Abstract
In recent years, the relationship between plants and nanoparticles (NPs) has been the subject of extensive research interest. Hence, an ever-increasing number of perspectives connected with both the positive and adverse consequences of NPs application in plants are highlighted in this review. On the other hand, nanoparticles and their interactive effects on plants have raised concern regarding their harmful aspects. There are complex mechanisms evolved in plants for controlling the uptake, accumulation, and mobilization of nanoparticles that need to be discussed further, especially in the context of proteomics and genetic level. To fill this gap, there is a dire need for the integration and application of multiomics in plants to identify sensitive biomarkers responding to engineered NPs and to provide mechanistic insights in order to design safer and enhanced nano-enabled products for agriculture. For this purpose, transcriptomic technologies have essentially contributed to understanding of the molecular systems in plants against nanoparticle stress by laying out an association between gene expression and cell response. In light of this background, the current article attempts to summarize a variety of recent transcriptomic and proteomic contributions that have been made to establishing the genetic basis of nanoparticle uptake and the mechanism of stress response. The present article also looks at recent proteomic and transcriptomic studies to learn more about the intricate regulatory network that connects plant and nanoparticle stress responses.
1. Introduction
Nanotechnology is a rapidly emerging and multifaceted science with great potential and applications. The concern of the scientific community regarding its impact on biota has grown along with the technology’s rapid growth [1,2,3]. Given that manufactured nanomaterials (MNMs) or engineered nanoparticles (ENPs) play a significant role in this rapidly emerging science, the remarkable physicochemical properties of manufactured nanoscale materials (ENMs) are studied in precision agriculture to (i) increase nutritional quality and crop productivity [4], (ii) apply the controlled release and targeted delivery of agrochemicals [5,6], (iii) protect crops against pathogens [7,8], (iv) enhance resilience to environmental stress [9,10], and (v) monitor soil quality and detect stressors using sensitive nanosensors [11,12]. In addition to their benefits to the targeted crop, it is also possible to significantly reduce unwarranted exposure to nontargeted organisms and leaching into environmental waters. The concern of the scientific community regarding its impact on biota has grown along with the technology’s significant role in a variety of consumer goods, including biomedicine, cosmetics, and many others, and their massive atmospheric release is inevitable. Plant communities are particularly exposed to MNMs due to their restricted distribution [13,14,15]. There is a lot of literature on nanoparticles’ (NPs’) beneficial as well as phytotoxic implications for plant communities [16,17]. However, the best way to use nanotechnology in agricultural applications has been limited by a lack of mechanistic understanding of how plants absorb, mobilize, and respond biologically to NPs. The type, particle size, composition, and specific surface area of these nanoparticles are the primary determinants of their effects on plants [18]. Plants’ reactions to NPs are exceptionally varied and subject to natural circumstances, plant species and age, duration and dose of exposure, and NP properties, such as structure, size, surface covering, morphology, stability, or disintegration, among different factors [19]. The integrated implications for specific NPs, including phytotoxic and beneficial effects on plants, have been precisely discussed in the literature [20,21,22]. Titanium oxide (TiO2) NPs, for example, lessen UV-B-induced oxidative stress [23]; they also stimulate the production of organic form of nitrogen from inorganic form, supporting nitrate assimilation and acquisition [24], and increasing photosynthesis rate [25]. In contrast, water acquisition and transpiration rate in corn plants is negatively impacted by TiO2 NPs [26]. In a similar manner, carbon nanotubes (CNTs) belong to a class of NPs that are capable of entering the seed coat and facilitating the acquisition of water by tomato plant seedlings, which is necessary for germination [27]. The synthesis of NPs, which exhibit fascinating effects on living systems and possess a variety of properties, has been attempted in a number of ways [28,29,30]. Likewise, many reports demonstrate the potential impacts of NPs on plants in terms of morphology and physiology [31], whereas there have been few endeavors examining the impacts of MNMs on plants at the genetic level. For example, soybean seedlings have been shown to be genotoxic to cerium oxide (CeO2) NPs, and onion seedlings have been shown to be genotoxic to silver (Ag) NPs; both of these nanoparticles have a gene-level effect on seedling growth [32]. In addition, the Oryza sativa seedling’s flowering time was extended when carbon (C) NPs were combined with natural organic matter [33]. Zinc oxide (ZnO) NPs, which harm the epidermal and cortical cells of Lolium perenne, are another example [34]. It is crucial to establish the interactive mechanism to study the relation between plants and their responses against nanoparticles, especially related nutrient flux, photosynthetic efficiency, and oxidative stress protections.
There are transcriptomics studies that assess the potential toxicity of nanomaterials in plants, and many of them focus on their impact on genes that respond to stress [35,36]. Because they show how NPs affect gene expression, the transcriptomic and proteomic studies add new information to our understanding of how plants interact with NPs. Carbon nanotubes (CNTs) were used in a study on tomato seedlings, and upregulation of various genes involved in stress-related processes was uncovered through microarray analysis [37].
A comparison examination of changes in total expression, however, points to a minimal effect on the transcriptome [38,39]. Biotic and abiotic stresses like pathogens, salinity, or drought may cause higher transcriptional reactions than nanoparticles [40,41]. It is obvious that plants did not have the opportunity to create unique gene regulation in response to novel nanomaterials; instead, they use regulatory circuits that are shared with other stress responses [42]. The suppression of genes for root development and pathogen response is a common consequence of abiotic stress [43]. There are reviews of further works that support these findings [44,45]. In a similar vein, additional research has shown that in Arabidopsis thaliana seedlings, gene expression for the peroxide-scavenging enzyme was influenced by CNTs [46]. Injection of single-walled carbon nanotubes (SWCNTs) to leaves increased transcription level for the two enzymes: ascorbate peroxidase (APX1) and manganese superoxide dismutase (Mn SOD1). Arabidopsis thaliana was the subject of a study using ZnO and TiO2 NPs, with the former exhibiting phytotoxicity in the aforementioned plant and the latter on depicting profoundly significant positive effect on the plant [24,34,47,48,49]. Researchers have utilized advanced techniques to find NP uptake in plant tissues upon root or foliar exposure [50,51]. Such examinations have really demonstrated that some NPs enter through roots or passes on and can cross natural hindrances to move inside plants, in contrast to their conventional analogues [52,53]. But the molecular drivers of the uptake and transport of NMs in plants remains inconclusive.
To fully utilize the potential of NPs for sustainable crop production, it is essential to have a comprehensive understanding of the biological networks at multiple levels [54]. In recent years, the examining of NP–plant interaction has developed from conventional, single endpoint measures to discovery-oriented, high-throughput system biology approaches, referred to as “omics”. Analytical methods and bioinformatics tools have increased in sensitivity and accuracy [55]. “Omics” refers to the undistinguished screening of biomolecules in an organism, specifically genes (genomics), mRNA (transcriptomics), and proteins (proteomics). With the rising need for mechanistic comprehension of complex agronomic qualities and crops’ reaction to NPs exposure, omics technologies have picked up speed in agriculture and nanotoxicity studies. This worldview helps in framing a hypothesis by checking the reaction of biomolecules upon perturbing biological cycles with NPs, followed by a combination of worldwide datasets onto pathways utilizing advanced bioinformatics [56,57]. In this manner, researchers are expected to provide more comprehensive knowledge to figure out the plant–NP interface. This review throws a wide beam of light over the previous endeavors made by researchers, and it also identifies a need for additional research; however, side effects such as genotoxic effects posed by nanoparticles on plants need to be discovered through relevant research studies. Acknowledgment of the underlying molecular systems in plants will provide cues to formulating NPs for explicit applications like expanding strength to ecological circumstances, designated delivery of nutrients, and NP-empowered biosensing.
2. Plant Interaction with Nanoparticles
Materials react substantially differently at the nanoscale than they do at larger dimensions, and they frequently exhibit unusual chemical and physical characteristics [41]. This makes the transformation of matter through nanotechnology a promising area for the creation of new goods and procedures. On the other hand, their distinct features enable innovative materials to interact with biological systems in unanticipated ways [42,43]. Different plants interact with NPs through chemical or physical contact (Figure 1). These interactions result in physical and chemical signaling, which generates new molecules, such as reactive oxygen species (ROS), which may pose serious damage to plants [58]. These ROS cause oxidative stress and damage [59], altering the movement of ions throughout the cell membrane [60], and peroxidation of fatty acids [61]. The key factors that determine plant–NP interaction are the nanoparticle’s characteristics, environmental factors, and plant species [62]. It is still unclear how the first NP contact point in plants chooses the traffic route. Nevertheless, a number of studies have suggested a few paths that could enhance our comprehension of nanoparticle trafficking. The well-known channels that determine the entry route are direct application, contaminated sediments or soil, accidental release, and atmospheric fallouts. Xylem, cortex, and pericycle play crucial roles in facilitating nanoparticles’ entry into the plant via the root [63]. Particle entry at various key checkpoints are controlled by a few key components. The cell wall is the first checkpoint, and it functions as a structural sieve, allowing only NPs with compatible pore sizes to enter. Only NPs with pore sizes comparable to those of the cell wall were able to successfully travel outside the cell membrane through the cell wall [13]. Through the root system, tiny NPs can quickly enter plant cells, while larger NPs can only enter through larger openings such as stomata, stigma, and hydathodes. The cell rarely receives larger NPs; as a result, there is no change in cell metabolism [64]. Metal-based NPs change into reactive metal ions when they enter plant cells, react with various functional protein groups, and disrupt the biochemical activity. These NPs easily adsorb various inorganic ions and molecules from the soil or nutrient medium due to their large surface area, leading to indirect toxicity symptoms such as wilting and chlorosis [65]. The reaction between NPs and plants determines their accumulation and transfer across various trophic levels.
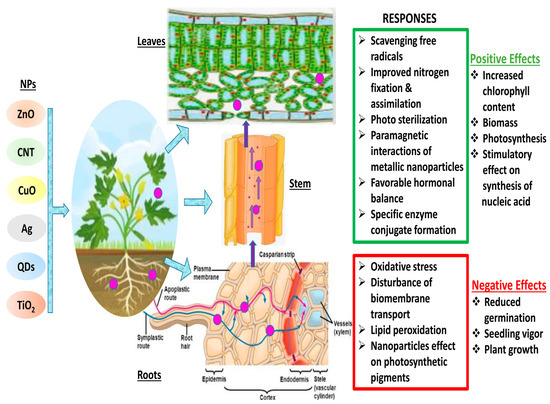
Figure 1.
