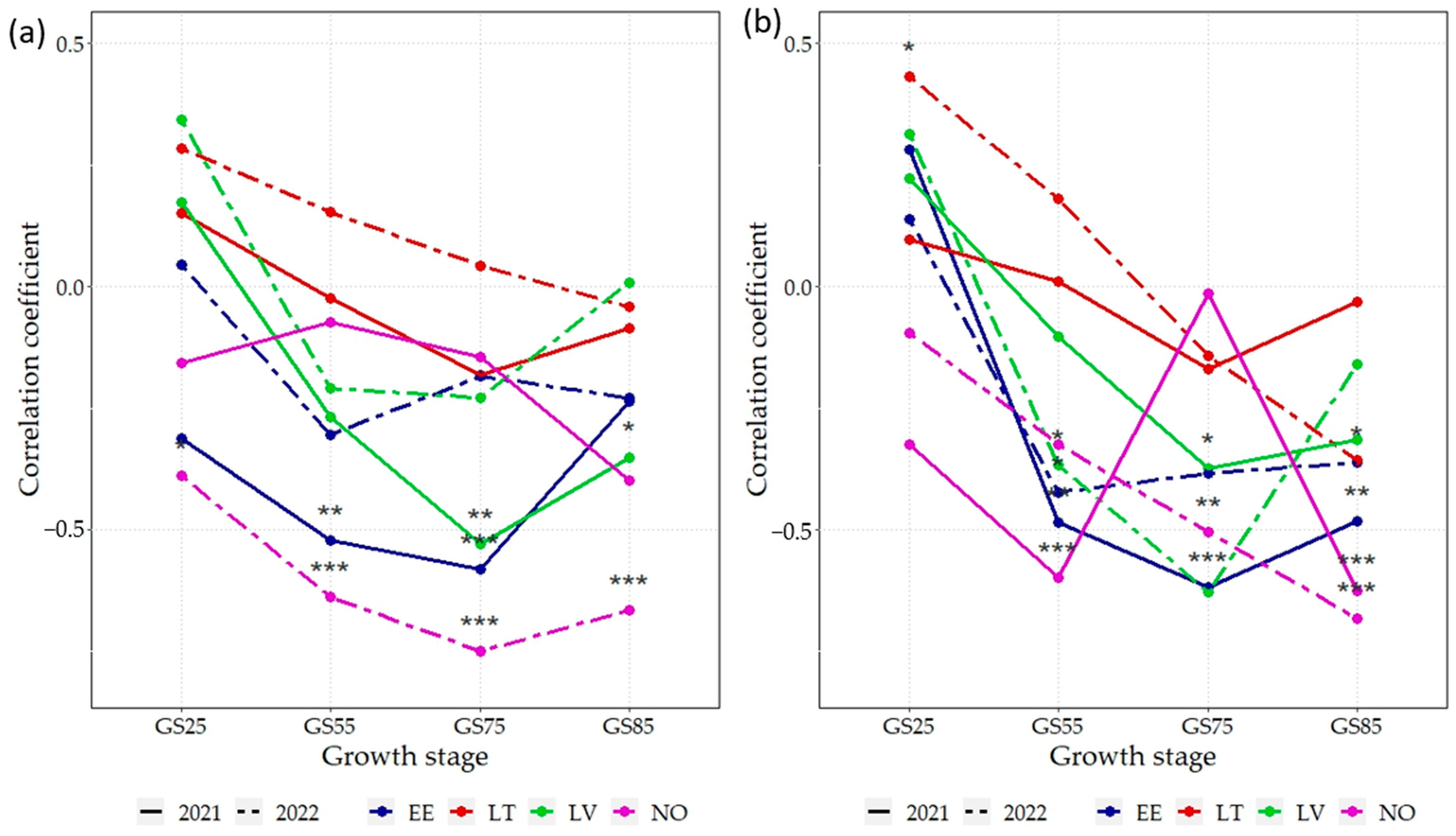
3.5. Correlations between NDVI and GY, GPC
3.6. Linear Relationships between NDVI and GY, GPC
4. Discussion
4.1. Variations in Meteorological Conditions across Trial Years and Locations Resulted in Differences in the GY and GPC
4.2. Spring Wheat Genotypes Grown under Contrasting N Rates Differed in Stability and Exhibited Different Patterns in NDVI Profiles during Seasons
4.3. Correlations and Linear Relationships between the GY, GPC, and UAV-Derived NDVI Highlight Key Crop Growth Stages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Crops and Livestock Production. 2023. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 12 June 2023).
- Wirehn, L. Nordic agriculture under climate change: A systematic review of challenges, opportunities and adaptation strategies for crop production. Land Use Policy 2018, 77, 63–74. [Google Scholar] [CrossRef]
- FAOSTAT. Crops and Livestock Production. 2021. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 30 November 2021).
