Screening the FIGS Set of Lentil (Lens culinaris Medikus) Germplasm for Tolerance to Terminal Heat and Combined Drought-Heat Stress

 , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Study Area
2.2. Treatments
2.3. Investigation and Calculation of Agronomic Traits
2.4. Statistical Analysis
3. Results
3.1. Effects of Heat Stress on Morphological, Phenological, and Yield Contributing Traits
3.2. Correlations among the Traits under Heat Stress and Combined Heat-Drought Stress
3.3. Classification of Genotypes Based on Heat Tolerance Index
3.4. Response to Combined Heat-Drought Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

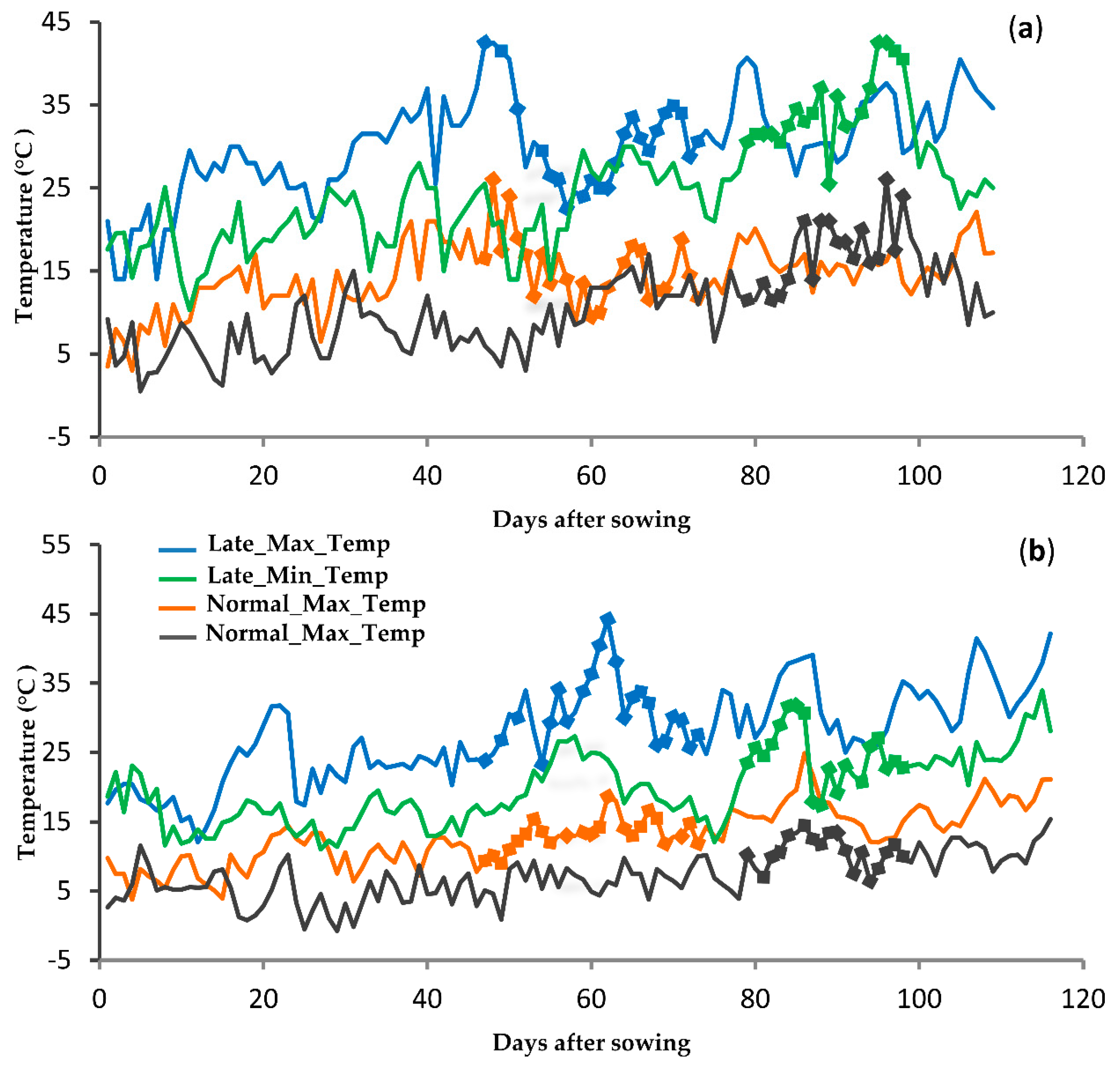{kind=link}
| Trait | Heat Stress Conditions | Heat-Drought Conditions | Normal Conditions | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tessaout | Marchouch | Tessaout | Marchouch | Tessaout | Marchouch | |||||||
| Min–Max | Mean ± SD | Min–Max | Mean ± SD | Min–Max | Mean ± SD | Min–Max | Mean ± SD | Min–Max | Mean ± SD | Min–Max | Mean ± SD | |
| PH | 15.00–28.0 | 20.32 ± 2.40 | 15.00–34.0 | 21.79 ± 3.13 | 12.75–28.90 | 17.67 ± 2.91 | 15.00–30.0 | 20.52 ± 2.83 | 17.00–37.0 | 24.95 ± 2.87 | 14.00–52 | 31.51 ± 6.09 |
| DF | 41.00–67.0 | 45.98 ± 4.28 | 47.00–73.0 | 54.58 ± 4.15 | 40.00–67.0 | 45.92 ± 4.70 | 48.50–73.0 | 54.62 ± 4.11 | 61.00–78.0 | 65.37 ± 1.90 | 79.00–98 | 87.62 ± 5.07 |
| DM | 80.00–103 | 88.21 ± 4.70 | 81.00–109 | 87.09 ± 4.82 | 77.00–101.0 | 82.97 ± 4.26 | 72.00–109.0 | 86.22 ± 4.39 | 100.00–119.0 | 115.63 ± 2.66 | 113.00–126 | 118.83 ± 2.48 |
| PBPP | 1.00–3.67 | 2.53 ± 0.58 | 1.00–4.67 | 2.35 ± 0.71 | 1.00–3.67 | 2.29 ± 0.59 | 1.00–5.00 | 1.81 ± 0.69 | 2.00–4.00 | 3.01 ± 0.68 | 1.00–5.00 | 2.59 ± 0.80 |
| SBPP | 2.00–17.10 | 9.42 ± 3.30 | 1.00–20.00 | 6.30 ± 3.31 | 2.00–16.50 | 8.02 ± 3.39 | 1.00–17.00 | 3.82 ± 2.62 | 4.00–25.00 | 13.10 ± 0.68 | 5.33–34.70 | 19.61 ± 5.98 |
| TBPP | 0.15–16.50 | 6.22 ± 3.43 | 0.20–8.90 | 2.54 ± 1.45 | 0.10–12.30 | 3.88 ± 3.22 | 0.00–9.00 | 1.22 ± 0.94 | 3.00–21.00 | 10.57 ± 2.79 | 1.00–30.70 | 14.34 ± 4.95 |
| NTPP | 2.29–126.0 | 29.88 ± 19.94 | 2.00–140.0 | 18.50 ± 15.89 | 1.00–70.75 | 21.22 ± 14.08 | 1.00–36.00 | 6.09 ± 3.79 | 6.00–160.45 | 56.68 ± 24.08 | 3.00–230.00 | 65.60 ± 40.69 |
| NUPP | 0.00–83.50 | 10.83 ± 9.13 | 0.00–38.0 | 3.44 ± 2.63 | 0.00–33.50 | 5.55 ± 3.47 | 0.00–14.00 | 1.82 ± 1.63 | 0.00–57.20 | 11.54 ± 7.65 | 0.00–82.67 | 17.11 ± 11.31 |
| NFPP | 0.50–96.20 | 19.06 ± 15.19 | 1.00–102.0 | 15.06 ± 12.11 | 0.00–62.75 | 15.66 ± 11.03 | 1.00–30.50 | 4.26 ± 2.13 | 2.00–129.45 | 45.14 ± 20.61 | 1.00–195.33 | 48.50 ± 35.15 |
| BPP | 0.25–15.80 | 3.99 ± 1.59 | 0.45–11.89 | 2.55 ± 1.92 | 0.50–32.21 | 3.72 ± 2.34 | 0.50–11.30 | 1.63 ± 1.03 | 1.40–39.60 | 12.86 ± 6.19 | 1.07–36.20 | 11.25 ± 4.08 |
| GYP | 0.13–3.94 | 0.88 ± 0.69 | 0.16–4.89 | 0.74 ± 0.68 | 0.12–2.78 | 0.66 ± 0.55 | 0.13–2.49 | 0.27 ± 0.20 | 0.15–7.71 | 2.74 ± 1.22 | 0.39–7.33 | 2.59 ± 1.49 |
| HSW | 0.90–5.00 | 2.50 ± 0.58 | 1.10–5.00 | 2.36 ± 0.59 | 0.70–4.20 | 1.94 ± 0.46 | 0.90–4.30 | 1.93 ± 0.52 | 0.50–3.70 | 1.54 ± 0.51 | 0.53–3.85 | 1.73 ± 0.62 |
| Trait | PH | DF | DM | PBPP | SBPP | TBPP | NTPP | NUPP | NFPP | BPP | GYP |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A. Under normal planting | |||||||||||
| DF | 0.156 ** | ||||||||||
| DM | 0.020ns | 0.141 * | |||||||||
| PBPP | −0.070ns | −0.067ns | −0.050ns | ||||||||
| SBPP | 0.242 ** | 0.152 ** | 0.015ns | −0.058ns | |||||||
| TBPP | 0.164 ** | 0.148 ** | 0.020ns | −0.060ns | 0.570 ** | ||||||
| NTPP | 0.080ns | −0.010ns | −0.055ns | 0.062ns | 0.428 ** | 0.372 ** | |||||
| NUPP | 0.090ns | −0.050ns | −0.132 * | 0.070ns | 0.199 ** | 0.163 ** | 0.521 ** | ||||
| NFPP | 0.058ns | 0.010ns | −0.010ns | 0.040ns | 0.409 ** | 0.360 ** | 0.931 ** | 0.174 ** | |||
| BPP | −0.008ns | 0.103ns | 0.079ns | −0.089ns | 0.086ns | 0.095ns | 0.131 * | 0.144 ** | 0.090ns | ||
| GYP | 0.056ns | 0.058ns | 0.011ns | 0.033ns | 0.400 ** | 0.396 ** | 0.873 ** | 0.148 ** | 0.944 ** | 0.112 * | |
| HSW | 0.119 * | 0.123 * | −0.120ns | −0.027ns | 0.084ns | 0.043ns | 0.061ns | 0.026ns | 0.059ns | −0.057ns | 0.070ns |
| B. Under heat stress condition | |||||||||||
| DF | 0.234 ** | ||||||||||
| DM | 0.146 ** | 0.363 ** | |||||||||
| PBPP | 0.070ns | 0.110ns | 0.090ns | ||||||||
| SBPP | 0.050ns | 0.05ns | 0.124 * | 0.147 ** | |||||||
| TBPP | 0.115 * | 0.030ns | 0.110ns | 0.226 ** | 0.657 ** | ||||||
| NTPP | 0.020ns | 0.010ns | 0.110ns | 0.129 * | 0.515 ** | 0.520 ** | |||||
| NUPP | 0.040ns | 0.040ns | 0.122 * | 0.127 * | 0.295 ** | 0.281 ** | 0.694 ** | ||||
| NFPP | 0.03ns | −0.020ns | 0.050ns | 0.090ns | 0.496 ** | 0.512 ** | 0.885 ** | 0.278 ** | |||
| BPP | 0.179 ** | 0.216 ** | 0.249 ** | 0.208 ** | 0.438 ** | 0.374 ** | 0.468 ** | 0.344 ** | 0.402 ** | ||
| GYP | 0.070ns | 0.020ns | 0.116 * | 0.110ns | 0.491 ** | 0.522 ** | 0.854 ** | 0.372 ** | 0.898 ** | 0.476 ** | |
| HSW | 0.182 ** | 0.139 * | 0.090ns | −0.010ns | −0.110ns | −0.030ns | −0.020ns | −0.060ns | 0.010ns | 0.040ns | 0.020ns |
| Trait | PH | DF | DM | PBPP | SBPP | TBPP | NTPP | NUPP | NFPP | BPP | GYP |
| C. Under heat-drought conditions | |||||||||||
| DF | 0.121 * | ||||||||||
| DM | 0.191 ** | 0.432 ** | |||||||||
| PBPP | 0.080ns | 0.010ns | 0.148 ** | ||||||||
| SBPP | 0.311 ** | 0.090ns | 0.203 ** | 0.135 * | |||||||
