NaCl and Na2SO4 Salinities Have Different Impact on Photosynthesis and Yield-Related Parameters in Rice (Oryza sativa L.)

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
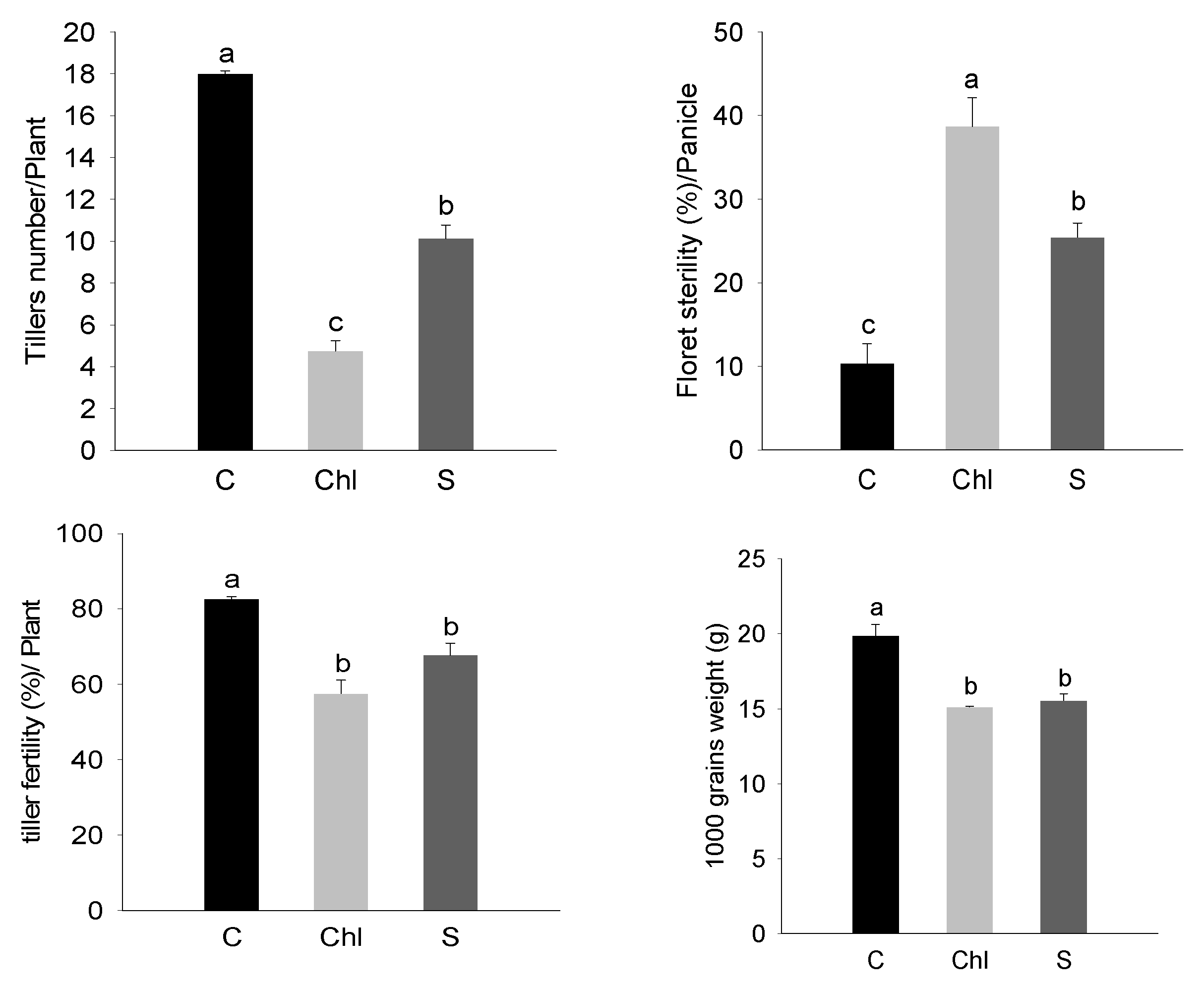
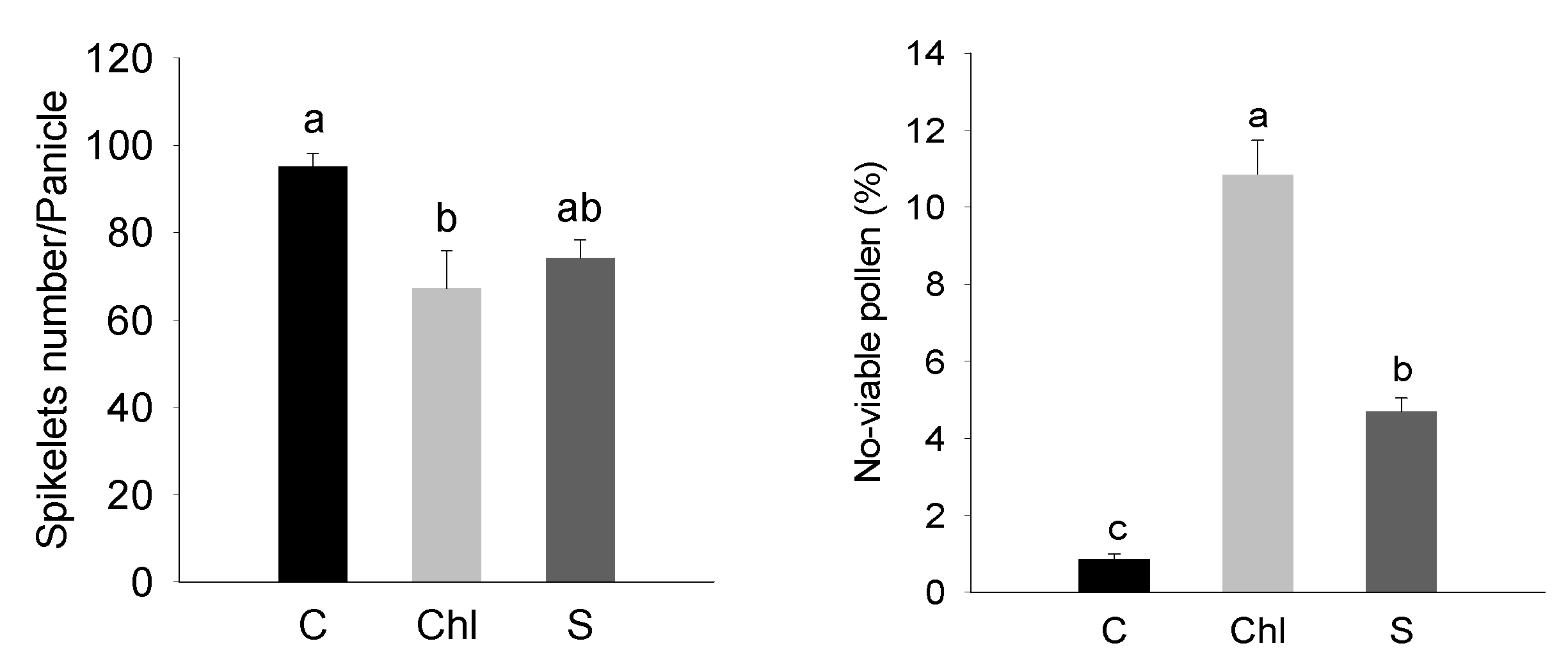
2.2. Estimation of Pollen Viability and Grain Ion Content
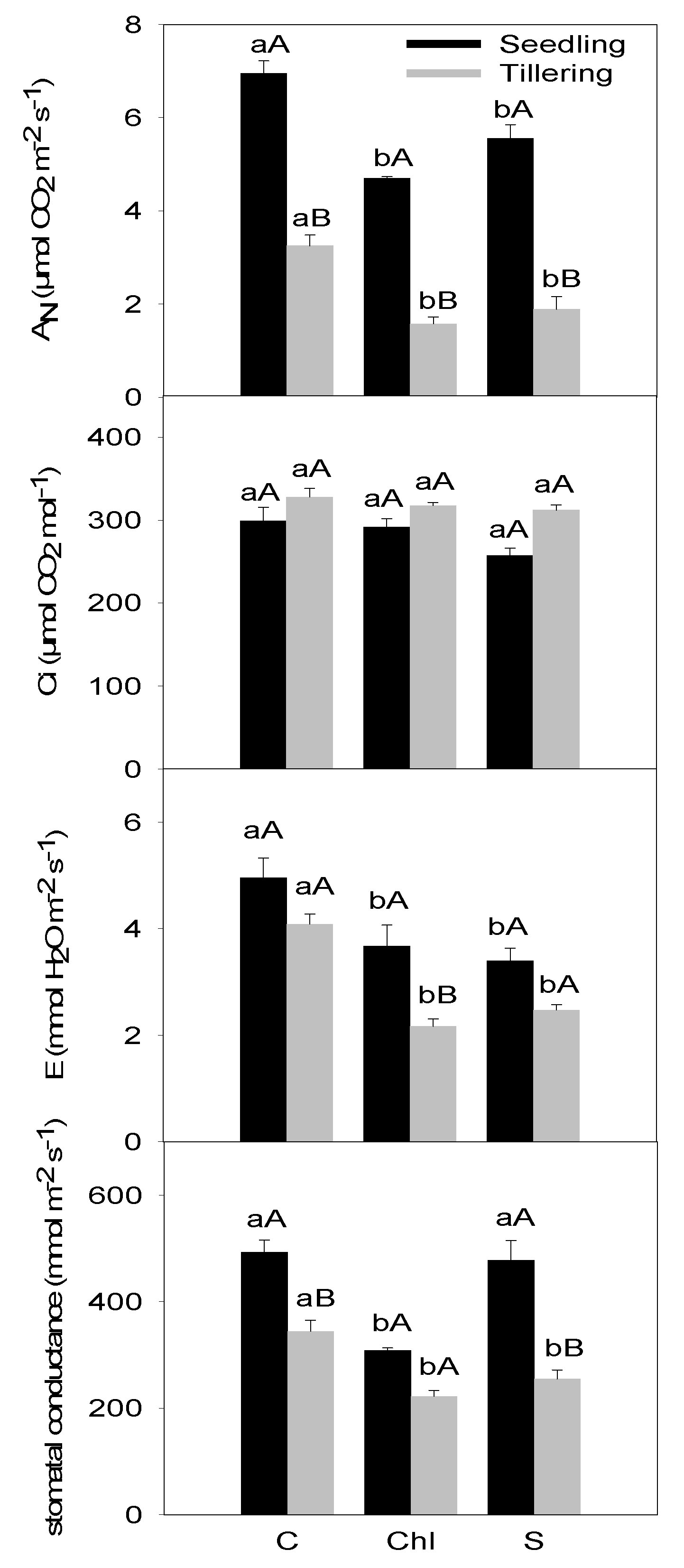
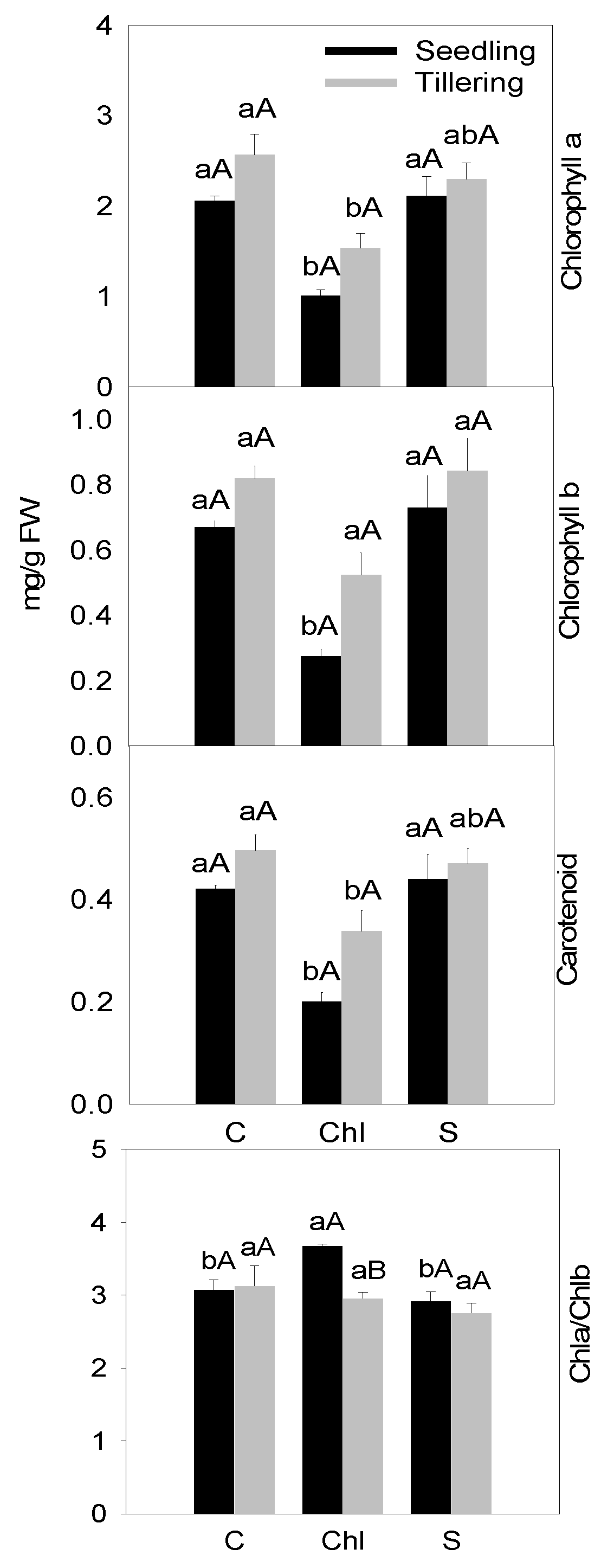
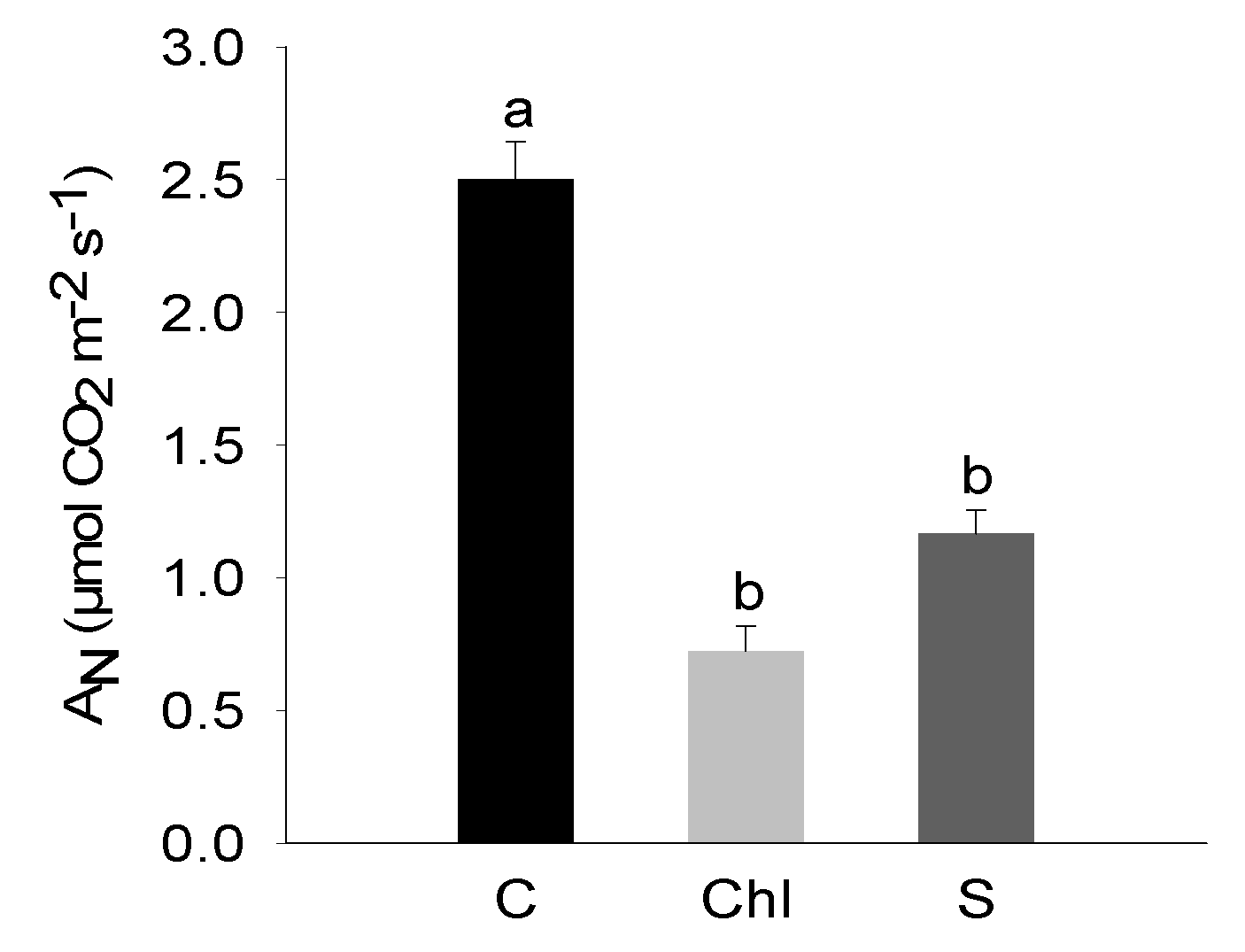
2.3. Photosynthesis-Related Parameters
2.4. Statistical Treatment of the Data
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bui, E. Soil salinity: A neglected factor in plant ecology and biogeography. J. Arid Environ. 2013, 92, 14–25. [Google Scholar] [CrossRef]
- Wassmann, R.; Nguyen, X.H.; Chu, T.H.; Tuong, T.P. Sea level rise affecting the Vietnamese Mekong Delta: Water elevation in the flood season and implications for rice production. Clim. Chang. 2004, 66, 89–107. [Google Scholar] [CrossRef]
- Grattan, S.R.; Zeng, L.; Shannon, M.C.; Roberts, S.R. Rice is more sensitive to salinity than previously thought. Calif. Agric. 2002, 56, 189–198. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot. 1995, 46, 1843–1852. [Google Scholar] [CrossRef]
- Moradi, F.; Ismail, A.M. Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging systems to salt stress during seedling and reproductive stages in rice. Ann. Bot. 2007, 99, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Shannon, M.C. Salinity effects on seedling growth and yield components of rice. Crop. Sci. 2000, 40, 996–1003. [Google Scholar] [CrossRef]
