Chromosome Engineering in Tropical Cash Crops


Abstract

1. Introduction
2. Fertility and Enhanced Meiotic Pairing

{kind=link}
{kind=link}
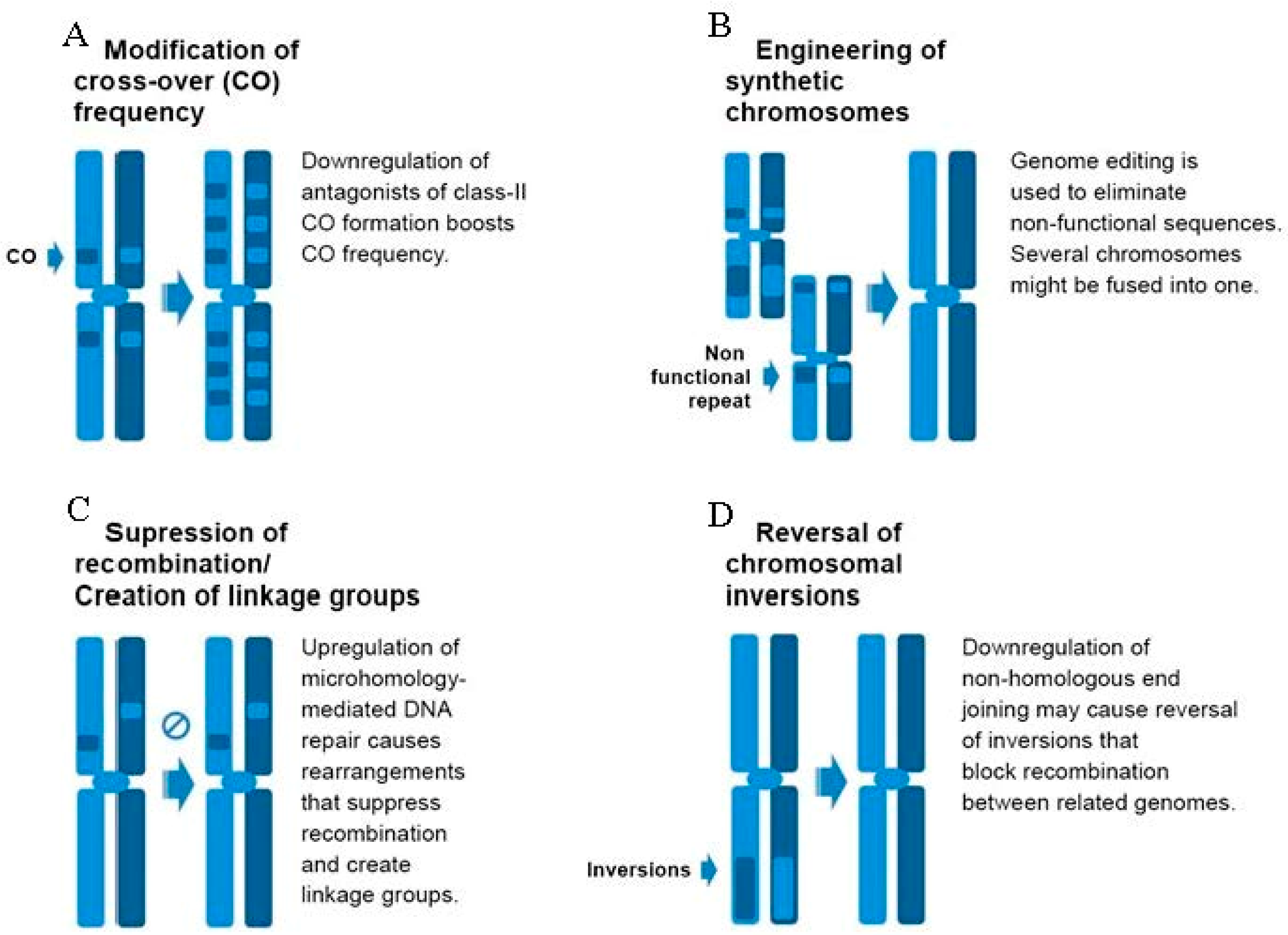{kind=link}
| Species | Genome Database | Suggested Method for Engineering | Estimated Time for Engineering and Regeneration of Plants |
|---|---|---|---|
| T. cacao L., Criollo genotype (B97-61/B2) | The Cocoa Genome Hub, http://cocoa-genome-hub.southgreen.fr/ [11] | Agrobacterium-mediated transient transformation with CRISPR/Cas9 of cacao leaves and cotyledon cells [17]. | Five years for seedlings to reach sexual maturity [16]. Time required to perform engineering is unknown. |
| C. canephora P., accession DH200-94 | Coffee Genome Hub, http://coffee-genome.org [20] | Cocultivation of embryogenic calli of Coffea canephora with Agrobacterium [26]. | Six to 8 years to reach sexual maturity in Coffea arabica [27]. In C. canephora, induction of primary calli takes 1 month and induction of embryogenic calli takes an additional eight months. The selection of transformants takes at least one month, whereas plantlet regeneration may take another month. Total minimum estimated time: 11 months [26]. |
| C. papaya, ‘Sun Up’ | Carica papaya ASGPBv0.4, https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Cpapaya [28] | Agrobacterium-mediated transformation of somatic and zygotic embryos [29,30]. | Four months for plantlets to reach maturity [8]. Transformation takes a long time [31]. Following field cultivation of line Tainung #2 for one year, papaya shoots were collected and rooted in vitro with indole-3-butyric acid (time was not reported) [31]. Shoots were then cultured for six weeks to harvest adventitious roots [31]. Roots were cultured in vitro for three months to obtain somatic embryos [31]. Embryos were cocultured for two days and selected for 80 days [31]. Regenerants were cultured for one week to induce shooting and transferred to vermiculite in the greenhouse [31]. Estimated time: approximately one year and seven months. |
3. Mechanisms of Meiosis and Fertility
4. Possibilities for Genome Editing in Coffee, Cacao and Papaya
5. Chromosome Structure Editing in Crops
6. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Dhankher, O.P.; Foyer, C.H. Climate Resilient Crops for Improving Global Food Security and Safety. Plant Cell Environ. 2018, 41, 877–884. [Google Scholar] [CrossRef]
- Tito, R.; Vasconcelos, H.L.; Feeley, K.J. Global Climate Change Increases Risk of Crop Yield Losses and Food Insecurity in the Tropical Andes. Glob. Chang. Biol. 2018, 24, e592–e602. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.M.; Hansen, J.; Rioux, J.; Stirling, C.M.; Twomlow, S. Wollenberg. Urgent Action to Combat Climate Change and Its Impacts (SDG 13): Transforming Agriculture and Food Systems. Curr. Opin. Environ. Sustain. 2018, 34, 13–20. [Google Scholar] [CrossRef]