| TBPP | 0.210 ** | −0.080ns | 0.110ns | 0.060ns | 0.694 ** | ||||||
| NTPP | 0.272 ** | 0.145 ** | 0.224 ** | 0.174 ** | 0.665 ** | 0.645 ** | |||||
| NUPP | 0.184 ** | 0.230 ** | 0.145 ** | 0.040ns | 0.337 ** | 0.238 ** | 0.522 ** | ||||
| NFPP | 0.241 ** | 0.080ns | 0.200 ** | 0.185 ** | 0.631 ** | 0.647 ** | 0.943 ** | 0.209 ** | |||
| BPP | 0.356 ** | 0.391 ** | 0.344 ** | 0.182 ** | 0.460 ** | 0.294 ** | 0.435 ** | 0.378 ** | 0.352 ** | ||
| GYP | 0.278 ** | 0.121 * | 0.238 ** | 0.186 ** | 0.598 ** | 0.589 ** | 0.902 ** | 0.223 ** | 0.948 ** | 0.391 ** | |
| HSW | 0.086ns | 0.159 ** | 0.144 ** | 0.019ns | −0.154 ** | −0.135 * | 0.020ns | 0.111 * | −0.023ns | 0.073ns | 0.014ns |
| PH | DF | DM | PBPP | SBPP | TBPP | NTPP | NUPP | NFPP | BPP | GYP | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A. Under normal planting | |||||||||||
| DF | 0.205 ** | ||||||||||
| DM | 0.321 ** | 0.462 ** | |||||||||
| PBPP | 0.043ns | −0.010ns | −0.008ns | ||||||||
| SBPP | 0.053ns | 0.022ns | 0.034ns | 0.032ns | |||||||
| TBPP | 0.142 * | −0.015ns | 0.057ns | 0.034ns | 0.670 ** | ||||||
| NTPP | −0.012ns | −0.086ns | 0.068ns | 0.073ns | 0.272 ** | 0.222 ** | |||||
| NUPP | −0.087ns | −0.064ns | −0.005ns | −0.052ns | 0.295 ** | 0.287 ** | 0.545 ** | ||||
| NFPP | 0.018ns | −0.076ns | 0.080ns | 0.110ns | 0.207 ** | 0.152 ** | 0.954 ** | 0.268 ** | |||
| BPP | 0.143 * | 0.001ns | 0.074ns | 0.114 * | 0.517 ** | 0.472 ** | 0.406 ** | 0.348 ** | 0.342 ** | ||
| GYP | 0.017ns | −0.126 * | 0.060ns | 0.090ns | 0.177 ** | 0.110ns | 0.861 ** | 0.228 ** | 0.908 ** | 0.302 ** | |
| HSW | 0.221 ** | −0.050ns | 0.030ns | 0.144 ** | 0.020ns | 0.050ns | −0.110ns | −0.129 * | −0.078ns | 0.146 ** | −0.060ns |
| B. Under heat stress condition | |||||||||||
| DF | 0.163 ** | ||||||||||
| DM | 0.358 ** | 0.619 ** | |||||||||
| PBPP | 0.030ns | 0.040ns | 0.150 ** | ||||||||
| SBPP | 0.148 ** | 0.054ns | 0.148 ** | 0.405 ** | |||||||
| TBPP | 0.069ns | 0.071ns | 0.073ns | 0.277 ** | 0.674 ** | ||||||
| NTPP | 0.223 ** | 0.030ns | 0.115 * | 0.357 ** | 0.531 ** | 0.431 ** | |||||
| NUPP | 0.118 * | 0.139 * | 0.257 ** | 0.287 ** | 0.407 ** | 0.307 ** | 0.684 ** | ||||
| NFPP | 0.228 ** | −0.010ns | 0.060ns | 0.335 ** | 0.504 ** | 0.417 ** | 0.975 ** | 0.504 ** | |||
| BPP | 0.293 ** | 0.251 ** | 0.341 ** | 0.360 ** | 0.535 ** | 0.445 ** | 0.648 ** | 0.616 ** | 0.579 ** | ||
| GYP | 0.248 ** | −0.023ns | 0.040ns | 0.314 ** | 0.511 ** | 0.422 ** | 0.931 ** | 0.511 ** | 0.946 ** | 0.584 ** | |
| HSW | 0.208 ** | 0.190 ** | 0.269 ** | 0.227 ** | 0.114 * | 0.103ns | 0.092ns | 0.145 ** | 0.065ns | 0.284 ** | 0.110ns |
| Trait | PH | DF | DM | PBPP | SBPP | TBPP | NTPP | NUPP | NFPP | BPP | GYP |
| C. Under heat-drought condition | |||||||||||
| DF | 0.092ns | ||||||||||
| DM | 0.125 * | 0.561 ** | |||||||||
| PBPP | 0.224 ** | −0.044ns | 0.118* | ||||||||
| SBPP | 0.180 ** | 0.041ns | 0.198 ** | 0.586 ** | |||||||
| TBPP | 0.115* | −0.010ns | 0.115* | 0.357 ** | 0.537 ** | ||||||
| NTPP | 0.188 ** | −0.010ns | 0.153 ** | 0.535 ** | 0.463 ** | 0.419 ** | |||||
| NUPP | 0.139* | −0.042ns | 0.060ns | 0.489 ** | 0.426 ** | 0.441 ** | 0.769 ** | ||||
| NFPP | 0.175 ** | 0.013ns | 0.177 ** | 0.439 ** | 0.379 ** | 0.307 ** | 0.913 ** | 0.443 ** | |||
| BPP | 0.237 ** | 0.070ns | 0.223 ** | 0.335 ** | 0.424 ** | 0.464 ** | 0.436 ** | 0.350 ** | 0.389 ** | ||
| GYP | 0.190 ** | 0.020ns | 0.168 ** | 0.417 ** | 0.402 ** | 0.371 ** | 0.890 ** | 0.482 ** | 0.942 ** | 0.408 ** | |
| HSW | 0.194 ** | 0.152 ** | 0.169 ** | 0.158 ** | 0.204 ** | 0.179 ** | 0.139 * | 0.139 * | 0.107ns | 0.228 ** | 0.123 * |
| Accession | DF | DM | Ys | Yp | HTI | Accession | DF | DM | Ys | Yp | HTI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Tessoaut Location | Marchouch Location | ||||||||||
| ILL 6359 | 47.00 | 89.00 | 1.96 | 2.96 | 0.94 | ILL 6363 | 54.00 | 85.00 | 2.38 | 3.91 | 0.97 |
| ILL 7295 | 46.00 | 88.50 | 1.91 | 2.71 | 0.94 | ILL 8025 | 54.50 | 86.50 | 1.91 | 1.70 | 0.87 |
| ILL 2524 | 43.00 | 87.00 | 1.95 | 2.91 | 0.94 | ILL 7819 | 51.50 | 84.50 | 2.00 | 2.43 | 0.85 |
| ILL 6053 | 42.50 | 88.50 | 1.68 | 1.66 | 0.90 | ILL 7223 | 51.50 | 88.00 | 2.05 | 3.80 | 0.75 |
| ILL 7389 | 46.00 | 88.00 | 2.00 | 3.57 | 0.88 | ILL 6361 | 53.00 | 85.50 | 2.10 | 4.21 | 0.74 |
| ILL 8019 | 43.00 | 83.50 | 1.93 | 3.14 | 0.88 | ILL 6053 | 52.50 | 85.00 | 1.91 | 3.13 | 0.72 |
| ILL 8018 | 45.50 | 88.50 | 1.80 | 2.60 | 0.86 | ILL 6075 | 55.00 | 86.00 | 1.71 | 2.15 | 0.69 |
| ILL 6094 | 45.00 | 95.00 | 1.93 | 3.41 | 0.84 | ILL 4902 | 68.00 | 90.00 | 1.78 | 3.70 | 0.64 |
| ILL 8025 | 44.50 | 87.00 | 1.88 | 3.16 | 0.84 | ILL 8061 | 61.50 | 93.00 | 1.42 | 1.25 | 0.61 |
| ILL 6095 | 45.00 | 90.00 | 1.46 | 1.02 | 0.82 | ILL 6362 | 57.00 | 88.00 | 2.01 | 5.30 | 0.59 |
| ILL 4605 | 50.84 | 84.59 | 1.95 | 3.81 | 0.81 | ILL 7806 | 55.50 | 88.00 | 1.59 | 2.65 | 0.57 |
| ILL 7932 | 44.00 | 84.50 | 1.21 | 0.76 | 0.64 | ILL 6359 | 51.00 | 86.50 | 1.27 | 0.90 | 0.51 |
| ILL 6364 | 44.50 | 86.50 | 1.74 | 3.64 | 0.62 | ILL 880 | 70.50 | 100.00 | 1.53 | 3.66 | 0.48 |
| ILL 5919 | 50.00 | 81.00 | 1.32 | 1.77 | 0.59 | ILL 504 | 67.00 | 106.00 | 1.38 | 2.80 | 0.45 |
| ILL 7250 | 44.00 | 90.00 | 1.39 | 2.16 | 0.56 | ||||||
| ILL 7796 | 48.50 | 88.00 | 1.64 | 3.65 | 0.55 | ||||||
| ILL 7817 | 43.00 | 85.00 | 1.75 | 4.12 | 0.55 | ||||||
