
Metabolomic Profiling of Pulmonary Neuroendocrine Neoplasms

 , ,
, ,  , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Metabolomic Profiling
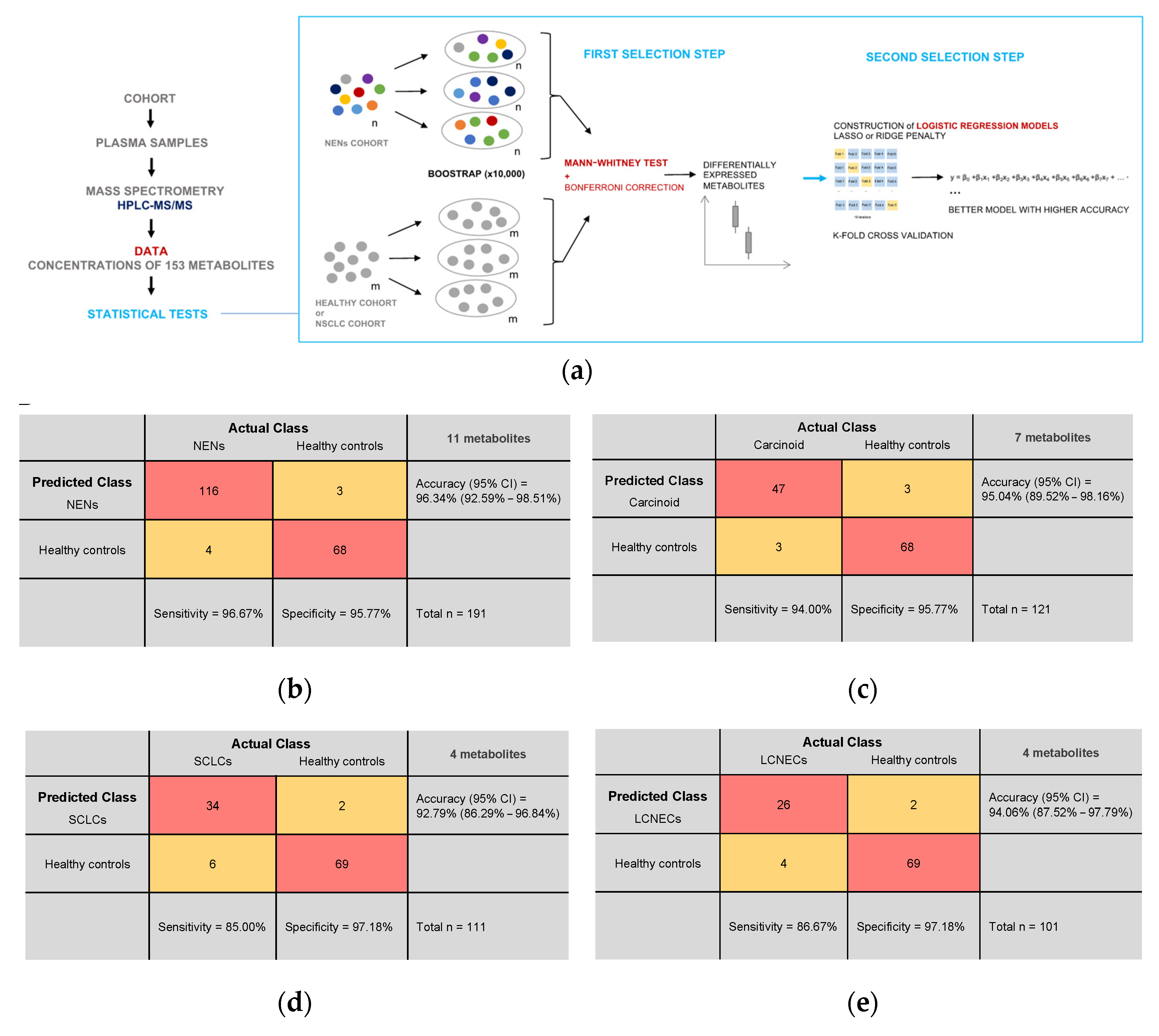
2.3. Statistical Analyses
2.4. Data Presentation
3. Results