- Zhu, G.; Kinet, J.M.; Lutts, S. Characterisation of rice (Oryza sativa) F3 populations selected for salt resistance. 2. Relationships between yield-related parameters and physiological properties. Austr. J. Exp. Agric. 2004, 44, 333–342. [Google Scholar] [CrossRef]
- Zhu, G.; Kinet, J.M.; Lutts, S. Characterization of rice (Oryza sativa L.) F3 populations selected for salt resistance. I. Physiological behaviour during vegetative growth. Euphytica 2001, 121, 251–263. [Google Scholar] [CrossRef]
- Abdullah, Z.; Khan, M.A.; Flowers, T. Causes of sterility in seed set of rice under salinity stress. J. Agron. Crop Sci. 2001, 187, 25–32. [Google Scholar] [CrossRef]
- Rao, P.S.; Mishra, B.; Gupta, S.R.; Rathore, A. Reproductive stage tolerance to salinity and alkalinity stresses in rice genotypes. Plant Breed. 2008, 127, 256–261. [Google Scholar] [CrossRef]
- Sultana, N.; Ikeda, T.; Itoh, R. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. Environ. Exp. Bot. 1999, 42, 211–220. [Google Scholar] [CrossRef]
- Zeng, L.; Lesch, S.M.; Grieve, C.M. Rice growth and yield respond to changes in water depth and salinity stress. Agric. Water Manag. 2003, 59, 67–75. [Google Scholar] [CrossRef]
- Zeng, L.; Shannon, M.C.; Lesch, S.M. Timing of salinity stress affects rice growth and yield components. Agric. Water Manag. 2001, 48, 191–206. [Google Scholar] [CrossRef]