- Lillemo, M.; Dieseth, J.A. Wheat breeding in Norway. In The World Wheat Book, A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., van Ginkel, M., Eds.; Lavoisier Publishing: Paris, France, 2011; Volume 2, pp. 45–75. [Google Scholar]
- Ruzgas, V.; Liatukas, Z. Wheat breeding in Lithuania. In The World Wheat Book, A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., van Ginkel, M., Eds.; Lavoisier Publishing: Paris, France, 2011; Volume 2, pp. 175–192. [Google Scholar]
- Strazdina, V.; Belicka, I.; Rashal, I.; Grauda, D. History of wheat breeding development in Latvia. In The World Wheat Book, A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., van Ginkel, M., Eds.; Lavoisier Publishing: Paris, France, 2011; Volume 2, pp. 213–238. [Google Scholar]
- Koppel, R.; Ingver, A.; Ardel, P.; Kangor, T.; Kennedy, H.J.; Koppel, M. The variability of yield and baking quality of wheat and suitability for export from Nordic-Baltic conditions. Acta Agric. Scand. Sect. B Soil Plant Sci. 2020, 70, 628–639. [Google Scholar] [CrossRef]
- Ross, A.S. Genetic and other factors affecting wheat quality. In Achieving Sustainable Cultivation of Wheat, Volume 1: Breeding, Quality Traits, Pests and Diseases; Langridge, P., Ed.; Burleigh Dodds Series in Agricultural Science; Burleigh Dodds Science Publishing Ltd.: Cambridge, UK, 2017; Volume 5, pp. 177–211. [Google Scholar]
- Cormier, F.; Faure, S.; Dubreuil, P.; Heumez, E.; Beauchêne, K.; Lafarge, S.; Praud, S.; Le Gouis, J. A multi-environmental study of recent breeding progress on nitrogen use efficiency in wheat (Triticum aestivum L.). Theor. Appl. Genet. 2013, 126, 3035–3048. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed]
- Kopecká, R.; Kameniarová, M.; Cerny, M.; Brzobohaty, B.; Novák, J. Abiotic Stress in Crop Production. Int. J. Mol. Sci. 2023, 24, 47. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J. Reducing the reliance on nitrogen fertilizer for wheat production. J. Cereal Sci. 2014, 59, 276–283. [Google Scholar] [CrossRef]
- Marsh, J.I.; Hu, H.F.; Gill, M.; Batley, J.; Edwards, D. Crop breeding for a changing climate: Integrating phenomics and genomics with bioinformatics. Theor. Appl. Genet. 2021, 134, 1677–1690. [Google Scholar] [CrossRef]
- Ma, J.J.; Zheng, B.Y.; He, Y. Applications of a Hyperspectral Imaging System Used to Estimate Wheat Grain Protein: A Review. Front. Plant Sci. 2022, 13, 10. [Google Scholar] [CrossRef]
- Radoglou-Grammatikis, P.; Sarigiannidis, P.; Lagkas, T.; Moscholios, I. A compilation of UAV applications for precision agriculture. Comput. Netw. 2020, 172, 18. [Google Scholar] [CrossRef]
- Xie, C.; Yang, C. A review on plant high-throughput phenotyping traits using UAV-based sensors. Comput. Electron. Agric. 2020, 178, 105731. [Google Scholar] [CrossRef]
- Fei, S.P.; Hassan, M.A.; He, Z.H.; Chen, Z.; Shu, M.Y.; Wang, J.K.; Li, C.C.; Xiao, Y.G. Assessment of Ensemble Learning to Predict Wheat Grain Yield Based on UAV-Multispectral Reflectance. Remote Sens. 2021, 13, 25. [Google Scholar] [CrossRef]
- Prey, L.; Hanemann, A.; Ramgraber, L.; Seidl-Schulz, J.; Noack, P.O. UAV-Based Estimation of Grain Yield for Plant Breeding: Applied Strategies for Optimizing the Use of Sensors, Vegetation Indices, Growth Stages, and Machine Learning Algorithms. Remote Sens. 2022, 14, 17. [Google Scholar] [CrossRef]
- Huang, S.; Tang, L.N.; Hupy, J.P.; Wang, Y.; Shao, G.F. A commentary review on the use of normalized difference vegetation index (NDVI) in the era of popular remote sensing. J. For. Res. 2021, 32, 1–6. [Google Scholar] [CrossRef]
