
Direct and Indirect Detoxification Effects of Humic Substances



Abstract
1. Introduction
2. Influence of HS on Living Organisms
3. Antioxidant Properties of HS via Bacterial Bioassay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orlov, D.S. Soil Humic Acids and General Theory of Humification; Moscow State University: Moscow, Russia, 1990; p. 364. (In Russian) [Google Scholar]
- Weber, J.H. Binding and transport of metals by humic materials. In Humic Substances and Their Role in the Environment; Frimmel, F.H., Christman, R.F., Eds.; J. Wiley & Sons Ltd: London, UK, 1988; pp. 165–178. [Google Scholar]
- Perminova, I.V.; Hatfield, K. Remediation chemistry of humic substances: Theory and implications for technology. In Use of Humic Substances to Remediate Polluted Environments: From Theory to Practice; Perminova, I.V., Ed.; Springer: Amsterdam, The Netherlands, 2005; pp. 3–36. [Google Scholar]
- Bezuglova, O.S.; Shestopalov, A.V. The Use of Humates for the Detoxification of Soils Contaminated with Heavy Metals. In Use of Humic Substances to Remediate Polluted Environments: From Theory to Practice; Perminova, I.V., Ed.; Springer: Amsterdam, The Netherlands, 2005; pp. 185–200. [Google Scholar]
- Stevenson, F.J. Humus Chemistry. Genesis, Composition, Reactions, 2nd ed.; Wiley: New York, NY, USA, 1994. [Google Scholar]
- Levinsky, B. All about Humate, 4th ed.; Кorf-Poligraf: Irkutsk, Russia, 2003. (In Russia) [Google Scholar]
- Steinberg, C.E.W. Ecology of Humic Substances in Freshwaters; Springer: Berlin/Heidelberg, Germany, 2003; p. 563. [Google Scholar]
- Weber., J. Humic Substances and their Role in the Environment. EC Agric. 2020, 1, 3–8. [Google Scholar]
- Lotosh, T.D. Experimental bases and prospects for the use of Humic Acid preparations from peat in medicine and agricultural production. Nauchnye Doki Vyss Shkoly Biol. Nauki. 1991, 1, 99–103. (In Russia) [Google Scholar]
- Piccolo, A.; Celano, G.; Pietramellara, G. Effects of fractions of coal-derived humic substances on seed germination and growth of seedlings (Lactuga sativa and Lycopersicum esculentum). Biol. Fertil Soil 1993, 16, 11–15. [Google Scholar] [CrossRef]
- Fedotov, G.N.; Shoba, S.A.; Fedotova, M.F.; Demin, V.V. On the Probable Nature of Biological Activity of Humic Substances. Eurasian Soil Sci. 2018, 51, 1034–1041. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Muscolo, A.; Vianello, A. Physiological effects of humic substances on higher plants. Soil Biol. Biochem. 2002, 34, 1527–1536. [Google Scholar] [CrossRef]