- Flowers, T.; Yeo, A.R. Variability in the resistance of sodium chloride salinity within rice (Oryza sativa L.) varieties. New Phytol. 1981, 88, 363–373. [Google Scholar] [CrossRef]
- Kaddah, M.T. Salinity effects on growth of rice at the seedling and inflorescence stages of development. Soil Sci. 1963, 96, 105–111. [Google Scholar] [CrossRef]
- Pardo, J.M. Biotechnology of water and salinity stress tolerance. Curr. Opin. Biotechnol. 2010, 21, 185–196. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. NaCl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Castillo, E.G.; Tuong, T.P.; Ismail, A.M.; Inubushi, K. Response to salinity in rice: Comparative effects of osmotic and ionic stresses. Plant Prod. Sci. 2007, 10, 159–170. [Google Scholar] [CrossRef]
- Khatun, S.; Flowers, T. Effects of salinity on seed set in rice. Plant Cell Environ. 1995, 18, 61–67. [Google Scholar] [CrossRef]
- Samson, M.E.; Fortin, J.; Pepin, S.; Caron, J. Impact of potassium sulfate salinity on growth and development of cranberry plants subjected to overhead and subirrigation. Can. J. Soil Sci. 2016, 97, 20–30. [Google Scholar] [CrossRef]
- Khatun, S.; Flowers, T. The estimation of pollen viability in rice. J. Exp. Bot. 1995, 46, 151–154. [Google Scholar] [CrossRef]
