
Bayesian Non-Parametric Thermal Thresholds for Helicoverpa armigera and Their Integration into a Digital Plant Protection System

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites, Monitoring and Heat Summations
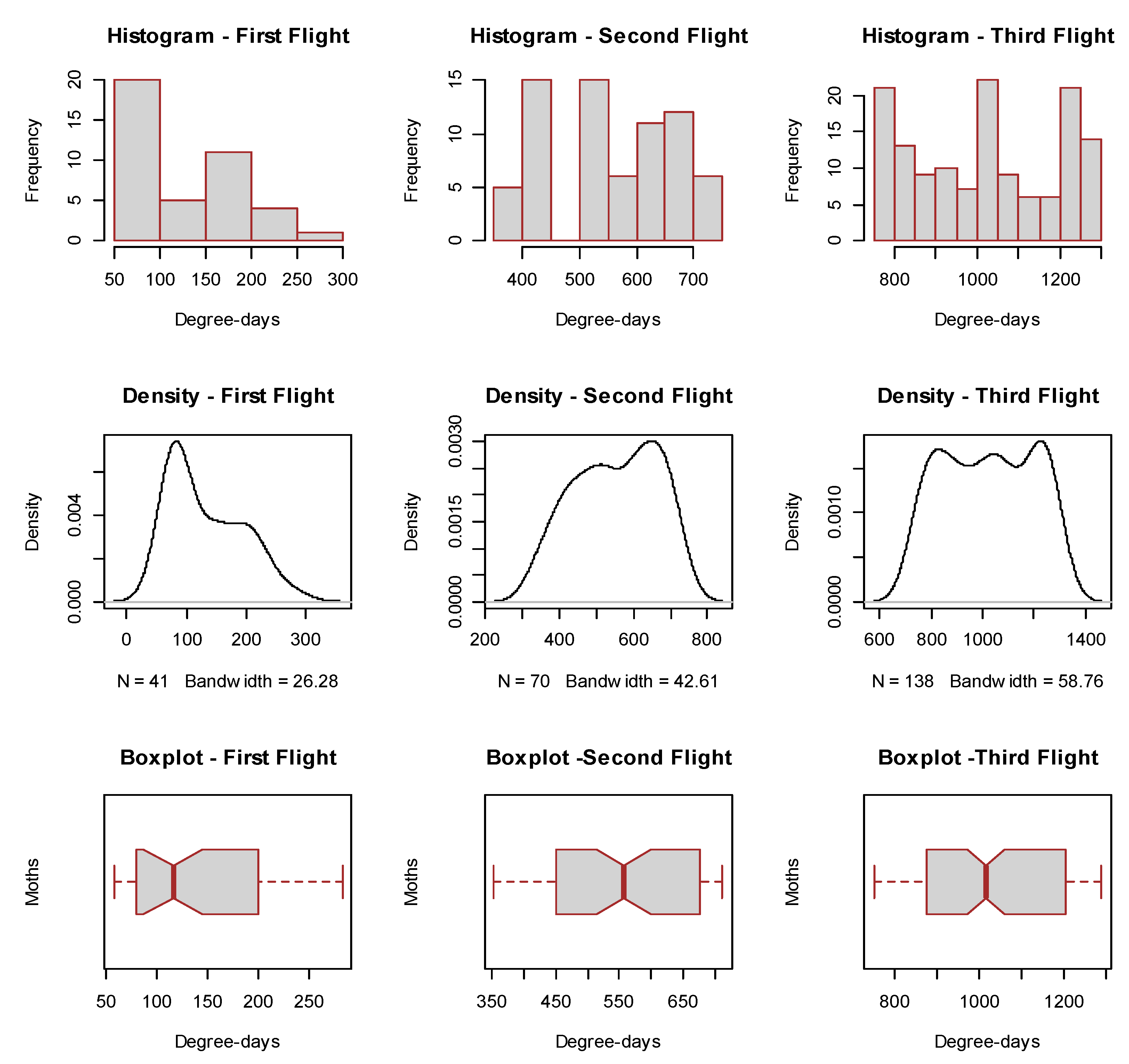
2.2. Determination of H. armigera Moth Phenology
2.3. Data Analysis
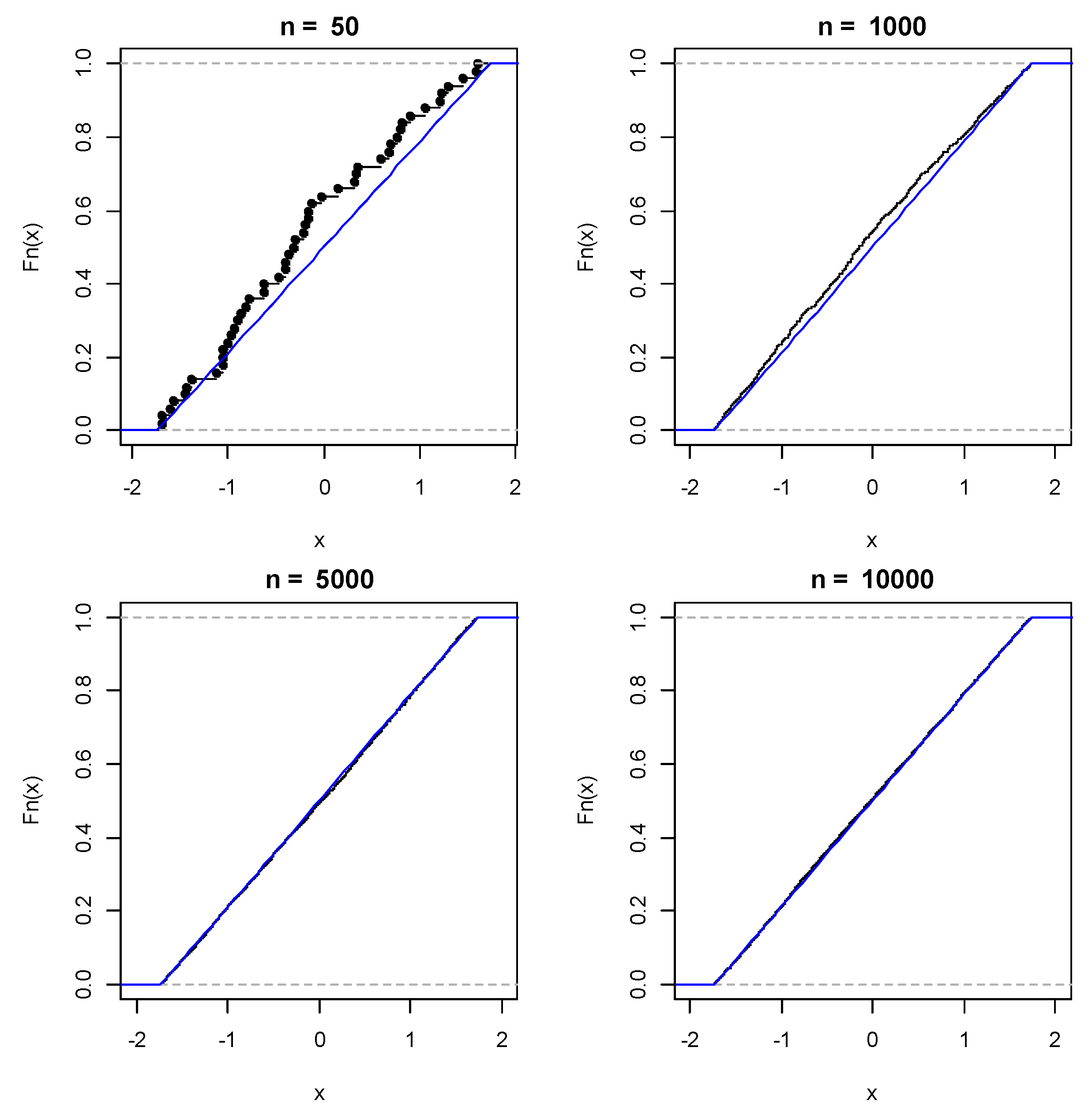
2.3.1. Non-Parametric Bootstrap with Replication
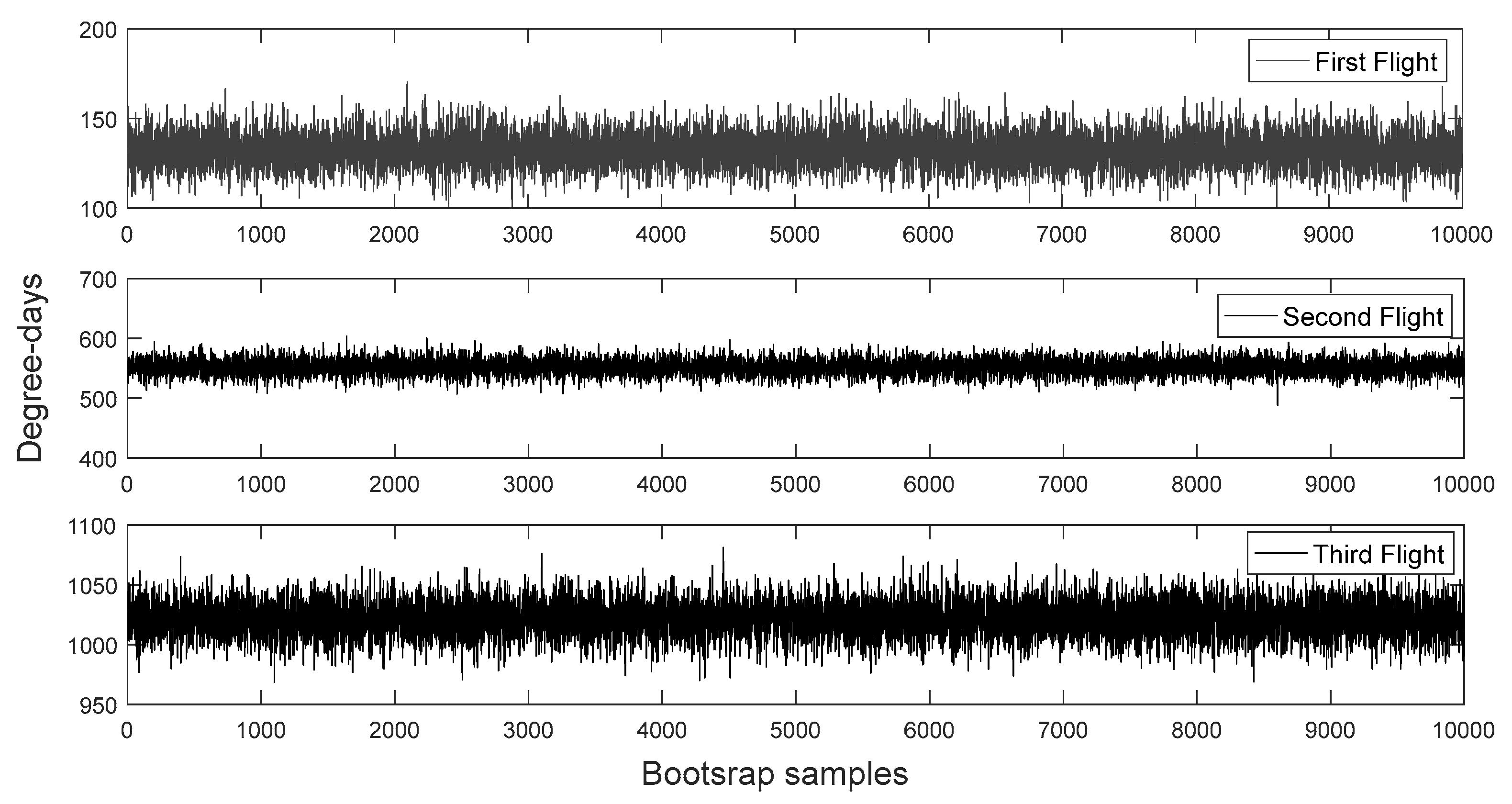
2.3.2. The Bayesian Bootstrap
2.3.3. Dirichlet Sampling to Weight the Data and Estimate the Prior
2.3.4. Confidence Intervals and Percentiles
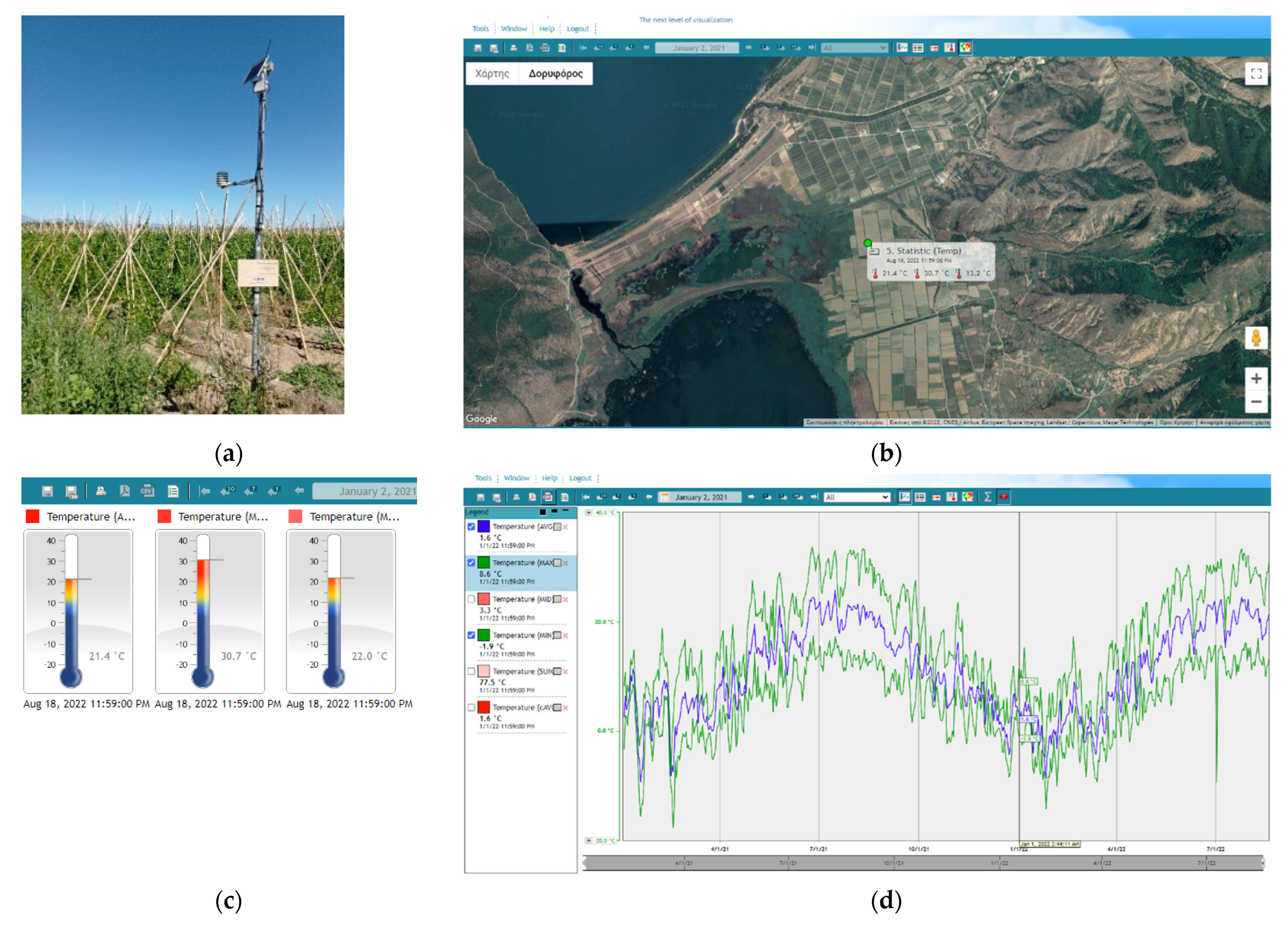
2.4. Meteorological Network, Whether Data and Software
3. Results
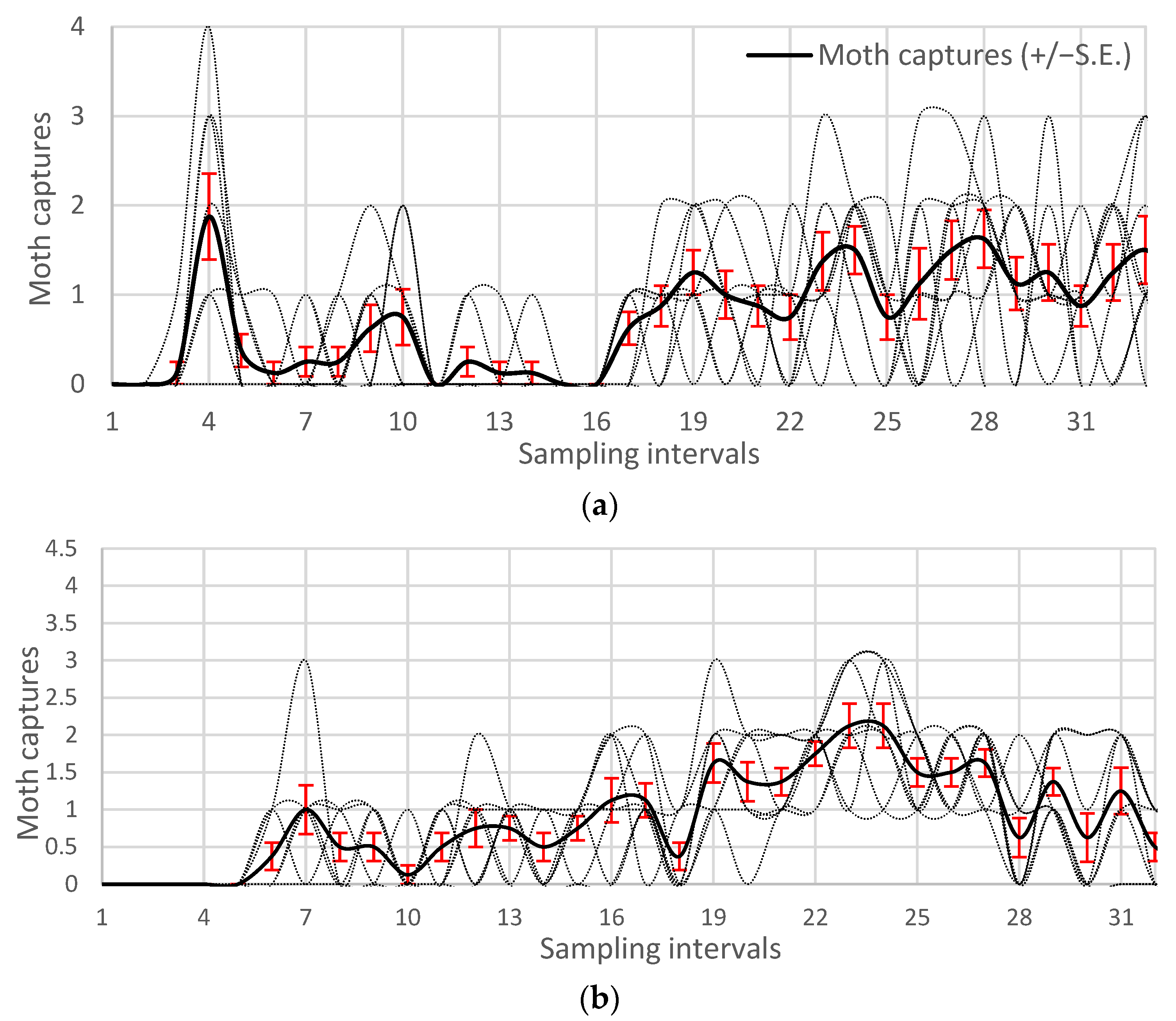
3.1. Moth Phenology and Population Density
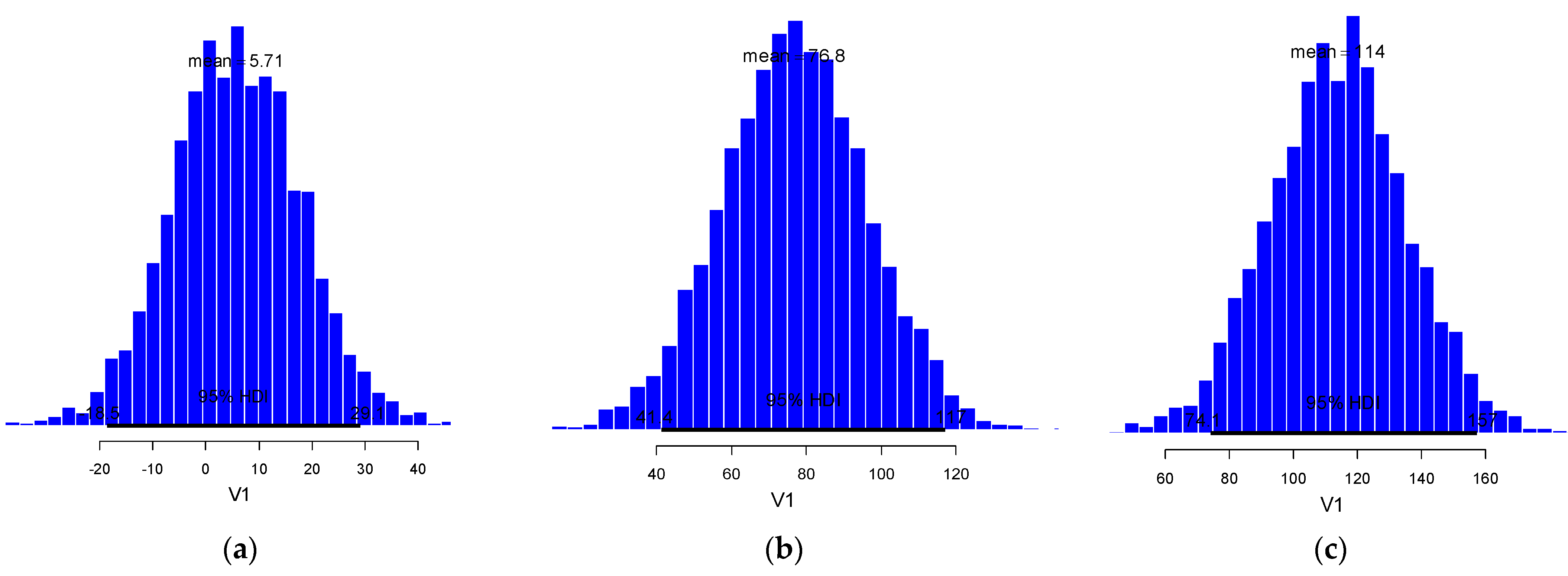
3.2. Bootstrapped Phenology and Bayesian Degree-Day Thresholds
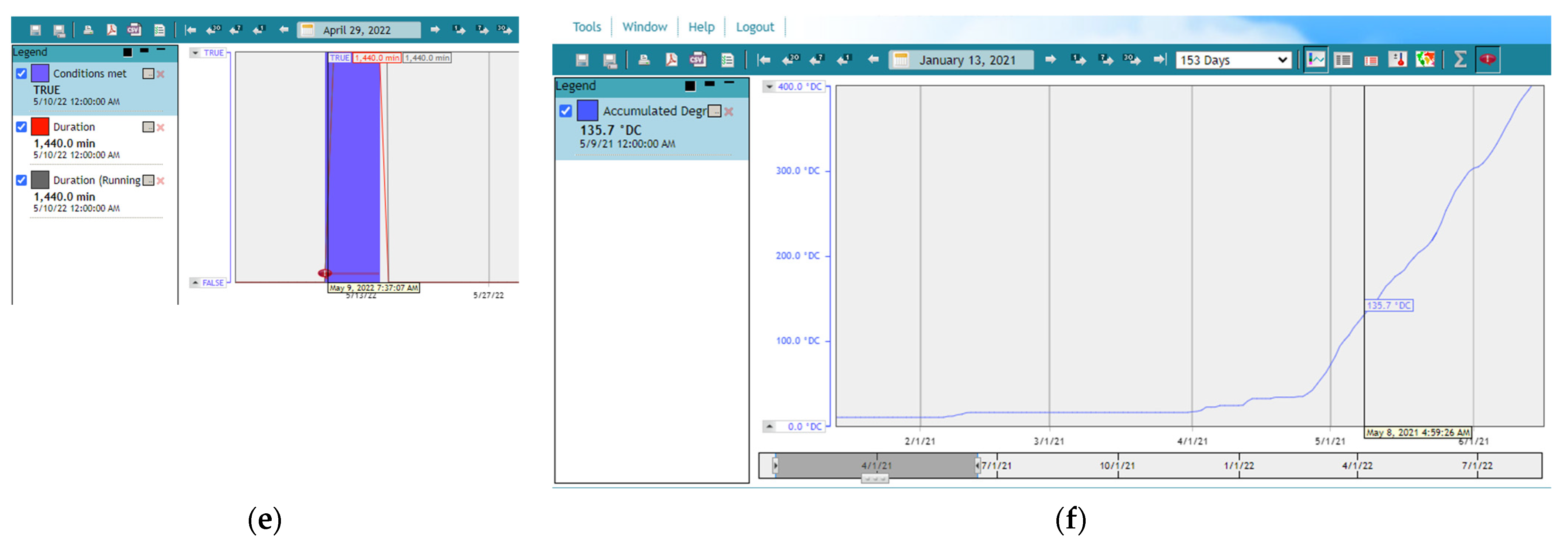
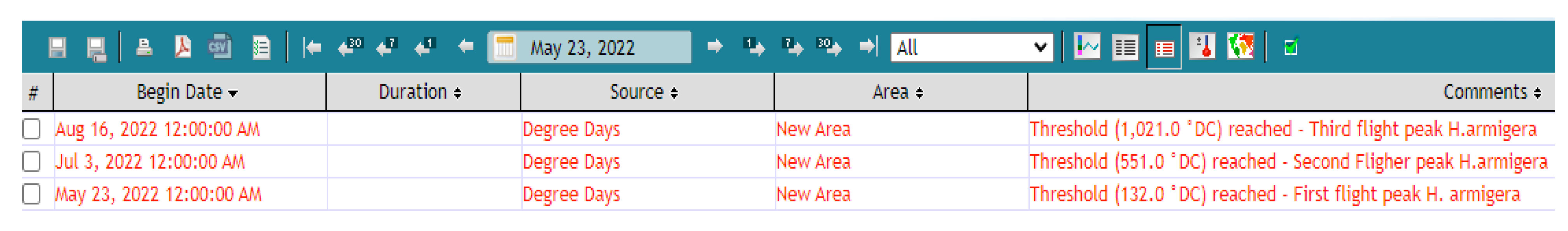
3.3. Decision Support System and H. armigera Forecasts
4. Discussion
5. Conclusions
Author Contributions
Funding

Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parde, V.D.; Sharma, H.C.; Kachole, M.S. Protease Inhibitors in Wild Relatives of Pigeonpea against the Cotton Bollworm/Legume Pod Borer, Helicoverpa armigera. Am. J. Plant Sci. 2001, 3, 70–72. [Google Scholar]
- Zalucki, M.P.; Daglish, G.; Firempong, S.; Twine, P.H. The biology and ecology of Heliothis armigera (Hubner) and H. punctigera Wallengren (Lepidoptera: Noctuidae) in Australia: What do we Know? Aust. J. Zool. 1986, 34, 779–814. [Google Scholar] [CrossRef]
- Sharma, H.C. Heliothis/Helicoverpa Management: Emerging Trends and Strategies for Future Research; Oxford and IBH Publishing Co., Pvt. Ltd.: New Delhi, India, 2005; p. 469. [Google Scholar]
- Smith, I.M.; McNamara, D.G.; Scott, P.R.; Holderness, M. Quarantine Pests for Europe; CAB International: Wallingford, UK, 1997. [Google Scholar]