- Davis, A.P.; Chadburn, H.; Moat, J.; O’Sullivan, R.; Hargreaves, S.; Lughadha, E.N. High Extinction Risk for Wild Coffee Species and Implications for Coffee Sector Sustainability. Sci. Adv. 2019, 5, eaav3473. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.D.; Rhiney, K.; Eitzinger, A.; Umaharan, P. Climate Adaptation in a Minor Crop Species: Is the Cocoa Breeding Network Prepared for Climate Change? Agroecol. Sustain. Food Syst. 2018, 42, 812–833. [Google Scholar] [CrossRef]
- Schroth, G.; Läderach, P.; Martinez-Valle, A.I.; Bunn, C.; Jassogne, L. Vulnerability to Climate Change of Cocoa in West Africa: Patterns, Opportunities and Limits to Adaptation. Sci. Total Environ. 2016, 556, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Van der Vossen, H.; Bertrand, B.; Charrier, A. Next Generation Variety Development for Sustainable Production of Arabica Coffee (Coffea Arabica, L.): A Review. Euphytica 2015, 204, 243–256. [Google Scholar] [CrossRef]
- Liao, Z.; Yu, Q.; Ming, R. Development of Male-Specific Markers and Identification of Sex Reversal Mutants in Papaya. Euphytica 2017, 213, 53. [Google Scholar] [CrossRef]
- Campostrini, E.; Schaffer, B.; Ramalho, J.D.C.; González, J.C.; Rodrigues, W.P.; Da Silva, J.R.; Lima, R.S.N. Environmental Factors Controlling Carbon Assimilation, Growth, and Yield of Papaya (Carica Papaya L.) Under Water-Scarcity Scenarios. In Water Scarcity and Sustainable Agriculture in Semiarid Environment; Elsevier: Amsterdam, The Netherlands, 2018; pp. 481–500. [Google Scholar] [CrossRef]
- Gaur, R.K.; Verma, R.K.; Khurana, S.M.P. Genetic Engineering of Horticultural Crops. In Genetic Engineering of Horticultural Crops; Academic Press: Cambridge, MA, USA, 2018; pp. 23–46. [Google Scholar] [CrossRef]
- Argout, X.; Martin, G.; Droc, G.; Fouet, O.; Labadie, K.; Rivals, E.; Aury, J.M.; Lanaud, C. The Cacao Criollo Genome v2.0: An Improved Version of the Genome for Genetic and Functional Genomic Studies. BMC Genom. 2017, 18, 730. [Google Scholar] [CrossRef]
- Argout, X.; Salse, J.; Aury, J.M.; Guiltinan, M.J.; Droc, G.; Gouzy, J.; Allegre, M.; Chaparro, C.; Legavre, T.; Maximova, S.N.; et al. The Genome of Theobroma Cacao. Nat. Genet. 2011, 43, 101–108. [Google Scholar] [CrossRef]
- Cornejo, O.E.; Yee, M.-C.; Dominguez, V.; Andrews, M.; Sockell, A.; Strandberg, E.; Livingstone, D.; Stack, C.; Romero, A.; Umaharan, P.; et al. Population Genomic Analyses of the Chocolate Tree, Theobroma cacao, L., Provide Insights into Its Domestication Process. Commun. Biol. 2018, 1, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, E.J.; Motamayor, J.C.; Cornejo, O.E. Genetic Differentiation and Intrinsic Genomic Features Explain Variation in Recombination Hotspots among Cocoa Tree Populations. bioRxiv 2018. [Google Scholar] [CrossRef]
- Dantas, L.G.; Guerra, M. Chromatin Differentiation between Theobroma Cacao L. and T. Grandiflorum Schum. Genet. Mol. Biol. 2010, 33, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Wickramasuriya, A.M.; Dunwell, J.M. Cacao Biotechnology: Current Status and Future Prospects. Plant Biotechnol. J. 2018, 16, 4–17. [Google Scholar] [CrossRef]