- Aghajanzadeh, T.A.; Reich, M.; Kopriva, S.; De kok, L.J. Impact of chloride (NaCl, KCl) and sulphate (Na2SO4, K2SO4) salinity on glucosinolate metabolism in Brassica rapa. J. Agron. Crop. Sci. 2018, 204, 137–146. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D.; Chen, Y. Differentiation of carbonate, chloride, and sulfate salinity responses in tall fescue. Sci. Hort. 2012, 139, 1–7. [Google Scholar] [CrossRef]
- Irakoze, W.; Vanpee, B.; Rufyikiri, G.; Dailly, H.; Nijimbere, S.; Lutts, S. Comparative effects of chloride and sulfate salinities on two contrasting rice cultivars (Oryza sativa L.) at the seedling stage. J. Plant Nutr. 2019, 42, 1001–1015. [Google Scholar] [CrossRef]
- Cock, J.; Yoshida, S.; Forno, D.A. Laboratory Manual for Physiological Studies of Rice; International Rice Research Institute: Los Baños, Philippines, 1976. [Google Scholar]
- Alexander, M. Differential staining of aborted and nonaborted pollen. Stain Technol. 1969, 44, 117–122. [Google Scholar] [CrossRef]
- Hamrouni, L.; Hanana, M.; Abdelly, C.; Ghorbel, C. Exclusion du chlorure et inclusion du sodium: Deux mécanismes concomitants de tolérance à la salinité chez la vigne sauvage Vitis vinifera subsp. sylvestris (var.’Séjnène.). Biotech. Agron. Soc. Environ. 2011, 15, 387. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 350–382. [Google Scholar] [CrossRef]
- Krishnamurthy, S.L.; Gautam, R.K.; Sharma, P.C.; Sharma, D.K. Effect of different salt stresses on agro-morphological traits and utilisation of salt stress indices for reproductive stage salt tolerance in rice. Field Crop. Res. 2016, 190, 26–33. [Google Scholar] [CrossRef]
