
Low-Cost Spore Traps: An Efficient Tool to Manage Fusarium Head Blight through Improved Cropping Systems



Abstract
1. Introduction
2. Materials and Methods
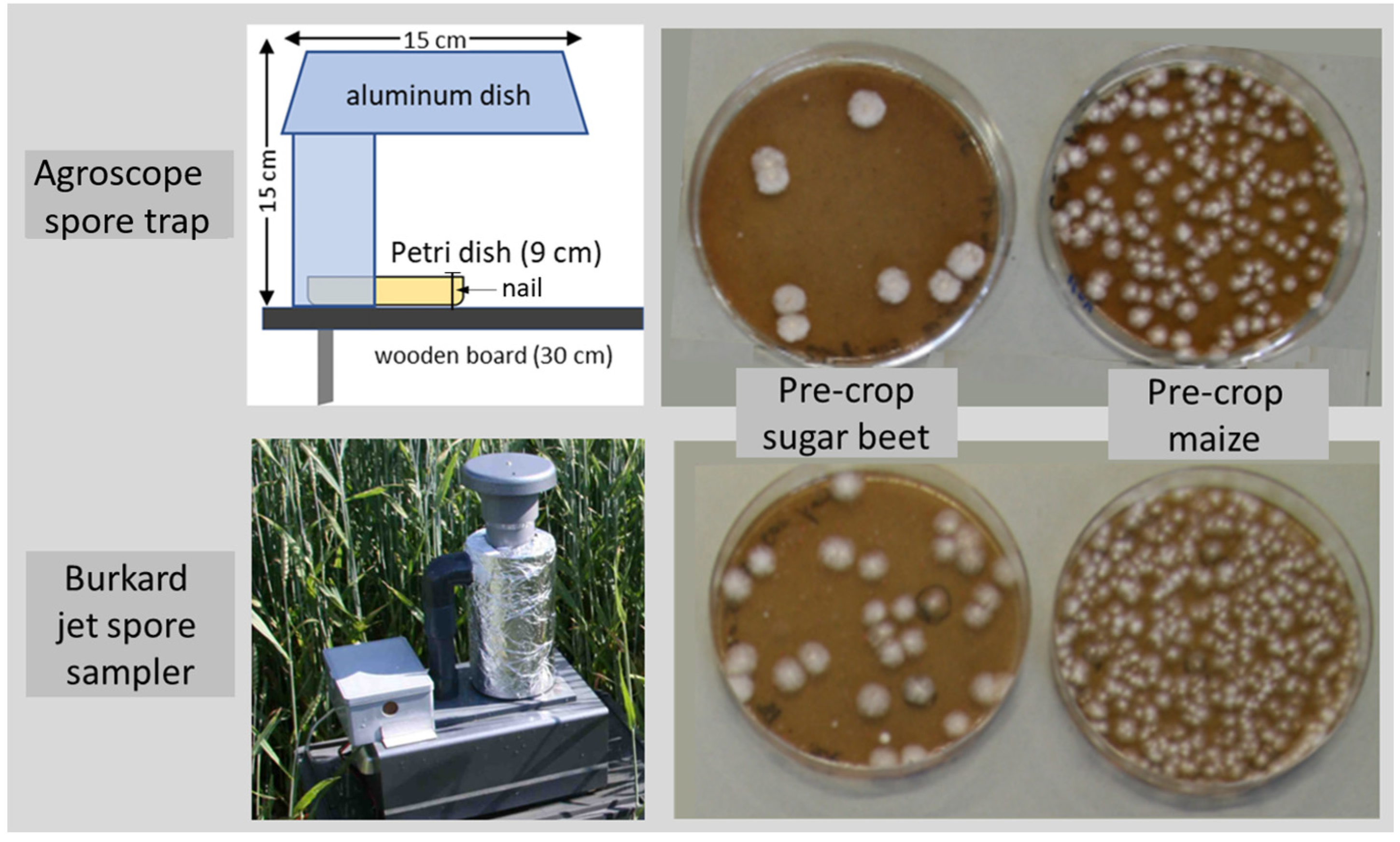
2.1. Spore Trap Construction and Utilisation
2.2. Deposition of Fusarium Colonies on Agar Plates and Species Identification
2.3. Field Experiments: Locations, Cropping Systems and Weather Data
2.4. Wheat Harvest and Analyses of Deoxynivalenol (DON)
2.5. Statistical Analysis
3. Results
3.1. Use and Potential of the Agroscope Spore Trap
3.1.1. Deposition of F. graminearum Colony Forming Units (CFU) in Different Cropping Systems and Correlation of the Number of CFUs Sampled at Different Heights in the Wheat Canopy (2008)
3.1.2. Deposition of F. graminearum Colony-Forming Units (CFU) during the Day and Night and Their Relationship with the Relative Humidity of the Air (2008)
3.2. Comparison of the Agroscope (AGS) Spore Trap with a High-Throughput Jet Spore Sampler
3.2.1. Deposition of F. graminearum Colony-Forming Units (CFU) from April until August 2010
3.2.2. Deposition of Various Fusarium Species in Three Periods from April until August 2010 on the Agroscope (AGS) Spore Trap and the High Throughput Jet Sampler (JSS)
3.3. Relationship between Deposited Spores on Agroscope (AGS) Spore Traps in No-Tillage on-Farm Field Experiments and Deoxynivalenol (DON) Content in Wheat Grains
4. Discussion
4.1. Spore Dispersal in Different Cropping Systems and Spore Deposition at Different Height in the Wheat Canopy
4.2. Dispersal of Fusarium graminearum Spores over Day and Night
4.3. Comparison of Spore Traps and Seasonal Dynamics of Airborne Fusarium graminearum and Other Fusarium Species
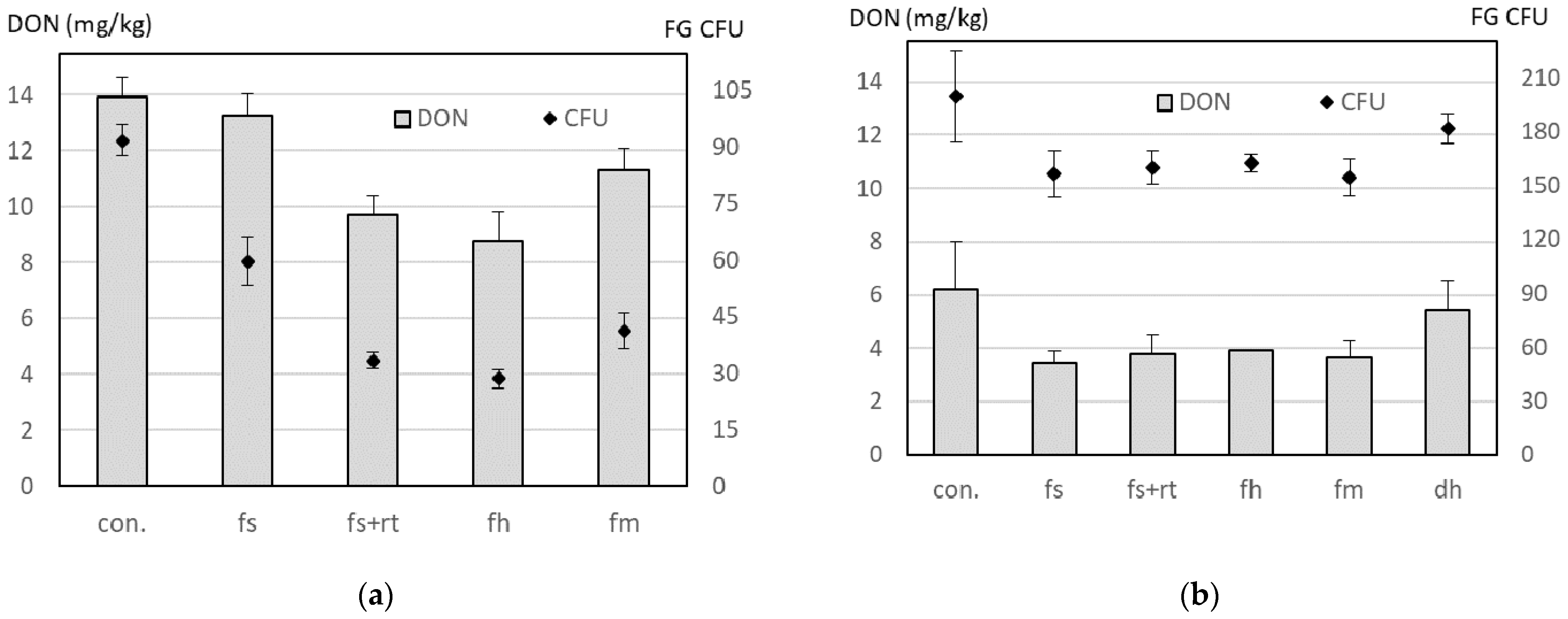
4.4. Correlation between Colony Forming Units Sampled with the Agroscope (AGS) Spore Trap and the Deoxynivalenol (DON) Content in Wheat Grains in On-Farm Experiments with Maize Residue Treatments
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernando, W.G.D.; Oghenekaro, A.O.; Tucker, J.R.; Badea, A. Building on a foundation: Advances in epidemiology, resistance breeding, and forecasting research for reducing the impact of fusarium head blight in wheat and barley. Can. J. Plant Pathol. 2021. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.-R. An eight-year survey of wheat shows distinctive effects of cropping factors on different Fusarium species and associated mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional oats. Food Addit. Contam., Part A. 2009, 26, 1063–1069. [Google Scholar] [CrossRef]