| ILL 6055 | 47.00 | 86.50 | 1.42 | 2.65 | 0.51 | ||||||
| ILL 8437 | 46.50 | 97.00 | 1.27 | 1.93 | 0.50 | ||||||
| ILL 6080 | 45.50 | 84.00 | 1.08 | 0.99 | 0.48 | ||||||
| ILL 8023 | 44.50 | 91.00 | 1.35 | 2.46 | 0.47 | ||||||
| ILL 7795 | 44.00 | 88.00 | 1.27 | 2.16 | 0.45 | ||||||
| ILL 6107 | 44.50 | 89.50 | 1.46 | 3.26 | 0.43 | ||||||
| ILL 7808 | 44.00 | 85.00 | 1.35 | 2.82 | 0.41 | ||||||
| ILL 7301 | 46.00 | 85.00 | 1.25 | 2.32 | 0.41 | ||||||
| ILL 7327 | 44.50 | 90.00 | 0.96 | 1.00 | 0.37 | ||||||
| Accession | DF | DM | Ys | Yp | HTI | Accession | DF | DM | Ys | Yp | HTI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Marchouch Station | Tessaout Station | ||||||||||
| Moderately heat tolerant | Moderately heat tolerant | ||||||||||
| ILL 7308 | 53.00 | 86.00 | 1.52 | 2.82 | 0.49 | ILL 206 | 44.50 | 92.00 | 1.21 | 3.68 | 0.14 |
| ILL 8029 | 55.00 | 85.00 | 1.43 | 2.76 | 0.45 | ILL 221 | 62.50 | 98.00 | 1.31 | 4.68 | 0.12 |
| ILL 3635 | 54.00 | 85.50 | 1.31 | 2.12 | 0.43 | ILL 1734 | 50.50 | 96.50 | 0.93 | 1.59 | 0.26 |
| ILL 7807 | 52.50 | 92.00 | 1.36 | 2.99 | 0.37 | ILL 1861 | 42.00 | 95.00 | 0.85 | 2.15 | 0.06 |
| ILL 7301 | 54.00 | 85.00 | 1.25 | 2.47 | 0.35 | ILL 3484 | 46.50 | 94.50 | 1.10 | 2.11 | 0.31 |
| ILL 7798 | 53.50 | 86.00 | 0.99 | 0.91 | 0.33 | ILL 3635 | 45.00 | 83.50 | 1.30 | 3.64 | 0.23 |
| ILL 7295 | 53.00 | 86.00 | 1.15 | 2.13 | 0.32 | ILL 4743 | 46.50 | 86.00 | 0.77 | 2.71 | −0.09 |
| ILL 6325 | 50.50 | 90.00 | 1.16 | 2.28 | 0.30 | ILL 4772 | 43.50 | 89.50 | 0.70 | 2.01 | −0.04 |
| ILL 6095 | 53.50 | 84.50 | 0.87 | 0.55 | 0.28 | ILL 4902 | 65.00 | 101.00 | 0.99 | 2.76 | 0.17 |
| ILL 729 | 66.50 | 106.00 | 1.24 | 3.78 | 0.26 | ILL 5918 | 46.50 | 81.50 | 0.68 | 1.72 | 0.00 |
| ILL 6086 | 53.00 | 87.00 | 1.07 | 2.22 | 0.25 | ILL 5929 | 51.00 | 90.00 | 1.04 | 2.77 | 0.16 |
| ILL 6364 | 51.00 | 81.00 | 1.20 | 3.16 | 0.24 | ILL 5958 | 45.00 | 89.50 | 0.69 | 1.53 | 0.04 |
| ILL 7286 | 53.50 | 85.50 | 0.85 | 0.90 | 0.24 | ILL 6059 | 51.00 | 88.50 | 0.81 | 2.48 | 0.00 |
| ILL 6337 | 51.00 | 87.00 | 1.30 | 3.89 | 0.24 | ILL 6074 | 46.00 | 88.00 | 0.61 | 1.13 | 0.04 |
| ILL 6094 | 53.50 | 85.00 | 1.01 | 2.03 | 0.23 | ILL 6075 | 43.00 | 89.00 | 1.22 | 4.40 | 0.02 |
| ILL 7830 | 54.50 | 85.50 | 0.90 | 1.38 | 0.23 | ILL 6077 | 48.50 | 88.50 | 0.79 | 1.21 | 0.19 |
| ILL 5958 | 53.50 | 84.50 | 0.92 | 1.75 | 0.20 | ILL 6079 | 45.00 | 84.50 | 0.78 | 2.41 | −0.03 |
| ILL 8019 | 53.00 | 85.50 | 1.08 | 2.87 | 0.20 | ILL 6088 | 43.50 | 85.00 | 0.93 | 1.94 | 0.18 |
| ILL 7344 | 54.00 | 87.00 | 1.12 | 3.33 | 0.18 | ILL 6092 | 46.00 | 86.50 | 0.81 | 1.03 | 0.23 |
| ILL 7309 | 52.00 | 85.00 | 0.71 | 0.70 | 0.16 | ILL 6096 | 44.50 | 90.50 | 1.09 | 3.11 | 0.13 |
| ILL 8017 | 54.50 | 87.00 | 1.20 | 4.14 | 0.16 | ILL 6101 | 45.50 | 88.50 | 1.23 | 3.11 | 0.25 |
| ILL 7250 | 53.00 | 87.50 | 0.87 | 2.05 | 0.14 | ILL 6102 | 45.00 | 90.50 | 1.05 | 2.96 | 0.11 |
| ILL 7238 | 53.50 | 86.00 | 0.80 | 1.60 | 0.14 | ILL 6325 | 44.50 | 94.00 | 0.74 | 1.83 | 0.03 |
| ILL 7300 | 57.50 | 88.00 | 0.82 | 2.00 | 0.13 | ILL 6337 | 43.50 | 88.00 | 0.75 | 1.71 | 0.05 |
| ILL 1861 | 52.50 | 100.00 | 0.56 | 0.50 | 0.08 | ILL 6346 | 46.00 | 90.50 | 0.90 | 2.38 | 0.09 |
| ILL 7380 | 53.00 | 86.00 | 0.75 | 2.15 | 0.05 | ILL 6356 | 45.50 | 87.50 | 1.17 | 3.26 | 0.17 |
| Heat sensitive | ILL 6361 | 45.00 | 86.50 | 1.01 | 2.25 | 0.20 | |||||
| ILL 7290 | 51.00 | 83.50 | 0.71 | 1.14 | 0.11 | ILL 6363 | 47.00 | 85.00 | 0.80 | 2.56 | −0.03 |
| ILL 8056 | 62.00 | 93.00 | 0.68 | 1.43 | 0.10 | ILL 6385 | 60.50 | 91.00 | 0.94 | 2.24 | 0.20 |
| ILL 7264 | 53.00 | 87.00 | 0.60 | 0.67 | 0.09 | ILL 7223 | 45.00 | 89.00 | 1.16 | 2.40 | 0.31 |
| ILL 7312 | 51.00 | 85.50 | 0.70 | 1.35 | 0.09 | ILL 7290 | 45.50 | 88.50 | 0.87 | 2.96 | −0.05 |
| ILL 6057 | 52.00 | 88.00 | 0.63 | 0.95 | 0.08 | ILL 7303 | 54.00 | 98.00 | 0.88 | 3.22 | −0.05 |
| ILL 7305 | 52.00 | 87.00 | 0.78 | 2.00 | 0.08 | ILL 7304 | 44.50 | 86.50 | 0.70 | 2.12 | −0.06 |
| ILL 7824 | 53.50 | 85.50 | 1.10 | 4.24 | 0.08 | ILL 7306 | 43.50 | 92.00 | 1.10 | 2.10 | 0.31 |
| ILL 6091 | 55.00 | 86.00 | 0.68 | 1.46 | 0.08 | ILL 7308 | 44.50 | 83.50 | 0.74 | 1.91 | 0.02 |
| ILL 4743 | 49.50 | 86.00 | 0.71 | 1.47 | 0.08 | ILL 7309 | 46.50 | 84.00 | 0.92 | 2.00 | 0.17 |
| ILL 7310 | 54.00 | 85.00 | 0.60 | 0.90 | 0.07 | ILL 7312 | 44.00 | 84.50 | 0.68 | 2.06 | −0.07 |
| ILL 6088 | 51.50 | 85.00 | 0.80 | 2.45 | 0.05 | ILL 7313 | 44.50 | 87.00 | 0.80 | 1.72 | 0.10 |
| ILL 7831 | 53.00 | 86.00 | 0.82 | 2.62 | 0.05 | ILL 7316 | 44.50 | 85.00 | 1.31 | 4.55 | 0.08 |
| ILL 8016 | 53.00 | 85.00 | 0.62 | 1.30 | 0.04 | ILL 7326 | 44.50 | 87.00 | 0.93 | 2.45 | 0.09 |
| ILL 8012 | 54.00 | 85.00 | 0.75 | 2.24 | 0.04 | ILL 7380 | 44.50 | 91.50 | 0.76 | 2.51 | −0.07 |
| ILL 6322 | 50.00 | 84.00 | 0.94 | 3.50 | 0.03 | ILL 7383 | 46.50 | 87.00 | 1.04 | 3.31 | 0.05 |
| ILL 7316 | 53.00 | 87.00 | 0.72 | 2.10 | 0.03 | ILL 7798 | 45.50 | 91.50 | 1.05 | 2.81 | 0.14 |
| ILL 6087 | 51.00 | 85.00 | 0.65 | 1.63 | 0.02 | ILL 7799 | 43.00 | 88.00 | 1.43 | 4.00 | 0.27 |
| ILL 8022 | 53.50 | 86.00 | 0.62 | 1.55 | 0.02 | ILL 7801 | 45.00 | 85.50 | 0.43 | 0.86 | −0.09 |
| ILL 206 | 54.00 | 86.00 | 0.69 | 2.11 | 0.02 | ILL 7806 | 44.50 | 90.50 | 0.90 | 0.93 | 0.33 |
| ILL 6077 | 54.50 | 85.00 | 0.79 | 2.99 | 0.00 | ILL 7807 | 45.00 | 86.50 | 1.20 | 4.86 | −0.07 |