- Damos, P. Current issues in integrated pest management of Lepidoptera pest threats in Industrial crop models. In Lepidoptera: Ecology, Behavior and Management; Guerritore, E., DeSare, J., Eds.; NovaScience: New York, NY, USA, 2012; ISBN 978-1-62417-249-6. [Google Scholar]
- Fitt, G.P. The ecology of Heliothis species in relation toagro-ecosystems. Ann. Rev. Entomol. 1989, 34, 17–52. [Google Scholar] [CrossRef]
- Damos, P.; Tsikos, E.; Louta, M.; Papathanasiou, F. Towards the Development of a Smart Plant Protection Solution for Improved Pest Management of Dry Beans (Phaseolus vulgaris L.) in Northern Greece. In Proceedings of the 1st International Electronic Conference on Entomology, Basel, Switzerland, 1–15 July 2021. [Google Scholar] [CrossRef]
- Papadopoulos, I.; Papathanasiou, F.; Vakali, C.; Kazoglou, I.; Tamoutsidis, E. Local landraces of dry beans (Phaseolus vulgaris L.): A valuable resource for organic production in Greece. Acta Hortic. 2012, 933, 75–81. [Google Scholar] [CrossRef]
- Papathanasiou, F.; Barbayiorgis, A.; Papadopoulou, V.; Kareklas, E.; Galaitsis, D.; Papadopoulou, F.; Tamoutsidis, E.; Papadopoulos, I. Physiological performance and yield of dry bean (Phaseolus vulgaris L.) genotypes under water deficit. In Proceedings of the Book of Abstracts of the AgriBalkan-Balkan Agricultural Congress, Edirne, Turkey, 8–11 September 2014; p. 398. [Google Scholar]
- Zalucki, M.P. Heliothis: Research Methods and Prospects; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Carvalho, F.P. Pecticides, environment and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Pal, S.; Chatterjee, H.; Senapati, S.K. Monitoring of Helicoverpa armigera using pheromone traps and relationship of moth activity with larval infestation on Carnation (Dianthus caryophyllus) in Darjeeling Hills. J. Entomol. Res. 2014, 38, 23–26. [Google Scholar]
- Kumar Sharma, P.; Kumar, U.; Vyas, S.; Sharma, S.; Shrivastava, S. Monitoring of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) Through Pheromone Traps In Chickpea (Cicer arietinum) Crop and influence of Some abiotic factors on insect population. IOSR J. Environ. Sci. Toxicol. Food Technol. (IOSR-JESTFT) 2012, 1, 44–46. [Google Scholar] [CrossRef]