- Lee, Y.S.; Bartlett, R.J. Stimulation of plant growth by humic substances. Soil Sci. Soc. Am. J. 1976, 40, 876–879. [Google Scholar] [CrossRef]
- Quaggiotti, S.; Ruperti, B.; Pizzeghello, D.; Francioso, O.; Tugnoli, V.; Nardi, S. Effect of low molecular size humic substances on the expression of genes involved in nitrate transport and reduction in maize (Zea mays L.). J. Exp. Bot 2004, 55, 803–813. [Google Scholar] [CrossRef]
- Muscolo, A.; Bovalo, F.; Gionfriddo, F.; Nardi, S. Earthworm humic matter produces auxin-like effects on Daucus carota cell growth and nitrate metabolism. Soil Biol. Biochem. 1999, 3, 1303–1311. [Google Scholar] [CrossRef]
- Dell’Agnola, G.; Nardi, S. Hormone-like effect and enhanced nitrate uptake induced by depolycondensed humic fractions obtained from Allobophora rosea and A. caliginosa faeces. Biol. Fertil Soils 1987, 4, 115–118. [Google Scholar]
- Vaughan, D.; Ord, B.G. Uptake and incorporation of 14C-labelled soil organic matter by roots of Pisum sativum L. J. Exp. Bot 1981, 32, 679–687. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Okorokova-Façanha, A.L.; Façanha, A.R. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence and plasma membrane H+ -ATPase activity in maize roots. Plant Physiol. 2002, 130, 1951–1957. [Google Scholar] [CrossRef] [PubMed]
- Canellas, L.P.; Teixeira, L.R.L., Jr.; Dobbss, L.B.; Silva, C.A.; Medici, L.O.; Zandonadi, D.B.; Façanha, A.R. Humic acids crossinteractions with root and orgnic acids. Ann. Appl. Biol. 2008, 153, 157–166. [Google Scholar]
- Clapp, C.E.; Chen, Y.; Hayes, M.H.B.; Cheng, H.H. Plant growth promoting activity of humic substances. In Understanding and Managing Organic Matter in Soils, Sediments and Waters; Swift, R.S., Sparks, K.M., Eds.; International Humic Science Society: St. Paul, MN, USA, 2001; pp. 243–555. [Google Scholar]
- Dobbss, L.B.; Medici, L.O.; Peres, L.E.P.; Pino-Nunes, L.E.; Rumjianek, V.M.; Façanha, A.R.; Canellas, L.P. Changes in root development of Arabidopsis promoted by organic matter from oxisols. Ann. Appl. Biol. 2007, 151, 199–211. [Google Scholar] [CrossRef]
- O’Connor, N.E.; Emmerson, M.C.; Crowe, T.P.; Donohue, I. Distinguishing between direct and indirect effects of predators in complex ecosystems. J. Anim. Ecol. 2013, 82, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Mora, V.; Olaetxea, M.; Bacaicoa, E.; Baigorri, R.; Fuentes, M.; Zamarreno, A.M.; García-Mina, J.M. Abiotic stress tolerance in plants: Exploring the role of nitric oxide and humic substances. In Nitric Oxide in Plants: Metabolism and Role in Stress Physiology; Khan, N.M., Mobin, M., Firoz, M., Corpas, F.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 243–264. [Google Scholar]