- Devinar, G.; Llanes, A.; Masciarelli, O.; Luna, V. Different relative humidity conditions combined with chloride and sulfate salinity treatments modify abscisic acid and salicylic acid levels in the halophyte Prosopis strombulifera. Plant Growth Regul. 2013, 70, 247–256. [Google Scholar] [CrossRef]
- Han, L.; Gao, Y.; Li, D. Ion uptake in tall fescue as affected by carbonate, chloride, and sulfate salinity. PLoS ONE 2014, 9, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Reich, M.; Aghajanzadeh, T.; Helm, J.; Parmar, S.; Hawkesford, M.J.; De kok, L.J. Chloride and sulfate salinity differently affect biomass, mineral nutrient composition and expression of sulfate transport and assimilation genes in Brassica rapa. Plant Soil 2017, 411, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.; Grieve, C.; Shannon, M. The response of lucerne (Medicago sativa L.) to sodium sulphate and chloride salinity. Plant Soil 1998, 202, 271–280. [Google Scholar] [CrossRef]
- Bilski, J.J.; Nelson, D.; Conlon, R.L. Response of six wild potato species to chloride and sulfate salinity. Amer. Pot. J. 1988, 65, 605–611. [Google Scholar] [CrossRef]
- Chunthaburee, S.; Sanitchon, J.; Pattanagul, W.; Theerakulpisut, P. Effects of salt stress after late booting stage on yield and antioxidant capacity in pigmented rice grains and alleviation of the salt-induced yield reduction by exogenous spermidine. Plant Prod. Sci. 2015, 18, 32–42. [Google Scholar] [CrossRef]