- McGeachie, W. The effects of moonlight illuminance, temperature and wind speed on light-trap catches of moths. Bull. Entmol. Res. 1989, 79, 185–192. [Google Scholar] [CrossRef]
- Choi, S.W. The effects of weather factors on the abundance and diversity of moths in a temperate deciduous mixed forest of Korea. Zool. Sci. 2018, 25, 53–58. [Google Scholar] [CrossRef]
- Nibouche, S.; Gozé, E.; Babin, R.; Beyo, J.; Brévault, T. Modeling Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) damages on cotton. Environ. Entomol. 2007, 36, 151–156. [Google Scholar] [CrossRef]
- Dalal, P.K.; Arora, R. Model-based phenology prediction of Helicoverpa armigera (Hübner) (Noctuidae: Lepidoptera) on tomato crop. J. Plant Dis. Prot. 2019, 126, 281–291. [Google Scholar] [CrossRef]
- Jallow, F.A.M.; Masaya, M. Infuence of temperature on the rate of development of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Appl. Entomol. Zool. 2001, 36, 427–430. [Google Scholar] [CrossRef]
- Amer, A.E.A.; El-Sayed, A. Effect of different host plants and artificial diet on Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) development and growth index. J. Entomol. 2014, 11, 299–305. [Google Scholar] [CrossRef]
- Damos, P.; Savopoulou-Soultani, M. Temperature driven models for Insect development and Vital Thermal Requirements. Psyche 2012, 2012, 123405. [Google Scholar] [CrossRef]
- Damos, P.; Karabatakis, S. Real time pest modeling trhough the world wide web: Decision making from theory to praxis. IOBC-WPRS Bull. 2013, 91, 253–258. [Google Scholar]
- Edwars, C.; Crone, E.E. Fitting phenological curves with GLMMs. Oikos 2022, 130, 1335–1345. [Google Scholar]
- Damos, P. A stepwise algorithm to detect significant time lags in ecological time series in terms of autocorrelation functions and ARMA model optimisation of pest population seasonal outbreaks. Stoch. Environ. Res. Risk Assess. 2016, 30, 1961–1980. [Google Scholar] [CrossRef]