| ILL 7826 | 58.00 | 90.00 | 0.56 | 1.59 | −0.01 | ILL 7818 | 43.50 | 87.00 | 1.08 | 2.95 | 0.14 |
| ILL 6079 | 58.00 | 87.00 | 0.49 | 1.25 | −0.02 | ILL 7826 | 49.50 | 87.50 | 0.81 | 1.87 | 0.10 |
| ILL 7796 | 54.00 | 88.00 | 0.44 | 0.77 | −0.02 | ILL 7827 | 43.50 | 88.50 | 0.93 | 2.73 | 0.05 |
| ILL 8021 | 54.50 | 85.00 | 0.40 | 0.67 | −0.04 | ILL 7831 | 45.00 | 85.50 | 0.85 | 2.84 | −0.04 |
| ILL 8026 | 52.50 | 85.00 | 0.85 | 3.69 | −0.04 | ILL 8013 | 46.50 | 90.00 | 1.30 | 4.18 | 0.14 |
| ILL 7303 | 58.50 | 104.50 | 0.92 | 4.40 | −0.04 | ILL 8014 | 48.00 | 93.50 | 0.96 | 1.90 | 0.23 |
| ILL 4758 | 47.50 | 83.00 | 0.78 | 3.10 | −0.05 | ILL 8016 | 44.00 | 83.50 | 1.04 | 2.13 | 0.24 |
| ILL 7795 | 55.00 | 86.00 | 0.37 | 0.60 | −0.05 | ILL 8020 | 45.00 | 82.00 | 0.95 | 2.16 | 0.17 |
| ILL 221 | 55.00 | 88.00 | 0.65 | 2.52 | −0.05 | ILL 8024 | 44.50 | 85.00 | 1.06 | 2.83 | 0.15 |
| ILL 7339 | 54.00 | 87.50 | 0.52 | 1.73 | −0.06 | ILL 8026 | 45.00 | 85.50 | 0.97 | 3.43 | −0.04 |
| ILL 8015 | 53.50 | 85.00 | 0.46 | 1.34 | −0.06 | ILL 8028 | 43.50 | 87.00 | 1.11 | 3.51 | 0.07 |
| ILL 7836 | 52.00 | 84.00 | 0.70 | 2.95 | −0.07 | ILL 8056 | 47.50 | 95.00 | 0.94 | 3.01 | 0.02 |
| ILL 7389 | 54.50 | 84.50 | 0.92 | 4.55 | −0.07 | ILL 8061 | 60.50 | 93.00 | 1.34 | 3.76 | 0.30 |
| ILL 6074 | 53.00 | 86.00 | 0.40 | 1.00 | −0.07 | Heat sensitive | |||||
| ILL 7306 | 54.00 | 86.00 | 0.70 | 3.13 | −0.08 | ILL 247 | 43.00 | 88.50 | 0.58 | 2.01 | −0.15 |
| ILL 1734 | 54.50 | 94.50 | 0.56 | 2.23 | −0.08 | ILL 5505 | 49.50 | 85.00 | 0.64 | 2.46 | −0.15 |
| ILL 5918 | 51.00 | 83.00 | 0.38 | 0.95 | −0.09 | ILL 5957 | 47.00 | 80.00 | 0.28 | 0.58 | −0.17 |
| ILL 7827 | 53.00 | 85.00 | 0.66 | 2.96 | −0.09 | ILL 5964 | 43.50 | 85.00 | 0.32 | 0.94 | −0.20 |
| ILL 7325 | 51.00 | 81.00 | 0.38 | 1.10 | −0.10 | ILL 6057 | 44.00 | 88.50 | 0.13 | 1.01 | −0.39 |
| ILL 6059 | 65.00 | 98.00 | 0.32 | 1.35 | −0.12 | ILL 6058 | 42.50 | 86.50 | 0.16 | 1.05 | −0.37 |
| ILL 6099 | 66.50 | 95.00 | 0.17 | 0.39 | −0.12 | ILL 6060 | 46.00 | 87.00 | 0.50 | 2.23 | −0.25 |
| ILL 5940 | 54.00 | 87.00 | 0.37 | 1.30 | −0.12 | ILL 6086 | 45.00 | 87.00 | 0.59 | 2.97 | −0.30 |
| ILL 8437 | 52.00 | 102.50 | 0.76 | 3.93 | −0.12 | ILL 6087 | 45.00 | 83.50 | 0.55 | 2.76 | −0.30 |
| ILL 6058 | 65.00 | 98.00 | 0.23 | 0.80 | −0.12 | ILL 6099 | 46.00 | 89.00 | 0.42 | 1.58 | −0.22 |
| ILL 6319 | 50.00 | 82.50 | 0.80 | 4.23 | −0.13 | ILL 6104 | 44.00 | 87.50 | 0.43 | 1.92 | −0.27 |
| ILL 7818 | 54.50 | 86.00 | 0.49 | 2.31 | −0.14 | ILL 6105 | 45.50 | 85.00 | 0.84 | 3.41 | −0.15 |
| ILL 6096 | 55.00 | 84.00 | 0.35 | 1.38 | −0.14 | ILL 6320 | 43.00 | 85.50 | 0.75 | 3.26 | −0.22 |
| ILL 5957 | 48.00 | 81.00 | 0.51 | 2.24 | −0.14 | ILL 6332 | 51.00 | 92.00 | 0.37 | 2.31 | −0.37 |
| ILL 4605 | 49.34 | 82.65 | 0.77 | 4.07 | −0.14 | ILL 6338 | 47.00 | 91.00 | 0.59 | 1.96 | −0.12 |
| ILL 3484 | 51.50 | 85.00 | 0.57 | 2.82 | −0.14 | ILL 6360 | 44.50 | 87.50 | 0.71 | 3.37 | −0.27 |
| ILL 7289 | 56.00 | 89.00 | 0.33 | 1.40 | −0.15 | ILL 7238 | 46.00 | 96.50 | 0.96 | 4.91 | −0.30 |
| ILL 7817 | 55.00 | 85.00 | 0.21 | 0.59 | −0.15 | ILL 7264 | 45.00 | 87.00 | 0.84 | 3.56 | −0.18 |
| ILL 6100 | 52.00 | 84.00 | 0.50 | 2.45 | −0.15 | ILL 7286 | 44.00 | 88.00 | 0.69 | 3.16 | −0.25 |
| ILL 7304 | 53.50 | 85.00 | 0.37 | 1.70 | −0.16 | ILL 7310 | 43.00 | 86.50 | 0.40 | 2.49 | −0.40 |
| ILL 7327 | 53.00 | 85.00 | 0.45 | 2.25 | −0.17 | ILL 7311 | 50.00 | 89.50 | 0.40 | 2.00 | −0.29 |
| ILL 7307 | 54.00 | 85.00 | 0.41 | 2.07 | −0.17 | ILL 7314 | 44.00 | 85.00 | 0.47 | 2.03 | −0.26 |
| ILL 247 | 54.50 | 88.00 | 0.26 | 1.10 | −0.17 | ILL 7317 | 45.50 | 90.00 | 0.66 | 2.94 | −0.23 |
| ILL 7812 | 53.00 | 85.00 | 0.50 | 2.68 | −0.17 | ILL 7325 | 44.50 | 90.00 | 0.53 | 1.66 | −0.13 |
| ILL 8013 | 54.00 | 95.50 | 0.81 | 4.85 | −0.17 | ILL 7328 | 54.50 | 95.00 | 1.08 | 4.69 | −0.12 |
| ILL 6356 | 51.50 | 84.50 | 0.86 | 5.10 | −0.17 | ILL 7344 | 44.00 | 87.00 | 0.63 | 3.17 | −0.31 |
| ILL 6332 | 73.00 | 98.00 | 0.19 | 1.31 | −0.17 | ILL 7813 | 49.50 | 84.50 | 0.55 | 2.81 | −0.29 |
| ILL 5955 | 56.00 | 87.00 | 0.20 | 0.80 | −0.18 | ILL 7815 | 48.00 | 93.00 | 0.61 | 2.43 | −0.18 |
| ILL 7838 | 50.50 | 83.50 | 0.32 | 1.39 | −0.18 | ILL 7816 | 44.00 | 92.50 | 0.75 | 3.03 | −0.17 |
| ILL 8028 | 58.00 | 90.00 | 0.52 | 3.08 | −0.18 | ILL 7819 | 43.50 | 86.00 | 0.41 | 1.19 | −0.16 |
| ILL 7328 | 65.00 | 90.00 | 0.24 | 1.42 | −0.18 | ILL 7820 | 44.50 | 87.50 | 0.80 | 3.03 | −0.12 |
| ILL 6055 | 50.50 | 84.00 | 0.56 | 3.10 | −0.18 | ILL 7830 | 44.50 | 86.00 | 0.67 | 3.78 | −0.37 |
| ILL 6092 | 58.00 | 89.00 | 0.41 | 2.38 | −0.18 | ILL 7836 | 45.50 | 84.50 | 0.26 | 0.54 | −0.19 |
| ILL 6360 | 51.00 | 83.00 | 0.42 | 2.20 | −0.18 | ILL 7837 | 42.50 | 82.50 | 0.66 | 2.71 | −0.20 |
| ILL 8018 | 52.00 | 82.00 | 0.44 | 2.55 | −0.20 | ILL 8015 | 48.00 | 84.50 | 0.68 | 4.01 | −0.39 |
| ILL 7311 | 57.00 | 86.00 | 0.25 | 1.48 | −0.21 | ILL 8017 | 45.00 | 84.00 | 0.55 | 2.71 | −0.30 |
| ILL 6102 | 54.00 | 88.00 | 0.22 | 1.17 | −0.21 | ILL 8021 | 45.00 | 87.00 | 0.60 | 2.97 | −0.30 |
| ILL 7313 | 53.00 | 87.00 | 0.30 | 1.70 | −0.21 | ILL 8054 | 44.50 | 88.00 | 0.61 | 3.49 | −0.38 |
| ILL 8023 | 51.50 | 85.00 | 0.61 | 3.85 | −0.22 | ILL 8280 | 50.00 | 91.00 | 0.22 | 1.66 | −0.40 |
| ILL 8054 | 52.50 | 82.50 | 0.24 | 1.41 | −0.22 | Highly heat sensitive | |||||