- Chen, Y.; Aviad, T. Effects of humic substances on plant growth. In Humic Substances in Soil and Crop Science: Selected Readings; Mac-Carthy, P., Malcom, R.E., Clapp, C.E., Bloom, P.R., Eds.; Americal Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 1990; pp. 161–187. [Google Scholar]
- Trevisan, S.; Francioso, O.; Quaggiotti, S.; Serenella, N. Humic substances biological activity at the plant-soil interface from environmental aspects to molecular factors. Plant Signal. Behav. 2010, 5–6, 635–643. [Google Scholar] [CrossRef]
- Pflugmacher, S.; Spangenber, M.; Steinberg, C.E.W. Dissolved organic matter (DOM) and effects on the aquatic macrophyte Ceratophyllum demersum in relation to photosynthesis, pigment pattern and activity detoxication enzymes. J. Appl. Bot. 1999, 73, 184–190. [Google Scholar]
- Pflugmacher, S.; Tidwell, L.F.; Stainberg, C.E.W. Dissolved humic substances can directly affect freshwater organisms. Acta Hydrochim. Hydrobiol. 2001, 29, 34–40. [Google Scholar] [CrossRef]
- Glover, C.N.; Wood, C.M. The disruption of Daphnia magna sodium metabolism by humic substances. Physiol. Biochem. Zool. 2005, 78, 1005–1016. [Google Scholar] [CrossRef]
- Glover, C.N.; Pane, E.F.; Wood, C.M. Humic substances influence sodium metabolism in the freshwater crustacean Daphnia magna. Physiol. Biochem. Zool. 2005, 78, 405–416. [Google Scholar] [CrossRef]
- McCarthy, J.F.; Jimenez, B.D.; Barbee, T. Effect of dissolved humic material on accumulation of polycyclic aromatic hydrocarbons: Structure-activity relationships. Aquat. Toxicol. 1985, 7, 15–24. [Google Scholar] [CrossRef]
- Mézin, L.C.; Hale, R.C. Effect of humic acids on toxicity of DDT and chlorpyrifos to freshwater and estuarine invertebrates. Environ. Toxicol. Chem. 2003, 23, 583–590. [Google Scholar]
- Meinelt, T.; Schreckenbach, K.; Knopf, K.; Wienke, A.; Stüber, A.; Steinberg, C.E.W. Humic substances increase the constitution of swordtail (Xiphophorus helleri). Aquat. Sci. 2004, 66, 239–245. [Google Scholar]
- Höss, S.; Bergtold, M.; Haitzer, M.; Traunspurger, W.; Steinberg, C.E.W. Refractory dissolved organic matter can influence the reproduction of Caenorhabditis elegans (Nematoda). Freshw. Biol. 2001, 46, 1–10. [Google Scholar] [CrossRef]
- Timofeyev, M.A.; Wiegand, C.; Burnison, B.K.; Shatilina, Z.M.; Pflugmacher, S.; Steinberg, C.E.W. Direct impact of natural organic matter (NOM) on freshwater amphipods. Sci. Total Environ. 2004, 319, 115–121. [Google Scholar] [CrossRef]