- Schöneberg, T.; Musa, T.; Forrer, H.-R.; Mascher, F.; Bucheli, T.D.; Bertossa, M.; Keller, B.; Vogelgsang, S. Infection conditions of Fusarium graminearum in barley are variety specific and different from those in wheat. Eur. J. Plant Pathol. 2018, 151, 975–989. [Google Scholar] [CrossRef]
- Xu, X.M.; Berrie, A.M. Epidemiology of mycotoxigenic fungi associated with Fusarium ear blight and apple blue mould: A review. Food Addit. Contam. 2005, 22, 290–301. [Google Scholar] [CrossRef] [PubMed]
- van der Lee, T.; Zhang, H.; van Diepeningen, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. Part A. 2015, 32, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Dill-Macky, R.; Jones, R.K. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Khonga, E.B.; Sutton, J.C. Inoculum production and survival of Gibberella zeae in maize and wheat residues. Can. J. Plant Pathol. 1988, 10, 232–239. [Google Scholar] [CrossRef]
- Blandino, M.; Haidukowski, M.; Pascale, M.; Plizzari, L.; Scudellari, D.; Reyneri, A. Integrated strategies for the control of Fusarium head blight and deoxynivalenol contamination in winter wheat. Field Crops Res. 2012, 133, 139–149. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Hecker, A.; Musa, T.; Dorn, B.; Forrer, H.R. On-farm experiments over five years in a grain maize—winter wheat rotation: Effect of maize residue treatments on Fusarium graminearum infection and deoxynivalenol contamination in wheat. Mycotoxin Res. 2011, 27, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the post genomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Fernando, W.G.D.; Miller, J.D.; Seaman, W.L.; Seifen, K.; Paulitz, T.C. Daily and seasonal dynamics of airborne spores of Fusarium graminearum and other Fusarium species sampled over wheat plots. Can. J. Plant Pathol. 2000, 78, 497–505. [Google Scholar]