| ILL 6385 | 70.00 | 89.00 | 0.21 | 1.94 | −0.23 | ILL 504 | 66.50 | 93.50 | 0.18 | 2.22 | −0.47 |
| ILL 6080 | 54.00 | 84.00 | 0.35 | 2.36 | −0.24 | ILL 729 | 63.50 | 98.50 | 0.66 | 4.83 | −0.49 |
| Highly heat sensitive | ILL 880 | 45.50 | 86.50 | 0.23 | 3.11 | −0.65 | |||||
| ILL 5919 | 51.50 | 83.00 | 0.52 | 3.61 | −0.26 | ILL 4758 | 42.00 | 89.00 | 0.31 | 2.91 | −0.56 |
| ILL 5929 | 55.00 | 88.00 | 0.18 | 1.60 | −0.27 | ILL 4910 | 43.50 | 95.00 | 0.29 | 3.36 | −0.65 |
| ILL 8014 | 61.00 | 88.50 | 0.30 | 2.70 | −0.28 | ILL 5940 | 47.00 | 93.00 | 0.42 | 3.56 | −0.55 |
| ILL 5964 | 55.00 | 85.00 | 0.27 | 2.28 | −0.28 | ILL 5943 | 46.00 | 88.50 | 0.31 | 2.96 | −0.55 |
| ILL 7837 | 54.50 | 86.50 | 0.61 | 4.58 | −0.28 | ILL 5955 | 43.50 | 85.50 | 0.19 | 1.81 | −0.47 |
| ILL 7317 | 55.50 | 89.00 | 0.17 | 1.71 | −0.29 | ILL 6054 | 44.50 | 86.50 | 0.22 | 2.48 | −0.56 |
| ILL 7266 | 56.50 | 89.00 | 0.23 | 2.20 | −0.29 | ILL 6072 | 46.00 | 88.00 | 0.33 | 3.68 | −0.66 |
| ILL 8020 | 54.00 | 86.00 | 0.40 | 3.27 | −0.29 | ILL 6081 | 43.50 | 86.50 | 0.23 | 1.87 | −0.45 |
| ILL 7808 | 52.50 | 84.00 | 0.27 | 2.30 | −0.29 | ILL 6089 | 45.00 | 88.50 | 0.27 | 2.40 | −0.50 |
| ILL 7832 | 51.00 | 84.00 | 0.38 | 3.05 | −0.29 | ILL 6091 | 45.50 | 85.50 | 0.55 | 4.09 | −0.53 |
| ILL 6105 | 57.00 | 89.00 | 0.26 | 2.50 | −0.30 | ILL 6100 | 44.50 | 96.50 | 0.28 | 3.20 | −0.62 |
| ILL 7314 | 54.00 | 86.00 | 0.29 | 2.62 | −0.30 | ILL 6319 | 45.00 | 90.50 | 0.32 | 2.56 | −0.48 |
| ILL 6101 | 55.00 | 84.00 | 0.21 | 2.25 | −0.31 | ILL 6322 | 44.00 | 91.00 | 0.26 | 2.41 | −0.51 |
| ILL 7326 | 54.00 | 86.00 | 0.46 | 3.95 | −0.32 | ILL 6362 | 43.50 | 90.50 | 0.55 | 3.85 | −0.50 |
| ILL 7813 | 50.00 | 84.00 | 0.55 | 4.45 | −0.32 | ILL 7232 | 43.50 | 89.50 | 0.53 | 3.43 | −0.44 |
| ILL 7800 | 52.50 | 87.00 | 0.41 | 3.75 | −0.34 | ILL 7266 | 44.50 | 87.50 | 0.36 | 3.41 | −0.59 |
| ILL 7232 | 57.00 | 88.00 | 0.25 | 2.85 | −0.34 | ILL 7289 | 44.50 | 83.00 | 0.20 | 3.48 | −0.75 |
| ILL 7797 | 55.00 | 87.00 | 0.31 | 3.20 | −0.34 | ILL 7300 | 43.50 | 87.00 | 0.49 | 3.61 | −0.51 |
| ILL 7815 | 51.00 | 84.00 | 0.47 | 4.16 | −0.35 | ILL 7305 | 44.50 | 85.00 | 0.13 | 2.10 | −0.57 |
| ILL 7801 | 55.00 | 83.00 | 0.21 | 2.60 | −0.35 | ILL 7307 | 45.00 | 87.00 | 0.32 | 2.51 | −0.47 |
| ILL 7820 | 54.50 | 84.00 | 0.29 | 3.16 | −0.35 | ILL 7339 | 44.00 | 86.50 | 0.35 | 4.07 | −0.71 |
| ILL 6346 | 54.00 | 86.00 | 0.37 | 3.65 | −0.35 | ILL 7797 | 47.50 | 92.00 | 0.24 | 3.52 | −0.71 |
| ILL 7383 | 55.00 | 83.50 | 0.46 | 4.30 | −0.35 | ILL 7800 | 44.50 | 84.50 | 0.26 | 2.41 | −0.51 |
| ILL 4910 | 55.00 | 86.00 | 0.21 | 2.70 | −0.36 | ILL 7804 | 43.00 | 83.50 | 0.53 | 3.37 | −0.44 |
| ILL 6089 | 55.50 | 86.00 | 0.22 | 2.80 | −0.36 | ILL 7812 | 43.50 | 85.00 | 0.53 | 3.54 | −0.46 |
| ILL 8024 | 53.00 | 88.00 | 0.23 | 2.83 | −0.37 | ILL 7824 | 45.50 | 85.00 | 0.74 | 4.74 | −0.47 |
| ILL 6060 | 53.00 | 88.00 | 0.20 | 2.69 | −0.37 | ILL 7829 | 44.00 | 84.00 | 0.43 | 3.61 | −0.57 |
| ILL 6072 | 54.00 | 88.00 | 0.48 | 4.67 | −0.38 | ILL 7833 | 45.00 | 85.00 | 0.23 | 3.28 | −0.68 |
| ILL 7814 | 54.00 | 85.00 | 0.20 | 2.78 | −0.38 | ILL 7838 | 44.00 | 83.00 | 0.43 | 2.95 | −0.45 |
| ILL 6107 | 53.00 | 87.00 | 0.50 | 4.80 | −0.38 | ILL 8012 | 43.00 | 84.50 | 0.41 | 2.84 | −0.45 |
| ILL 8280 | 55.00 | 85.00 | 0.20 | 2.85 | −0.38 | ILL 8022 | 47.50 | 93.00 | 0.46 | 4.21 | −0.62 |
| ILL 2524 | 54.00 | 83.00 | 0.21 | 2.97 | −0.39 | ||||||
| ILL 5943 | 53.00 | 85.00 | 0.19 | 3.09 | −0.42 | ||||||
| ILL 7804 | 58.00 | 88.00 | 0.20 | 3.35 | −0.42 | ||||||
| ILL 7829 | 55.00 | 84.00 | 0.28 | 3.80 | −0.42 | ||||||
| ILL 4772 | 53.00 | 84.00 | 0.24 | 3.54 | −0.43 | ||||||
| ILL 6081 | 54.00 | 86.00 | 0.30 | 4.17 | −0.45 | ||||||
| ILL 7799 | 55.00 | 82.00 | 0.22 | 3.80 | −0.46 | ||||||
| ILL 7816 | 61.50 | 89.50 | 0.21 | 4.95 | −0.56 | ||||||
| ILL 6054 | 52.00 | 86.00 | 0.25 | 4.85 | −0.56 | ||||||
| ILL 5505 | 58.50 | 109.00 | 0.17 | 5.08 | −0.61 | ||||||
| ILL 6320 | 54.00 | 88.50 | 0.17 | 7.20 | −0.83 | ||||||
References
- Kumar, S.; Rajendran, K.; Kumar, J.; Hamwieh, A.; Baum, M. Current knowledge in lentil genomics and its application for crop improvement. Front. Plant Sci. 2015, 6, 78. [Google Scholar] [CrossRef] [PubMed]
- Migliozzi, M.; Thavarajah, D.; Thavarajah, P.; Smith, P. Lentil and kale: Complementary nutrient-rich whole food sources to combat micronutrient and calorie malnutrition. Nutrients 2015, 7, 9285–9298. [Google Scholar] [CrossRef] [PubMed]
- Hamwieh, A.; Udupa, S.; Choumane, W.; Sarker, A.; Dreyer, F.; Jung, C.; Baum, M. A genetic linkage map of Lens sp. based on microsatellite and AFLP markers and the localization of fusarium vascular wilt resistance. Theor. Appl. Genet. 2005, 110, 669–677. [Google Scholar] [CrossRef] [PubMed]
- FAO. Statistical Database. Food and Agriculture Organization of the United Nations, Rome, Italy. 2019. Available online: http://www.fao.org/faostat.org (accessed on 20 April 2020).
- Kumar, S.; Barpete, S.; Kumar, J.; Gupta, P.; Sarker, A. Global lentil production: Constraints and strategies. SATSA Mukhapatra Annu. Tech. 2013, 17, 1–13. [Google Scholar]
- Giss/NASA. Robert B Schmunk. National Aeronautics and Space Administration Goddard Institute for Space Studies. 2019. Available online: https://data.giss.nasa.gov/gistemp/ (accessed on 21 November 2019).