- Munholland, P.L.; Kalbfleish, J.D.; Dennis, B. A stochastic model for insect life history data. In Estimating and Analysis of Insect Populations. Lecture Notes in Statistics; McDonald, L.L., Manly, B.F.L., Lochwood, J.A., Logan, J.A., Eds.; Springer: New York, NY, USA, 1988; Volume 55. [Google Scholar]
- Maurer, J.A.; Shepard, J.H.; Crabo, L.G.; Hammond, P.C.; Zack, R.S.; Peterson, M.A. Phenological responses of 215 moth species to interannual climate variation in the Pacific Northwest from 1895 through 2013. PLoS ONE. 2018, 13, e0202850. [Google Scholar] [CrossRef] [PubMed]
- Damos, P.; Soulopoulou, P. Do Insect Populations Die at Constant Rates as They Become Older? Contrasting Demographic Failure Kinetics with Respect to Temperature According to the Weibull Model. PLoS ONE 2015, 10, e0127328. [Google Scholar]
- Moussus, J.P.; Jiquet, J.F. Featuring phenological estimators using simulated data. Methods Ecol. Evol. 2010, 1, 140–150. [Google Scholar] [CrossRef]
- Damos, P. Demography and randomized life table statistics for the peach twig borer Anarsia lineatella (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2013, 106, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Gershman, S.J.; Blei, D.M. A tutorial on Bayesian nonparametric models. J. Math. Psychol. 2012, 56, 1–12. [Google Scholar] [CrossRef]
- Shao, J.; Tu, D. Bayesoan bootrsrap and random weighting. In The Jacknife and Bootrsap. Springer Series in Statistics; Springer: New York, NY, USA, 1995. [Google Scholar]
- Muliere, P.; Secchi, P. Bayesian nonparametric inference and boostrap techniques. Ann. Inst. Stat. Math. 1996, 48, 663–673. [Google Scholar] [CrossRef]
- Galvani, M.; Bardelli, C.; Figini, S.; Muliere, P. A Bayesian Nonparametric Learning Approach to Ensemble Models Using the Proper Bayesian Bootstrap. Algorithms 2021, 14, 11. [Google Scholar] [CrossRef]
- Ellison, A.M. Bayesian inference in ecology. Ecol. Lett. 2004, 7, 509–520. [Google Scholar] [CrossRef]
