
Impact of Collembola on the Winter Wheat Growth in Soil Infected by Soil-Borne Pathogenic Fungi


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
- Plant: wheat, wheat, and pea;
- Collembola number per test vessel: 100 individuals, 50 individuals, 0 individuals;
- The presence of the plant pathogenic fungus: present, absent.
2.2. Soil Preparation
2.3. Organisms and Incubation
2.4. Analyzes after Incubation
2.5. Data Analysis
3. Results
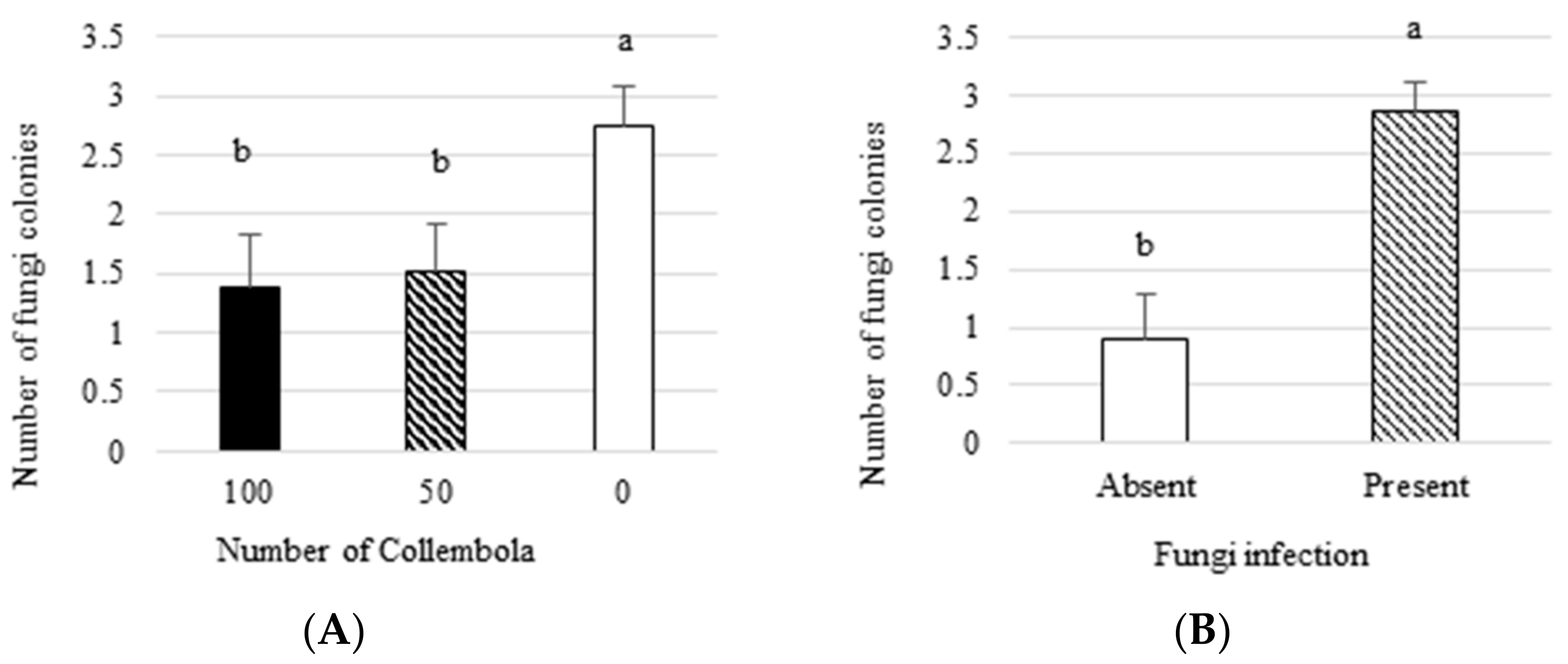
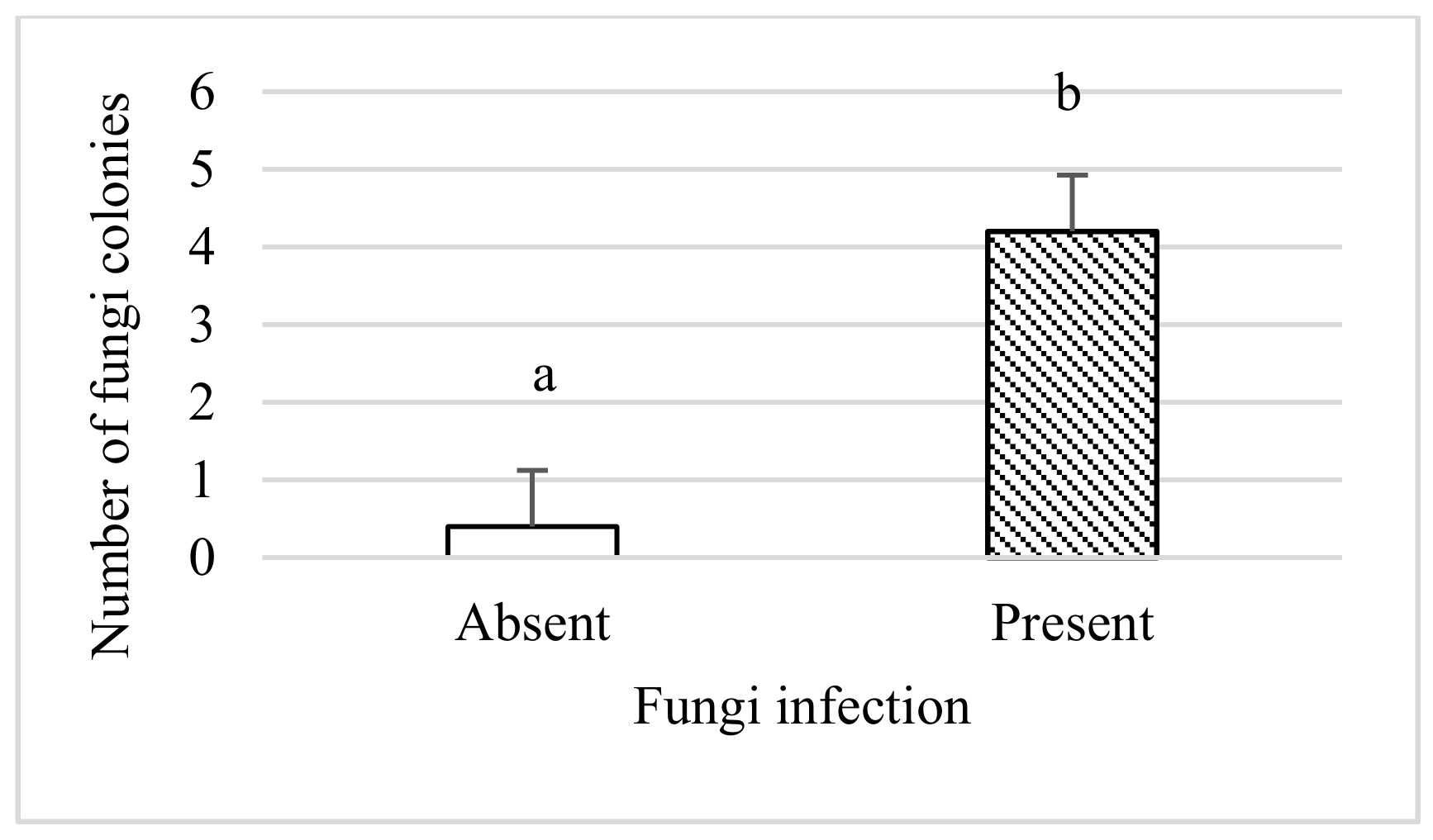
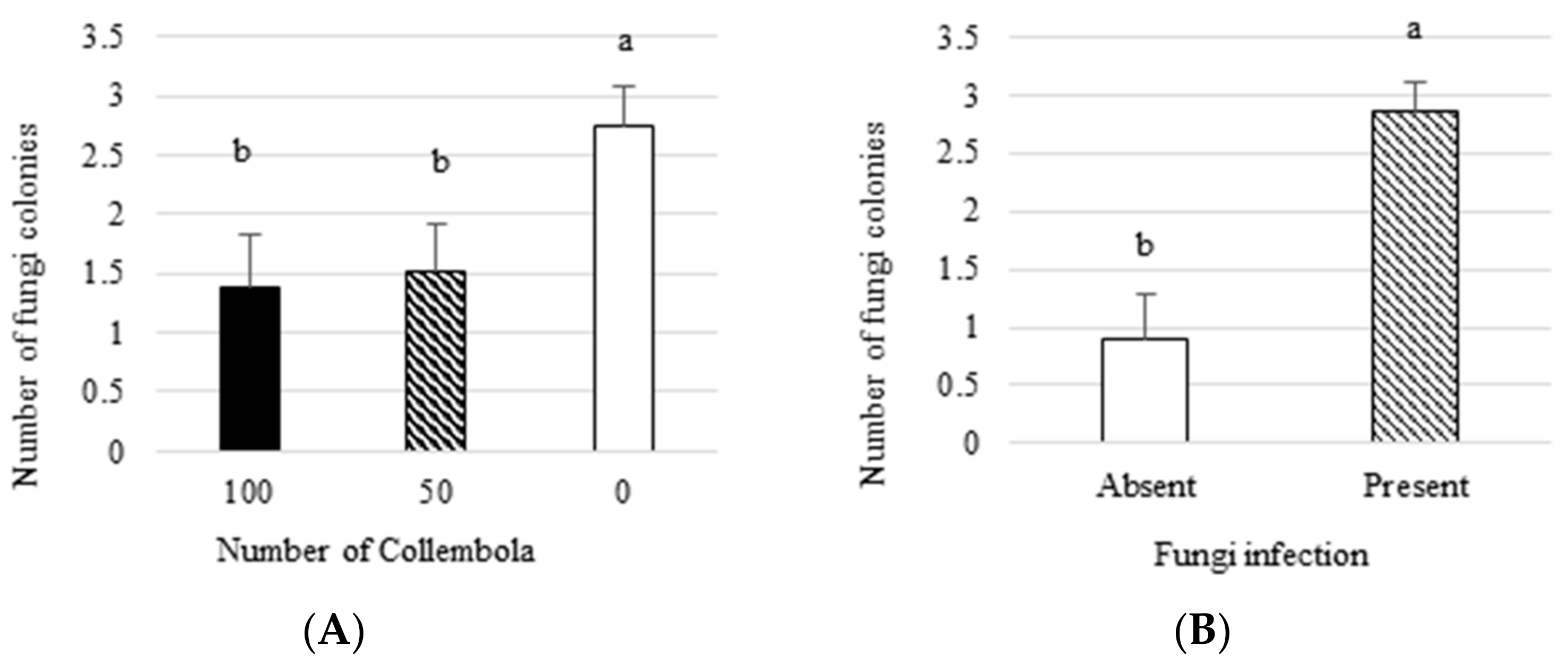
3.1. Effect on Plant Growth Parameters and Number of Fungal Colonies