- Hellin, P.; Duvivier, M.; Dedeurwaerder, G.; Bataille, C.; De Proft, M.; Legrève, A. Evaluation of the temporal distribution of Fusarium graminearum airborne inoculum above the wheat canopy and its relationship with Fusarium head blight and DON concentration. Eur. J. Plant Pathol. 2018, 151, 1049–1064. [Google Scholar] [CrossRef]
- Papavizas, G.C. Evaluation of various media and antimicrobial agents for isolation of Fusarium from soil. Phytopathology 1967, 57, 848–852. [Google Scholar]
- Nirenberg, H. A simplified method for identifying Fusarium spp. occurring in wheat. Can. J. Bot. 1981, 59, 1599–1609. [Google Scholar] [CrossRef]
- Singh, K.; Frisvad, J.C.; Thrane, U.; Mathur, S.B. An Illustrated Manual on Identification of some Seed-borne Aspergilli, Fusarium, Penicillia and their Mycotoxins; Danish Government, Institute of Seed Pathology for Developing Countries: Copenhagen, Denmark, 1991. [Google Scholar]
- Nelson, P.E.; Toussoun, T.A.; Marassas, W.F.O. Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: State College, PA, USA, 1983. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006; p. 388. [Google Scholar]
- Musa, T.; Hecker, A.; Vogelgsang, S.; Forrer, H.R. Forecasting of Fusarium head blight and deoxynivalenol content in winter wheat with FusaProg. EPPO Bull. 2007, 37, 283–289. [Google Scholar] [CrossRef]
- Forrer, H.R.; Hecker, A.; Musa, T.; Schwab, F.; Bucheli, T.D.; Wettstein, F.E.; Vogelgsang, S. Fusarium head blight control and prevention of mycotoxin contamination in wheat with botanicals and tannic acid. Toxins 2014, 6, 830–849. [Google Scholar] [CrossRef] [PubMed]
- Vogelgsang, S.; Sullyok, M.; Hecker, A.; Jenny, E.; Krska, R.; Schuhmacher, R.; Forrer, H.-R. Toxigenicity and pathogenicity of Fusarium poae and Fusarium avenaceum on wheat. Eur. J. Plant. Pathol. 2008, 122, 265–276. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals (maize, sorghum, forage grasses and dicotyledonous crops). Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- West, J.S.; Kimber, R.B.E. Innovations in air sampling to detect plant pathogens. Ann. Appl. Biol. 2015, 166, 4–17. [Google Scholar] [CrossRef]