- Lekklar, C.; Pongpanich, M.; Suriya-arunroj, D.; Chinpongpanich, A.; Tsai, H.; Comai, L.; Chadchawan, S.; Buaboocha, T. Genome-wide association study for salinity tolerance at the flowering stage in a panel of rice accessions from Thailand. BMC Genom. 2019, 20, 76–85. [Google Scholar] [CrossRef]
- Thitisaksakul, M.; Tananuwong, K.; Shoemaker, C.F.; Chun, A.; Tanadul, O.; Labavitch, J.M.; Beckles, D.M. Effects of timing and severity of salinity stress on rice (Oryza sativa L.) yield, grain composition, and starch functionality. J. Agric. Food Chem. 2015, 63, 2296–2304. [Google Scholar] [CrossRef]
- Ma, Y.; Shabala, S.; Li, C.; Liu, C.; Zhang, W.; Zhou, M. Quantitative trait loci for salinity tolerance identified under drained and waterlogged conditions and their association with flowering time in barley (Hordeum vulgare L). PLoS ONE 2015, 10, 456–469. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A role for GIGANTEA: Keeping the balance between flowering and salinity stress tolerance. Plant Signal. Behav. 2013, 8, 24820. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Elteren, J.V.; Albacete, A.; Quinet, M.; Martínez-Andújar, C.; Kinet, J.M.; Pérez-Alfocea, F.; Lutts, S. Impact of salinity on early reproductive physiology of tomato (Solanum lycopersicum) in relation to a heterogeneous distribution of toxic ions in flower organs. Funct. Plant Biol. 2009, 36, 125–136. [Google Scholar] [CrossRef]