- Morevie, M.A.; Davison, A.C.; Pasquier, D.; Charmillot, P.J. Bayesian forecasting of grape moth emergence. Ecol. Mod. 2006, 197, 478–489. [Google Scholar] [CrossRef]
- Wesner, J.S.; Pomeranz, J.P.F. Choosing priors in Bayesian ecological models by simulating from the prior predictive distribution. Ecosphere 2021, 12, e03739. [Google Scholar] [CrossRef]
- Sanbasivan, R.; Das, S.; Sahu, S.K. A Bayesian perspective of statistical machine learning for big data. Comput. Stat. 2020, 35, 893–930. [Google Scholar] [CrossRef]
- Singh, N.; Gupta, N. Bayesian network for decision-support on pest management of tomato fruit borer, H. armigera. Int. J. Eng. Technol. 2017, 6, 168–170. [Google Scholar] [CrossRef]
- Apel, H.; Herrmann, A.; Richter, O. A decision support system for integrated pest managment of Helicoverpa armigera in the tropics and subtropics by means of a rule-based Fuzzy model. Z. Fur Agrainformatik 1999, 7, 83–90. [Google Scholar]
- Narava, R.; Sai Ram Kumar, D.V.; Jaba, J.; Kumar, A.P.; Rao, R.G.V.; Rao, S.R.; Mishra, S.P.; Kukanur, V. Development of Temporal Model for Forecasting of Helicoverpa armigera (Noctuidae: Lepidopetra) Using Arima and Artificial Neural Networks. J. Insect Sci. 2022, 22, 2. [Google Scholar] [CrossRef] [PubMed]
- Nanushi, O.; Sitokonstantinou, V.; Tsoumas, I.; Konteos, C. Pest presence prediction using interpretable machine learning. arXiv 2022, arXiv:2205.07723. [Google Scholar]
- Almalki, F.A.; Soufiene, B.O.; Alsamhi, S.H.; Sakli, H. A Low-Cost Platform for Environmental Smart Farming Monitoring System Based on IoT and UAVs. Sustainability 2021, 13, 5908. [Google Scholar] [CrossRef]
- Xu, M.; David, J.M.; Kim, S.H. The fourth industrial revolution: Opportunities and challenges. Int. J. Financ. Res. 2018, 9, 90–95. [Google Scholar] [CrossRef]
- Tinte, M.M.; Chele, K.H.; van der Hooft, J.J.J.; Tugizimana, F. Metabolomics-Guided Elucidation of Plant Abiotic Stress Responses in the 4IR Era: An Overview. Metabolites 2021, 11, 445. [Google Scholar] [CrossRef]