Schematic diagram of plant—NPs interactions and various responses. (Red color indicates the negative impacts of NPs interaction in plants, whereas green color shows the positive interactions of NPs in plant cells)
3. Uptake and Translocation of NPs in Plants
Through roots or stomata or epidermal absorption in leaves, NPs enter the plant because of their quite small size. Through the apoplastic or symplastic pathway, NPs enter the cells and reach various plant systems in the case of roots. NPs enter the cell membrane through the cell wall in the root’s epidermis, where they are symplastically transported to the vascular system. Pore size is a factor in movement through the cell membrane or cell wall [66,67]. In addition to travelling through the symplastic pathways, NPs also travel via the apoplastic pathway to reach the vascular system. After they enter, NPs are moved to the plant’s leaves, where they come in contact with chloroplast, which results in the disruption of the plant’s physiology [68]. A brief explanation of how NPs make their way to a plant’s system is provided in the schematic illustration in Figure 1.
3.1. Leaf
Leaves contain two entry points: one is via absorption through the leaf’s epidermis and the other through the stomata. In a study, it was demonstrated through confocal microscopy that NPs enter Vicia faba at a specific size through the stomata; NPs of 1.1 μm did not penetrate the cell system; those of 43 nm did. Cerium (37 nm) enter the plant system but are not translocated to the stem, as observed in leaves of Zea mays using electronic and confocal microscopy and mass spectrometry [69]; however, this study was unable to determine whether the stomata or the leaf epidermis were the entry points [70]. Similarly, in another study using X-ray and electron fluorescence microscopy on Arabidopsis seedlings, NPs entered the epidermis through stomata and were absorbed by endocytic vessels to reach the palisade layer [71,72]. Several studies have been carried out to investigate entry through the leaf’s epidermis in addition to those focused on stomatal entry. The work carried out by some researchers, using nonionic colloidal solutions of iron, zinc, and manganese, demonstrated that these are translocated after being absorbed by the leaf epidermis [73]. Another study found that both bulk salt and Ag NPs (both with 47.9 nm hydrodynamic diameter) entered Lactuga via absorption through the cuticle and stomata before being transported to the vascular system. The study also unveiled that Ag+ is interconverted into Ag NPs by binding with thiol groups [74].
3.2. Roots
Maize, rapeseed, spinach, and desert plants have been the subject of numerous studies [73,75,76,77,78,79,80] and it has been demonstrated that NPs make their way to the root system through the epidermis and then translocate to the stems and leaves [81]. In another study, the entry of iron oxide (Fe2O3) NPs through the root epidermis was found in pumpkin plants grown in an aqueous medium, and further translocation to shoots and leaves was observed as well [82]. In another study, 2.8 nm sized Ti nanoconjugates were found to be penetrated into root cells and transported to nucleus and vacuoles in Arabidopsis thaliana [71]. Transmission electron microscopy revealed the adherence and accumulation of Ti NPs in the cell wall of roots, as well as further translocation of particles in the cortex and then to the vacuoles [83]. In addition, a number of experiments using confocal microscopy, X-ray fluorescence, and coupled plasma optical emission spectrometry reported that NPs entered the plant via the roots before being transferred to the shoots via the apoplastic pathway [84,85,86]. Additionally, analysis on soil systems averred that organic substances in the soil influence NP mobility.
4. Mechanism of Phytotoxicity Induced by Nanoparticle–Plant Interaction
The phytotoxicity caused by NPs increases the production of ROS like hydrogen peroxide (H2O2), superoxide radical (O2•−), singlet oxygen (1O2), and hydroxyl radical (OH•). The distortion in flow of electrons results in the production of ROS in the chloroplast and mitochondria. Resultantly, oxygen takes the electron that has escaped from the electron transport chain and generates the superoxide radical O2•−. Through the action of the enzyme SOD, O2− becomes H2O2. Although ROS function as a signaling molecule at a low level, an increase in ROS causes negative effects known as oxidative stress. When the amount of ROS exceeds the defense mechanisms, oxidative stress occurs, and it poses serious threat to cells, i.e., damage to DNA, lipid peroxidation and protein oxidation, membrane damage, electrolyte leakage, and, consequently, cell death [87,88,89]. The hydroxyl radical is the most reactive ROS, which is produced by a Fenton reaction. Because it has one unpaired electron, it reacts with all biological molecules causing cellular damage like membrane and protein damages [90,91]. Damage to the fundamental structure of the cell membrane results in membrane leakage when lipids are peroxidized [92]. It can be said that NPs may speed up lipid peroxidation and increase the production of ROS. It was proposed in a study that phytotoxicity is caused by lipid peroxidation and ROS formation in research on ZnO NPs in ryegrass [34]. Later, it was discovered that the accumulation of H2O2 in plant roots treated with La2O3 and CeO2 NPs using the 3,3′-diaminobenzidine-a ROS-sensitive dye demonstrated that a deep brown-colored insoluble product was produced when NPs were used [93]. Similarly, treatment with Ag NPs up to 10 mg mL−1 produced ROS, which in turn decreased the number of viable cells [94]. Plants have an antioxidant system that includes nonenzymatic (ascorbate, nonprotein thiol, glutathione, cysteine, proline) and enzymatic (guaiacol peroxidase, catalase (CAT), SOD, APX, glutathione reductase (GR), glutathione peroxidase (GPx), monodehydro ascorbate reductase, glutathione S-transferase) elements. Researchers also found that treated castor bean (Ricinus communis) seeds produced more ROS and that POD (peroxidase) and SOD (superoxide dismutase) activity also increased. SOD, which converts O2− into less harmful H2O2, is the first line of defense among enzymatic antioxidants [95]. It is also reported that 0–1000 mg L−1 nickle oxide (NiO) NPs had a different impact on SOD, at the low levels of treatment, SOD level increased, while at a high level of treatment, SOD activity decreased [96]. Similar to that, in a finding, SOD activity increased at low exposure levels to CeO2 NPs, but that enzyme activity did not change at high treatment levels [97]. Some researchers utilized 100 mg L−1 Al2O3 NPs exposed on onion root and produced similar results [98]. Moreover, CAT also is an essential enzyme that is involved in stress-induced ROS detoxification and produces O2 and H2O from H2O2 [99]. The ascorbate–glutathione (AsA–GSH) cycle, which includes GR, DHAR, and APX, controls the excessive production of ROS in plants, and is centrally regulated by APX, which also targets H2O2 but at the cost of ascorbate [100]. It also is an essential enzyme that is involved in stress-induced ROS detoxification and produces O2 and H2O from H2O2. Similarly, it was reported that treatment with Fe3O4 NPs, CeO2 NPs, and Co3O4 NPs increased the activity of CAT, while treatment with CuO NPs, Au NPs, Fe3O4 NPs, MnO2 NPs, Co3O4 NPs increased the activity of GPX, and treatment with CeO2 NPs and Pt NPs accelerate the activity of SOD [101]. It was also found that treatment with NiO NPs increased SOD and CAT activity in tomato depending on the dose increase [96]. Additionally, in Phaseolus vulgaris, phytotoxicity was caused by exposure to CeO2 NP, which disrupted the antioxidant defense system [102]. CeO2 NP exposure significantly improved the activities of POD, CAT, SOD, and APX in A. thaliana, while an increase was only observed for POD and SOD [97]. Likewise, Nd2O3 NPs decreased pumpkin growth, nutrient levels (Ca, K, S, and Mg), and APX, CAT, and SOD activity [103]. Nonenzymatic antioxidants like GSH and ASC participate in the ROS detoxification process, just like enzymatic antioxidants do. On treatment with CuO NPs, consistent increases in AsA were observed [104]. GSH is another important thiol group metabolite that helps to bind pesticides and metals. Through the AsA–GSH cycle, GSH also aids in AsA regeneration [105]. According to several studies on metal and metal-based NP phytotoxicity, it can be concluded that NPs can cause oxidative stress in many plant species by producing ROS that are balanced by the plant’s defense system [104,106,107,108,109]. In recent years, the study of NP phytotoxicity has made rapid progress; however, our understanding of ROS-dependent damage and its impacts on plants are incomplete and ambiguous in many ways. As a result, it is necessary to investigate the entire antioxidant defense system and enzymatic mechanism of nanotoxicity in plants.
5. Nanoparticle–Plant Interaction: Transcriptomic and Proteomic Debate
Plants are portrayed as the most important link between the environment and the biosphere; therefore, in order to conduct pertinent environmental assessments, it is necessary to understand how nanoparticles affect them. Compared to their bulk metal counterparts, Ag, ZnO, and SnO2 NPs can alter growth rate, cell viability, and chlorophyll at the cellular level [110,111,112]. The proteomics platforms that have evolved over the past 20 years—gel, label, gel free/label free, targeted—are used to describe protein profiles, posttranslational changes, and interactions [45]. Isolation, separation, visualization, and precise protein identification are among the many aspects of any proteomic technique that ultimately determine its effectiveness [44]. Despite recent developments, more attention needs to be paid to protein extraction procedures, especially for proteins that are hydrophobic and large molecular mass proteins [39].
Changes in organelles like chloroplasts and vacuoles, as well as the aggregation of NPs close to the plasma membrane and cell wall, have been observed at the subcellular level [113,114]. When compared to bulk metals, NPs are considered to significantly increase the concentration of ROS and peroxidation of lipids at the molecular level [111,115,116,117]. Studying higher plants physiologically, it was discovered that 40 nm sized NPs are uptaken by the root via the symplastic pathway through the vascular system; however, the apoplastic pathway is where larger NPs accumulate. After being exposed to clay NPs and TiO2, the cell walls of maize roots had smaller pores, reduced transpiration, and suppressed hydraulic conductivity [26]. Rice has demonstrated C-based NMs translocation, uptake, and generational transmission [33]. New studies on the plant Arabidopsis thaliana about its gene expression have shown the novel interactions mechanisms between plants and nanoparticles [118,119]. When subjects were exposed to fullerene of different sizes for the long duration, TiO2 NPs, Ag NPs, and ZnO NPs, these studies observed changes in gene expression. In addition, oxidative stress and antioxidant enzymes were found to unveil the mechanisms underlying ZnO NP toxicity [120]. In this context, the major advancements that have been made in plants’ transcriptomic and proteomic levels since the use of nanoparticles are highlighted in the sections that follow.