- Kudryasheva, N. Bioluminescence and exogenous compounds: Physicochemical basis for bioluminescence assay. J. Photochem. Photobiol. 2006, 83, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.S. Nonspecific effects of exogenous compounds on bacterial bioluminescent enzymes: Fluorescence study. Curr. Enzym. Inhib. 2006, 2, 363–372. [Google Scholar] [CrossRef]
- Vetrova, E.V.; Kudryasheva, N.S.; Kratasyuk, V.A. Redox compounds influence on the NAD(P)H: FMN-oxidoreductase-luciferase bioluminescent system. Photochem. Photobiol. Sci. 2007, 6, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, L. Bioassays as a tool for evaluating advanced oxidation processes in water and wastewater treatment. Water Res. 2011, 45, 4311–4340. [Google Scholar] [CrossRef]
- Agrawal, A.; Kumari, S.; Sahu, K.K. Iron and copper recovery/removal from industrial wastes: A review. Ind. Eng. Chem. Res. 2009, 48, 6145–6161. [Google Scholar] [CrossRef]
- Paisio, C.E.; González, P.S.; Gerbaudo, A.; Bertuzzi, M.L.; Agostini, E. Toxicity of phenol solutions treated with rapeseed and tomato hairy roots. Desalination 2010, 263, 23–28. [Google Scholar] [CrossRef]
- Shourian, M.; Noghabi, K.A.; Zahiri, H.S.; Bagheri, T.; Karballaei, G.; Mollaei, M.; Rad, I.; Ahadi, S.; Raheb, J.; Abbasi, H. Efficient phenol degradation by a newly characterized Pseudomonas sp. SA01 isolated from pharmaceutical wastewaters. Desalination 2009, 246, 577–594. [Google Scholar] [CrossRef]
- Ren, S. Phenol mechanism of toxic action classification and prediction: A decision tree approach. Toxicol. Lett. 2003, 144, 313–323. [Google Scholar] [CrossRef]
- Tarasova, A.S.; Stom, D.I.; Kudryasheva, N.S. Bioluminescence as a tool for studying detoxification processes in metal salt solutions involving humic substances. Environ. Toxicol. Chem. 2011, 30, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Katafias, A.; Impert, O.; Kita, P. Hydrogen peroxide as a reductant of hexacyanoferrate (III) in alkaline solutions: Kinetic studies. Transit. Met. Chem. 2008, 33, 1041–1046. [Google Scholar] [CrossRef]
- Skogerboe, R.; Wilson, A. Reduction of ionic species by fulvic acid. Anal. Chem. 1981, 53, 228–232. [Google Scholar] [CrossRef]
- Matthiessen, A. Kinetic aspects of the reduction of mercury ions by humic substances. Fresenius J. Anal. Chem. 1996, 354, 747–749. [Google Scholar] [CrossRef] [PubMed]