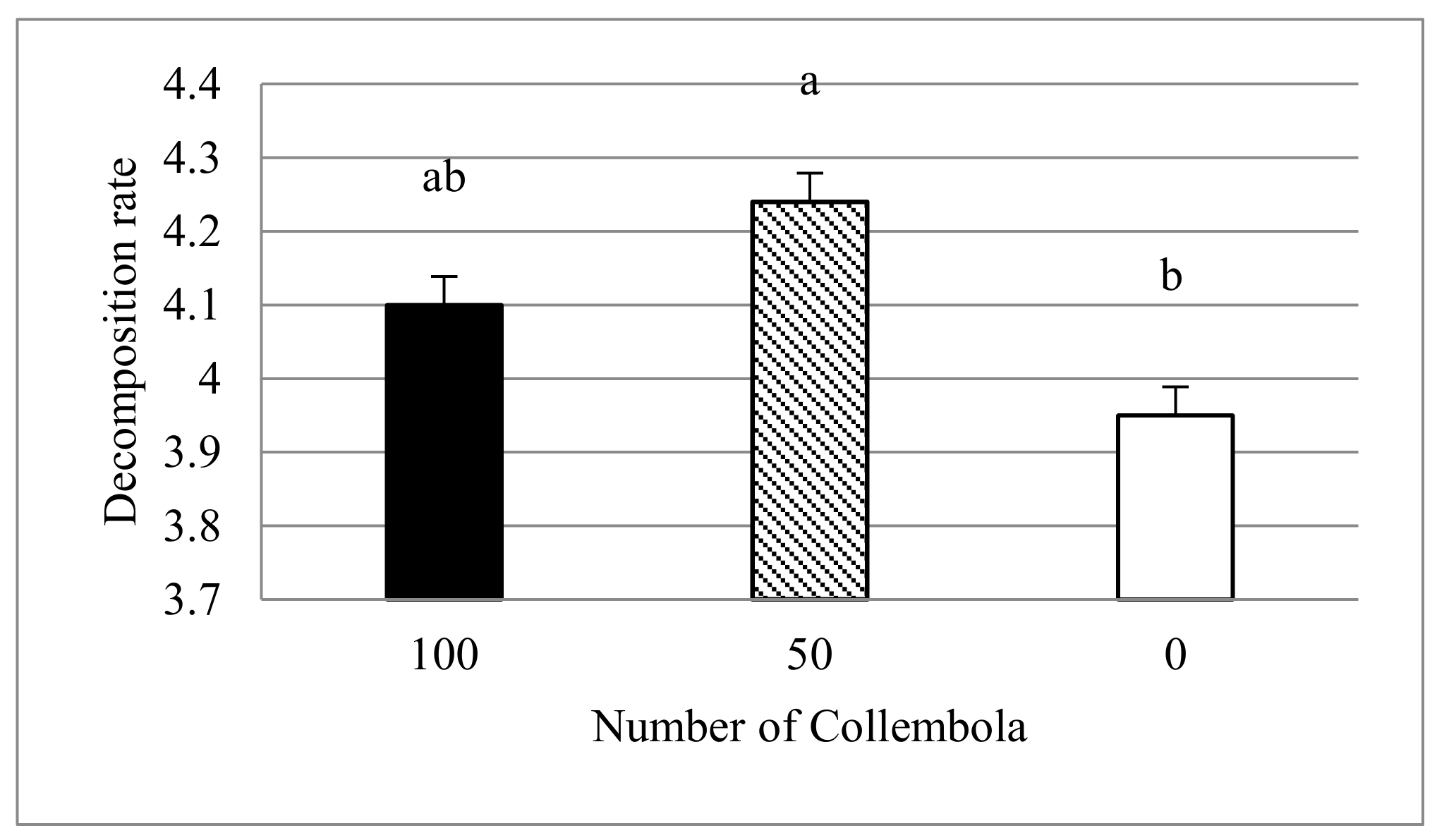
3.2. Collembolan Feeding on Fungus and Decomposition Rates
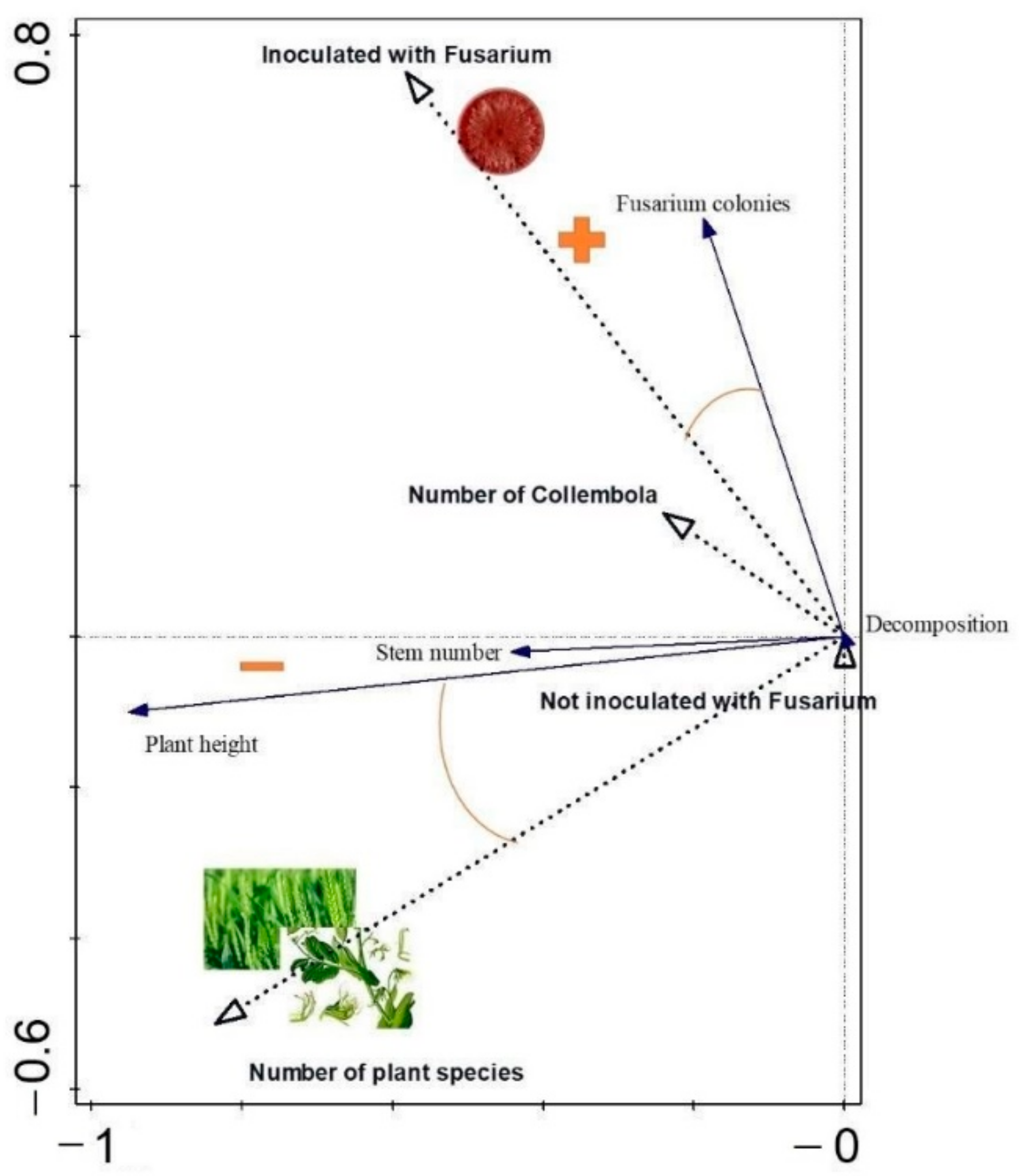
3.3. Link between the Plant Growth Parameters, Decomposition Rate, and Number of Fungus Colonies
4. Discussion
- Collembola decreased the number of fungus colonies isolated from plants by 45%.
- Fusarium culmorum was isolated from the bodies of Collembola, indicating the ability to feed on this fungus.