- Barteková, A.; Praslička, J. The effect of ambient temperature on the development of cotton bollworm (Helicoverpa armigera Hübner, 1808). Plant Prot. Sci. 2006, 42, 135–138. [Google Scholar] [CrossRef]
- Mironidis, G.K.; Savopoulou-Soultani, M. Development, Survivorship and Reproduction of Helicoverpa armigera (Lepidoptera: Noctuidae) Under Constant and Alternating Temperatures. Environ. Entomol. 2008, 37, 16–28. [Google Scholar] [CrossRef]
- Damos, P.T.; Savopoulou-Soultani, M. Development and statistical evaluation of models in forecasting moth phenology of major lepidopterous peach pest complex for Integrated Pest Management programs. Crop Prot. 2010, 29, 1190–1199. [Google Scholar] [CrossRef]
- Chernick, M.R. Boostrap Methods: A Practitioner’s Guide (Wiley Series in Probability and Statistics), 1st ed.; Wiley-Interscience: New York, NY, USA, 2007. [Google Scholar]
- Efron, B. The Jackknife, the Bootstrap and Other Resampling Plans; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1982. [Google Scholar]
- Efron, B.; Tibshirani, R. Software (Bootstrap, Cross-Validation, Jackknife) and Data for the Book “An Introduction to the Bootstrap”, In Package Bootrstap; Chapman and Hall: London, UK, 1993. [Google Scholar]
- Alfaro, M.E.; Zoller, S.; Lutzoni, F. Bayes or boostrap? A simulation study comparing the performance of Bayesian Markov chain Monte Carlo sampling and bootstrapping in assessing phylogenetic confidence. Mol. Biol. Evol. 2003, 20, 255–266. [Google Scholar] [CrossRef]
- Rubin, D.B. The bayesian boostrap. Ann. Stat. 1981, 9, 130–134. [Google Scholar] [CrossRef]
- Calatayud, J.; Jornet, M.; Mateu, J. A stochastic Bayesian bootstrapping model for COVID-19 data. Stoch. Environ. Res. Risk Assess. 2022, 36, 2907–2917. [Google Scholar] [CrossRef] [PubMed]
- Dirichletprocess Package Dirichletprocess: An R Package for Fitting Complex Bayesian Nonparametric Models. Available online: https://cran.r-project.org/web/packages/dirichletprocess/vignettes/dirichletprocess.pdf (accessed on 10 August 2022).