- West, J.S.; Canning, G.G.M.; Perryman, S.A.; King, K. Novel technologies for the detection of Fusarium head blight disease and airborne inoculum. Trop. Plant Pathol. 2017, 42, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Manstretta, V.; Rossi, V. Effects of temperature and moisture on development of Fusarium graminearum perithecia in maize stalk residues. Appl. Environ. Microbiol. 2016, 82, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Ramirez, S.L.; Schmale, D.G.I.; Shields, E.J.; Bergstrom, G.C. The relative abundance of viable spores of Gibberella zeae in the planetary boundary layer suggests the role of long-distance transport in regional epidemics of Fusarium head blight. Agric. For. Meterol. 2005, 132, 20–27. [Google Scholar] [CrossRef]
- Paul, P.A.; El-Allaf, S.M.; Lipps, P.E.; Madden, L.V. Rain splash dispersal of Gibberella zeae within wheat canopies in Ohio. Phytopathology 2004, 94, 1342–1349. [Google Scholar] [CrossRef] [PubMed]
- Manstretta, V.; Rossi, V. Effects of weather variables on ascospore discharge from Fusarium graminearum perithecia. PLoS ONE 2015, 10, e0138860. [Google Scholar] [CrossRef]
- Trail, F.; Xu, H.; Loranger, R.; Gadoury, D. Physiological and environmental aspects of ascospore discharge in Gibberella zeae (anamorph Fusarium graminearum). Mycologia 2002, 94, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Paulitz, T.C. Diurnal release of ascospores by Gibberella zeae in inoculated wheat plots. Plant Dis. 1996, 80, 674–678. [Google Scholar] [CrossRef]
- Inch, S.; Fernando, W.G.D.; Gilbert, J. Seasonal and daily variation in the airborne concentration of Gibberella zeae (Schw. Petch) spores in Manitoba. Can. J. Plant Pathol. 2005, 27, 357–363. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Meca, G.; Torrijos, R.; Marty, A.; Kägi, A.; Jenny, E.; Forrer, H.-R.; Six, J.; Vogelgsang, S. Control of Fusarium graminearum in wheat with mustard-based botanicals: From in vitro to in planta. Front. Microbiol. 2020, 11, 1595. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, A.; Kägi, A.; Drakopoulos, D.; Bänziger, I.; Lehmann, E.; Forrer, H.-R.; Keller, B.; Vogelgsang, S. From laboratory to the field: Biological control of Fusarium graminearum on infected maize crop residues. J. Appl. Microbiol. 2020, 129, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, A.; Leimgruber, M.; Kägi, A.; Jenny, E.; Vogelgsang, S. UV protection and shelf life of the biological control agent Clonostachys rosea against Fusarium graminearum. Biol. Control 2021, 158, 104600. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field Experiment Number/Year and Site | Plot Code | Geographic Location | Wheat Variety (FHB Resistance) a | Previous Crop and Tillage | Experimental Question | Period of Spore Trapping |