- Fister, A.S.; Landherr, L.; Maximova, S.N.; Guiltinan, M.J. Transient Expression of CRISPR/Cas9 Machinery Targeting TcNPR3 Enhances Defense Response in Theobroma Cacao. Front. Plant Sci. 2018, 9, 268. [Google Scholar] [CrossRef]
- Denoeud, F.; Carretero-Paulet, L.; Dereeper, A.; Droc, G.; Guyot, R.; Pietrella, M.; Zheng, C.; Alberti, A.; Anthony, F.; Aprea, G.; et al. The Coffee Genome Provides Insight into the Convergent Evolution of Caffeine Biosynthesis. Science 2014, 345, 1181–1184. [Google Scholar] [CrossRef]
- Pinto-Maglio, C.A.F. Cytogenetics of Coffee. Braz. J. Plant Physiol. 2006, 18, 37–44. [Google Scholar] [CrossRef]
- Dereeper, A.; Bocs, S.; Rouard, M.; Guignon, V.; Ravel, S.; Tranchant-Dubreuil, C.; Poncet, V.; Garsmeur, O.; Lashermes, P.; Droc, G. The Coffee Genome Hub: A Resource for Coffee Genomes. Nucleic Acids Res. 2015, 43, D1028–D1035. [Google Scholar] [CrossRef]
- Herrera, J.C.; D’Hont, A.; Lashermes, P. Use of Fluorescence in Situ Hybridization as a Tool for Introgression Analysis and Chromosome Identification in Coffee (Coffea Arabica L.). Genome 2007, 50, 619–626. [Google Scholar] [CrossRef]
- Moncada, M.D.P.; Tovar, E.; Montoya, J.C.; González, A.; Spindel, J.; McCouch, S. A Genetic Linkage Map of Coffee (Coffea Arabica L.) and QTL for Yield, Plant Height, and Bean Size. Tree Genet. Genomes 2016, 12, 1–17. [Google Scholar] [CrossRef]
- Ming, R.; Hou, S.; Feng, Y.; Yu, Q.; Dionne-Laporte, A.; Saw, J.H.; Senin, P.; Wang, W.; Ly, B.V.; Lewis, K.L.T.; et al. The Draft Genome of the Transgenic Tropical Fruit Tree Papaya (Carica Papaya Linnaeus). Nature 2008, 452, 991–997. [Google Scholar] [CrossRef] [PubMed]
- VanBuren, R.; Ming, R. Dynamic Transposable Element Accumulation in the Nascent Sex Chromosomes of Papaya. Mob. Genet. Elem. 2013, 3, e23462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Jiang, J. Molecular Cytogenetics of Papaya. In Genetics and Genomics of Papaya, Plant Genetics and Genomics: Crops and Models 10; Ming, R., Moore, P.H., Eds.; Springer Science + Business Media: New York, NY, USA, 2014; pp. 157–168. [Google Scholar] [CrossRef]
- Breitler, J.C.; Dechamp, E.; Campa, C.; Zebral Rodrigues, L.A.; Guyot, R.; Marraccini, P.; Etienne, H. CRISPR/Cas9-Mediated Efficient Targeted Mutagenesis Has the Potential to Accelerate the Domestication of Coffea Canephora. Plant Cell Tissue Organ Cult. 2018, 134, 383–394. [Google Scholar] [CrossRef]
- Sera, T. Coffee Genetic Breeding at IAPAR. Crop Breed. Appl. Biotechnol. 2001, 1, 179–199. [Google Scholar] [CrossRef]
- Jiménez, V.M.; Mora-Newcomer, E.; Gutiérrez-Soto, M.V. Biology of the Papaya Plant. In Genetics and Genomics of Papaya, Plant Genetics and Genomics: Crops and Models 10; Ming, R., Moore, P.H., Eds.; Springer Science + Business Media: New York, NY, USA, 2014; pp. 17–33. [Google Scholar] [CrossRef]
- Geetika, S.; Ruqia, M.; Harpreet, K.; Neha, D.; Shruti, K.; Singh, S.P. Genetic Engineering in Papaya. In Genetic Engineering of Horticultural Crops; Academic Press: Cambridge, MA, USA, 2018; pp. 137–154. [Google Scholar] [CrossRef]