- The decomposition was accelerated by Collembola. However, this was not related to better plant growth.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Potapov, A.; Bellini, B.; Chown, S.; Deharveng, L.; Janssens, F.; Kováč, Ľ.; Kuznetsova, N.; Ponge, J.F.; Potapov, M.; Querner, P.; et al. Towards a global synthesis of Collembola knowledge: Challenges and potential solutions. Soil Org. 2020, 92, 161–188. [Google Scholar] [CrossRef]
- Menta, C. Soil Fauna Diversity—Function, Soil Degradation, Biological Indices, Soil Restoration. In Biodiversity Conservation and Utilization in a Diverse World; Lameed, G.A., Ed.; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Verma, D.; Paliwal, A.K. Effects of springtails community on plant-growth. Biol. Forum Int. J. 2010, 2, 70–72. [Google Scholar]
- Kuťáková, E.; Cesarz, S.; Münzbergová, Z.; Eisenhauer, N. Soil microarthropods alter the outcome of plant-soil feedback experiments. Sci. Rep. 2018, 8, 11898. [Google Scholar] [CrossRef] [PubMed]
- Filser, J. The role of Collembola in carbon and nitrogen cycling in soil. Pedobiologia 2002, 46, 234–245. [Google Scholar] [CrossRef]
- Sabatini, M.A.; Innocenti, G. Effects of Collembola on plant-pathogenic fungus interactions in simple experimental systems. Biol. Fertil. Soils 2001, 33, 62–66. [Google Scholar] [CrossRef]
- Innocenti, G.; Sabatini, M.A. Collembola and plant pathogenic, antagonistic and arbuscular mycorrhizal fungi: A review. Bull. Insect. 2018, 71, 71–76. [Google Scholar]
- Sabatini, M.A.; Innocenti, G. Functional relationships between Collembola and plant pathogenic fungi of agricultural soils. Pedobiologia 2000, 44, 467–475. [Google Scholar] [CrossRef]
- Sabatini, M.A.; Innocenti, G. Soil-borne plant pathogenic fungi in relation to some collembolan species under laboratory con-ditions. Mycol. Res. 2000, 104, 1197–1201. [Google Scholar] [CrossRef]
- Berg, M.P.; Stoffer, M.; Heuvel, H.H.V.D. Feeding guilds in Collembola based on digestive enzymes. Pedobiologia 2004, 48, 589–601. [Google Scholar] [CrossRef]
- Chahartaghi, M.; Langel, R.; Scheu, S.; Ruess, L. Feeding guilds in Collembola based on nitrogen stable isotope ratios. Soil Biol. Biochem. 2005, 37, 1718–1725. [Google Scholar] [CrossRef]
- Potapov, A.M.; Tiunov, A. Stable isotope composition of mycophagous collembolans versus mycotrophic plants: Do soil invertebrates feed on mycorrhizal fungi? Soil Biol. Biochem. 2016, 93, 115–118. [Google Scholar] [CrossRef]
- Fountain, M.T.; Hopkin, S.P. FOLSOMIA CANDIDA (COLLEMBOLA): A “Standard” Soil Arthropod. Annu. Rev. Èntomol. 2005, 50, 201–222. [Google Scholar] [CrossRef] [Green Version]
- Bärlocher, F. Leaf mass loss estimated by litter bag technique. In Methods to Study Litter Decomposition: Practical Guide; Graca, M.A.S., Bärlocher, F., Gessner, M., Eds.; Springer: Amsterdam, The Netherland, 2005; pp. 37–42. [Google Scholar]
- Rovira, P.; Rovira, R. Fitting litter decomposition datasets to mathematical curves: Towards a generalised exponential approach. Geoderma 2010, 155, 329–343. [Google Scholar] [CrossRef]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Penn State University: University Park, PY, USA, 1990. [Google Scholar]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Kampichler, C.; Bruckner, A. The role of microarthropods in terrestrial decomposition: A meta-analysis of 40 years of litterbag studies. Biol. Rev. 2009, 84, 375–389. [Google Scholar] [CrossRef]