- Bellón, B.; Bégué, A.; Lo Seen, D.; de Almeida, C.A.; Simoes, M. A Remote Sensing Approach for Regional-Scale Mapping of Agricultural Land-Use Systems Based on NDVI Time Series. Remote Sens. 2017, 9, 600. [Google Scholar] [CrossRef]
- Zhu, X.L.; Liu, D.S. Improving forest aboveground biomass estimation using seasonal Landsat NDVI time-series. ISPRS J. Photogramm. Remote Sens. 2015, 102, 222–231. [Google Scholar] [CrossRef]
- Beisel, N.S.; Callaham, J.B.; Sng, N.J.; Taylor, D.J.; Paul, A.-L.; Ferl, R.J. Utilization of single-image normalized difference vegetation index (SI-NDVI) for early plant stress detection. Appl. Plant Sci. 2018, 6, e01186. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Guzman, J.; Atkinson, P.M.; Dash, J.; Rioja-Nieto, R. Spatiotemporal Variation in Mangrove Chlorophyll Concentration Using Landsat 8. Remote Sens. 2015, 7, 14530–14558. [Google Scholar] [CrossRef]
- Cabrera-Bosquet, L.; Molero, G.; Stellacci, A.M.; Bort, J.; Nogués, S.; Araus, J.L. NDVI as a Potential Tool for Predicting Biomass, Plant Nitrogen Content and Growth in Wheat Genotypes Subjected to Different Water and Nitrogen Conditions. Cereal Res. Commun. 2011, 39, 147–159. [Google Scholar] [CrossRef]
- Hitz, K.; Clark, A.J.; Van Sanford, D.A. Identifying nitrogen-use efficient soft red winter wheat lines in high and low nitrogen environments. Field Crops Res. 2017, 200, 1–9. [Google Scholar] [CrossRef]
- Naser, M.A.; Khosla, R.; Longchamps, L.; Dahal, S. Using NDVI to Differentiate Wheat Genotypes Productivity Under Dryland and Irrigated Conditions. Remote Sens. 2020, 12, 17. [Google Scholar] [CrossRef]
- Guan, S.L.; Fukami, K.; Matsunaka, H.; Okami, M.; Tanaka, R.; Nakano, H.; Sakai, T.; Nakano, K.; Ohdan, H.; Takahashi, K. Assessing Correlation of High-Resolution NDVI with Fertilizer Application Level and Yield of Rice and Wheat Crops using Small UAVs. Remote Sens. 2019, 11, 19. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Zobel, R.; Wright, M.; Gauch, H. Statistical Analysis of a Yield Trial. Agron. J. 1988, 80, 388–393. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.C.; da Silva, J.A.G.; Marchioro, V.S.; de Souza, V.Q.; Jost, E. Mean Performance and Stability in Multi- Environment Trials I: Combining Features of AMMI and BLUP Techniques. Agron. J. 2019, 111, 2949–2960. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 15 July 2023).
- Wan, C.; Dang, P.; Gao, L.; Wang, J.; Tao, J.; Qin, X.; Feng, B.; Gao, J. How Does the Environment Affect Wheat Yield and Protein Content Response to Drought? A Meta-Analysis. Front. Plant Sci. 2022, 13, 896985. [Google Scholar] [CrossRef]
- Asseng, S.; Martre, P.; Maiorano, A.; Roetter, R.P.; O’Leary, G.J.; Fitzgerald, G.J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M.A.; et al. Climate change impact and adaptation for wheat protein. Glob. Chang. Biol. 2019, 25, 155–173. [Google Scholar] [CrossRef]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S.T.; Cui, Y.; Liu, Y.; Zahoor, R.; Jiang, D.; Dai, T. Nitrogen Nutrition Improves the Potential of Wheat (Triticum aestivum L.) to Alleviate the Effects of Drought Stress during Vegetative Growth Periods. Front. Plant Sci. 2016, 7, 981. [Google Scholar] [CrossRef]
- Giménez, V.D.; Miralles, D.J.; García, G.A.; Serrago, R.A. Can crop management reduce the negative effects of warm nights on wheat yield? Field Crops Res. 2021, 261, 108010. [Google Scholar] [CrossRef]