- Tarasova, A.S.; Kislan, S.L.; Fedorova, E.S.; Kuznetsov, A.M.; Mogilnaya, O.A.; Stom, D.I.; Kudryasheva, N.S. Bioluminescence as a tool for studying detoxification processes in metal salt solutions involving humic substances. J. Photochem. Photobiol. B 2012, 117, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.S.; Tarasova, A.S. Pollutant toxicity and detoxification by humic substances: Mechanisms and quantitative assessment via luminescent biomonitoring. Environ. Sci. Pollut. Res. 2015, 22, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Deprez, K.; Robbens, J.; Nobels, I.; Vanparys, C.; Vanermen, G.; Tirez, K.; Michiels, L.; Weltens, R. DISCRISET: A battery of tests for fast waste classification—application of tests on waste extracts. Waste Manag. 2012, 32, 2218–2228. [Google Scholar] [CrossRef] [PubMed]
- Tigini, V.; Giansanti, P.; Mangiavillano, A.; Pannocchia, A.; Varese, G.C. Evaluation of toxicity, genotoxicity and environmental risk of simulated textile and tannery wastewaters with a battery of biotests. Ecotoxicol. Environ. Saf. 2011, 74, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.S.; Kratasyuk, V.; Esimbekova, E.N.; Vetrova, E.V.; Kudinova, I.Y.; Nemtseva, E.V. Development of the bioluminescent bioindicators for analyses of pollutions. Field Anal. Chem. Technol. 1998, 5, 277–280. [Google Scholar] [CrossRef]
- Bulich, A.A.; Isenberg, D.L. Use of the luminescent bacterial system for rapid assessment of aquatic toxicity. ISA Trans. 1981, 20, 29–33. [Google Scholar]
- Roda, A.; Guardigli, M.; Michelini, E.; Mirasoni, M. Bioluminescence in analytical chemistry and in vivo imaging. TrAC Trends Anal. Chem. 2009, 28, 307–322. [Google Scholar] [CrossRef]
- Roda, A.; Pasini, P.; Mirasoni, M.; Michchelini, E.; Guardigli, M. Biotechnological application of bioluminescence and chemiluminescence. Trends Biotechnol. 2004, 22, 295–303. [Google Scholar] [CrossRef]
- Girotti, S.; Ferri, E.N.; Fumo, M.G.; Maiolini, E. Monitoring of environmental pollutants by bioluminescent bacteria. Anal. Chim. Acta 2008, 608, 2–21. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Adil, M.; Ehtisham-ul-Haque, S.; Munir, B.; Yameen, M.; Ghaffar, A.; Shar, G.A.; Asif Tahir, M.; Iqbal, M. Vibrio fischeri bioluminescence inhibition assay for ecotoxicity assessment: A review. Sci. Total Environ. 2018, 626, 1295–1309. [Google Scholar] [CrossRef]
- Ivask, A.; Rolova, T.; Kahru, A. A suite of recombinant luminescent bacterial strains for the quantification of bioavailable heavy metals and toxicity testing. BMC Biotechnol. 2009, 9, 1–15. [Google Scholar] [CrossRef]
- Deryabin, D.G.; Aleshina, E.S. Natural and recombinant luminescent microorganisms in biotoxicity testing of mineral waters. Appl. Biochem. Microbiol. 2008, 44, 378–381. [Google Scholar] [CrossRef]