5.1. Genomic Insight into Nanoparticle–Plant Interaction
Since nanotechnology has been recognized as the most cutting-edge, rapidly developing, and labor-saving technology by the scientific community as of late, it now has applications in almost every field of science. The toxicity caused by the constant deposition of NPs in the environment is not ignorable, especially in terms of plant performance. NPs interaction at genomic level is shown in Figure 2. A bulk of the literature demonstrates the morphological and physiological phytotoxic effects of NPs, but very little demonstrates the gene-level toxicity of NPs. Researchers reported that CuO NPs were toxic to some agricultural crops like Loliumrigidum, Loliumperenne and Raphanussativus, Lolium rigidum, Lolium perenne, and Raphanus sativus, and after prolonged NP exposure, DNA was destroyed. The deposition of compounds that have been altered by oxidation led to the formation of mutagenic DNA lesions, which in turn caused rapid disruptions in plant growth [121]. Similarly, ZnO NP toxicity in wheat seedlings was examined and it was discovered that nitric oxide (NO) mitigates this toxicity in wheat effectively [122]. Additionally, figuring out the NPs’ genotoxic endpoints is a concern because they have been linked to metal’s genotoxic effects in plants [123], but any generalizations regarding NP-induced phytogenotoxicity must be handled with care.
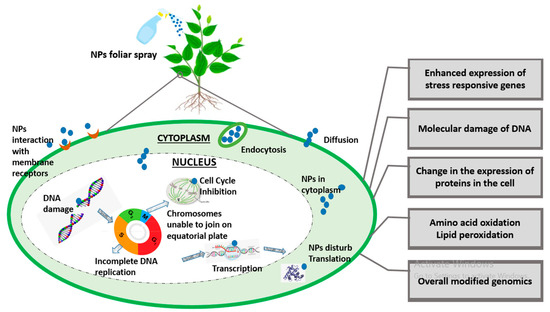
Figure 2.
A brief illustration of genomic insight into plant–-NPs interaction.
5.2. Genes Regulation with Respect to NP-Induced Genotoxicity
At the molecular level, the basis of nanoparticle-induced phytotoxicity in higher plants is still poorly understood. However, transcriptomic analysis, upon NP exposure, has presented information about the up- and downregulation of different genes, which has provided some knowledge regarding NP toxicity at the genetic level in higher plants. For example, it was experimented that when maize root tissue was exposed to single-walled carbon nanotubes (SWCNTs), the genes for RTCS and SLR1 were upregulated and the genes for RTH3 and RTH1 were downregulated. Aquaporin also plays a significant role in nanoparticle acquisition. A variety of stress parameters control or regulate many stress-inducible genes in plants [124]. In another study, it was found that an increase in aquaporin production in tomato seedlings upregulated the water channel and NtLRX1 genes [125]. Some researchers treated Eruca sativa with Ag NPs, recorded the proteomic profile of Ag-treated plants, and different gene and protein expressions. Ag NPs effected the number of proteins in the plant cell’s vacuole and endoplasmic reticulum (ER), indicating that these organelles are Ag NPs’ primary targets [126]. When Arabidopsis plants were treated with varying concentrations of gold (Au) nanoparticles, similar expression results for micro RNAs (miRs) were observed [127]. When Au NP exposure was compared to a control, expression of some microRNAs, especially miR164, miR395, miR167, miR408, and miR398, were found to be downregulated. However, this study’s findings are in direct opposition to those of Burklew, Ashlock [128] regarding tomato plant seedlings in significant ways. This further proves that NPs have a species-specific phytotoxicity. Moreover, NPs alter gene expression even at very low concentrations, suggesting that the impact of NP-caused toxicity at cellular level can be gauged using gene expression analysis [129,130]. In this way, gene expression analysis assists in the identification and development of transgenic plants that are sensitive or tolerant to certain stresses [131,132]. As a result, genomics analysis is profoundly useful for determining the mechanism of NP toxicity and relating any physiological or morphological changes at the genetic level in plants. The fundamental processes of electron transport chain signaling are disrupted by the NP-induced phytotoxicity, which ultimately affects the organism’s cell cycle. Moreover, the experimental proofs of the NPs induced gene regulation is summarized in the Table 1.

Table 1.
Genomic analysis of different nanomaterials’ interaction with plants [133].
5.3. Transcriptomic Insight into Nanoparticle–Plant Interaction
Numerous researchers have reported evidence of nanoparticle-induced alterations in plant gene expression. It is averred that exposure to ZnO and TiO2 altered the gene expression found in Arabidopsis thaliana roots. They concluded that ZnO NPs are the most toxic NPs among the three abovementioned NPs because they induced downregulation and upregulation of 826 and 660 genes for stress regulation, respectively, although the least toxic TiO2 NPs only had an impact on 74 and 80 genes, respectively [118]. Despite the researchers’ clear demonstration of up- and downregulation of genes caused by the phytotoxicity of nanoparticles, comprehensive details have not yet been portrayed. In a recently conducted study, they exposed A. thaliana to various sizes and types of NPs under abiotic and biotic stresses to observe the early changes in gene expression. As a general response to nanoparticle-induced stress, they observed transcriptional repression and downregulation of gene expression [134]. In another study, examination of proteomic, ionomic, and transcriptomic shifts have put Arabidopsis into focus to determine how Au NPs affect growth and development. The expression of the MYB, BHLH, and WRKY gene families, which are involved in homeostasis of some essential metals and Fe, differed according to the transcriptome analysis of Arabidopsis roots [153]. It was inferred that Au impacted the normal homeostasis of essential elements like Mn, Fe, and Zn. By observing genetic and physiological responses of plants to Au, it has also been discovered that Au is uptaken by Arabidopsis and then transported to the shoot. In addition, it has been observed to have an effect on the transcription of a specific group of genes, particularly cation transporters, with 869 genes being upregulated and 851 genes being downregulated. IRT1 and IRT2, iron-regulated transporters, have shown the lowest expression of genes. Downregulation of eleven aquaporin genes was also reported in this study. Metal uptake, particularly that of heavy metals, is thought to involve a number of these genes [154]. Numerous studies have looked into the different kinetics that NMs use to acquire, translocate, and internalize NMs, as well as the cytotoxicity that is recorded at the level of gene expression. These studies also found that NMs affected plant growth and development, along with a number of other morphological and physiological effects. Although the genetic processes underlying these effects have not yet been discovered, it has been observed that these effects are species-specific. A summary of the various other transcriptomic analyses that have been carried out with NPs, as well as their effects on cell systems, can be seen in Table 2. The transcripts of genes involved in vacuolar proton or cation exchange, hormonal responses, antioxidant systems, pathogen-responsive genes, decreases in photosynthesis genes’ transcript formation, aquaporin genes, as well as biogenesis and cell organization genes, were found to be upregulated in NP-mediated toxicity.
Table 2.
Transcriptomic analysis of different nanomaterials’ interaction with plants.
5.4. Proteomic Insight into NP–Plant Interaction
Over the past few decades, various multidisciplinary studies on the impacts of Ag NPs on plants have shown that they are toxic at many levels—cellular, morphological, physiological, etc. However, only a small number of studies have examined the detrimental proteomic effects of Ag NP stress on plants. In an effort to carry away the toxicity posed by Ag NPs, a gel-based proteomic study of Oryza sativa seedlings was recently conducted [157]. Ag NP-receptive proteins of were reported to be primarily associated with Ca2+ signaling and regulation, the oxidative stress response pathway, transcription, protein degradation, cell wall synthesis, cell division, and apoptosis. Specific defense-related proteins, i.e., SOD, glutathione S-transferase (GST), and L-ascorbate peroxidase, were found to be overexpressed after exposure to Ag NP toxicity. Resultantly, this accelerated the generation of reactive oxygen species (ROS) under Ag NP treatment. Previous research has suggested that secreted ions or Ag NPs could disturb the cell’s metabolism by binding to Ca2+/Na+ATPases and Ca2+ ion channels—the secondary messenger receptors. In a similar manner, a proteomic analysis of Eruca sativa roots exposed to AgNO3 and Ag NP stress revealed that both bulk metal and NPs can alter the structure of proteins associated with redox regulation and ultimately disrupt cellular homeostasis [126]. However, the fact that the exposure of Ag NPs is sufficient to alter the membrane proteins of vacuoles and endoplasmic reticulum suggests that these NPs primarily target both of these specific organelles. According to these findings, Ag NP phytotoxicity is caused by its unique chemical and physical properties rather than its continuous release to the atmosphere [126]. The agricultural sector is an important field in terms of the application of gold nanoparticles. After exposing Arabidopsis thaliana to Au NPs, a proteome analysis revealed the induction of glutathione S-transferases in the shoot. Additionally, researchers have also suggested that auxin, the plant hormone, plays a significant role in shaping Au-induced root system architecture [153]. In a similar way, Ag NPs at three distinct particle sizes (2, 15, and 50–80 nm) and concentrations (0.2, 2, and 20 ppm) were used in a series of experiments on soybean seedlings [158]. They demonstrated that, under flood stress, Ag NPs of 15 nm particle size promoted soybean seedling growth rate and germination potential in comparison to larger and smaller Ag NPs. The proteins altered by the exposure of Ag NPs were mostly linked to stress signaling, the detoxification pathway, and cellular metabolism. The concentration of these proteins increased under flood stress, but the proteins rapidly decreased when exposed to Ag NPs. Additionally, treatment with Ag NPs demonstrated a shift in metabolism from fermentative to cellular processes. According to the findings, soybean seedlings subjected to Ag NP treatment under flooding stress experience less oxygen loss as a result of treatment with Ag NPs, which have a particle size of 15 nm and 2 ppm. Additionally, soybean seedlings were found to be damaged at elevated Ag NP concentrations with particle sizes of 15 nm [158]. A comparative study with Ag, Al2O3, and ZnO NPs on two-day-old soybean seedlings subjected to flooding stress was recently conducted [159]. Surprisingly, treatment with 50 ppm Al2O3 NPs resulted in increased vigor and growth in soybeans. The primary functions of these Al2O3NP-responsive proteins were lipid metabolism, glycolysis, protein degradation, and protein synthesis. Under treatment with Al2O3 NPs, a fivefold increase in the negative NmrA-like transcriptional regulatory protein family was also observed. The proteomic results suggest that the growth of flood-stressed soybean seedlings may be attributed to regulation of energy metabolism and reduced proliferation of root cells. Similar to Ag and Au NPs, various cell systems have been used to investigate other NP toxicities. For instance, it has been demonstrated that CeO2 NPs inhibit maize growth by hampering stress-responsive protein expressions like CAT, heat shock protein, and APX [84]. A brief description of the effects that some other NPs have on cell systems is provided in Table 3. Mediation of NP toxicity has been reported by the decline and destabilization in cell membrane potential, lowering ATP levels in cells, elevating stress-responsive and quorum-sensing proteins, expression of oxidative stress-tolerant proteins, Ca2+ release and its signaling, transcriptional proteins, as well as downregulating essential proteins related to growth-regulating mechanisms like cell metabolism, photosystem-II functioning, and sulfur assimilation. A brief description of NPs interaction at transcriptomic and proteomic level is shown in Figure 3.