- Ru, C.; Hu, X.T.; Chen, D.Y.; Wang, W.E.; Zhen, J.B.; Song, T.Y. Individual and combined effects of heat and drought and subsequent recovery on winter wheat (Triticum aestivum L.) photosynthesis, nitrogen metabolism, cell osmoregulation, and yield formation. Plant Physiol. Biochem. 2023, 196, 222–235. [Google Scholar] [CrossRef] [PubMed]
- Forkel, M.; Carvalhais, N.; Verbesselt, J.; Mahecha, M.D.; Neigh, C.S.R.; Reichstein, M. Trend Change Detection in NDVI Time Series: Effects of Inter-Annual Variability and Methodology. Remote Sens. 2013, 5, 2113–2144. [Google Scholar] [CrossRef]
- Thapa, S.; Rudd, J.C.; Xue, Q.W.; Bhandari, M.; Reddy, S.K.; Jessup, K.E.; Liu, S.Y.; Devkota, R.N.; Baker, J.; Baker, S. Use of NDVI for characterizing winter wheat response to water stress in a semi-arid environment. J. Crop Improv. 2019, 33, 633–648. [Google Scholar] [CrossRef]
- Yousfi, S.; Gracia-Romero, A.; Kellas, N.; Kaddour, M.; Chadouli, A.; Karrou, M.; Araus, J.L.; Serret, M.D. Combined Use of Low-Cost Remote Sensing Techniques and δ13C to Assess Bread Wheat Grain Yield under Different Water and Nitrogen Conditions. Agronomy 2019, 9, 285. [Google Scholar] [CrossRef]
- Laperche, A.; Brancourt-Hulmel, M.; Heumez, E.; Gardet, O.; Hanocq, E.; Devienne-Barret, F.; Le Gouis, J. Using genotype x nitrogen interaction variables to evaluate the QTL involved in wheat tolerance to nitrogen constraints. Theor. Appl. Genet. 2007, 115, 399–415. [Google Scholar] [CrossRef]
- Mroz, T.; Dieseth, J.A.; Lillemo, M. Historical grain yield genetic gains in Norwegian spring wheat under contrasting fertilization regimes. Crop Sci. 2022, 62, 997–1010. [Google Scholar] [CrossRef]
- Zörb, C.; Ludewig, U.; Hawkesford, M.J. Perspective on Wheat Yield and Quality with Reduced Nitrogen Supply. Trends Plant Sci. 2018, 23, 1029–1037. [Google Scholar] [CrossRef]
- Weiner, J.; Du, Y.L.; Zhao, Y.M.; Li, F.M. Allometry and Yield Stability of Cereals. Front. Plant Sci. 2021, 12, 5. [Google Scholar] [CrossRef]
- Ali, M. Stability Analysis of Bread Wheat Genotypes under Different Nitrogen Fertilizer Levels. J. Plant Prod. 2017, 8, 261–275. [Google Scholar] [CrossRef]
- Hassan, M.A.; Yang, M.J.; Fu, L.P.; Rasheed, A.; Zheng, B.Y.; Xia, X.C.; Xiao, Y.G.; He, Z.H. Accuracy assessment of plant height using an unmanned aerial vehicle for quantitative genomic analysis in bread wheat. Plant Methods 2019, 15, 12. [Google Scholar] [CrossRef]
- Duan, T.; Chapman, S.C.; Guo, Y.; Zheng, B. Dynamic monitoring of NDVI in wheat agronomy and breeding trials using an unmanned aerial vehicle. Field Crops Res. 2017, 210, 71–80. [Google Scholar] [CrossRef]
- Sultana, S.R.; Ali, A.; Ahmad, A.; Mubeen, M.; Zia-Ul-Haq, M.; Ahmad, S.; Ercisli, S.; Jaafar, H.Z.E. Normalized Difference Vegetation Index as a Tool for Wheat Yield Estimation: A Case Study from Faisalabad, Pakistan. Sci. World J. 2014, 2014, 725326–725328. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, K.S.; Mihalyov, P.D.; Lewien, M.J.; Pumphrey, M.O.; Carter, A.H. Combining Genomic and Phenomic Information for Predicting Grain Protein Content and Grain Yield in Spring Wheat. Front. Plant Sci. 2021, 12, 14. [Google Scholar] [CrossRef]
- Walsh, O.S.; Marshall, J.M.; Nambi, E.; Jackson, C.A.; Ansah, E.O.; Lamichhane, R.; McClintick-Chess, J.; Bautista, F. Wheat Yield and Protein Estimation with Handheld and Unmanned Aerial Vehicle-Mounted Sensors. Agronomy 2023, 13, 14. [Google Scholar] [CrossRef]