- Zonia, L.; Cordeiro, S.; Tupý, J.; Feijó, J.A. Oscillatory chloride efflux at the pollen tube apex has a role in growth and cell volume regulation and is targeted by inositol 3, 4, 5, 6-tetrakisphosphate. Plant Cell 2002, 14, 2233–2249. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, N.; Usuda, H.; Nakamoto, H.; Ishihara, K. Changes in the rate of photosynthesis during grain filling and the enzymatic activities associated with the photosynthetic carbon metabolism in rice panicles. Plant Cell Physiol. 1990, 31, 835–844. [Google Scholar] [CrossRef]
- Vromman, D.; Lutts, S.; Lefèvre, I.; Somer, L.; De Vreese, O.; Šlejkovec, Z.; Quinet, M. Effects of simultaneous arsenic and iron toxicities on rice (Oryza sativa L.) development, yield-related parameters and As and Fe accumulation in relation to As speciation in the grains. Plant Soil 2013, 371, 199–217. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mmol CO2 Uptake/mmol H2O Loss | ||||||
|---|---|---|---|---|---|---|
| Seedling | Tillering | |||||
| C | Chl | S | C | Chl | S | |
| WUE | 1.4 ± 0.09 aA | 1.3 ± 0.14 Aa | 1.6 ± 0.12 aA | 0.8 ± 0.08 aB | 0.7 ± 0.09 aB | 0.8 ± 0.08 aB |
| µmol g−1 DW | ||||||
|---|---|---|---|---|---|---|
| Grain Hull | Grain | |||||
| Ion | C | Chl | S | C | Chl | S |
| Na+ | 42.8 ± 3.5 c | 503.4 ± 3.2 a | 314 ± 36.5 b | 5.4 ± 0.06 c | 51.7 ± 2.3 a | 22.9 ± 3.5 b |