- Giorgi, F. Climate change hot-spots. Geophys. Res. Lett. 2006, 33, 1–4. [Google Scholar] [CrossRef]
- Öktem, H.; Eyidoðan, F.; Demirba, D.; Bayraç, A.; Öz, M.; Özgür, E.; Selçuk, F.; Yücel, M. Antioxidant Responses of Lentil to Cold and Drought Stress. J. Plant Biochem. Biotechnol. 2008, 17, 15–21. [Google Scholar] [CrossRef]
- Ibrahim, H.M. Heat stress in food legumes: Evaluation of membrane thermostability methodology and use of infra-red thermometry. Euphytica 2011, 180, 99–105. [Google Scholar] [CrossRef]
- Bhandari, K.; Siddique, K.H.; Turner, N.C.; Kaur, J.; Singh, S.; Agrawal, S.K.; Nayyar, H. Heat Stress at Reproductive Stage Disrupts Leaf Carbohydrate Metabolism, Impairs Reproductive Function, and Severely Reduces Seed Yield in Lentil. J. Crop Improv. 2016, 30, 118–151. [Google Scholar] [CrossRef]
- Delahunty, A.; Nuttall, J.; Nicolas, M.; Brand, J. Genotypic heat tolerance in lentil. In Proceedings of the 17th ASA Conference, Hobart, Australia, 20–24 September 2015; pp. 20–24. [Google Scholar]
- Kumar, J.; Kant, R.; Kumar, S.; Basu, P.S.; Sarker, A.; Singh, N.P. Heat tolerance in lentil under field conditions. Legume Genom. Genet. 2016, 7, 1–11. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; HanumanthaRao, B.; Nair, R.M.; Vara Prasad, P.V.; Kumar, S.; Gaur, P.M.; Farooq, M.; Siddique, K.H.; Varshney, R.K.; et al. Food legumes and rising temperatures: Effects, adaptive functional mechanisms specific to reproductive growth stage and strategies to improve heat tolerance. Front. Plant Sci. 2017, 8, 1658. [Google Scholar] [CrossRef] [PubMed]
- Idrissi, O.; Chafika, H.; Nsarellah, N. Comparaison de lignées avancées de lentille sous stress hydrique durant la phase de floraison et formation des gousses. Nat. Technol. 2013, 8, 53A. [Google Scholar]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Bhandari, K.; Kumar, S.; Kumar, J.; Vara Prasad, P.V.; Siddique, K.H.; Nayyar, H. Influence of drought and heat stress, applied independently or in combination during seed development, on qualitative and quantitative aspects of seeds of lentil (Lens culinaris Medikus) genotypes, differing in drought sensitivity. Plant Cell Environ. 2019, 42, 198–211. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Effects of drought, heat and their interaction on the growth, yield and photosynthetic function of lentil (Lens culinaris Medikus) genotypes varying in heat and drought sensitivity. Front. Plant Sci. 2017, 8, 1776. [Google Scholar] [CrossRef]
- Hamidou, F.; Halilou, O.; Vadez, V. Assessment of groundnut under combined heat and drought stress. J. Agron. Crop Sci. 2013, 199, 1–11. [Google Scholar] [CrossRef]
- Awasthi, R.; Gaur, P.; Turner, N.C.; Vadez, V.; Siddique, K.H.; Nayyar, H. Effects of individual and combined heat and drought stress during seed filling on the oxidative metabolism and yield of chickpea (Cicer arietinum) genotypes differing in heat and drought tolerance. Crop Pasture Sci. 2017, 68, 823–841. [Google Scholar] [CrossRef]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Siddique, K.H.; Nayyar, H. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. 2014, 41, 1148–1167. [Google Scholar] [CrossRef]
- Canci, H.; Toker, C. Evaluation of yield criteria for drought and heat resistance in chickpea (Cicer arietinum L.). J. Agron. Crop Sci. 2009, 195, 47–54. [Google Scholar] [CrossRef]
- Savin, R.; Nicolas, M.E. Effects of short periods of drought and high temperature on grain growth and starch accumulation of two malting barley cultivars. Funct. Plant Biol. 1996, 23, 201–210. [Google Scholar] [CrossRef]
- Hossain, A.; da Silva, J.A.; Lozovskaya, M.V.; Zvolinsky, V.P. High temperature combined with drought affect rainfed spring wheat and barley in South-Eastern Russia: I. Phenology and growth. Saudi J. Biol. Sci. 2012, 19, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.V.; Pisipati, S.R.; Momčilović, I.; Ristic, Z. Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat. J. Agron. Crop Sci. 2011, 197, 430–441. [Google Scholar] [CrossRef]
- Zhang, X.; Cai, J.; Wollenweber, B.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Multiple heat and drought events affect grain yield and accumulations of high molecular weight glutenin subunits and glutenin macropolymers in wheat. J. Cereal Sci. 2013, 57, 134–140. [Google Scholar] [CrossRef]
- Kumar, J.; Basu, P.S.; Srivastava, E.; Chaturvedi, S.K.; Nadarajan, N.; Kumar, S. Phenotyping of traits imparting drought tolerance in lentil. Crop Pasture Sci. 2012, 63, 547–554. [Google Scholar] [CrossRef]
- Krishnamurthy, L.; Kashiwagi, J.; Gaur, P.M.; Upadhyaya, H.D.; Vadez, V. Sources of tolerance to terminal drought in the chickpea (Cicer arietinum L.) minicore germplasm. Field Crops Res. 2010, 119, 322–330. [Google Scholar] [CrossRef]
- Krishnamurthy, L.; Gaur, P.M.; Basu, P.S.; Chaturvedi, S.K.; Tripathi, S.; Vadez, V.; Rathore, A.; Varshney, R.K.; Gowda, C.L. Large genetic variation for heat tolerance in the reference collection of chickpea (Cicer arietinum L.) germplasm. Plant Genet. Resour. 2011, 9, 59–69. [Google Scholar] [CrossRef]
- Devasirvatham, V.; Gaur, P.M.; Raju, T.N.; Trethowan, R.M.; Tan, D.K. Field response of chickpea (Cicer arietinum L.) to high temperature. Field Crops Res. 2015, 172, 59–71. [Google Scholar] [CrossRef]
- Ali, M.B.; El-Sadek, A.N. Evaluation of drought tolerance indices for wheat (Triticum aestivum L.) under irrigated and rainfed conditions. Commun. Biometry Crop Sci. 2016, 11, 77–89. [Google Scholar]
- Naveed, M.; Nadeem, M.; Shafiq, M.; Rafiq, C.M.; Zahid, M.A. Selection of promising chickpea (cicer arietinum l.) genotypes using drought tolerance indices. JAPS J. Anim. Plant Sci. 2019, 29, 278–290. [Google Scholar]
- Kakaei, M. Evaluation of Terminal Drought Stress Tolerance in Lentil (Lens Culinaris). Isfahan Univ. Technol. J. Crop Prod. Process. 2019, 8, 59–71. [Google Scholar] [CrossRef][Green Version]
- Siahsar, B.A.; Ganjali, S.; Allahdoo, M. Evaluation of drought tolerance indices and their relationship with grain yield of lentil lines in drought-stressed and irrigated environments. Aust. J. Basic Appl. Sci. 2010, 4, 4336–4346. [Google Scholar]
- Khan, A.A.; Kabir, M.R. Evaluation of spring wheat genotypes (Triticum aestivum L.) for heat stress tolerance using different stress tolerance indices. Cercet. Agron. Mold. 2015, 47, 49–63. [Google Scholar] [CrossRef]
- Jha, U.C.; Jha, R.; Singh, N.P.; Shil, S.; Kole, P.C. Heat Tolerance Indices and Their Role in Selection of Heat Stress Tolerant Chickpea (Cicer arietinum) Genotypes. Indian J. Agric. Sci. 2018, 88, 260–270. [Google Scholar]
- Benhin, J.K. Climate Change and South African Agriculture: Impacts and Adaptation Options; CEEPA Discussion Paper; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Dube, T.; Moyo, P.; Ncube, M.; Nyathi, D. The impact of climate change on agro-ecological based livelihoods in Africa: A review. J. Sustain. Dev. 2016, 9, 256–267. [Google Scholar] [CrossRef]
- Nhemachena, C.; Mano, R. Assessment of the Economic Impacts of Climate Change on Agriculture in Zimbabwe: A Ricardian Approach; The World Bank: Washington, DC, USA, 2007. [Google Scholar]
- Gaur, P.M.; Jukanti, A.K.; Samineni, S.; Chaturvedi, S.K.; Basu, P.S.; Babbar, A.; Jayalakshmi, V.; Nayyar, H.; Devasirvatham, V.; Mallikarjuna, N.; et al. Climate Change and Heat Stress Tolerance in Chickpea; Wiley Online Library: Hoboken, NJ, USA, 2013; pp. 839–855. [Google Scholar]
- Khazaei, H.; Street, K.; Bari, A.; Mackay, M.; Stoddard, F.L. The FIGS (Focused Identification of Germplasm Strategy) approach identifies traits related to drought adaptation in Vicia faba genetic resources. PLoS ONE 2013, 8, e63107. [Google Scholar] [CrossRef] [PubMed]
- Mackay, M.; Von Bothmer, R.; Skovmand, B. Conservation and utilization of plant genetic resources–future directions. Czech J. Genet. Plant Breed. 2005, 41. [Google Scholar] [CrossRef]
- El Bouhssini, M.; Street, K.; Amri, A.; Mackay, M.; Ogbonnaya, F.C.; Omran, A.; Abdalla, O.; Baum, M.; Dabbous, A.; Rihawi, F. Sources of resistance in bread wheat to Russian wheat aphid (Diuraphis noxia) in Syria identified using the Focused Identification of Germplasm Strategy (FIGS). Plant Breed. 2011, 130, 96–97. [Google Scholar] [CrossRef]
- Bari, A.; Street, K.; Mackay, M.; Endresen, D.T.; De Pauw, E.; Amri, A. Focused identification of germplasm strategy (FIGS) detects wheat stem rust resistance linked to environmental variables. Genet. Resour. Crop Evol. 2012, 59, 1465–1481. [Google Scholar] [CrossRef]
- Endresen, D.T.; Street, K.; Mackay, M.; Bari, A.; Amri, A.; De Pauw, E.; Nazari, K.; Yahyaoui, A. Sources of resistance to stem rust (Ug99) in bread wheat and durum wheat identified using focused identification of germplasm strategy. Crop Sci. 2012, 52, 764–773. [Google Scholar] [CrossRef]
- El Bouhssini, M.; Street, K.; Joubi, A.; Ibrahim, Z.; Rihawi, F. Sources of wheat resistance to Sunn pest, Eurygaster integriceps Puton, in Syria. Genet. Resour. Crop Evol. 2009, 56, 1065. [Google Scholar] [CrossRef]
- Elhaddoury, J.; Lhaloui, S.; Udupa, S.M.; Moatassim, B.; Taiq, R.; Rabeh, M.; Kamlaoui, M.; Hammadi, M. Registration of ‘Kharoba’: A bread wheat cultivar developed through doubled haploid breeding. J. Plant Regist. 2012, 6, 169–173. [Google Scholar] [CrossRef][Green Version]
- Moussadek, R.; Iaaich, H.; Elouadi, M. “Soil Survey Report of ICARDA farm- Marchouch.” Icarda-Inra; ICARDA: Rabat, Morocco, 2014. [Google Scholar]
- Agrawal, S.K. Lentil Ontology-Crop Ontology Curation Tool. Crop Ontology for Agricultural Data. CGIAR Centers. Available online: http://agroportal.lirmm.fr/ontologies/CO_339 (accessed on 20 January 2020).