- Yousfi, S.; Kellas, N.; Saidi, L.; Benlakehal, Z.; Chaou, L.; Siad, D.; Herda, F.; Karrou, M.; Vergara, O.; Gracia, A.; et al. Comparative performance of remote sensing methods in assessing wheat performance under Mediterranean conditions. Agric. Water Manag. 2016, 164, 137–147. [Google Scholar] [CrossRef]
- Feng, M.-C.; Xiao, L.-J.; Zhang, M.-J.; Yang, W.-D.; Ding, G.-W. Integrating Remote Sensing and GIS for Prediction of Winter Wheat (Triticum aestivum) Protein Contents in Linfen (Shanxi), China. PLoS ONE 2014, 9, e80989. [Google Scholar] [CrossRef]
- Stoy, P.C.; Khan, A.M.; Wipf, A.; Silverman, N.; Powell, S.L. The spatial variability of NDVI within a wheat field: Information content and implications for yield and grain protein monitoring. PLoS ONE 2022, 17, e0265243. [Google Scholar] [CrossRef]
- Barmeier, G.; Hofer, K.; Schmidhalter, U. Mid-season prediction of grain yield and protein content of spring barley cultivars using high-throughput spectral sensing. Eur. J. Agron. 2017, 90, 108–116. [Google Scholar] [CrossRef]
- Fu, Z.P.; Yu, S.S.; Zhang, J.Y.; Xi, H.; Gao, Y.; Lu, R.H.; Zheng, H.B.; Zhu, Y.; Cao, W.X.; Liu, X.J. Combining UAV multispectral imagery and ecological factors to estimate leaf nitrogen and grain protein content of wheat. Eur. J. Agron. 2022, 132, 15. [Google Scholar] [CrossRef]
- Xue, L.H.; Cao, W.X.; Yang, L.Z. Predicting grain yield and protein content in winter wheat at different N supply levels using canopy reflectance spectra. Pedosphere 2007, 17, 646–653. [Google Scholar] [CrossRef]
- Wang, K.; Huggins, D.R.; Tao, H.Y. Rapid mapping of winter wheat yield, protein, and nitrogen uptake using remote and proximal sensing. Int. J. Appl. Earth Obs. Geoinf. 2019, 82, 10. [Google Scholar] [CrossRef]
- Sánchez, N.; Plaza, J.; Criado, M.; Pérez-Sánchez, R.; Gómez-Sánchez, M.A.; Morales-Corts, M.R.; Palacios, C. The Second Derivative of the NDVI Time Series as an Estimator of Fresh Biomass: A Case Study of Eight Forage Associations Monitored via UAS. Drones 2023, 7, 19. [Google Scholar] [CrossRef]
- Marti, J.; Bort, J.; Slafer, G.A.; Araus, J.L. Can wheat, yield be assessed by early measurements of Normalized Difference Vegetation Index? Ann. Appl. Biol. 2007, 150, 253–257. [Google Scholar] [CrossRef]
- Chandel, N.S.; Tiwari, P.S.; Singh, K.P.; Jat, D.; Gaikwad, B.B.; Tripathi, H.; Golhani, K. Yield prediction in wheat (Triticum aestivum L.) using spectral reflectance indices. Curr. Sci. 2019, 116, 272–278. [Google Scholar] [CrossRef]
- Noureldin, N.A.; Saudy, H.S.; Ashmawy, F.; Saed, H.M. Grain yield response index of bread wheat cultivars as influenced by nitrogen levels. Ann. Agric. Sci. 2013, 58, 147–152. [Google Scholar] [CrossRef]
- Walsh, O.S.; Torrion, J.A.; Liang, X.; Shafian, S.; Yang, R.; Belmont, K.M.; McClintick-Chess, J.R. Grain yield, quality, and spectral characteristics of wheat grown under varied nitrogen and irrigation. Agrosyst. Geosci. Environ. 2020, 3, e20104. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage of Plant Development | LV | EE | LT | NO | |||||
|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | 2021 | 2022 | ||
| Middle of vegetative plant development | GS25 | 05.29 | 05.25 | 05.31 | 06.06 | 05.28 | 05.23 | 05.28 | 05.31 |
| Spike emergence or heading | GS55 | 06.21 | 06.30 | 06.28 | 06.30 | 06.24 | 06.27 | 06.23 | 06.29 |