- Innocenti, G.; Ganassi, S.; Montanari, M.; Branzanti, M.B.; Sabatini, M.A. Response of plant growth to Collembola, arbuscular mycorrhizal and plant pathogenic fungi interactions. Bull. Insect. 2009, 62, 191–195. [Google Scholar]
- Cortet, J.; Joffre, R.; Elmholt, S.; Krogh, P.H. Increasing species and trophic diversity of mesofauna affects fungal biomass, mesofauna community structure and organic matter decomposition processes. Biol. Fertil. Soils 2003, 37, 302–312. [Google Scholar] [CrossRef]
- Scheu, S.; Theenhaus, A.; Jones, T.H. Links between the detritivore and the herbivore system: Effects of earthworms and Collembola on plant growth and aphid development. Oecologia 1999, 119, 541–551. [Google Scholar] [CrossRef]
- Rich, S.M.; Watt, M. Soil conditions and cereal root system architecture: Review and considerations for linking Darwin and Weaver. J. Exp. Bot. 2013, 64, 1193–1208. [Google Scholar] [CrossRef] [Green Version]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Endlweber, K.; Scheu, S. Interactions between mycorrhizal fungi and Collembola: Effects on root structure of competing plant species. Biol. Fertil. Soils 2006, 43, 741–749. [Google Scholar] [CrossRef]
- Endlweber, K. Decomposer-Plant Interactions: Effects of Collembola on Plant Performance and Competitiveness. Ph.D. Thesis, Technical University of Darmstadt, Darmstadt, Germany, 2007. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Agricultural Soil | A Mixture Of Soil, Sand, and Peat |
|---|---|---|---|
| pH w H2O | 5.1 | 6.6 | |
| Salinity | g NaCl/dm3 | <0.2 (n) | <0.2 (n) |
| N-NO3 | mg/dm3 | <10 (5) | <10 (4) |
| P | 17 | 19 | |
| K | 30 | 14 | |
| Ca | 109 | 429 | |
| Na | <6 (4) | <6 (5) | |
| Mg | 17 | 47 | |
| Chlorides | 10 | 10 | |
| C- org | % dry mass | 0.62 | 0.69 |
| Organic matter | 1.07 | 1.19 |
| Effect | Plant Height | Number of Shoots | ||||
|---|---|---|---|---|---|---|
| DF * | F | p | Df | F | p | |
| Number of plant species | 1 | 6.12 | 0.01 | 1 | 5.47 | 0.02 |
| Number of Collembola | 2 | 0.23 | 0.80 | 2 | 0.13 | 0.88 |
| Presence of fungus | 1 | 0.01 | 0.90 | 1 | 2.48 | 0.12 |
| Effect | DF * | F | p |
|---|---|---|---|
| Number of Collembola | 2 | 4.28 | 0.02 |
| Presence of fungus | 1 | 18.49 | 0.0001 |
| Effect | DF * | F | p |
|---|---|---|---|
| Number of plant species | 2 | 0.2 | 0.82 |
| Number of Collembola | 2 | 6.44 | 0.007 |
| Presence of fungus | 1 | 0.82 | 0.37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruss, I.; Twardowski, J.; Matkowski, K.; Jurga, M. Impact of Collembola on the Winter Wheat Growth in Soil Infected by Soil-Borne Pathogenic Fungi. Agronomy 2022, 12, 1599. https://doi.org/10.3390/agronomy12071599
Gruss I, Twardowski J, Matkowski K, Jurga M. Impact of Collembola on the Winter Wheat Growth in Soil Infected by Soil-Borne Pathogenic Fungi. Agronomy. 2022; 12(7):1599. https://doi.org/10.3390/agronomy12071599
Chicago/Turabian StyleGruss, Iwona, Jacek Twardowski, Krzysztof Matkowski, and Marta Jurga. 2022. "Impact of Collembola on the Winter Wheat Growth in Soil Infected by Soil-Borne Pathogenic Fungi" Agronomy 12, no. 7: 1599. https://doi.org/10.3390/agronomy12071599
APA StyleGruss, I., Twardowski, J., Matkowski, K., & Jurga, M. (2022). Impact of Collembola on the Winter Wheat Growth in Soil Infected by Soil-Borne Pathogenic Fungi. Agronomy, 12(7), 1599. https://doi.org/10.3390/agronomy12071599