|---|---|---|---|---|---|---|
| 1/2008 Agroscope * | A | N 47° 25′ 44″ E 08° 30′ 51″ | Titlis (+) | Pasture plough | Comparison of cropping systems and deposition of F. graminearum CFUs in AGS traps at 20, 50 and 90 cm height. Correlation of CFU numbers and DON | 19–29 May |
| 1/2008 Agroscope * | B | N 47° 25′ 47″ E 08° 30′ 57″ | Titlis (+) | Maize plough | ||
| 1/2008 Agroscope ** | C | N 47° 25′ 47″ E 08° 30′ 42″ | Zinal (∅) | Maize no-tillage | ||
| 2/2008 Agroscope ** | C | “ | “ | “ | Day and night spore deposition (AGS trap, 2 h interval) and relation with rh% | 17 June (14:00) to 19 June (14:00) |
| 3/2010 Agroscope * | A | N 47° 25′ 44″ E 08° 30′ 51″ | Levis (-) | Wheat no-tillage | Comparison of CFUs and ratios of Fusarium species sampled with the AGS and the JSS spore traps | Beginning of April–end of August |
| 4/2007 Ballmoos (BE) | N 47° 02′ 24″ E 07° 28′ 32″ | Ludwig (∅) | Maize no-tillage | Correlation of spore deposition in the AGS spore trap and the DON content in wheat grains after maize residue treatments in on-farm field trials | 5 June | |
| 5/2008 Ballmoos (BE) | N 47° 02′ 41″ E 07° 28′ 03″ | Runal (∅) | Maize no-tillage | 2, 5, 9 and 12 June | ||
| 6/2009 Ossingen (ZH) | N 46° 57′ 04″ E 07° 26′ 19″ | Ludwig (∅) | Maize no-tillage | 8, 9 and 10 June |
| Date of Sampling | Height of Spore Trap Above Ground | FG CFU on AGS Spore Traps 1 from Plot | Correlation 2 with DON | ||
|---|---|---|---|---|---|
| A | B | C | |||
| 22 May 2008 | 20 cm | 5.3 | 11.0 | 435.0 | 0.944 * |
| 50 cm | 6.0 | 5.0 | 359.7 | 0.958 ** | |
| 90 cm | 8.3 | 7.0 | 130.7 | 0.997 ** | |
| Average between 15 and 29 May 2008 | 20 cm | 1.6 | 1.8 | 64.4 | 0.949 ** |
| 50 cm | 1.7 | 1.2 | 49.5 | 0.952 ** | |
| 90 cm | 1.7 | 1.4 | 20.2 | 0.991 ** | |
| DON (mg/kg) 1 in wheat grains | 0.077 | 0.132 | 2.072 | ||
| % Wheat grains with FG 1 | 0.3 | 0.0 | 29 | ||
| Period | 1 April–10 June | 11 June–30 July | 1 August–31 August | |||