| Middle of grain filling | GS75 | 07.13 | 07.22 | 07.12 | 07.21 | 07.09 | 07.19 | 07.13 | 07.21 |
| Late grain filling | GS85 | 07.27 | 08.12 | 07.26 | 08.09 | 07.23 | 08.08 | 07.26 | 08.09 |
| Year | Location | |||
|---|---|---|---|---|
| NO | EE | LV | LT | |
| GY, g m−2 | ||||
| 2021 | 477.5 ± 79.1 b | 466.8 ± 51.9 b | 490.0 ± 57.2 b | 533.7 ± 70.7 a |
| 2022 | 589.9 ± 94.6 a | 619.3 ± 50.5 a | 622.2 ± 75.7 a | 478.7 ± 73.3 b |
| GPC, % | ||||
| 2021 | 11.7 ± 1.7 b | 15.1 ± 1.1 a | 15.6 ± 0.8 a | 14.3 ± 0.9 a |
| 2022 | 13.8 ± 1.4 a | 12.5 ± 1.8 b | 14.5 ± 1.2 b | 10.9 ± 0.9 b |
| Year | Location | |||
|---|---|---|---|---|
| NO | EE | LV | LT | |
| GS25 | ||||
| 2021 | 0.536 ± 0.05 a | 0.637 ± 0.05 a | 0.479 ± 0.03 a | 0.544 ± 0.08 b |
| 2022 | 0.466 ± 0.10 b | 0.515 ± 0.09 b | 0.408 ± 0.04 b | 0.574 ± 0.06 a |
| GS55 | ||||
| 2021 | 0.889 ± 0.03 | 0.903 ± 0.01 a | 0.864 ± 0.02 b | 0.870 ± 0.03 b |
| 2022 | 0.882 ± 0.03 | 0.894 ± 0.02 b | 0.905 ± 0.01 a | 0.885 ± 0.02 a |
| GS75 | ||||
| 2021 | 0.821 ± 0.04 b | 0.791 ± 0.03 | 0.763 ± 0.05 b | 0.755 ± 0.06 a |
| 2022 | 0.837 ± 0.04 a | 0.782 ± 0.07 | 0.803 ± 0.04 a | 0.729 ± 0.06 b |
| GS85 | ||||
| 2021 | 0.478 ± 0.09 b | 0.445 ± 0.06 | 0.349 ± 0.04 b | 0.350 ± 0.04 a |
| 2022 | 0.577 ± 0.10 a | 0.425 ± 0.08 | 0.366 ± 0.05 a | 0.269 ± 0.03 b |
| Environment | GS25 | GS55 | GS75 | GS85 | ||||
|---|---|---|---|---|---|---|---|---|
| N75 | N150 | N75 | N150 | N75 | N150 | N75 | N150 | |
| EE_2021 | 0.613 b | 0.660 a | 0.898 b | 0.908 a | 0.784 | 0.797 | 0.430 b | 0.461 a |
| EE_2022 | 0.506 | 0.521 | 0.890 | 0.897 | 0.780 | 0.783 | 0.414 | 0.436 |
| LV_2021 | 0.477 | 0.475 | 0.857 b | 0.871 a | 0.748 b | 0.778 a | 0.335 b | 0.363 a |
| LV_2022 | 0.403 | 0.412 | 0.901 b | 0.910 a | 0.799 | 0.807 | 0.351 b | 0.382 a |
| LT_2021 | 0.546 | 0.542 | 0.868 | 0.872 | 0.753 | 0.757 | 0.323 b | 0.346 a |
| LT_2022 | 0.602 a | 0.546 b | 0.878 b | 0.891 a | 0.702 b | 0.755 a | 0.260 b | 0.277 a |
| NO_2021 | 0.520 b | 0.552 a | 0.866 b | 0.912 a | 0.791 b | 0.850 a | 0.421 b | 0.535 a |
| NO_2022 | 0.467 | 0.466 | 0.885 | 0.880 | 0.837 | 0.836 | 0.547 b | 0.606 a |
| Genotype | GY | WAASB_GY | GPC | WAASB_GPC | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N75 | N150 | N75 | N150 | N75 | N150 | N75 | N150 | |||||||||
| g m−2 | R | g m−2 | R | Value | R | Value | R | % | R | % | R | % | R | % | R | |
| DS-655-7-DH | 564.1 | 1 | 592.5 | 3 | 2.7 | 11 | 3.54 | 12 | 12.7 | 7 | 14.3 | 7 | 0.39 | 12 | 0.33 | 10 |
| DS-17-16-DH | 558.1 | 2 | 609.5 | 1 | 1.13 | 1 | 2.63 | 9 | 12.0 | 15 | 13.6 | 15 | 0.20 | 2 | 0.35 | 12 |
| DS-720-3-DH | 547.4 | 3 | 607.7 | 2 | 4.23 | 16 | 2.83 | 10 | 13.0 | 5 | 14.5 | 4 | 0.22 | 6 | 0.20 | 5 |
| DS-638-5-DH | 539.9 | 4 | 575.6 | 6 | 3.28 | 12 | 4.18 | 16 | 11.7 | 16 | 13.2 | 16 | 0.39 | 13 | 0.59 | 15 |
| Betong | 538.6 | 5 | 570.9 | 7 | 1.73 | 4 | 2.02 | 6 | 12.7 | 9 | 14.2 | 11 | 0.22 | 5 | 0.28 | 7 |
| 013-01 | 538.4 | 6 | 581.7 | 4 | 4.22 | 15 | 3.79 | 15 | 12.3 | 14 | 13.7 | 14 | 0.42 | 15 | 0.61 | 16 |
| 013-032 | 528.7 | 7 | 580.6 | 5 | 2.34 | 10 | 1.43 | 4 | 12.5 | 12 | 14.2 | 12 | 0.42 | 14 | 0.34 | 11 |
| Robijs | 523.6 | 8 | 554.8 | 9 | 3.7 | 14 | 3.68 | 14 | 12.4 | 13 | 14.1 | 13 | 0.30 | 8 | 0.33 | 9 |
| 013-074 | 521.4 | 9 | 564.4 | 8 | 1.98 | 7 | 2.30 | 7 | 12.5 | 11 | 14.5 | 6 | 0.17 | 1 | 0.17 | 3 |
| 990-2 | 507.4 | 10 | 528.6 | 13 | 3.48 | 13 | 1.04 | 1 | 12.7 | 8 | 14.3 | 8 | 0.31 | 10 | 0.13 | 1 |