Table 3.
Proteomic analysis of different nanomaterials interaction with plants.
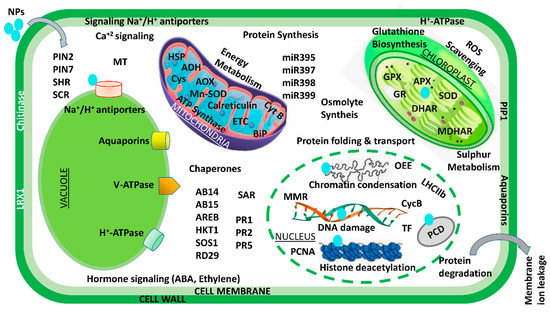
Figure 3.
An illustration of transcriptomic and proteomic analysis of the plant–-NPs interaction. ABI: ABA-insensitive; AOX: alternate oxidase; ADH: alcohol dehydrogenase; AREB: ABA-responsive element-binding proteins; APX: ascorbate peroxidase; ARGOS: auxin-regulated gene involved in organ size; CycB: cyclin B; BiP: binding immunoglobulin proteins; Cys: cysteine synthase; GR: glutathione reductase; ETC: electron transport chain; GlyII: glyoxalase II; HKT1: high-affinity K+ transporter 1; GPX: glutathione peroxidase; HSP: heat shock protein; miR: microRNA; MDHAR: monodehydroascorbate reductase; MMR: mismatch repair gene; OEE2: oxygen-evolving complex 2; MT: metallothionein; PCNA: proliferating cell nuclear antigen; PIN: peptidyl–prolyl cis–trans isomerase NIMA-interacting; PCS1: phytochelatin synthase 1; PIP1: plasma membrane intrinsic protein 1; RbcL: large subunit of Rubisco; PR: pathogenesis-related; RD29A: responsive to desiccation 29A; SAR: systemic acquired resistance; ROS: reactive oxygen species; SCR: scarecrow; SOS1: salt overly sensitive 1; TF: transcription factors; SOD: superoxide dismutase; SHR: short root. Modified from [133,160].
6. Conclusions and Future Perspectives
There connection between plants and nanomaterials in food and the agro-industry is gaining much interest and support due to the vast applications of nanomaterials. Due to their huge applications in various walks of life, i.e., household and industry, nanoparticles are detrimental to the environment and hazardous to human health. Although nanomaterials have been viewed as transformative agents, their potential long-term impact on environment and human health is not yet completely understood. Legislators still lack relevant data to build a regulatory scientific framework for the commercialization and release of nanoparticles. Various research studies, in particular, focus on the profound toxic impacts of nanoparticles in microorganisms, but toxic effects on plants are devoid of substantial research knowledge. The weak domain of plant studies in this regard is genotoxicity caused by nanoparticles. Hence, this article is an essay that unveils the role of transcriptomic and proteomic contributions in this need of hour to decipher the genetic basis of NP absorption and the stress response mechanism. However, a small study about proteomics was conducted in this regard, which showed the complex response of plants towards nanomaterial stress and the manifestation of differential expression of some proteins that augment biological reactions, for instance, detoxification, oxidation reduction, signal transduction, the hormonal signaling pathway, etc. Proteomic studies on silver nanoparticles have recently been performed, such as induced phytotoxicity depicted by particle size as a vital factor that impacts the magnitude of cell kinetics in plants. Every aforementioned piece of collected information provides further comprehensive, detailed, and elliptical patterns of plant responses against nanoparticle stress.
Author Contributions
Conceptualization, Z.A. and M.H.; writing—original draft preparation, N.M., W.G. and A.E.-K.; writing—review and editing, Z.A., M.H. and F.Z.; visualization, Z.A. and M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All information is available in this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Patel, M.; Patel, R.; Rai, M. Nanomedicines: Applications and Toxicological Concerns. In Nanotechnology in Medicine: Toxicity and Safety; Rai, M., Patel, M., Patel, R., Eds.; John Wiley & Sons Ltd.: West Sussex, UK, 2021; Volume 1, pp. 1–28. [Google Scholar]
- Abideen, Z.; Hanif, M.; Munir, N.; Nielsen, B.L. Impact of nanomaterials on the regulation of gene expression and metabolomics of plants under salt stress. Plants 2022, 11, 691. [Google Scholar] [CrossRef] [PubMed]
- Usman, M.; Farooq, M.; Wakeel, A.; Nawaz, A.; Cheema, S.A.; ur Rehman, H.; Ashraf, I.; Sanaullah, M. Nanotechnology in agriculture: Current status, challenges and future opportunities. Sci. Total Environ. 2020, 721, 137778. [Google Scholar] [CrossRef] [PubMed]
- Munir, N.; Hanif, M.; Dias, D.A.; Abideen, Z. The role of halophytic nanoparticles towards the remediation of degraded and saline agricultural lands. Environ. Sci. Pollut. Res. 2021, 28, 60383–60405. [Google Scholar] [CrossRef] [PubMed]
- Walker, G.W.; Kookana, R.S.; Smith, N.E.; Kah, M.; Doolette, C.L.; Reeves, P.T.; Lovell, W.; Anderson, D.J.; Turney, T.W.; Navarro, D.A. Ecological risk assessment of nano-enabled pesticides: A perspective on problem formulation. J. Agric. Food Chem. 2017, 66, 6480–6486. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; White, J.C.; Wang, Z.; Xing, B. Nano-enabled fertilizers to control the release and use efficiency of nutrients. Curr. Opin Plant Sci. Health 2018, 6, 77–83. [Google Scholar] [CrossRef]
- Munir, N.; Gulzar, W.; Abideen, Z.; Hancook, J.T.; El-Keblawy, A.; Radicetti, E. Nanotechnology improves disease resistance in plants for food security: Applications and challenges. Biocatal. Agric. Biotechnol. 2023, 51, 102781. [Google Scholar]
- Kah, M.; Tufenkji, N.; White, J. Nano-enabled strategies to enhance crop nutrition and protection. Nat. Nanotech. 2019, 14, 532–540. [Google Scholar] [CrossRef]
- Bora, K.A.; Hashmi, S.; Zulfiqar, F.; Abideen, Z.; Ali, H.; Siddiqui, Z.S.; Siddique, K.H. Recent progress in bio-mediated synthesis and applications of engineered nanomaterials for sustainable agriculture. Front. Plant Sci. 2022, 13, 999505. [Google Scholar]
- Jacobson, A.; Doxey, S.; Potter, M.; Adams, J.; Britt, D.; McManus, P.; McLean, J.; Anderson, A. Interactions between a plant probiotic and nanoparticles on plant responses related to drought tolerance. Indust. Biotech. 2018, 14, 148–156. [Google Scholar] [CrossRef]
- Giraldo, J.P.; Wu, H.; Newkirk, G.M.; Kruss, S. Nanobiotechnology approaches for engineering smart plant sensors. Nat. Nanotech. 2019, 14, 541–553. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Dev, A.; Karmakar, S. Nanosensors and nanobiosensors in food and agriculture. Environ. Chem. Lett. 2018, 16, 161–182. [Google Scholar] [CrossRef]
- Navarro, E.; Baun, A.; Behra, R.; Hartmann, N.B.; Filser, J.; Miao, A.-J.; Quigg, A.; Santschi, P.H. Environmental behavior and ecotoxicity of engineered nanoparticles to algae, plants, and fungi. Ecotoxicology 2008, 17, 372–386. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K. Silicon nanoparticles (SiNp) alleviate chromium (VI) phytotoxicity in Pisum sativum (L.) seedlings. Plant Physiol. Biochem. 2015, 96, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K. Silicon nanoparticles more efficiently alleviate arsenate toxicity than silicon in maize cultiver and hybrid differing in arsenate tolerance. Front. Environ. Sci. 2016, 4, 46. [Google Scholar] [CrossRef]
- Mathur, P.; Chakraborty, R.; Aftab, T.; Roy, S. Engineered nanoparticles in plant growth: Phytotoxicity concerns and the strategies for their attenuation. Plant Physiol. Biochem. 2023, 199, 107721. [Google Scholar] [PubMed]
- Fernández, M.D.; García-Gómez, C. Impact of Emerging Metal-Based NPs on Plants and Their Influence on the Phytotoxicity of Other Pollutants. In Emerging Contaminants and Plants: Interactions, Adaptations and Remediation Technologies; Springer: Berlin/Heidelberg, Germany, 2023; pp. 29–66. [Google Scholar]
- An, C.; Sun, C.; Li, N.; Huang, B.; Jiang, J.; Shen, Y.; Wang, C.; Zhao, X.; Cui, B.; Wang, C. Nanomaterials and nanotechnology for the delivery of agrochemicals: Strategies towards sustainable agriculture. J. Nanobiotech. 2022, 20, 11. [Google Scholar]
- Ma, C.; White, J.C.; Zhao, J.; Zhao, Q.; Xing, B. Uptake of engineered nanoparticles by food crops: Characterization, mechanisms, and implications. Annu. Rev. 2018, 9, 129–153. [Google Scholar] [CrossRef]
- Khan, M.; Khan, M.S.A.; Borah, K.K.; Goswami, Y.; Hakeem, K.R.; Chakrabartty, I. The potential exposure and hazards of metal-based nanoparticles on plants and environment, with special emphasis on ZnO NPs, TiO2 NPs, and AgNPs: A review. Environ. Adv. 2021, 6, 100128. [Google Scholar] [CrossRef]
- Yang, J.; Cao, W.; Rui, Y. Interactions between nanoparticles and plants: Phytotoxicity and defense mechanisms. J. Plant Interact. 2017, 12, 158–169. [Google Scholar] [CrossRef]
- Younis, S.A.; Kim, K.-H.; Shaheen, S.M.; Antoniadis, V.; Tsang, Y.F.; Rinklebe, J.; Deep, A.; Brown, R.J. Advancements of nanotechnologies in crop promotion and soil fertility: Benefits, life cycle assessment, and legislation policies. Renew. Sust. Energ. Rev. 2021, 152, 111686. [Google Scholar]