- Bidinger, F.R.; Mahalakshmi, V.; Rao, G.D. Assessment of drought resistance in pearl millet (Pennisetum americanum (L.) Leeke). II. Estimation of genotype response to stress. Aust. J. Agric. Res. 1987, 38, 49–59. [Google Scholar] [CrossRef]
- Devasirvatham, V.; Tan, D.K. Impact of high temperature and drought stresses on chickpea production. Agronomy 2018, 8, 145. [Google Scholar] [CrossRef]
- Fernandez, G.C. Effective selection criteria for assessing plant stress tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and other Food Crops in Temperature and Water Stress, Shanhua, Taiwan, 13–16 August 1992; pp. 257–270. [Google Scholar]
- Rosielle, A.A.; Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environment 1. Crop Sci. 1981, 21, 943–946. [Google Scholar] [CrossRef]
- Schneider, K.A.; Rosales-Serna, R.; Ibarra-Perez, F.; Cazares-Enriquez, B.; Acosta-Gallegos, J.A.; Ramirez-Vallejo, P.; Wassimi, N.; Kelly, J.D. Improving common bean performance under drought stress. Crop Sci. 1997, 37, 43–50. [Google Scholar] [CrossRef]
- Choudhury, D.R.; Tarafdar, S.; Das, M.; Kundagrami, S. Screening lentil (Lens culinaris Medik.) germplasms for heat tolerance. Trends Biosci. 2012, 5, 143–146. [Google Scholar]
- Kaur, R.; Bains, T.S.; Bindumadhava, H.; Nayyar, H. Responses of mungbean (Vigna radiata L.) genotypes to heat stress: Effects on reproductive biology, leaf function and yield traits. Sci. Hortic. 2015, 197, 527–541. [Google Scholar] [CrossRef]
- Sharma, L.; Priya, M.; Bindumadhava, H.; Nair, R.M.; Nayyar, H. Influence of high temperature stress on growth, phenology and yield performance of mungbean [Vigna radiata (L.) Wilczek] under managed growth conditions. Sci. Hortic. 2016, 213, 379–391. [Google Scholar] [CrossRef]
- Sita, K.; Sehgal, A.; Kumar, J.; Kumar, S.; Singh, S.; Siddique, K.H.; Nayyar, H. Identification of high-temperature tolerant lentil (Lens culinaris Medik.) genotypes through leaf and pollen traits. Front. Plant Sci. 2017, 8, 744. [Google Scholar] [CrossRef]
- Summerfield, R.J.; Roberts, E.H.; Erskine, W.; Ellis, R.H. Effects of temperature and photoperiod on flowering in lentils (Lens culinaris Medic.). Ann. Bot. 1985, 56, 659–671. [Google Scholar] [CrossRef]
- Erskine, W.; Ellis, R.H.; Summerfield, R.J.; Roberts, E.H.; Hussain, A. Characterization of responses to temperature and photoperiod for time to flowering in a world lentil collection. Theor. Appl. Genet. 1990, 80, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Upadhyaya, H.D.; Dronavalli, N.; Gowda, C.L.; Singh, S. Identification and evaluation of chickpea germplasm for tolerance to heat stress. Crop Sci. 2011, 51, 2079–2094. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S. Impact of elevated temperatures on growth and yield of chickpea (Cicer arietinum L.). Field Crops Res. 2014, 164, 90–97. [Google Scholar] [CrossRef]
- Abdelmula, A.A.; Abuanja, I.K. Genotypic responses, yield stability, and association between characters among some of Sudanese Faba bean (Vicia faba L.) genotypes under heat stress. In Proceedings of the Conference on International Agricultural Research for Development, Rome, Italy, 31 October 2007; pp. 9–11. [Google Scholar]
- Gan, Y.; Angadi, S.V.; Cutforth, H.; Potts, D.; Angadi, V.V.; McDonald, C.L. Canola and mustard response to short periods of temperature and water stress at different developmental stages. Can. J. Plant Sci. 2004, 84, 697–704. [Google Scholar] [CrossRef]
- Gaur, P.M.; Samineni, S.; Krishnamurthy, L.; Varshney, R.K.; Kumar, S.; Ghanem, M.E.; Beebe, S.E.; Rao, I.M.; Chaturvedi, S.K.; Basu, P.S.; et al. High Temperature Tolerance in Grain Legumes; Legume Perspectives. In Proceedings of the 6th International Food Legumes Research Conference (IFLRC VI) and 7th International Conference on Legume Genetics and Genomics (ICLGG VII), Saskatoon, SK, Canada, 7–11 Juiy 2014; pp. 23–24. [Google Scholar]
- Fang, X.; Turner, N.C.; Yan, G.; Li, F.; Siddique, K.H. Flower numbers, pod production, pollen viability, and pistil function are reduced and flower and pod abortion increased in chickpea (Cicer arietinum L.) under terminal drought. J. Exp. Bot. 2010, 61, 335–345. [Google Scholar] [CrossRef]
- Ohnishi, S.; Miyoshi, T.; Shirai, S. Low temperature stress at different flower developmental stages affects pollen development, pollination, and pod set in soybean. Environ. Exp. Bot. 2010, 69, 56–62. [Google Scholar] [CrossRef]
- Hendrix, J.E. Production-related assimilate transport and partitioning. Handb. Plant Crop Physiol. 2001, 18, 421. [Google Scholar]
- Younis, N.; Hanif, M.; Sadiq, S.; Abbas, G.; Asghar, M.J.; Haq, M.A. Estimates of genetic parameters and path analysis in lentil (Lens culinaris Medik). Pak. J. Agri. Sci. 2008, 45, 44–48. [Google Scholar]
- Ul Hussan, S.; Khuroo, N.S.; Lone, A.A.; Dar, Z.A.; Dar, S.A.; Dar, M.S. Study of variability and association analysis for various agromorphological traits in lentil (Lens culinaris M.). J. Pharmacogn. Phytochem. 2018, 7, 2172–2175. [Google Scholar]
- Ahmadi, A.; Dehaghi, M.A.; Fotokian, M.H.; Sedghi, M.; Far, C.M. Evaluation of Stress Tolerance Indices in a Number of Advanced Genotypes of Lentil (Lens Culinaris Medik) Under Rainfed and Low Irrigation Conditions. Plant Arch. 2019, 19, 490–499. [Google Scholar]
- Prasad, P.V.; Staggenborg, S.A.; Ristic, Z. Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants. In Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes; ASA, CSSA, SSSA: Phoenix, AZ, USA, 2008; pp. 301–355. [Google Scholar] [CrossRef]
- Tullu, A.; Kusmenoglu, I.; McPhee, K.E.; Muehlbauer, F.J. Characterization of core collection of lentil germplasm for phenology, morphology, seed and straw yields. Genet. Resour. Crop Evol. 2001, 48, 143–152. [Google Scholar] [CrossRef]
- Chakherchaman, S.A.; Mostafaei, H.; Imanparast, L.; Eivazian, M.R. Evaluation of drought tolerance in lentil advanced genotypes in Ardabil region, Iran. J. Food Agric. Environ. 2009, 7, 283–288. [Google Scholar]
- Kumar, J.; Gupta, S.; Gupta, P.; Dubey, S.; Tomar, R.S.; Kumar, S. Breeding strategies to improve lentil for diverse agro-ecological environments. Indian J. Genet. Plant Breed 2016, 76, 530–549. [Google Scholar] [CrossRef]
- Saxena, N.P. Screening for adaptation to drought: Case studies with chickpea and pigeonpea [a review]. In Proceedings of the Consultants’ Workshop: Adaptation of Chickpea and Pigeonpea to Abiotic Stresses, Patancheru, India, 19–21 December 1984. [Google Scholar]
- Berger, J.D.; Milroy, S.P.; Turner, N.C.; Siddique, K.H.; Imtiaz, M.; Malhotra, R. Chickpea evolution has selected for contrasting phenological mechanisms among different habitats. Euphytica 2011, 180, 1–5. [Google Scholar] [CrossRef]
- Erdemci, I. Evaluation of Drought Tolerance Selection Indices Using Grain Yield in Chickpea (Cicer arietinum L.). Not. Sci. Biol. 2018, 10, 439–446. [Google Scholar] [CrossRef]
- Shabani, A.; Zebarjadi, A.; Mostafaei, A.; Saeidi, M.; Poordad, S.S. Evaluation of drought stress tolerance in promising lines of chickpea (Cicer arietinum L.) using drought resistance indices. Environ. Stresses Crop Sci. 2018, 11, 289–299. [Google Scholar]
- Rad, M.R.; Ghasemi, A.; Arjmandinejad, A. Study of limit irrigation on yield of lentil (Lens culinaris) genotypes of national plant gene bank of Iran by drought resistance indices. Am. Eur. J. Agric. Environ. Sci. 2009, 6, 352–355. [Google Scholar]
- Mishra, B.K.; Srivastava, J.P.; Lal, J.P.; Sheshshayee, M.S. Physiological and biochemical adaptations in lentil genotypes under drought stress. Russ. J. Plant Physiol. 2016, 63, 695–708. [Google Scholar] [CrossRef]
- Grzesiak, S.; Hordyńska, N.; Szczyrek, P.; Grzesiak, M.T.; Noga, A.; Szechyńska-Hebda, M. Variation among wheat (Triticum easativum L.) genotypes in response to the drought stress: I–selection approaches. J. Plant Interact. 2019, 14, 30–44. [Google Scholar] [CrossRef]
- Ajalli, J.; Salehi, M. Evaluation of drought stress indices in barley (Hordeum vulgare L.). Ann. Biol. Res. 2012, 3, 5515–5520. [Google Scholar]
- Zare, M. Evaluation of drought tolerance indices for the selection of Iranian barley (Hordeum vulgare) cultivars. Afr. J. Biotechnol. 2012, 11, 15975–15981. [Google Scholar]
- Ahari, D.S.; Kashi, A.K.; Hassandokht, M.R.; Amri, A.; Alizadeh, K. Assessment of drought tolerance in Iranian fenugreek landraces. J. Food Agri. Environ. 2009, 7, 414–419. [Google Scholar]
- Akcura, M.; Ceri, S. Evaluation of drought tolerance indices for selection of Turkish oat (Avena sativa L.) landraces under various environmental conditions. Zemdirb. Agric. 2011, 98, 157–166. [Google Scholar]