- Hamim, I.; Borth, W.B.; Marquez, J.; Green, J.C.; Melzer, M.J.; Hu, J.S. Transgene-Mediated Resistance to Papaya Ringspot Virus: Challenges and Solutions. Phytoparasitica 2018, 46, 1–18. [Google Scholar] [CrossRef]
- Kung, Y.J.; Yu, T.A.; Huang, C.H.; Wang, H.C.; Wang, S.L.; Yeh, S.D. Generation of Hermaphrodite Transgenic Papaya Lines with Virus Resistance via Transformation of Somatic Embryos Derived from Adventitious Roots of in Vitro Shoots. Transgenic Res. 2010, 19, 621–635. [Google Scholar] [CrossRef]
- VanBuren, R.; Zeng, F.; Chen, C.; Zhang, J.; Wai, C.M.; Han, J.; Aryal, R.; Gschwend, A.R.; Wang, J.; Na, J.K.; et al. Origin and Domestication of Papaya Yh Chromosome. Genome Res. 2015, 25, 524–533. [Google Scholar] [CrossRef]
- Chen, J.R.; Urasaki, N.; Matsumura, H.; Chen, I.C.; Lee, M.J.; Chang, H.J.; Chung, W.C.; Ku, H.M. Dissecting the All-Hermaphrodite Phenomenon of a Rare X Chromosome Mutant in Papaya (Carica Papaya L.). Mol. Breed. 2019, 39, 14. [Google Scholar] [CrossRef]
- Lee, C.Y.; Lin, H.J.; Viswanath, K.K.; Lin, C.P.; Chang, B.C.H.; Chiu, P.H.; Chiu, C.T.; Wang, R.H.; Chin, S.W.; Chen, F.C. The Development of Functional Mapping by Three Sex-Related Loci on the Third Whorl of Different Sex Types of Carica papaya L. PLoS ONE 2018, 13, e0194605. [Google Scholar] [CrossRef]
- Fayos, I.; Mieulet, D.; Petit, J.; Meunier, A.C.; Périn, C.; Nicolas, A.; Guiderdoni, E. Engineering Meiotic Recombination Pathways in Rice. Plant Biotechnol. J. 2019, 1–16. [Google Scholar] [CrossRef]
- Wang, Y.; Copenhaver, G.P. Meiotic Recombination: Mixing It up in Plants. Annu. Rev. Plant Biol. 2018, 69, 577–609. [Google Scholar] [CrossRef] [PubMed]
- Mieulet, D.; Aubert, G.; Bres, C.; Klein, A.; Droc, G.; Vieille, E.; Rond-Coissieux, C.; Sanchez, M.; Dalmais, M.; Mauxion, J.-P.; et al. Unleashing Meiotic Crossovers in Crops. Nat. Plants 2018, 4, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Que, Q.; Chen, Z.; Kelliher, T.; Skibbe, D.; Dong, S.; Chilton, M.-D. Nd Their Applications in Genome Engineering. In Plant Genome Editing with CRISPR Systems; Qi, Y., Ed.; Springer Science + Business Media, LLC: New York, NY, USA, 2019; pp. 3–24. [Google Scholar] [CrossRef]
- Que, Q.; Chilton, M.D.M.; Elumalai, S.; Zhong, H.; Dong, S.; Shi, L. Repurposing Macromolecule Delivery Tools for Plant Genetic Modification in the Era of Precision Genome Engineering. In Methods in Molecular Biology; Springer Science + Business Media, LLC: New York, NY, USA, 2019; pp. 3–18. [Google Scholar] [CrossRef]
- Schmidt, C.; Schindele, P.; Puchta, H. From Gene Editing to Genome Engineering: Restructuring Plant Chromosomes via CRISPR/Cas. aBIOTECH 2019. [Google Scholar] [CrossRef]
- Richardson, S.M.; Mitchell, L.A.; Stracquadanio, G.; Yang, K.; Dymond, J.S.; DiCarlo, J.E.; Lee, D.; Huang, C.L.V.; Chandrasegaran, S.; Cai, Y.; et al. Design of a Synthetic Yeast Genome. Science 2017, 355, 1040–1044. [Google Scholar] [CrossRef]