- Fedorova, E.; Kudryasheva, N.; Kuznetsov, A.; Mogil’naya, O.; Stom, D. Bioluminescent monitoring of detoxification processes: Activity of humic substances in quinone solutions. J. Photochem. Photobiol. 2007, 88, 131–136. [Google Scholar] [CrossRef]
- Rozhko, T.V.; Kudryasheva, N.S.; Kuznetsov, A.M.; Vydryakova, G.A.; Bondareva, L.G.; Bolsunovsky, A.Y. Effect of low-level α-radiation on bioluminescent assay systems of various complexity. Photochem. Photobiol. Sci. 2007, 6, 67–70. [Google Scholar] [CrossRef]
- Selivanova, M.A.; Mogilnaya, O.A.; Badun, G.A.; Vydryakova, G.A.; Kuznetsov, A.M.; Kudryasheva, N.S. Effect of tritium on luminousmarine bacteria and enzyme reactions. J. Environ. Radioact. 2013, 120, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Wang, X.C.; Ngo, H.H.; Guo, W.; Wu, M.N.; Wang, N. Bioassay based luminescent bacteria: Interferences, improvements, and applications. Sci. Total Environ. 2014, 468–469, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kudryasheva, N.S.; Kovel, E.S. Monitoring of Low-intensity Exposures via Luminescent Bioassays of Different Complexity: Cells, Enzyme Reactions and Fluorescent Proteins. Int. J. Mol. Sci. 2019, 20, 4451. [Google Scholar] [CrossRef]
- Kudryasheva, N.S.; Kovel, E.S.; Sachkova, A.S.; Vorobeva, A.A.; Isakova, V.G.; Churilov, G.N. Bioluminescent enzymatic assay as a tool for studying antioxidant activity and toxicity of bioactive compounds. J. Photochem. Photobiol. 2017, 93, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Kovel, E.S.; Sachkova, A.S.; Vnukova, N.G.; Churilov, G.N.; Knyazeva, E.M.; Kudryasheva, N.S. Antioxidant activity and toxicity of fullerenols via bioluminescence signaling: Role of oxygen substituents. Int. J. Mol. Sci. 2019, 20, 2324. [Google Scholar] [CrossRef]
- Bondarenko, L.S.; Kovel, E.S.; Kydralieva, K.A.; Dzhardimalieva, G.I.; Illé, E.; Tombácz, E.; Kicheeva, A.G.; Kudryasheva, N.S. Effects of modified magnetite nanoparticles on bacterial cells and enzyme reactions. Nanomaterials 2020, 10, 1499. [Google Scholar] [CrossRef] [PubMed]
- Sachkova, A.S.; Kovel, E.S.; Churilov, G.N.; Stom, D.I.; Kudryasheva, N.S. Biological activity of carbonic nano-structures—Comparison via enzymatic bioassay. J. Soils Sediments 2019, 19, 2689–2696. [Google Scholar] [CrossRef]
- Grabert, E.; Kossler, F. About the effects of nutrients on the luminescent bacteria test. In Bioluminescence and Chemiluminescence; Hastings, J.W., Kricka, L.J., Stanley, P.E., Eds.; John Wiley & Sons: Chichester, UK, 1997; pp. 291–294. [Google Scholar]