|---|---|---|---|---|---|---|
| Type of Spore Trap | AGS | JSS | AGS | JSS | AGS | JSS |
| F. graminearum | 57 | 58 | 55 | 72 | 85 | 96 |
| F. avenaceum | 20 | 18 | 16 | 8 | 4 | 1 |
| F. equiseti | 4 | 4 | 6 | 5 | 2 | 0 |
| F. cerealis | 1 | 1 | 10 | 2 | 1 | 0 |
| F. oxysporum | 4 | 3 | 1 | 2 | 0 | 0 |
| F. tricinctum | 1 | 2 | 4 | 1 | 0 | 0 |
| F. poae | 0 | 0 | 2 | 1 | 0 | 0 |
| F. spp. | 12 | 15 | 6 | 9 | 7 | 2 |
| Total nb 1 of colonies | 230 | 108 | 3′772 | 2′373 | 17′084 | 8′734 |
| Total nb 1 of isolates * | 214 | 108 | 588 | 351 | 328 | 284 |
| Correlation AGS/JSS ** | 0.997 | 0.977 | 0.998 | |||
| Year | Date of Sampling | Correlation between CFU and DON (Coefficient) | |||
|---|---|---|---|---|---|
| GS | 50 cm | 90 cm | Mean of 50 and 90 cm | ||
| 2007 | 5 June | 65–69 | 0.860 *** | 0.650 ** | 0.808 *** |
| 2008 | 2 June | 57 | 0.846 *** | 0.662 ** | 0.864 *** |
| 5 June | 59 | 0.610 ** | 0.567 * | 0.679 ** | |
| 9 June | 63 | 0.476 * | 0.592 ** | 0.666 ** | |
| 12 June | 69 | 0.514 * | 0.727 *** | 0.742 *** | |
| Ø 2–9 June (3 d) | 57–63 | 0.856 **** | 0.761 *** | 0.890 **** | |
| Ø 2–12 June (4 d) | 57–69 | 0.879 **** | 0.802 **** | 0.897 **** | |
| 2009 | Ø 8–10 June (3 d) | 69–71 | 0.449 ns | 0.669 *** | 0.606 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forrer, H.-R.; Pflugfelder, A.; Musa, T.; Vogelgsang, S. Low-Cost Spore Traps: An Efficient Tool to Manage Fusarium Head Blight through Improved Cropping Systems. Agronomy 2021, 11, 987. https://doi.org/10.3390/agronomy11050987
Forrer H-R, Pflugfelder A, Musa T, Vogelgsang S. Low-Cost Spore Traps: An Efficient Tool to Manage Fusarium Head Blight through Improved Cropping Systems. Agronomy. 2021; 11(5):987. https://doi.org/10.3390/agronomy11050987
Chicago/Turabian StyleForrer, Hans-Rudolf, Annegret Pflugfelder, Tomke Musa, and Susanne Vogelgsang. 2021. "Low-Cost Spore Traps: An Efficient Tool to Manage Fusarium Head Blight through Improved Cropping Systems" Agronomy 11, no. 5: 987. https://doi.org/10.3390/agronomy11050987
APA StyleForrer, H.-R., Pflugfelder, A., Musa, T., & Vogelgsang, S. (2021). Low-Cost Spore Traps: An Efficient Tool to Manage Fusarium Head Blight through Improved Cropping Systems. Agronomy, 11(5), 987. https://doi.org/10.3390/agronomy11050987