- van Strien, A.J.; Plantenga, W.F.; Sodaat, L.L.; van Swaay, C.A.M.; WallisDeVries, M.F. Bias in phenology assessments based on first appearance data of butterlies. Oecologia 2008, 156, 227–235. [Google Scholar] [CrossRef]
- Miller-Rushing, J.A.; Inouye, D.W.; Primack, R.B. How well do first flowering dates measure plant responses to climate change? The effects of population size and sampling frequency. J. Ecol. 2008, 96, 1289–1296. [Google Scholar]
- Inouye, B.D.; Ehrlén, J.; Undeewoos, N. Phenology as a process rather than an event: From individual reaction norms to community metrics. Ecol. Monog. 2019, 89, e01352. [Google Scholar] [CrossRef]
- Steward, T.J. Experience with a Byesian bootstrap method incorporating proper prior information. Commun. Stat. Theory Meth. 1986, 15, 3205–3225. [Google Scholar] [CrossRef]
- Damos, P.; Bonsignore, C.P.; Gardi, F.; Avtzis, D.N. Phenological responses and a comperative phylogenetic insight of Anarsia lineatella and Grapholita molesta between distinct geographical regions within the Mediterranean basin. J. Appl. Entomol. 2014, 138, 528–538. [Google Scholar] [CrossRef]
- EPPO/CABI. European and Mediterranean Plant Protection Organization/CAB International. Helicoverpa armigera. In Quarantine Pests for Europe, 2nd ed.; Smith, I.M., McNamara, D.G., Scott, P.R., Holderness, M., Eds.; CAB International: Wallingford, UK, 1997; pp. 289–294. [Google Scholar]
- Damos, P.; Mantzoukas, S.; Theoharis, X.; Zaggos, G.; Staurakoulis, N.; Karanastasi, E.; Perdikis, D. Development and first evaluation of seasonal and spatial models of Helicoverpa armigera and Tuta absoluta in industrial tomato cultivations in the prefectures of Ilia and Achaia. In Proceedings of the 16th PanHellenic Conference on Entomology, Heraklion, Greece, 20–30 October 2015. [Google Scholar]
- Mironidis, G.K.; Stamopoulos, D.C.; Savopoulou-Soultani, M. Overwintering survival and spring emergence of Helicoverpa armigera (Lepidoptera: Noctuidae) in northern Greece. Environ. Entomol. 2010, 39, 1068–1084. [Google Scholar] [CrossRef]
- Baker, G.H.; Tann, C.R.; Fitt, G.P. A tale of two trapping methods: Helicoverpa spp. (Lepidoptera, Noctuidae) in pheromone and light traps in Australian cotton production systems. Bull. Entomol. Res. 2011, 101, 9–23. [Google Scholar] [CrossRef]
- Zhou, X.; Applebaum, S.W.; Coll, M. Overwintering and spring migration in the bollworm Helicoverpa armigera (Lepidoptera: Noctuidae) in Israel. Environ. Entomol. 2000, 29, 1289–1294. [Google Scholar] [CrossRef]
- Qureshi, M.H.; Murai, T.; Yoshida, H.; Shiraga, T.; Tsumuki, H. Population variation in diapause induction and termination of Helicoverpa armigera (Lepideptera: Noctuidae). Appl. Entomol. Zool. 2000, 35, 357–360. [Google Scholar] [CrossRef]
- Jokar, M. A thermal forecasting model for the overwintering generation of cotton bollworm by remote sensing in the southeast of Caspian Sea. Span. J. Agric. Res. 2022, 20, e1001. [Google Scholar] [CrossRef]
- Liu, B.; Yang, L.; Yang, F.; Wang, Q.; Yang, Y.Z.; Lu, Y.H.; Gardiner, M.M. Landscape diversity enhances parasitism of cotton bollworm (Helicoverpa armigera) eggs by Trichogramma chilonis in cotton. Biol. Control. 2016, 93, 15–23. [Google Scholar] [CrossRef]
- Bheemanna, M.; Ashoka, J.; Wali, V.; Dave, S.; Bheemanna, M.; Ashoka, J.; Shivayogiyappa, P.; Lim, K.S.; Chapman, J.W.; Sane, S.P. Evidence for facultative migratory flight behavior in Helicoverpa armigera (Noctuidae: Lepidoptera) in India. PLoS ONE 2021, 16, e0245665. [Google Scholar]
- Behere, G.T.; Tay, W.T.; Russell, D.A.; Kranthi, K.R.; Batterham, P. Population genetic structure of the cotton bollworm Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) in India as inferred from EPIC-PCR DNA markers. PLoS ONE 2013, 8, e53448. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.Q.; Wu, K.M.; Ni, Y.X.; Cheng, D.F.; Guo, Y.Y. High-Altitude Windborne Transport of Helicoverpa armigera (Lepidoptera: Noctuidae) in Mid-Summer in Northern China. J Insect Behav 2005, 18, 335–349. [Google Scholar] [CrossRef]
- Aheer, G.M.; Ali, A.; Akram, M. Effect of weather factors on populations of Helicoverpa armigera moths at cotton-based agro-ecological sites. Entomol. Res. 2009, 29, 36–42. [Google Scholar] [CrossRef]
- Huang, J. Effects of climate change on different geographical populations of the cotton bollworm Helicoverpa armigera (Lepidoptera, Noctuidae). Ecol. Evol. 2021, 11, 18357–18368. [Google Scholar] [CrossRef]
- Rousselet, G.A.; Pernet, C.R.; Wilcox, R.R. The percentile Bootstrap: A primer with step-by-step instructions in R. Adv. Meth. Pract. Psych. Sci. 2021, 4, 2515245920911881. [Google Scholar] [CrossRef]
- Bååth, R. The Non-Parametric Bootstrap as a Bayesian Model. Publishable Stuff. Available online: http://www.sumsar.net/ blog/2015/04/the-non-parametric-bootstrap-as-a-bayesianmodel/ (accessed on 18 April 2015).
- Canty, A.; Ripley, B.D. Boot: Bootstrap Functions, Version 1.3-25; Comprehensive R Archive Network. Available online: https://CRAN.R-project.org/package=boot (accessed on 5 August 2022).