- Rozhko, T.; Bondareva, L.; Mogilnaya, O.; Vydryakova, G.; Bolsunovsky, A.; Stom, D.; Kudryasheva, N. Detoxification of Am-241 solutions by humic substances: Bioluminescent monitoring. Anal. Bioanal. Chem. 2011, 400, 29–334. [Google Scholar] [CrossRef]
- Reiller, P.; Moulin, V.; Dautel, C.; Casanova, F. Complexation of Th(IV) by humic, substances. In FZKA 6, 524, Wissenschaftliche Beerichte; Buckau, G., Ed.; Forschungszentrum Karlsruhe: Karlsruhe, Germany, 2000; pp. 121–129. [Google Scholar]
- Chopin, G.R. Role of humics in actinide behavior in ecosystems. In Chemical Separation Technologies and Relates Methods of Nuclear Waste Management; Chopin, G.R., Khankhasaev, M., Eds.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1999; pp. 247–260. [Google Scholar]
- Rozhko, T.; Kolesnik, O.; Badun, G.; Stom, D.; Kudryasheva, N. Humic substances mitigate the impact of tritium on luminous marine bacteria. Involvement of Reactive Oxygen Species. Int. J. Mol. Sci. 2020, 21, 6783. [Google Scholar] [CrossRef]
- Rozhko, T.; Nogovitsyna, E.; Badun, G.; Lukyanchuk, A.; Kudryasheva, N. Reactive oxygen species and low-dose effects of tritium on bacterial cells. J. Environ. Radioact. 2019, 208–209, 106035. [Google Scholar] [CrossRef]
- Rozhko, T.; Nemtseva, E.; Gardt, M.; Raikov, A.; Lisitsa, A.; Badun, G.; Kudryasheva, N. Enzymatic responses to low-intensity radiation of tritium. Int. J. Mol. Sci. 2020, 21, 8464. [Google Scholar] [CrossRef]
- Bondareva, L.; Fedorov, N. Accumulation of Am-241 by the submerged macrophyte Elodea canadensis and assessment of radionuclide toxicity with direct effects. In International Research in Environment, Geography and Earth Science; Department of Water Engineering, College of Agriculture, Isfahan University of Technology: Isfahan, Iran, 2020; Volume 3, pp. 78–91. [Google Scholar]
- Costerton, J.W. Structure and plasticity at various organization levels in the bacterial cell. Can. J. Microbiol. 1988, 34, 513–552. [Google Scholar] [CrossRef]
- Yakimenko, O.S.; Terekhova, V.A. Humic preparations and the assessment of their biological activity for certification purposes. Eurasian. Soil Sci. 2011, 44, 1222–1230. [Google Scholar] [CrossRef]
- Pukalchik, M.; Mercl, F.; Panova, M.; Břendová, K.; Terekhova, V.; Tlustoš, P. The improvement of multi-contaminated sandy loam soil chemical and biological properties by the biochar, wood ash, and humic substances amendments. Environ. Pollut. 2017, 229, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Pukalchik, M.; Panova, M.; Karpuhin, M.; Yakimenko, O.; Kydralieva, K.; Terekhova, V. Using humic products as amendments to restore Zn and Pb polluted soil: A case study using rapid screening phytotest endpoint. J. Soils Sediments 2018, 18, 750–761. [Google Scholar] [CrossRef]