- Wang, J.; Li, M.; Feng, J.; Yan, X.; Chen, H.; Han, R. Effects of TiO2-NPs pretreatment on UV-B stress tolerance in Arabidopsis thaliana. Chemosphere 2021, 281, 130809. [Google Scholar] [CrossRef]
- Yang, F.; Hong, F.; You, W.; Liu, C.; Gao, F.; Wu, C.; Yang, P.J. Influence of nano-anatase TiO2 on the nitrogen metabolism of growing spinach. Bio. Trace Element Res. 2006, 110, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Tighe-Neira, R.; Reyes-Díaz, M.; Nunes-Nesi, A.; Recio, G.; Carmona, E.; Corgne, A.; Rengel, Z.; Inostroza-Blancheteau, C. Titanium dioxide nanoparticles provoke transient increase in photosynthetic performance and differential response in antioxidant system in Raphanus sativus L. Sci. Hortic. 2020, 269, 109418. [Google Scholar] [CrossRef]
- Asli, S.; Neumann, P.M. Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ. 2009, 32, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Das, A.K.; Gantait, S.; Kumar, V.; Gurel, E. Applications of carbon nanomaterials in the plant system: A perspective view on the pros and cons. Sci. Total Environ. 2019, 667, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Faraz, M.; Pandey, R.; Shakir, M.; Fatma, T.; Varma, A.; Barman, I.; Prasad, R. Facile algae-derived route to biogenic silver nanoparticles: Synthesis, antibacterial, and photocatalytic properties. Langmuir 2015, 31, 11605–11612. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Pandey, R.; Barman, I.; Prasad, R.J. Leveraging the attributes of Mucor hiemalis-derived silver nanoparticles for a synergistic broad-spectrum antimicrobial platform. Front. Microbiol. 2016, 7, 1984. [Google Scholar] [CrossRef]
- Prasad, R.; Pandey, R.; Barman, I. Nanobiotechnology. Engineering tailored nanoparticles with microbes: Quo vadis? Wiley Interdiscip. Rev. Nanomed. Nanobiotech. 2016, 8, 316–330. [Google Scholar] [CrossRef] [PubMed]
- Amin, H.H.; Elsayed, A.B.; Maswada, H.F.; Elsheery, N.I. Enhancing Sugar Beet Plant Health with Zinc Nanoparticles: A Sustainable Solution for Disease Management. J. Soil Plant Environ. 2023, 2, 1–20. [Google Scholar] [CrossRef]
- Kumari, M.; Mukherjee, A.; Chandrasekaran, N. Genotoxicity of silver nanoparticles in Allium cepa. Sci. Total Environ. 2009, 407, 5243–5246. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Reppert, J.; Hu, Q.; Hudson, J.S.; Reid, M.L.; Ratnikova, T.A.; Rao, A.M.; Luo, H.; Ke, P.C. Uptake, translocation, and transmission of carbon nanomaterials in rice plants. Small 2009, 5, 1128–1132. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. technology. Root uptake and phytotoxicity of ZnO nanoparticles. Environ. Sci. Tech. 2008, 42, 5580–5585. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Li, N.; Zhang, S.; Han, Y.; Zhang, Q. Integration of proteome and metabolome profiling to reveal heat stress response and tolerance mechanisms of Serratia sp. AXJ-M for the bioremediation of papermaking black liquor. J. Hazard. Mater. 2023, 450, 131092. [Google Scholar]
- Zhang, Q.; Cheng, Z.; Wang, Y.; Fu, L. Dietary protein-phenolic interactions: Characterization, biochemical-physiological consequences, and potential food applications. Crit. Rev. Food Sci. Nutr. 2021, 61, 3589–3615. [Google Scholar] [CrossRef]
- Hossain, Z.; Yasmeen, F.; Komatsu, S. Nanoparticles: Synthesis, morphophysiological effects, and proteomic responses of crop plants. Int. J. Mol. Sci. 2020, 21, 3056. [Google Scholar] [PubMed]
- Cheng, M.; Cui, Y.; Yan, X.; Zhang, R.; Wang, J.; Wang, X. Effect of dual-modified cassava starches on intelligent packaging films containing red cabbage extracts. Food Hydrocoll. 2022, 124, 107225. [Google Scholar] [CrossRef]
- Jiao, Y.; Lan, S.; Ma, D. Ultra-stable and multistimuli-responsive nanoparticles coated with zwitterionic pillar [n] arene for enhanced cellular uptake. Chin. Chem. Lett. 2021, 32, 1025–1028. [Google Scholar] [CrossRef]
- Zhou, L.; Yan, X.; Pei, X.; Du, J.; Ma, R.; Qian, J. The role of NiFe2O4 nanoparticle in the anaerobic digestion (AD) of waste activated sludge (WAS). Chin. Chem. Lett. 2022, 33, 428–433. [Google Scholar] [CrossRef]
- Fu, H.; Guo, Y.; Yu, J.; Shen, Z.; Zhao, J.; Xie, Y.; Ling, Y.; Ouyang, S.; Li, S.; Zhang, W. Tuning the shell thickness of core-shell α-Fe2O3@ SiO2 nanoparticles to promote microwave absorption. Chin. Chem. Lett. 2022, 33, 957–962. [Google Scholar]
- Wang, W.; Liu, H.; Huang, Z.; Fu, F.; Wang, W.; Wu, L.; Huang, Y.; Wu, C.; Pan, X. The effect of organic ligand modification on protein corona formation of nanoscale metal organic frameworks. Chin. Chem. Lett. 2022, 33, 4185–4190. [Google Scholar] [CrossRef]
- Zou, G.; Zhang, K.; Yang, W.; Liu, C.; Fang, Z.; Zhou, X. 5-Formyluracil targeted biochemical reactions with proteins inhibit DNA replication, induce mutations and interference gene expression in living cells. Chin. Chem. Lett. 2021, 32, 3252–3256. [Google Scholar] [CrossRef]
- Zheng, L.; Cao, W.; Dou, B.; Zeng, X.; Cao, M.; Wang, J.; Li, X. Ac36deoGlcNAz could selectively label O-GlcNAc modified proteins with minimal S-glyco-modification. Chin. Chem. Lett. 2023, 34, 107598. [Google Scholar] [CrossRef]
- Li, Y.; Xia, C.; Zhao, H.; Xie, Y.; Zhang, Y.; Zhang, W.; Yu, Y.; Wang, J.; Qin, W. A new photolabeling probe for efficient enrichment and deep profiling of cell surface membrane proteome by mass spectrometry. Chin. Chem. Lett. 2023, 34, 107377. [Google Scholar] [CrossRef]
- Shen, C.X.; Zhang, Q.F.; Li, J.; Bi, F.C.; Yao, N. Induction of programmed cell death in Arabidopsis and rice by single-wall carbon nanotubes. Am. J. Bot. 2010, 97, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollution 2007, 150, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Mahendra, S.; Zodrow, K.; Li, D.; Tsai, Y.C.; Braam, J.; Alvarez, P.J. Developmental phytotoxicity of metal oxide nanoparticles to Arabidopsis thaliana. Environ. Toxicolog. Chem. 2010, 29, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Mingyu, S.; Xiao, W.; Chao, L.; Chunxiang, Q.; Liang, C.; Hao, H.; Xiaoqing, L.; Fashui, H. Antioxidant stress is promoted by nano-anatase in spinach chloroplasts under UV-B radiation. Biolog. Trace Element Res. 2008, 121, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Avellan, A.; Yun, J.; Zhang, Y.; Spielman-Sun, E.; Unrine, J.M.; Thieme, J.; Li, J.; Lombi, E.; Bland, G.; Lowry, G.V. Nanoparticle size and coating chemistry control foliar uptake pathways, translocation, and leaf-to-rhizosphere transport in wheat. ACS Publ. 2019, 13, 5291–5305. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Michel, H.A.; Larue, C.; Del Real, A.E.P.; Cotte, M.; Sarret, G. Practical review on the use of synchrotron based micro-and nano-X-ray fluorescence mapping and X-ray absorption spectroscopy to investigate the interactions between plants and engineered nanomaterials. Plant Physiol. Biochem. 2017, 110, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; White, J.C.; Dhankher, O.P.; Xing, B. Metal-based nanotoxicity and detoxification pathways in higher plants. Environ. Sci. Tech. 2015, 49, 7109–7122. [Google Scholar] [CrossRef] [PubMed]
- Sanzari, I.; Leone, A.; Ambrosone, A.J.F.i.B. Biotechnology. Nanotechnology in plant science: To make a long story short. Front. Bioeng. Biotechnol. 2019, 7, 120. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, Y.; Gilbertson, L. Opportunities to advance sustainable design of nano-enabled agriculture identified through a literature review. Environ. Sci. Nano 2018, 5, 11–26. [Google Scholar]
- Quanbeck, S.M.; Brachova, L.; Campbell, A.A.; Guan, X.; Perera, A.; He, K.; Rhee, S.Y.; Bais, P.; Dickerson, J.A.; Dixon, P. Metabolomics as a hypothesis-generating functional genomics tool for the annotation of Arabidopsis thaliana genes of “unknown function”. Front. Plant Sci. 2012, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Pagano, L.; Wohlschlegel, J.A.; Villani, M.; Zappettini, A.; White, J.C.; Keller, A.A. Proteomic, gene and metabolite characterization reveal the uptake and toxicity mechanisms of cadmium sulfide quantum dots in soybean plants. Environ. Sci.: Nano 2019, 6, 3010–3026. [Google Scholar] [CrossRef]
- Ruotolo, R.; Maestri, E.; Pagano, L.; Marmiroli, M.; White, J.C.; Marmiroli, N. Plant response to metal-containing engineered nanomaterials: An omics-based perspective. Environ. Sci. Technol. 2018, 52, 2451–2467. [Google Scholar] [CrossRef] [PubMed]
- Elbasiouny, H.; Elbehiry, F.; El-Ramady, H. Toxic effects of nanoparticles under combined stress on plants. In Toxicity of Nanoparticles in Plants; Elsevier: Amsterdam, The Netherlands, 2022; pp. 109–129. [Google Scholar]