- Louta, M.; Papathanasiou, F.; Damos, P.; PLoSkas, N.; Dasygenis, M.; Kyriakidis, T.; Dimokas, N.; Balafas, V.; Chatzisavvas, A.; Karampelia, I.; et al. Intelligent pesticide and irrigation management in precision agriculture: The case of VELOS project. HAICTA. In Proceedings of the 10th International Conference on ICT in Agriculture, Food & Environment, Athens, Greece, 22–25 September 2022. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
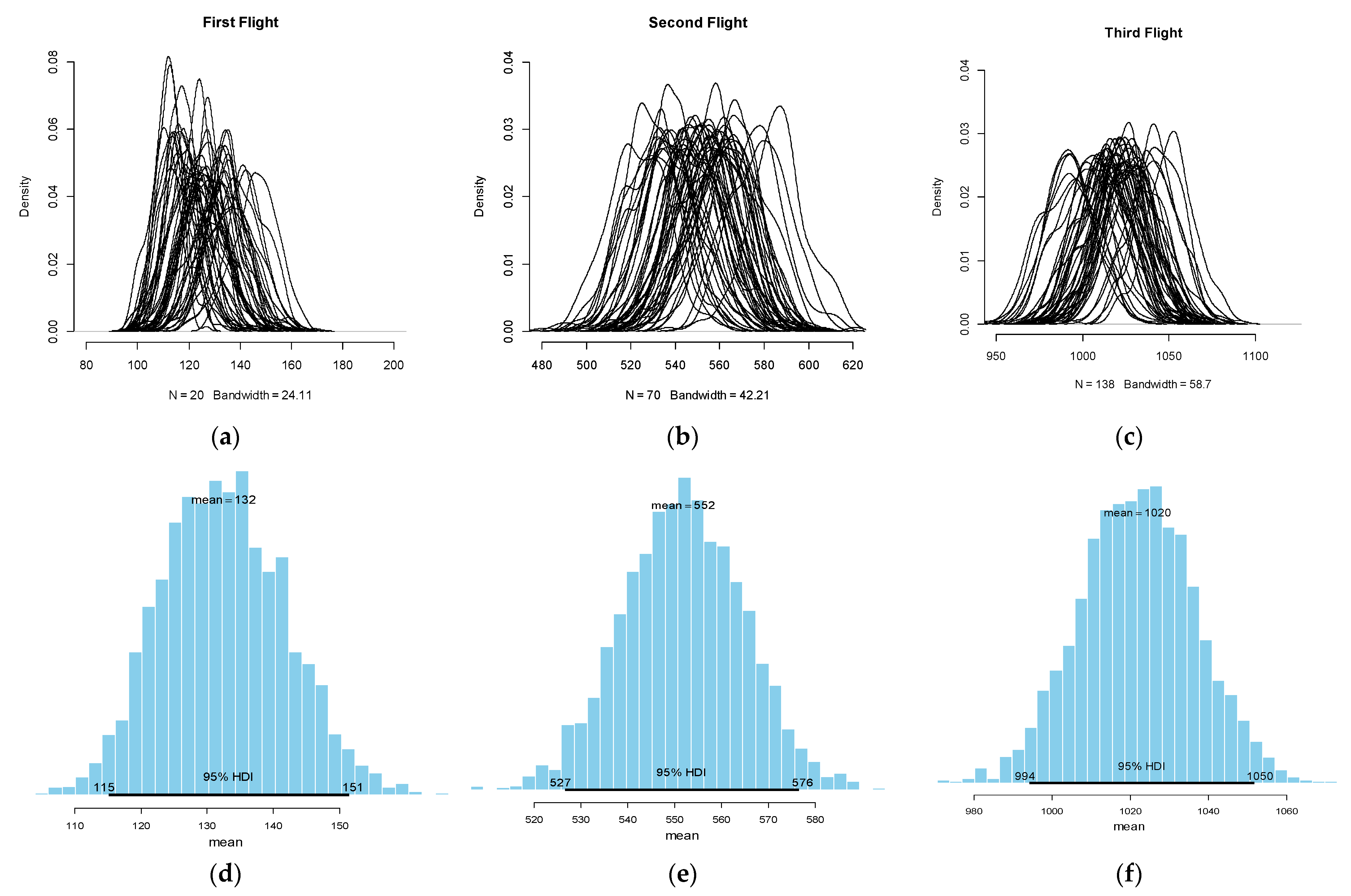
| Summary Statistics of the Posterior | ||||||
| Flight Generation | Draws | HDI *. Low | Mean | HDI. High | sd | Midrange |
| First (F1) | 10,000 | 113.13 | 132.13 | 149.75 | 9.32 | 170 |
| Second (F2) | 10,000 | 527.78 | 551.72 | 579.27 | 13.2 | 531.9 |
| Third (F3) | 10,000 | 992.2 | 1021.6 | 1049.6 | 14.8 | 1020.3 |
| Quantiles and tendencies | ||||||
| Q2.5% | Q25% | median | Q75% | Q97.5% | mode | |
| First (F1) | 114.04 | 125.76 | 131.83 | 138.21 | 150.96 | 79.4 |
| Second (F2) | 525.83 | 542.56 | 551.72 | 561.05 | 577.6 | 675.6 |
| Third (F3) | 992.7 | 1011.5 | 1021.56 | 1031.82 | 1050.09 | 1049.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damos, P.; Papathanasiou, F.; Tsikos, E.; Kyriakidis, T.; Louta, M. Bayesian Non-Parametric Thermal Thresholds for Helicoverpa armigera and Their Integration into a Digital Plant Protection System. Agronomy 2022, 12, 2474. https://doi.org/10.3390/agronomy12102474
Damos P, Papathanasiou F, Tsikos E, Kyriakidis T, Louta M. Bayesian Non-Parametric Thermal Thresholds for Helicoverpa armigera and Their Integration into a Digital Plant Protection System. Agronomy. 2022; 12(10):2474. https://doi.org/10.3390/agronomy12102474
Chicago/Turabian StyleDamos, Petros, Fokion Papathanasiou, Evaggelos Tsikos, Thomas Kyriakidis, and Malamati Louta. 2022. "Bayesian Non-Parametric Thermal Thresholds for Helicoverpa armigera and Their Integration into a Digital Plant Protection System" Agronomy 12, no. 10: 2474. https://doi.org/10.3390/agronomy12102474
APA StyleDamos, P., Papathanasiou, F., Tsikos, E., Kyriakidis, T., & Louta, M. (2022). Bayesian Non-Parametric Thermal Thresholds for Helicoverpa armigera and Their Integration into a Digital Plant Protection System. Agronomy, 12(10), 2474. https://doi.org/10.3390/agronomy12102474