| Hiie | 505.9 | 11 | 511.5 | 15 | 1.20 | 2 | 3.20 | 11 | 12.6 | 10 | 14.2 | 10 | 0.21 | 3 | 0.14 | 2 |
| Zombi | 496.6 | 12 | 533.1 | 11 | 1.49 | 3 | 1.14 | 3 | 13.1 | 3 | 14.5 | 5 | 0.30 | 9 | 0.28 | 8 |
| Caress | 493.9 | 13 | 548.4 | 10 | 1.85 | 6 | 1.86 | 5 | 12.7 | 6 | 14.3 | 9 | 0.21 | 4 | 0.22 | 6 |
| Voore | 489.3 | 14 | 532.1 | 12 | 2.13 | 9 | 1.06 | 2 | 13.1 | 4 | 14.7 | 3 | 0.24 | 7 | 0.19 | 4 |
| 876 | 477.9 | 15 | 516.4 | 14 | 1.77 | 5 | 2.43 | 8 | 14.0 | 1 | 16.0 | 1 | 0.52 | 16 | 0.48 | 13 |
| Runar | 422.3 | 16 | 454.5 | 16 | 2.09 | 8 | 3.59 | 13 | 13.5 | 2 | 15.6 | 2 | 0.32 | 11 | 0.52 | 14 |
| Mean | 515.8 a | × | 553.9 b | × | 2.46 | × | 2.54 | × | 12.7 b | × | 14.4 a | × | 0.3 | × | 0.32 | × |
| LSD | 29.03 | × | 34.2 | × | × | × | × | × | 0.26 | × | 0.38 | × | × | × | × | × |
| Genotype * | PH, cm | R | DM, Days | R | NDVI | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GS25 | R | GS55 | R | GS75 | R | GS85 | R | |||||
| DS-17-16-DH | 86.3 | 12 | 106.9 | 5 | 0.512 | 11 | 0.886 | 8 | 0.809 | 5 | 0.427 | 5 |
| DS-655-7-DH | 91.4 | 5 | 107.8 | 4 | 0.533 | 3 | 0.896 | 4 | 0.823 | 3 | 0.433 | 4 |
| DS-720-3-DH | 90.1 | 7 | 108.6 | 2 | 0.526 | 6 | 0.897 | 3 | 0.822 | 4 | 0.449 | 3 |
| 013-01 | 88.5 | 8 | 108.4 | 3 | 0.493 | 15 | 0.904 | 2 | 0.837 | 1 | 0.491 | 2 |
| DS-638-5-DH | 85.5 | 14 | 111.0 | 1 | 0.504 | 13 | 0.910 | 1 | 0.833 | 2 | 0.511 | 1 |
| Betong | 86.1 | 13 | 104.9 | 10 | 0.518 | 8 | 0.877 | 12 | 0.770 | 11 | 0.379 | 10 |
| 013-032 | 86.4 | 11 | 106.6 | 6 | 0.505 | 12 | 0.891 | 6 | 0.805 | 6 | 0.405 | 8 |
| 013-074 | 88.2 | 9 | 104.8 | 11 | 0.523 | 7 | 0.882 | 10 | 0.767 | 12 | 0.371 | 12 |
| Robijs | 94.0 | 3 | 106.1 | 8 | 0.570 | 1 | 0.894 | 5 | 0.795 | 7 | 0.410 | 6 |
| Caress | 80.4 | 15 | 105.4 | 9 | 0.499 | 14 | 0.883 | 9 | 0.780 | 9 | 0.382 | 9 |
| 990-2 | 91.1 | 6 | 106.2 | 7 | 0.472 | 16 | 0.876 | 14 | 0.780 | 8 | 0.407 | 7 |
| Zombi | 76.9 | 16 | 104.7 | 12 | 0.526 | 5 | 0.889 | 7 | 0.775 | 10 | 0.369 | 13 |
| Voore | 87.5 | 10 | 104.1 | 14 | 0.515 | 10 | 0.876 | 13 | 0.754 | 14 | 0.376 | 11 |
| Hiie | 95.5 | 2 | 104.5 | 13 | 0.532 | 4 | 0.879 | 11 | 0.758 | 13 | 0.360 | 14 |
| 876 | 93.0 | 4 | 102.8 | 16 | 0.516 | 9 | 0.871 | 15 | 0.738 | 15 | 0.357 | 16 |
| Runar | 97.7 | 1 | 103.1 | 15 | 0.568 | 2 | 0.870 | 16 | 0.716 | 16 | 0.360 | 15 |
| LSD | 1.81 | × | 0.48 | × | 0.031 | × | 0.001 | × | 0.019 | × | 0.020 | × |
| Mean | 88.7 | × | 106.0 | × | 0.519 | × | 0.886 | × | 0.785 | × | 0.405 | × |
| Environment | Grain Yield (GY) | Grain Protein Content (GPC) | ||||||
|---|---|---|---|---|---|---|---|---|
| RMSE | adj. R2 | RMSE | adj. R2 | |||||
| N75 | N150 | N75 | N150 | N75 | N150 | N75 | N150 | |
| GS25 | ||||||||
| EE_2021 | 35.09 | 46.49 | 0.26 ** | 0.16 * | 0.65 | 0.72 | −0.03 | 0.01 |
| EE_2022 | 32.00 | 31.62 | −0.03 | −0.03 | 0.56 | 0.71 | −0.03 | −0.01 |
| LV_2021 | 52.16 | 50.52 | 0.01 | −0.03 | 0.51 | 0.73 | 0.00 | 0.02 |
| LV_2022 | 79.26 | 61.47 | −0.01 | 0.14 * | 0.93 | 0.93 | 0.09 | 0.07 |
| LT_2021 | 57.96 | 50.18 | 0.10 * | 0.49 *** | 0.69 | 0.7 | −0.01 | −0.02 |
| LT_2022 | 70.02 | 71.97 | 0.04 | −0.03 | 0.63 | 0.58 | 0.05 | 0.16 * |
| NO_2021 | 46.24 | 66.82 | 0.00 | 0.00 | 0.93 | 1.48 | −0.01 | 0.08 |
| NO_2022 | 70.96 | 91.32 | 0.11 * | 0.23 ** | 1.23 | 1.18 | 0.12 * | −0.02 |
| GS55 | ||||||||
| EE_2021 | 38.68 | 43.98 | 0.1 * | 0.25 ** | 0.57 | 0.65 | 0.21 ** | 0.18 ** |
| EE_2022 | 31.81 | 24.68 | −0.02 | 0.37 *** | 0.53 | 0.65 | 0.06 | 0.15 * |