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive oxygen species-sources, functions, oxidative damage. Polski merkuriusz lekarski: Organ Polskiego Towarzystwa Lekarskiego 2020, 48, 124–127. [Google Scholar] [PubMed]
- Auffan, M.; Achouak, W.; Rose, J.; Roncato, M.-A.; Chanéac, C.; Waite, D.T.; Masion, A.; Woicik, J.C.; Wiesner, M.R.; Bottero, J.Y. Relation between the redox state of iron-based nanoparticles and their cytotoxicity toward Escherichia coli. Environ. Sci. Tech. 2008, 42, 6730–6735. [Google Scholar] [CrossRef]
- Bano, A.; Gupta, A.; Rai, S.; Fatima, T.; Sharma, S.; Pathak, N. Mechanistic role of reactive oxygen species and its regulation via the antioxidant system under environmental stress. In Plant Stress Physiology—Perspectives in Agriculture; Mirza, H., Nahar, K., Eds.; Intech Open: London, UK, 2021; pp. 1–18. [Google Scholar] [CrossRef]
- Aqeel, U.; Aftab, T.; Khan, M.M.A.; Naeem, M.; Khan, M.N. A comprehensive review of impacts of diverse nanoparticles on growth, development and physiological adjustments in plants under changing environment. Chemosphere 2022, 291, 132672. [Google Scholar] [PubMed]
- Dev, A.; Srivastava, A.K.; Karmakar, S. Uptake and toxicity of nanomaterials in plants. Nanosci. Food Agric. 2017, 5, 169–204. [Google Scholar]
- Verano-Braga, T.; Miethling-Graff, R.; Wojdyla, K.; Rogowska-Wrzesinska, A.; Brewer, J.R.; Erdmann, H.; Kjeldsen, F. Insights into the cellular response triggered by silver nanoparticles using quantitative proteomics. ACS Nano 2014, 8, 2161–2175. [Google Scholar] [CrossRef]
- Dev, A.; Srivastava, A.K.; Karmakar, S. Nanomaterial toxicity for plants. Environ. Chem. Lett. 2018, 16, 85–100. [Google Scholar]
- Shukla, P.K.; Misra, P.; Kole, C. Uptake, translocation, accumulation, transformation, and generational transmission of nanoparticles in plants. In Plant Nanotechnology; Kumar, C., Khodakovskaya, D.M., Eds.; Springer: Cham, Switzerland, 2016; pp. 183–218. [Google Scholar]
- Ali, S.; Mehmood, A.; Khan, N. Uptake, translocation, and consequences of nanomaterials on plant growth and stress adaptation. J. Nanomater. 2021, 2021, 6677616. [Google Scholar]
- Kang, M.E.; Weng, Y.; Liu, Y.; Wang, H.; Ye, L.; Gu, Y.; Bai, X. A Review on the Toxicity Mechanisms and Potential Risks of Engineered Nanoparticles to Plants. Rev. Environ. Contam. Toxicol. 2023, 261, 5. [Google Scholar]
- Eichert, T.; Kurtz, A.; Steiner, U.; Goldbach, H.E. Size exclusion limits and lateral heterogeneity of the stomatal foliar uptake pathway for aqueous solutes and water-suspended nanoparticles. Physiol. Plant. 2008, 134, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Birbaum, K.; Brogioli, R.; Schellenberg, M.; Martinoia, E.; Stark, W.J.; Günther, D.; Limbach, L.K. No evidence for cerium dioxide nanoparticle translocation in maize plants. Environ. Sci. Technol. 2010, 44, 8718–8723. [Google Scholar] [CrossRef] [PubMed]
- Kurepa, J.; Paunesku, T.; Vogt, S.; Arora, H.; Rabatic, B.M.; Lu, J.; Wanzer, M.B.; Woloschak, G.E.; Smalle, J.A. Uptake and distribution of ultrasmall anatase TiO2 Alizarin red S nanoconjugates in Arabidopsis thaliana. Nano Lett. 2010, 10, 2296–2302. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-N.; Tarafdar, J.C.; Biswas, P. Nanoparticle synthesis and delivery by an aerosol route for watermelon plant foliar uptake. J. Nanoparticle Res. 2013, 15, 1417. [Google Scholar] [CrossRef]
- Taran, N.; Batsmanova, L.; Konotop, Y.; Okanenko, A. Redistribution of elements of metals in plant tissues under treatment by non-ionic colloidal solution of biogenic metal nanoparticles. Nanoscale Res. Lett. 2014, 9, 354. [Google Scholar] [CrossRef] [PubMed]
- Larue, C.; Castillo-Michel, H.; Sobanska, S.; Cécillon, L.; Bureau, S.; Barthès, V.; Ouerdane, L.; Carrière, M.; Sarret, G. Foliar exposure of the crop Lactuca sativa to silver nanoparticles: Evidence for internalization and changes in Ag speciation. J. Hazard. Mater. 2014, 264, 98–106. [Google Scholar] [CrossRef]
- Zheng, L.; Hong, F.; Lu, S.; Liu, C. Effect of nano-TiO2 on strength of naturally aged seeds and growth of spinach. Biolog. Trace Element Res. 2005, 104, 83–91. [Google Scholar] [CrossRef]
- Racuciu, M.; Creanga, D. Cytogenetical changes induced by β-cyclodextrin coated nanoparticles in plant seeds. Roman J. Phys. 2009, 54, 2. [Google Scholar]
- Stampoulis, D.; Sinha, S.K.; White, J.C. technology. Assay-dependent phytotoxicity of nanoparticles to plants. Environ. Sci. Tech. 2009, 43, 9473–9479. [Google Scholar] [CrossRef]
- Mousavi Kouhi, S.M.; Lahouti, M.; Ganjeali, A.; Entezari, M.H. Comparative phytotoxicity of ZnO nanoparticles, ZnO microparticles, and Zn2+ on rapeseed (Brassica napus L.): Investigating a wide range of concentrations. Toxicol. Environ. Chem. 2014, 96, 861–868. [Google Scholar] [CrossRef]
- Pokhrel, L.R.; Dubey, B. Evaluation of developmental responses of two crop plants exposed to silver and zinc oxide nanoparticles. Sci. Total Environ. 2013, 452, 321–332. [Google Scholar] [CrossRef] [PubMed]
- De La Rosa, G.; Lopez-Moreno, M.L.; Hernandez-Viezcas, J.A.; Montes, M.O.; Peralta-Videa, J.; Gardea-Torresdey, J. Toxicity and biotransformation of ZnO nanoparticles in the desert plants Prosopis juliflora-velutina, Salsola tragus and Parkinsonia florida. Int. J. Nanotechnol. 2011, 8, 492. [Google Scholar] [CrossRef]
- Rico, C.M.; Majumdar, S.; Duarte-Gardea, M.; Peralta-Videa, J.R.; Gardea-Torresdey, J. Interaction of nanoparticles with edible plants and their possible implications in the food chain. J. Agric. Food Chem. 2011, 59, 3485–3498. [Google Scholar] [CrossRef]
- Zhu, H.; Han, J.; Xiao, J.Q.; Jin, Y. Uptake, translocation, and accumulation of manufactured iron oxide nanoparticles by pumpkin plants. J. Environ. Monit. 2008, 10, 713–717. [Google Scholar] [CrossRef]
- Du, W.; Sun, Y.; Ji, R.; Zhu, J.; Wu, J.; Guo, H. TiO2 and ZnO nanoparticles negatively affect wheat growth and soil enzyme activities in agricultural soil. J. Environ. Monit. 2011, 13, 822–828. [Google Scholar] [CrossRef]
- Zhao, L.; Peng, B.; Hernandez-Viezcas, J.A.; Rico, C.; Sun, Y.; Peralta-Videa, J.R.; Tang, X.; Niu, G.; Jin, L.; Varela-Ramirez, A. Stress response and tolerance of Zea mays to CeO2 nanoparticles: Cross talk among H2O2, heat shock protein, and lipid peroxidation. ACS Nano 2012, 6, 9615–9622. [Google Scholar] [CrossRef]
- Zhao, L.; Peralta-Videa, J.R.; Varela-Ramirez, A.; Castillo-Michel, H.; Li, C.; Zhang, J.; Aguilera, R.J.; Keller, A.A.; Gardea-Torresdey, J.L. Effect of surface coating and organic matter on the uptake of CeO2 NPs by corn plants grown in soil: Insight into the uptake mechanism. J. Hazard. Mater. 2012, 225, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Peralta-Videa, J.R.; Peng, B.; Bandyopadhyay, S.; Corral-Diaz, B.; Osuna-Avila, P.; Montes, M.O.; Keller, A.A.; Gardea-Torresdey, J.L. Alginate modifies the physiological impact of CeO2 nanoparticles in corn seedlings cultivated in soil. J. Environ. Sci. 2014, 26, 382–389. [Google Scholar] [CrossRef]
- Meriga, B.; Reddy, B.K.; Rao, K.R.; Reddy, L.A.; Kishor, P.K. Aluminium-induced production of oxygen radicals, lipid peroxidation and DNA damage in seedlings of rice (Oryza sativa). J. Plant Physiol. 2004, 161, 63–68. [Google Scholar] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lopez-Delgado, H.; Dat, J.F.; Scott, I.M. Hydrogen peroxide-and glutathione-associated mechanisms of acclimatory stress tolerance and signalling. Physiol. Plant. 1997, 100, 241–254. [Google Scholar] [CrossRef]
- Freinbichler, W.; Colivicchi, M.A.; Stefanini, C.; Bianchi, L.; Ballini, C.; Misini, B.; Weinberger, P.; Linert, W.; Varešlija, D.; Tipton, K.F. Highly reactive oxygen species: Detection, formation, and possible functions. Cell. Mol. Life Sci. 2011, 68, 2067–2079. [Google Scholar] [PubMed]
- Halliwell, B.; Gutteridge, J.M. Free Radicals in Biology and Medicine; Oxford University Press: New York, NY, USA, 2015. [Google Scholar]
- Tarasenko, V.; Garnik, E.Y.; Shmakov, V.; Konstantinov, Y.M. Modified alternative oxidase expression results in different reactive oxygen species contents in Arabidopsis cell culture but not in whole plants. Biol. Plant. 2012, 56, 635–640. [Google Scholar] [CrossRef]
- Oukarroum, A.; Barhoumi, L.; Pirastru, L.; Dewez, D. Silver nanoparticle toxicity effect on growth and cellular viability of the aquatic plant Lemna gibba. Environ. Toxicol. Chem. 2013, 32, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Yasur, J.; Rani, P.U. Environmental effects of nanosilver: Impact on castor seed germination, seedling growth, and plant physiology. Environ. Sci. Pollut. Res. 2013, 20, 8636–8648. [Google Scholar] [CrossRef] [PubMed]