| K+ | 920 ± 30 a | 534.9 ± 15 b | 550 ± 24 b | 226 ± 1.5 a | 141 ± 3.6 b | 146 ± 7 b |
| Cl− | 199.3 ± 13 b | 1455.6 ± 27 a | 188 ± 14 b | 6.7 ± 0.08 b | 64.6 ± 3 a | 7 ± 0.7 b |
| S6+ | 48.7 ± 5.6 b | 57.2 ± 4.3 b | 284 ± 17 a | 16.4 ± 1 b | 15 ± 0.1 b | 25 ± 2 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irakoze, W.; Prodjinoto, H.; Nijimbere, S.; Rufyikiri, G.; Lutts, S. NaCl and Na2SO4 Salinities Have Different Impact on Photosynthesis and Yield-Related Parameters in Rice (Oryza sativa L.). Agronomy 2020, 10, 864. https://doi.org/10.3390/agronomy10060864
Irakoze W, Prodjinoto H, Nijimbere S, Rufyikiri G, Lutts S. NaCl and Na2SO4 Salinities Have Different Impact on Photosynthesis and Yield-Related Parameters in Rice (Oryza sativa L.). Agronomy. 2020; 10(6):864. https://doi.org/10.3390/agronomy10060864
Chicago/Turabian StyleIrakoze, Willy, Hermann Prodjinoto, Séverin Nijimbere, Gervais Rufyikiri, and Stanley Lutts. 2020. "NaCl and Na2SO4 Salinities Have Different Impact on Photosynthesis and Yield-Related Parameters in Rice (Oryza sativa L.)" Agronomy 10, no. 6: 864. https://doi.org/10.3390/agronomy10060864
APA StyleIrakoze, W., Prodjinoto, H., Nijimbere, S., Rufyikiri, G., & Lutts, S. (2020). NaCl and Na2SO4 Salinities Have Different Impact on Photosynthesis and Yield-Related Parameters in Rice (Oryza sativa L.). Agronomy, 10(6), 864. https://doi.org/10.3390/agronomy10060864