| LV_2021 | 47.66 | 43.2 | 0.17 * | 0.25 ** | 0.5 | 0.74 | 0.04 | −0.02 |
| LV_2022 | 76.99 | 66.32 | 0.04 | 0.00 | 0.97 | 0.91 | 0.01 | 0.11 * |
| LT_2021 | 45.60 | 42.24 | 0.44 *** | 0.64 *** | 0.7 | 0.64 | −0.03 | 0.13 * |
| LT_2022 | 41.58 | 67.02 | 0.66 *** | 0.10 * | 0.65 | 0.63 | −0.01 | 0 ns |
| NO_2021 | 42.29 | 49.90 | 0.16 * | 0.44 *** | 0.94 | 1.25 | −0.03 | 0.34 *** |
| NO_2022 | 52.59 | 66.73 | 0.51 *** | 0.59 *** | 1.02 | 1.12 | 0.39 *** | 0.08 |
| GS75 | ||||||||
| EE_2021 | 39.68 | 39.40 | 0.05 | 0.4 *** | 0.53 | 0.54 | 0.32 *** | 0.44 *** |
| EE_2022 | 32.00 | 23.56 | −0.03 | 0.43 *** | 0.55 | 0.66 | 0.00 | 0.12 * |
| LV_2021 | 43.17 | 36.41 | 0.32 *** | 0.47 *** | 0.44 | 0.69 | 0.26 ** | 0.11 * |
| LV_2022 | 70.83 | 61.00 | 0.19 ** | 0.16 * | 0.96 | 0.76 | 0.02 | 0.37 *** |
| LT_2021 | 44.18 | 47.45 | 0.47 *** | 0.54 *** | 0.69 | 0.65 | 0.00 | 0.12 * |
| LT_2022 | 29.96 | 53.13 | 0.82 *** | 0.44 *** | 0.66 | 0.63 | −0.03 | −0.01 |
| NO_2021 | 39.62 | 66.75 | 0.27 ** | 0.00 | 0.93 | 1.56 | −0.01 | −0.03 |
| NO_2022 | 36.08 | 53.46 | 0.77 *** | 0.74 *** | 0.88 | 1.02 | 0.55 *** | 0.23 ** |
| GS85 | ||||||||
| EE_2021 | 41.24 | 51.58 | −0.03 | −0.03 | 0.6 | 0.72 | 0.11 * | 0.01 |
| EE_2022 | 31.93 | 27.36 | −0.03 | 0.23 ** | 0.54 | 0.67 | 0.02 | 0.1 * |
| LV_2021 | 52.28 | 39.91 | 0.00 | 0.36 *** | 0.49 | 0.71 | 0.09 * | 0.07 |
| LV_2022 | 79.74 | 64.50 | −0.03 | 0.06 | 0.99 | 0.96 | −0.03 | −0.01 |
| LT_2021 | 60.01 | 62.53 | 0.03 | 0.2 ** | 0.7 | 0.67 | −0.03 | 0.07 |
| LT_2022 | 68.10 | 71.68 | 0.09 | −0.02 | 0.66 | 0.6 | −0.03 | 0.1 * |
| NO_2021 | 33.68 | 46.16 | 0.47 *** | 0.52 *** | 0.87 | 1.22 | 0.13 * | 0.37 *** |
| NO_2022 | 52.13 | 83.35 | 0.52 *** | 0.36 *** | 0.99 | 0.86 | 0.42 *** | 0.45 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jansone, Z.; Rendenieks, Z.; Lapāns, A.; Tamm, I.; Ingver, A.; Gorash, A.; Aleliūnas, A.; Brazauskas, G.; Shafiee, S.; Mróz, T.; et al. Phenotypic Variation and Relationships between Grain Yield, Protein Content and Unmanned Aerial Vehicle-Derived Normalized Difference Vegetation Index in Spring Wheat in Nordic–Baltic Environments. Agronomy 2024, 14, 51. https://doi.org/10.3390/agronomy14010051
Jansone Z, Rendenieks Z, Lapāns A, Tamm I, Ingver A, Gorash A, Aleliūnas A, Brazauskas G, Shafiee S, Mróz T, et al. Phenotypic Variation and Relationships between Grain Yield, Protein Content and Unmanned Aerial Vehicle-Derived Normalized Difference Vegetation Index in Spring Wheat in Nordic–Baltic Environments. Agronomy. 2024; 14(1):51. https://doi.org/10.3390/agronomy14010051
Chicago/Turabian StyleJansone, Zaiga, Zigmārs Rendenieks, Andris Lapāns, Ilmar Tamm, Anne Ingver, Andrii Gorash, Andrius Aleliūnas, Gintaras Brazauskas, Sahameh Shafiee, Tomasz Mróz, and et al. 2024. "Phenotypic Variation and Relationships between Grain Yield, Protein Content and Unmanned Aerial Vehicle-Derived Normalized Difference Vegetation Index in Spring Wheat in Nordic–Baltic Environments" Agronomy 14, no. 1: 51. https://doi.org/10.3390/agronomy14010051
APA StyleJansone, Z., Rendenieks, Z., Lapāns, A., Tamm, I., Ingver, A., Gorash, A., Aleliūnas, A., Brazauskas, G., Shafiee, S., Mróz, T., Lillemo, M., Kollist, H., & Bleidere, M. (2024). Phenotypic Variation and Relationships between Grain Yield, Protein Content and Unmanned Aerial Vehicle-Derived Normalized Difference Vegetation Index in Spring Wheat in Nordic–Baltic Environments. Agronomy, 14(1), 51. https://doi.org/10.3390/agronomy14010051