- Ostrov, N.; Beal, J.; Ellis, T.; Gordon, D.B.; Karas, B.J.; Lee, H.H.; Lenaghan, S.C.; Schloss, J.A.; Stracquadanio, G.; Trefzer, A.; et al. Technological Challenges and Milestones for Writing Genomes. Science 2019, 366, 310–312. [Google Scholar] [CrossRef]
- Shao, Y.; Lu, N.; Xue, X.; Qin, Z. Creating Functional Chromosome Fusions in Yeast with CRISPR–Cas9. Nat. Protoc. 2019, 14, 2521–2545. [Google Scholar] [CrossRef]
- Xu, J.; Hua, K.; Lang, Z. Genome Editing for Horticultural Crop Improvement. Hortic. Res. 2019, 6, 113. [Google Scholar] [CrossRef]
- Xu, X.; Duan, D.; Chen, S.J. CRISPR-Cas9 Cleavage Efficiency Correlates Strongly with TargetsgRNA Folding Stability: From Physical Mechanism to off-Target Assessment. Sci. Rep. 2017, 7, 143. [Google Scholar] [CrossRef]
- Feng, C.; Su, H.; Bai, H.; Wang, R.; Liu, Y.; Guo, X.; Liu, C.; Zhang, J.; Yuan, J.; Birchler, J.A.; et al. High-Efficiency Genome Editing Using a Dmc1 Promoter-Controlled CRISPR/Cas9 System in Maize. Plant Biotechnol. J. 2018, 16, 1848–1857. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-Replace Genome Editing without Double-Strand Breaks or Donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Ribas, A.F.; Dechamp, E.; Champion, A.; Bertrand, B.; Combes, M.C.; Verdeil, J.L.; Lapeyre, F.; Lashermes, P.; Etienne, H. Agrobacterium-Mediated Genetic Transformation of Coffea arabica (L.) Is Greatly Enhanced by Using Established Embryogenic Callus Cultures. BMC Plant Biol. 2011, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Pacher, M.; Puchta, H. Efficient Induction of Heritable Inversions in Plant Genomes Using the CRISPR/Cas System. Plant J. 2019, 98, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Park, C.Y.; Sung, J.J.; Kim, D.W. Genome Editing of Structural Variations: Modeling and Gene Correction. Trends Biotechnol. 2016, 34, 548–561. [Google Scholar] [CrossRef] [PubMed]
- Narducci Da Silva, E.; Neto, M.F.; Pereira, T.N.S.; Pereira, M.G. Meiotic Behavior of Wild Caricaceae Species Potentially Suitable for Papaya Improvement. Crop Breed. Appl. Biotechnol. 2012, 12, 52–59. [Google Scholar] [CrossRef][Green Version]
- Khanday, I.; Skinner, D.; Yang, B.; Mercier, R.; Sundaresan, V. A Male-Expressed Rice Embryogenic Trigger Redirected for Asexual Propagation through Seeds. Nature 2019, 565, 91. [Google Scholar] [CrossRef]


© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolaños-Villegas, P. Chromosome Engineering in Tropical Cash Crops. Agronomy 2020, 10, 122. https://doi.org/10.3390/agronomy10010122
Bolaños-Villegas P. Chromosome Engineering in Tropical Cash Crops. Agronomy. 2020; 10(1):122. https://doi.org/10.3390/agronomy10010122
Chicago/Turabian StyleBolaños-Villegas, Pablo. 2020. "Chromosome Engineering in Tropical Cash Crops" Agronomy 10, no. 1: 122. https://doi.org/10.3390/agronomy10010122
APA StyleBolaños-Villegas, P. (2020). Chromosome Engineering in Tropical Cash Crops. Agronomy, 10(1), 122. https://doi.org/10.3390/agronomy10010122