- Faisal, M.; Saquib, Q.; Alatar, A.A.; Al-Khedhairy, A.A.; Hegazy, A.K.; Musarrat, J. Phytotoxic hazards of NiO-nanoparticles in tomato: A study on mechanism of cell death. J. Hazard. Mater. 2013, 250, 318–332. [Google Scholar] [CrossRef]
- Ma, C.; Liu, H.; Guo, H.; Musante, C.; Coskun, S.H.; Nelson, B.C.; White, J.C.; Xing, B.; Dhankher, O.P. Defense mechanisms and nutrient displacement in Arabidopsis thaliana upon exposure to CeO2 and In2O3 nanoparticles. ACS Sustain. Chem. Engin. 2016, 3, 1369–1379. [Google Scholar] [CrossRef]
- Rajeshwari, A.; Kavitha, S.; Alex, S.A.; Kumar, D.; Mukherjee, A.; Chandrasekaran, N.; Mukherjee, A. Cytotoxicity of aluminum oxide nanoparticles on Allium cepa root tip—Effects of oxidative stress generation and biouptake. Environ. Sci. Pollut. Res. 2015, 22, 11057–11066. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Singh, A.; Mehta, S.; Yadav, S.; Nagar, G.; Ghosh, R.; Roy, A.; Chakraborty, A.; Singh, I.K. How to cope with the challenges of environmental stresses in the era of global climate change: An update on ROS stave off in plants. Int. J. Mol. Sci. 2022, 23, 1995. [Google Scholar] [CrossRef]
- Wei, H.; Wang, E. Nanomaterials with enzyme-like characteristics (nanozymes): Next-generation artificial enzymes. Chem. Soc. Rev. 2013, 42, 6060–6093. [Google Scholar] [PubMed]
- Majumdar, S.; Peralta-Videa, J.R.; Bandyopadhyay, S.; Castillo-Michel, H.; Hernandez-Viezcas, J.-A.; Sahi, S.; Gardea-Torresdey, J.L. Exposure of cerium oxide nanoparticles to kidney bean shows disturbance in the plant defense mechanisms. J. Hazard. Mater. 2014, 278, 279–287. [Google Scholar] [CrossRef]
- Chen, G.; Ma, C.; Mukherjee, A.; Musante, C.; Zhang, J.; White, J.C.; Dhankher, O.P.; Xing, B. Tannic acid alleviates bulk and nanoparticle Nd2O3 toxicity in pumpkin: A physiological and molecular response. Nanotoxicology 2016, 10, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.K.; Hossain, Z. Impact of nano-CuO stress on rice (Oryza sativa L.) seedlings. Chemosphere 2013, 93, 906–915. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant. 2003, 119, 355–364. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; McLean, J.E.; Latta, D.E.; Manangón, E.; Britt, D.W.; Johnson, W.P.; Boyanov, M.I.; Anderson, A.J. CuO and ZnO nanoparticles: Phytotoxicity, metal speciation, and induction of oxidative stress in sand-grown wheat. J. Nanoparticle Res. 2012, 14, 1125. [Google Scholar] [CrossRef]
- Cui, D.; Zhang, P.; Ma, Y.; He, X.; Li, Y.; Zhang, J.; Zhao, Y.; Zhang, Z. Effect of cerium oxide nanoparticles on asparagus lettuce cultured in an agar medium. Environ. Sci. Nano J. 2014, 1, 459–465. [Google Scholar] [CrossRef]
- Zhang, P.; Ma, Y.; Zhang, Z.; He, X.; Li, Y.; Zhang, J.; Zheng, L.; Zhao, Y. Species-specific toxicity of ceria nanoparticles to Lactuca plants. Nanotoxicology 2015, 9, 1–8. [Google Scholar] [CrossRef]
- Zhang, P.; Ma, Y.; Liu, S.; Wang, G.; Zhang, J.; He, X.; Zhang, J.; Rui, Y.; Zhang, Z. Phytotoxicity, uptake and transformation of nano-CeO2 in sand cultured romaine lettuce. Environ. Pollut. 2017, 220, 1400–1408. [Google Scholar] [CrossRef]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of silver nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Tech. 2008, 42, 8959–8964. [Google Scholar] [CrossRef] [PubMed]
- Oukarroum, A.; Bras, S.; Perreault, F.; Popovic, R. Inhibitory effects of silver nanoparticles in two green algae, Chlorella vulgaris and Dunaliella tertiolecta. Ecotoxicol. Environ. Saf. 2012, 78, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Manzo, S.; Miglietta, M.L.; Rametta, G.; Buono, S.; Di Francia, G. Toxic effects of ZnO nanoparticles towards marine algae Dunaliella tertiolecta. Sci. Total Environ. 2013, 445, 371–376. [Google Scholar] [CrossRef]
- Perreault, F.; Bogdan, N.; Morin, M.; Claverie, J.; Popovic, R. Interaction of gold nanoglycodendrimers with algal cells (Chlamydomonas reinhardtii) and their effect on physiological processes. Nanotoxicology 2012, 6, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Geisler-Lee, J.; Wang, Q.; Yao, Y.; Zhang, W.; Geisler, M.; Li, K.; Huang, Y.; Chen, Y.; Kolmakov, A.; Ma, X. Phytotoxicity, accumulation and transport of silver nanoparticles by Arabidopsis thaliana. Nanotoxicology 2012, 7, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Van Hoecke, K.; De Schamphelaere, K.A.; Van der Meeren, P.; Lcucas, S.; Janssen, C.R. Ecotoxicity of silica nanoparticles to the green alga Pseudokirchneriella subcapitata: Importance of surface area. Environ. Toxicol. Chem. 2008, 27, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Geiser-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.F.; Domingos, R.F.; Hauser, C.; Hutchins, C.M.; Zerges, W.; Wilkinson, K. Transcriptome sequencing (RNA-seq) analysis of the effects of metal nanoparticle exposure on the transcriptome of Chlamydomonas reinhardtii. ASM J. 2013, 79, 4774–4785. [Google Scholar] [CrossRef]
- Landa, P.; Vankova, R.; Andrlova, J.; Hodek, J.; Marsik, P.; Storchova, H.; White, J.C.; Vanek, T. Nanoparticle-specific changes in Arabidopsis thaliana gene expression after exposure to ZnO, TiO2, and fullerene soot. J. Hazard. Mater. 2012, 241, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Kaveh, R.; Li, Y.-S.; Ranjbar, S.; Tehrani, R.; Brueck, C.L.; Van Aken, B. Changes in Arabidopsis thaliana gene expression in response to silver nanoparticles and silver ions. Environ. Sci. Tech. 2013, 47, 10637–10644. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, M.; Jana, A.; Sinha, S.; Jothiramajayam, M.; Nag, A.; Chakraborty, A.; Mukherjee, A.; Mukherjee, A.; Mutagenesis, E. Effects of ZnO nanoparticles in plants: Cytotoxicity, genotoxicity, deregulation of antioxidant defenses, and cell-cycle arrest. Genet. Toxicol. Environ. Mutagen. 2016, 807, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Atha, D.H.; Wang, H.; Petersen, E.J.; Cleveland, D.; Holbrook, R.D.; Jaruga, P.; Dizdaroglu, M.; Xing, B.; Nelson, B.C. Copper oxide nanoparticle mediated DNA damage in terrestrial plant models. Environ. Sci. Tech. 2012, 46, 1819–1827. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Mishra, R.K.; Singh, S.; Singh, S.; Vishwakarma, K.; Sharma, S.; Singh, V.P.; Singh, P.K.; Prasad, S.M.; Dubey, N.K. Nitric oxide ameliorates zinc oxide nanoparticles phytotoxicity in wheat seedlings: Implication of the ascorbate–glutathione cycle. Front. Plant Sci. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.; Azevedo, R.; Fernandes, P.; Santos, C. Cr (VI) induces DNA damage, cell cycle arrest and polyploidization: A flow cytometric and comet assay study in Pisum sativum. Chem. Res. Toxicol. 2011, 24, 1040–1047. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Lien, H.-L.; Koel, B.E.; Zhang, W. Impacts. Iron nanoparticles for environmental clean-up: Recent developments and future outlook. Environ. Sci. Process. Impacts 2013, 15, 63–77. [Google Scholar] [CrossRef]
- Khodakovskaya, M.; Dervishi, E.; Mahmood, M.; Xu, Y.; Li, Z.; Watanabe, F.; Biris, A.S. Carbon nanotubes are able to penetrate plant seed coat and dramatically affect seed germination and plant growth. ACS Nano 2009, 3, 3221–3227. [Google Scholar] [CrossRef]
- Vannini, C.; Domingo, G.; Onelli, E.; Prinsi, B.; Marsoni, M.; Espen, L.; Bracale, M. Morphological and proteomic responses of Eruca sativa exposed to silver nanoparticles or silver nitrate. PLoS ONE 2013, 8, e68752. [Google Scholar] [CrossRef]
- Kumar, V.; Guleria, P.; Kumar, V.; Yadav, S.K. Gold nanoparticle exposure induces growth and yield enhancement in Arabidopsis thaliana. Sci. Total Environ. 2013, 461, 462–468. [Google Scholar] [CrossRef]
- Burklew, C.E.; Ashlock, J.; Winfrey, W.B.; Zhang, B. Effects of aluminum oxide nanoparticles on the growth, development, and microRNA expression of tobacco (Nicotiana tabacum). PLoS ONE 2012, 7, e34783. [Google Scholar] [CrossRef] [PubMed]
- Poma, A.; Di Giorgio, M.L. Toxicogenomics to improve comprehension of the mechanisms underlying responses of in vitro and in vivo systems to nanomaterials: A review. Curr. Genom. 2008, 9, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Poynton, H.C.; Vulpe, C.D. Ecotoxicogenomics: Emerging technologies for emerging contaminants. J. Am. Water Resour. Assoc. 2009, 45, 83–96. [Google Scholar] [CrossRef]
- Merrick, B.A.; Bruno, M.E. Genomic and proteomic profiling for biomarkers and signature profiles of toxicity. Curr. Opin. Molecul. Ther. 2004, 6, 600–607. [Google Scholar]