- Papathanasiou, F.; Dordas, C.; Gekas, F.; Pankou, C.; Ninou, E.; Mylonas, I.; Tsantarmas, K.; Sistanis, I.; Sinapidou, E.; Lithourgidis, A.; et al. The use of stress tolerance indices for the selection of tolerant inbred lines and their correspondent hybrids under normal and water-stress conditions. Procedia Environ. Sci. 2015, 29, 274–275. [Google Scholar] [CrossRef]
- El Sabagh, A.; Barutcular, C.; Hossain, A.; Islam, M.S. Response of maize hybrids to drought tolerance in relation to grain weight. Fresenius Environ. Bull. 2018, 27, 2476–2482. [Google Scholar]
- Farshadfar, E.; Elyasi, P. Screening quantitative indicators of drought tolerance in bread wheat (Triticum aestivum L.) landraces. Eur. J. Exp. Biol. 2012, 2, 577–584. [Google Scholar]
- Ganjali, A.; Bagheri, A.; Porsa, H. Evaluation of Chickpea (Cicer Arietinum L.) Germplasm for Drought Resistance. J. Agron. Res. Iran 2009, 7, 183–194. [Google Scholar]
- Majidi, M.M.; Tavakoli, V.; Mirlohi, A.; Sabzalian, M.R. Wild safflower species (‘Carthamus oxyacanthus’ Bieb.): A possible source of drought tolerance for arid environments. Aust. J. Crop Sci. 2011, 5, 1055. [Google Scholar]
- El-Mohsen, A.A.; El-Shafi, M.A.; Gheith, E.M.; Suleiman, H.S. Using different statistical procedures for evaluating drought tolerance indices of bread wheat genotypes. Adv. Agric. Biol. 2015, 4, 19–30. [Google Scholar]
- Farshadfar, E.; Poursiahbidi, M.M.; Safavi, S.M. Assessment of drought tolerance in land races of bread wheat based on resistance/tolerance indices. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 143–158. [Google Scholar]
- Talebi, R.; Baghebani, N.; Karami, E.; Ensafi, M.H. Defining selection indices for drought tolerance in chickpea under terminal drought stresses. J. Appl. Biol. Sci. 2011, 5, 33–38. [Google Scholar]

| Source | df | PH | DF | DM | PBPP | SBPP | TBPP | NTPP | NUPP | NFPP | BPP | GYP | HSW |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession (A) | 161 | 3179.93 ** | 9020.56 ** | 7926.79 ** | 85.73 * | 2809.87 ** | 3116.53 ** | 134,681.09 ** | 17,979.63 ** | 97,275.47 ** | 6602.61 ** | 240.04 ** | 128.57 ** |
| Treatment (T) | 2 | 8801.50 ** | 81,401.01 ** | 194,468.02 ** | 87.07 ** | 4447.67 ** | 7474.59 ** | 221,759.77 ** | 6956.78 ** | 169,051.73 ** | 17,534.56 ** | 847.36 ** | 153.11 ** |
| A × T | 322 | 1373.34 ns | 3564.90 ** | 9245.98 ** | 109.86 ns | 2796.58 * | 2834.41 ns | 158,553.22 * | 30,640.38 ** | 101,053.29 * | 9055.03 ns | 235.05 ** | 84.96 ** |
| Error | 486 | 2734.02 | 1675.88 | 4595.96 | 181.53 | 3506.01 | 3745.33 | 180,293.52 | 31,008.66 | 125,776.68 | 11,795.35 | 258.51 | 53.9 |
| R2 | 0.83 ** | 0.98 ** | 0.98 ** | 0.61 ** | 0.74 ** | 0.78 ** | 0.74 ** | 0.66 ** | 0.75 ** | 0.74 ** | 0.84 ** | 0.87 ** |
| Source | df | PH | DF | DM | PBPP | SBPP | TBPP | NTPP | NUPP | NFPP | BPP | GYP | HSW |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (A) | 161 | 5998.87 ** | 12,577.23 ** | 8763.72 ** | 179.10 ** | 4794.79 ** | 2163.08 ** | 217,688.81 ** | 16,449.68 ** | 164,517.17 ** | 4042.86 ** | 297.13 ** | 168.53 ** |
| (T) | 2 | 23,422.11 ** | 235,449.92 ** | 223,732.48 ** | 103.03 ** | 46,726.49 ** | 33,818.93 ** | 638,843.72 ** | 45,673.66 ** | 344,672.12 ** | 18,250.69 ** | 976.43 ** | 67.07 ** |
| A × T | 322 | 6986.42 ** | 6242.97 ** | 5907.75 ** | 222.12 ** | 7827.42 ** | 4388.39 ** | 324,520.74 ** | 35,015.62 ** | 238,902.17 ** | 6559.96 ** | 382.04 ** | 86.57 ** |
| Error | 486 | 4714.99 | 503.09 | 952.59 | 122.45 | 4694.98 | 2275.94 | 160,769.5 | 14,911.52 | 121,599.93 | 2839.61 | 217.94 | 74.55 |
| R2 | 0.88 ** | 0.99 ** | 0.99 ** | 0.8 ** | 0.93 ** | 0.95 ** | 0.88 ** | 0.87 ** | 0.86 ** | 0.91 ** | 0.88 ** | 0.81 ** |
| Marchouch | Tessaout | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Location | Accession | DF | DM | GYP | HTI | DF | DM | GYP | HTI |
| Marchouch | ILL 7833 | 53 | 85.00 | 4.33 | 1.97 | 45 | 80.00 | 0.23 | −0.68 |
| ILL 6338 | 54 | 86.00 | 3.16 | 1.52 | 47 | 83.00 | 0.59 | −0.12 | |
| ILL 7835 | 54 | 86.00 | 3.02 | 1.39 | 43.5 | 81.00 | 3.52 | 1.95 | |
| ILL 6104 | 53 | 85.00 | 2.52 | 1.11 | 44 | 82.20 | 0.43 | −0.27 | |
| Mean | 53.5 | 85.5 | 3.26 | 1.49 | 44.87 | 81.55 | 1.19 | 0.22 | |
| SD | 0.5 | 0.5 | 0.66 | 0.31 | 1.34 | 1.14 | 1.34 | 1.02 | |
| Tessaout | ILL 7835 | 54 | 86.00 | 3.02 | 1.39 | 43.5 | 81.00 | 3.52 | 1.95 |
| ILL 7814 | 54 | 85.00 | 0.2 | −0.38 | 43.5 | 82.50 | 3.26 | 1.83 | |
| ILL 8029 | 55 | 85.00 | 1.43 | 0.44 | 47.5 | 83.50 | 2.65 | 1.36 | |
| Mean | 54.33 | 85.33 | 1.55 | 0.48 | 44.83 | 82.33 | 3.14 | 1.71 | |
| SD | 0.47 | 0.47 | 1.15 | 0.72 | 1.88 | 1.02 | 0.36 | 0.25 | |
| Location | Accession | DF | DM | Ys | Yp | STI | GMP | MP | TOL | HARM |
|---|---|---|---|---|---|---|---|---|---|---|
| Marchouch | ILL 7835 | 59.50 | 89.50 | 1.88 | 3.98 | 1.12 | 2.73 | 2.93 | 2.10 | 2.55 |
| ILL 6075 | 58.00 | 87.00 | 1.99 | 2.15 | 0.64 | 2.07 | 2.07 | 0.16 | 2.07 | |
| ILL 6362 | 59.00 | 87.50 | 0.77 | 5.30 | 0.61 | 2.02 | 3.04 | 4.53 | 1.34 | |
| ILL 7819 | 55.00 | 87.50 | 1.54 | 2.43 | 0.56 | 1.93 | 1.98 | 0.89 | 1.88 | |
| ILL 7266 | 56.00 | 88.00 | 1.20 | 2.20 | 0.40 | 1.62 | 1.70 | 1.00 | 1.55 | |
| ILL 6361 | 59.00 | 89.00 | 0.60 | 4.21 | 0.38 | 1.59 | 2.40 | 3.61 | 1.05 | |
| ILL 880 | 64.00 | 98.00 | 0.67 | 3.66 | 0.37 | 1.56 | 2.16 | 2.99 | 1.13 | |
| ILL 4605 | 49.00 | 83.30 | 0.59 | 4.07 | 0.36 | 1.55 | 2.33 | 3.48 | 1.03 | |
| ILL 6088 | 57.50 | 87.50 | 0.90 | 2.45 | 0.33 | 1.48 | 1.67 | 1.55 | 1.31 | |
| ILL 7815 | 58.50 | 85.50 | 0.48 | 4.16 | 0.30 | 1.41 | 2.32 | 3.69 | 0.85 | |
| Mean | 57.55 | 88.28 | 1.062 | 3.461 | 0.507 | 1.796 | 2.26 | 2.4 | 1.476 | |
| SD | 3.64 | 3.64 | 0.53 | 1.02 | 0.23 | 0.38 | 0.43 | 1.38 | 0.51 | |
| Tessaout | ILL 7814 | 42.50 | 83.00 | 2.71 | 4.63 | 1.68 | 3.54 | 3.67 | 1.92 | 3.42 |
| ILL 7835 | 42.50 | 81.50 | 2.28 | 5.30 | 1.61 | 3.47 | 3.79 | 3.02 | 3.19 | |
| ILL 7804 | 43.00 | 82.50 | 2.53 | 3.37 | 1.14 | 2.91 | 2.95 | 0.84 | 2.89 | |
| ILL 6101 | 45.00 | 86.00 | 2.20 | 3.11 | 0.91 | 2.62 | 2.66 | 0.91 | 2.58 | |
| ILL 6100 | 45.00 | 83.50 | 2.22 | 3.20 | 0.95 | 2.67 | 2.71 | 0.98 | 2.62 | |
| ILL 7807 | 45.50 | 83.50 | 1.40 | 4.86 | 0.91 | 2.61 | 3.13 | 3.46 | 2.18 | |
| ILL 6091 | 47.00 | 82.50 | 1.23 | 4.09 | 0.67 | 2.24 | 2.66 | 2.86 | 1.88 | |
| ILL 8029 | 46.50 | 83.50 | 1.11 | 4.25 | 0.63 | 2.17 | 2.68 | 3.14 | 1.76 | |
| ILL 4605 | 51.58 | 81.07 | 1.15 | 3.81 | 0.59 | 2.09 | 2.48 | 2.66 | 1.77 | |
| ILL 8061 | 66.50 | 97.00 | 1.16 | 3.76 | 0.58 | 2.08 | 2.46 | 2.60 | 1.77 | |
| Mean | 47.51 | 84.41 | 1.79 | 4.04 | 0.97 | 2.64 | 2.92 | 2.24 | 2.41 | |
| SD | 6.82 | 4.39 | 0.61 | 0.69 | 0.38 | 0.51 | 0.45 | 0.95 | 0.59 |
| Location | Stress Parameter | Ys | Yp | STI | GMP | MP | TOL | HARM |
|---|---|---|---|---|---|---|---|---|
| Tessaout | Yp | 0.192 * | 1 | |||||
| STI | 0.862 ** | 0.613 ** | 1 | |||||
| GMP | 0.862 ** | 0.613 ** | 1.00 ** | 1 | ||||
| MP | 0.471 ** | 0.941 ** | 0.817 ** | 0.817 ** | 1 | |||
| TOL | −0.157 * | 0.905 ** | 0.293 ** | 0.293 ** | 0.726 ** | 1 | ||
| HARM | 0.979 ** | 0.334 ** | 0.936 ** | 0.230 ** | 0.595 ** | 0.051 ns | 1 | |
| Marchouch | Yp | 0.137 * | 1 | |||||
| STI | 0.544 ** | 0.831 ** | 1 | |||||
| GMP | 0.544 ** | 0.831 ** | 1.00 ** | 1 | ||||
| MP | 0.241 ** | 0.978 ** | 0.907 ** | 0.907 ** | 1 | |||
| TOL | 0.30 ns | 0.977 ** | 0.728 ** | 0.728 ** | 0.916 ** | 1 | ||
| HARM | 0.950 ** | 0.373 ** | 0.728 ** | 0.728 ** | 0.468 ** | 0.265 ** | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El haddad, N.; Rajendran, K.; Smouni, A.; Es-Safi, N.E.; Benbrahim, N.; Mentag, R.; Nayyar, H.; Maalouf, F.; Kumar, S. Screening the FIGS Set of Lentil (Lens culinaris Medikus) Germplasm for Tolerance to Terminal Heat and Combined Drought-Heat Stress. Agronomy 2020, 10, 1036. https://doi.org/10.3390/agronomy10071036
El haddad N, Rajendran K, Smouni A, Es-Safi NE, Benbrahim N, Mentag R, Nayyar H, Maalouf F, Kumar S. Screening the FIGS Set of Lentil (Lens culinaris Medikus) Germplasm for Tolerance to Terminal Heat and Combined Drought-Heat Stress. Agronomy. 2020; 10(7):1036. https://doi.org/10.3390/agronomy10071036
Chicago/Turabian StyleEl haddad, Noureddine, Karthika Rajendran, Abdelaziz Smouni, Nour Eddine Es-Safi, Nadia Benbrahim, Rachid Mentag, Harsh Nayyar, Fouad Maalouf, and Shiv Kumar. 2020. "Screening the FIGS Set of Lentil (Lens culinaris Medikus) Germplasm for Tolerance to Terminal Heat and Combined Drought-Heat Stress" Agronomy 10, no. 7: 1036. https://doi.org/10.3390/agronomy10071036
APA StyleEl haddad, N., Rajendran, K., Smouni, A., Es-Safi, N. E., Benbrahim, N., Mentag, R., Nayyar, H., Maalouf, F., & Kumar, S. (2020). Screening the FIGS Set of Lentil (Lens culinaris Medikus) Germplasm for Tolerance to Terminal Heat and Combined Drought-Heat Stress. Agronomy, 10(7), 1036. https://doi.org/10.3390/agronomy10071036