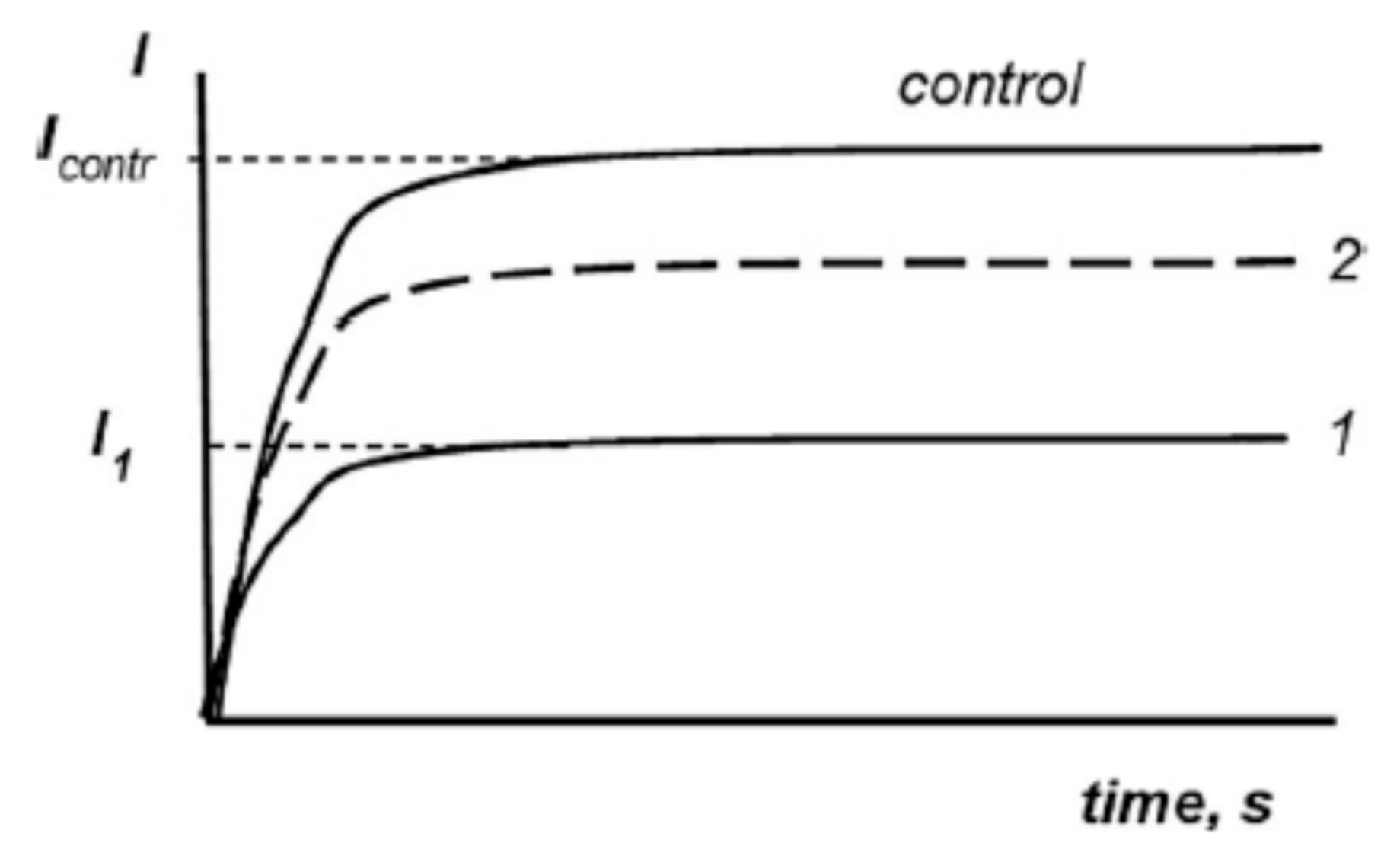{kind=link}
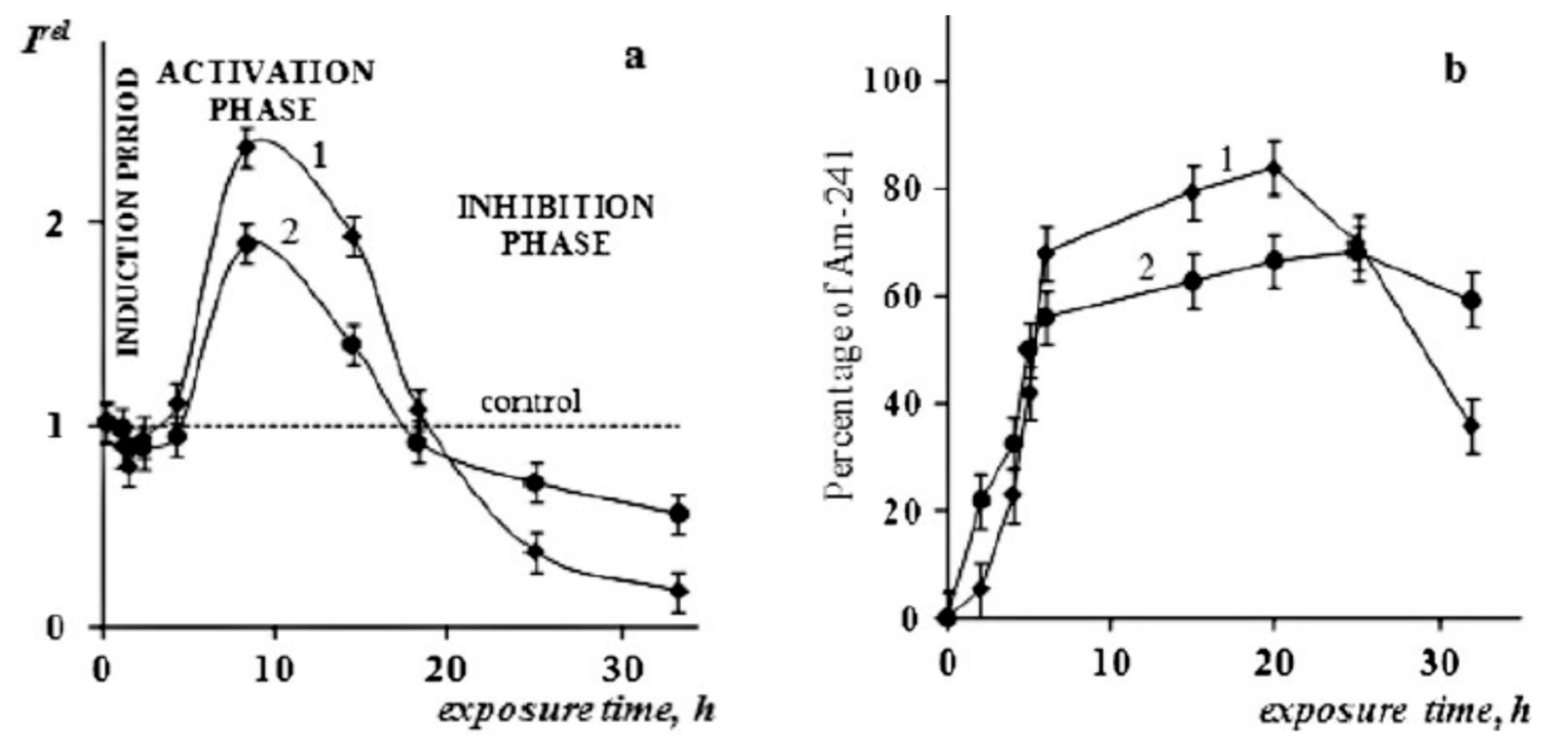{kind=link}
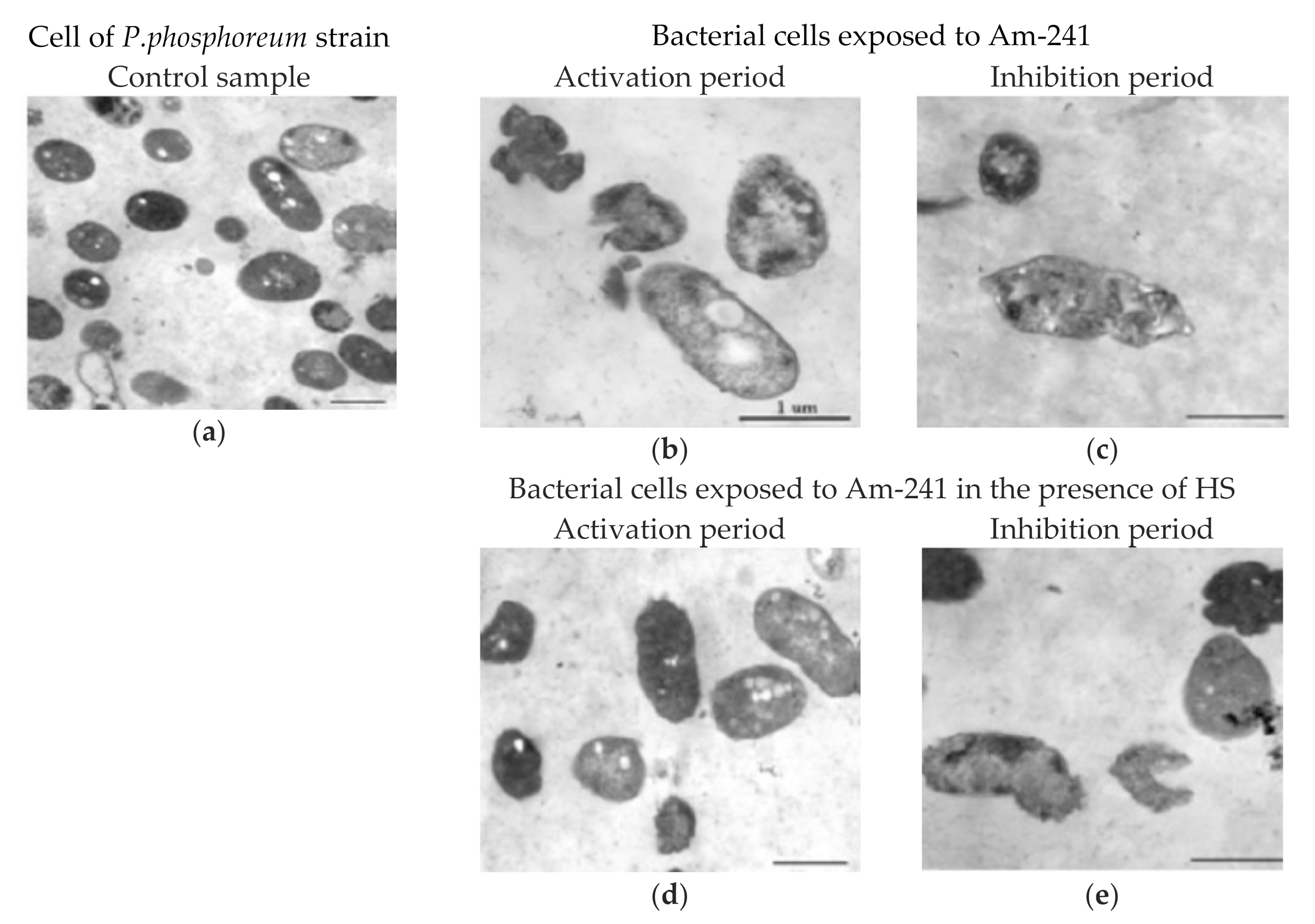{kind=link}
{kind=link}
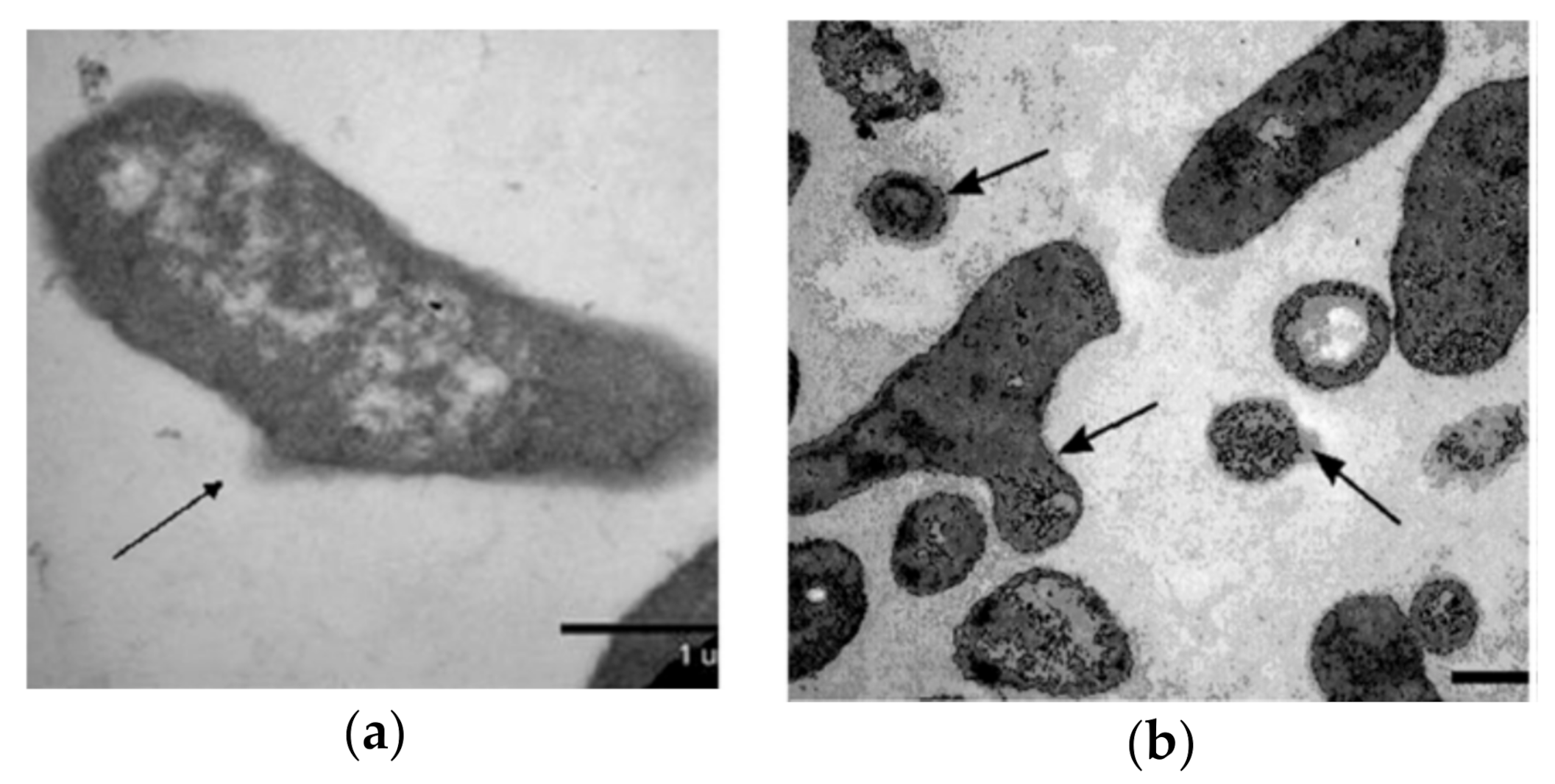{kind=link}
| Issue | Role of Humic Substances |
|---|---|
| Carbon cycling | Major C pool, transformations, transport and accumulation |
| Light penetration into waters | Absorption and attenuation of light by humic chromophores |
| Soil warming | Absorption of solar radiation by soil humic matter |
| Soil and water acidification | Binding of protons, aluminium and base cations in soils and water |
| Nutrient source | Reservoir of carbon, nitrogen, phosphorous, sulphur and chlorine |
| Nutrient control | Binding of iron and phosphate |
| Microbial metabolism | Substrate for microbes |
| Weathering | Enhancement of mineral dissolution rate |
| Soil formation (podzolisation) | Translocation of dissolved humic substances and associated metals (Al, Fe) |
| Properties of fine sediments | Adsorption at surfaces and alteration of colloidal properties |
| Soil structure | Aggregation effect on soil minerals solids |
| Photochemistry | Mediation of light-driven reactions |
| Heavy metals | Binding, transport, influence on bioavailability, redox reactions |
| Pesticides, xenobiotics | Binding, transport, influence on bioavailability |
| Radioactive waste disposal | Binding and transport of radionuclide ions in groundwaters |
| Ecosystem buffering | Control of proton and metal ions concentrations, persistence |
| Direct biological effect | Uptake and direct interaction with living organisms |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bondareva, L.; Kudryasheva, N. Direct and Indirect Detoxification Effects of Humic Substances. Agronomy 2021, 11, 198. https://doi.org/10.3390/agronomy11020198
Bondareva L, Kudryasheva N. Direct and Indirect Detoxification Effects of Humic Substances. Agronomy. 2021; 11(2):198. https://doi.org/10.3390/agronomy11020198
Chicago/Turabian StyleBondareva, Lydia, and Nadezhda Kudryasheva. 2021. "Direct and Indirect Detoxification Effects of Humic Substances" Agronomy 11, no. 2: 198. https://doi.org/10.3390/agronomy11020198
APA StyleBondareva, L., & Kudryasheva, N. (2021). Direct and Indirect Detoxification Effects of Humic Substances. Agronomy, 11(2), 198. https://doi.org/10.3390/agronomy11020198