- Thomas, C.R.; George, S.; Horst, A.M.; Ji, Z.; Miller, R.J.; Peralta-Videa, J.R.; Xia, T.; Pokhrel, S.; Mädler, L.; Gardea-Torresdey, J.L.; et al. Nanomaterials in the environment: From materials to high-throughput screening to organisms. ACS Nano 2011, 5, 13–20. [Google Scholar] [CrossRef]
- Bashri, G.; Parihar, P.; Singh, R.; Patel, A.; Prasad, S.M. Plant and nanoparticle interface at the molecular level: An integrated overview. In Nanomaterials in Plants, Algae, and Microorganisms: Concepts and Controversies; Tripathi, K.D., Ahmad, P., Dubey, N.K., Eds.; Elsevier: London, UK, 2018; Volume 1, pp. 325–344. [Google Scholar]
- García-Sánchez, S.; Bernales, I.; Cristobal, S. Early response to nanoparticles in the Arabidopsis transcriptome compromises plant defence and root-hair development through salicylic acid signalling. BMC Genom. 2015, 16, 347. [Google Scholar] [CrossRef] [PubMed]
- Tumburu, L.; Andersen, C.P.; Rygiewicz, P.T.; Reichman, J.R. Phenotypic and genomic responses to titanium dioxide and cerium oxide nanoparticles in Arabidopsis germinants. Environ. Toxicol. 2015, 34, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Frazier, T.P.; Burklew, C.E.; Zhang, B. Titanium dioxide nanoparticles affect the growth and microRNA expression of tobacco (Nicotiana tabacum). Funct. Integr. Genom. 2014, 14, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Landa, P.; Prerostova, S.; Petrova, S.; Knirsch, V.; Vankova, R.; Vanek, T. The transcriptomic response of Arabidopsis thaliana to zinc oxide: A comparison of the impact of nanoparticle, bulk, and ionic zinc. Environ. Sci. Technol. 2015, 49, 14537–14545. [Google Scholar] [CrossRef]
- Wang, Q.; Zhao, S.; Zhao, Y.; Rui, Q.; Wang, D. Toxicity and translocation of graphene oxide in Arabidopsis plants under stress conditions. RSC Adv. 2014, 4, 60891–60901. [Google Scholar] [CrossRef]
- Kohan-Baghkheirati, E.; Geisler-Lee, J. Gene expression, protein function and pathways of Arabidopsis thaliana responding to silver nanoparticles in comparison to silver ions, cold, salt, drought, and heat. Nanotoxicology 2015, 5, 436–467. [Google Scholar] [CrossRef]
- Gopalakrishnan Nair, P.M.; Chung, I. Cell cycle and mismatch repair genes as potential biomarkers in Arabidopsis thaliana seedlings exposed to silver nanoparticles. Bull. Environ. Contam. 2014, 92, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.; Chung, I. Assessment of silver nanoparticle-induced physiological and molecular changes in Arabidopsis thaliana. Environ. Sci. Pollut. Res. 2014, 21, 8858–8869. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; McLean, J.E.; Martineau, N.; Britt, D.W.; Haverkamp, R.; Anderson, A. Silver nanoparticles disrupt wheat (Triticum aestivum L.) growth in a sand matrix. Environ. Sci. Tech. 2013, 47, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Patlolla, A.K.; Berry, A.; May, L.; Tchounwou, P.B. Genotoxicity of silver nanoparticles in Vicia faba: A pilot study on the environmental monitoring of nanoparticles. Int. J. Environ. Res. Public Health 2012, 9, 1649–1662. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.; Chung, I. Study on the correlation between copper oxide nanoparticles induced growth suppression and enhanced lignification in Indian mustard (Brassica juncea L.). Ecotoxicol. Environ. Safety 2015, 113, 302–313. [Google Scholar] [CrossRef]
- Chu, H.; Kim, H.-J.; Kim, J.S.; Kim, M.-S.; Yoon, B.-D.; Park, H.-J.; Kim, C.Y. A nanosized Ag–silica hybrid complex prepared by γ-irradiation activates the defense response in Arabidopsis. Radiation Phys. Chem. 2012, 81, 180–184. [Google Scholar] [CrossRef]
- Chen, C.; Unrine, J.M.; Judy, J.D.; Lewis, R.W.; Guo, J.; McNear, D.H.; Tsyusko, O.V. Toxicogenomic responses of the model legume Medicago truncatula to aged biosolids containing a mixture of nanomaterials (TiO2, Ag, and ZnO) from a pilot wastewater treatment plant. Environ. Sci. Tech. 2015, 49, 8759–8768. [Google Scholar] [CrossRef] [PubMed]
- Lahiani, M.H.; Dervishi, E.; Chen, J.; Nima, Z.; Gaume, A.; Biris, A.S.; Khodakovskaya, M.V. Impact of carbon nanotube exposure to seeds of valuable crops. ACS Appl. Mater. Interfaces 2013, 5, 7965–7973. [Google Scholar] [CrossRef] [PubMed]
- Khodakovskaya, M.V.; De Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon nanotubes induce growth enhancement of tobacco cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef]
- Lahiani, M.H.; Chen, J.; Irin, F.; Puretzky, A.A.; Green, M.J.; Khodakovskaya, M.V. Interaction of carbon nanohorns with plants: Uptake and biological effects. Carbon 2015, 81, 607–619. [Google Scholar] [CrossRef]
- Yan, S.; Zhao, L.; Li, H.; Zhang, Q.; Tan, J.; Huang, M.; He, S.; Li, L. Single-walled carbon nanotubes selectively influence maize root tissue development accompanied by the change in the related gene expression. J. Hazard. Mater. 2013, 246, 110–118. [Google Scholar] [CrossRef]
- Ma, C.; Chhikara, S.; Xing, B.; Musante, C.; White, J.C.; Dhankher, O.P. Physiological and molecular response of Arabidopsis thaliana (L.) to nanoparticle cerium and indium oxide exposure. ACS Sustain. Chem. Engin. 2013, 1, 768–778. [Google Scholar] [CrossRef]
- López-Moreno, M.L.; de la Rosa, G.; Hernández-Viezcas, J.Á.; Castillo-Michel, H.; Botez, C.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J. Evidence of the differential biotransformation and genotoxicity of ZnO and CeO2 nanoparticles on soybean (Glycine max) plants. Environ. Sci. Tech. 2010, 44, 7315–7320. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Krishnamurthy, S.; Shukla, D.; Kiiskila, J.; Jain, A.; Datta, R.; Sharma, N.; Sahi, S.V. Comparative transcriptome and proteome analysis to reveal the biosynthesis of gold nanoparticles in Arabidopsis. Sci. Rep. 2016, 6, 21733. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.F.; Rylott, E.L.; Anderson, C.W.; Bruce, N.C. Investigating the toxicity, uptake, nanoparticle formation and genetic response of plants to gold. PLoS ONE 2014, 9, e93793. [Google Scholar] [CrossRef] [PubMed]
- Marmiroli, M.; Pagano, L.; Savo Sardaro, M.L.; Villani, M.; Marmiroli, N. Genome-wide approach in Arabidopsis thaliana to assess the toxicity of cadmium sulfide quantum dots. Environ. Sci. Tech. 2014, 48, 5902–5909. [Google Scholar] [CrossRef] [PubMed]
- Khodakovskaya, M.V.; de Silva, K.; Nedosekin, D.A.; Dervishi, E.; Biris, A.S.; Shashkov, E.V.; Galanzha, E.I.; Zharov, V.P. Complex genetic, photothermal, and photoacoustic analysis of nanoparticle-plant interactions. Appl. Biol. Sci. 2011, 108, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Mirzajani, F.; Askari, H.; Hamzelou, S.; Schober, Y.; Römpp, A.; Ghassempour, A.; Spengler, B. Proteomics study of silver nanoparticles toxicity on Oryza sativa L. Ecotoxicol. Environ. Saf. 2014, 108, 335–339. [Google Scholar] [CrossRef]
- Mustafa, G.; Sakata, K.; Hossain, Z.; Komatsu, S. Proteomic study on the effects of silver nanoparticles on soybean under flooding stress. J. Proteom. 2015, 122, 100–118. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, G.; Komatsu, S. Insights into the response of soybean mitochondrial proteins to various sizes of aluminum oxide nanoparticles under flooding stress. J. Proteome Res. 2016, 15, 4464–4475. [Google Scholar] [CrossRef] [PubMed]
- Jha, S.; Pudake, R.N. Molecular mechanism of plant–nanoparticle interactions. In Plant Nanotechnology Principles and Practices; Kole, C., Kumar, D.S., Khodakovskaya, M.V., Eds.; Springer: Cham, Switzerland, 2016; pp. 155–181. [Google Scholar]
- Lok, C.N.; Ho, C.M.; Chen, R.; He, Q.Y.; Yu, W.Y.; Sun, H.; Tam, P.K.; Chiu, J.F.; Che, C.M. Proteomic analysis of the mode of antibacterial action of silver nanoparticles. J. Proteome Res. 2006, 5, 916–924. [Google Scholar] [CrossRef] [PubMed]
- Sohm, B.; Immel, F.; Bauda, P.; Pagnout, C. Insight into the primary mode of action of TiO2 nanoparticles on Escherichia coli in the dark. Proteomics 2015, 15, 98–113. [Google Scholar] [CrossRef] [PubMed]
- Gambino, M.; Marzano, V.; Villa, F.; Vitali, A.; Vannini, C.; Landini, P.; Cappitelli, F. Effects of sublethal doses of silver nanoparticles on Bacillus subtilis planktonic and sessile cells. J. Appl. Microbiol. 2015, 118, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Vannini, C.; Domingo, G.; Onelli, E.; De Mattia, F.; Bruni, I.; Marsoni, M.; Bracale, M. Phytotoxic and genotoxic effects of silver nanoparticles exposure on germinating wheat seedlings. J. Plant Physiol. 2014, 171, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, A. The effects of silver nitrate and silver nanoparticles on Chlamydomonas reinhardtii: A proteomic approach. Degree Proj. Master Sci. Ecotoxicol. 2014, 9, 1–34. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).