3.1. Demographic and Clinical Characteristics
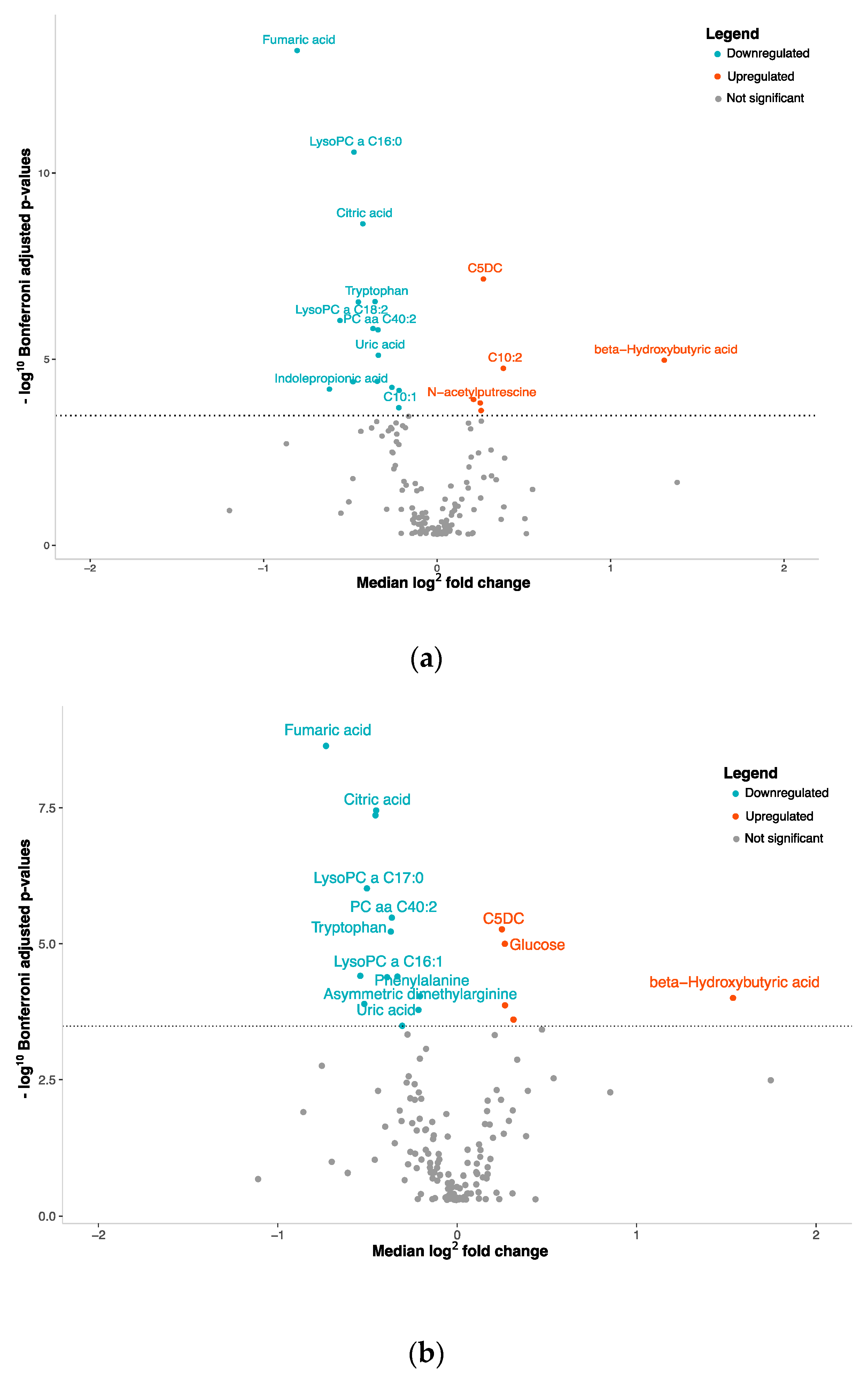
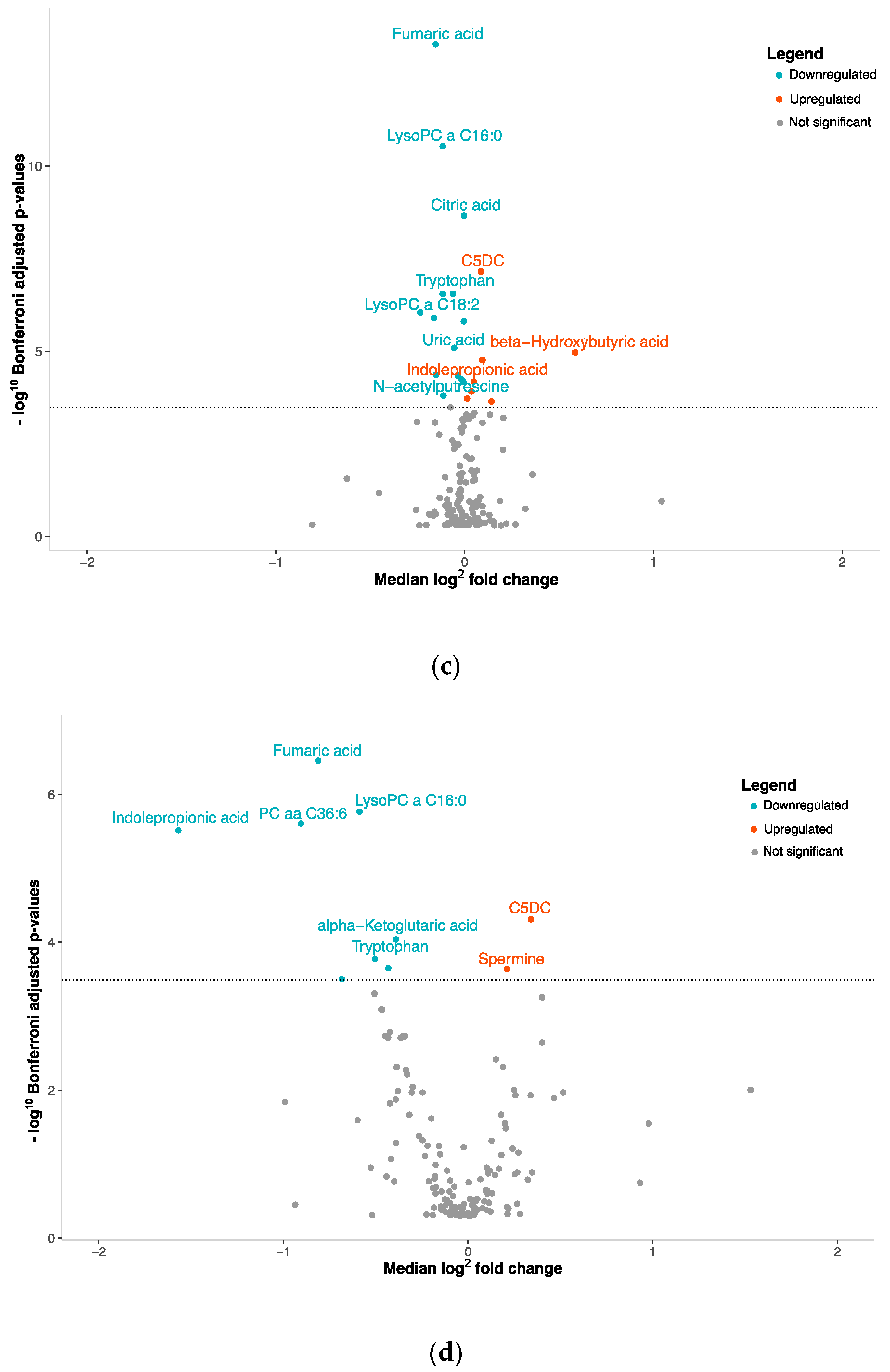
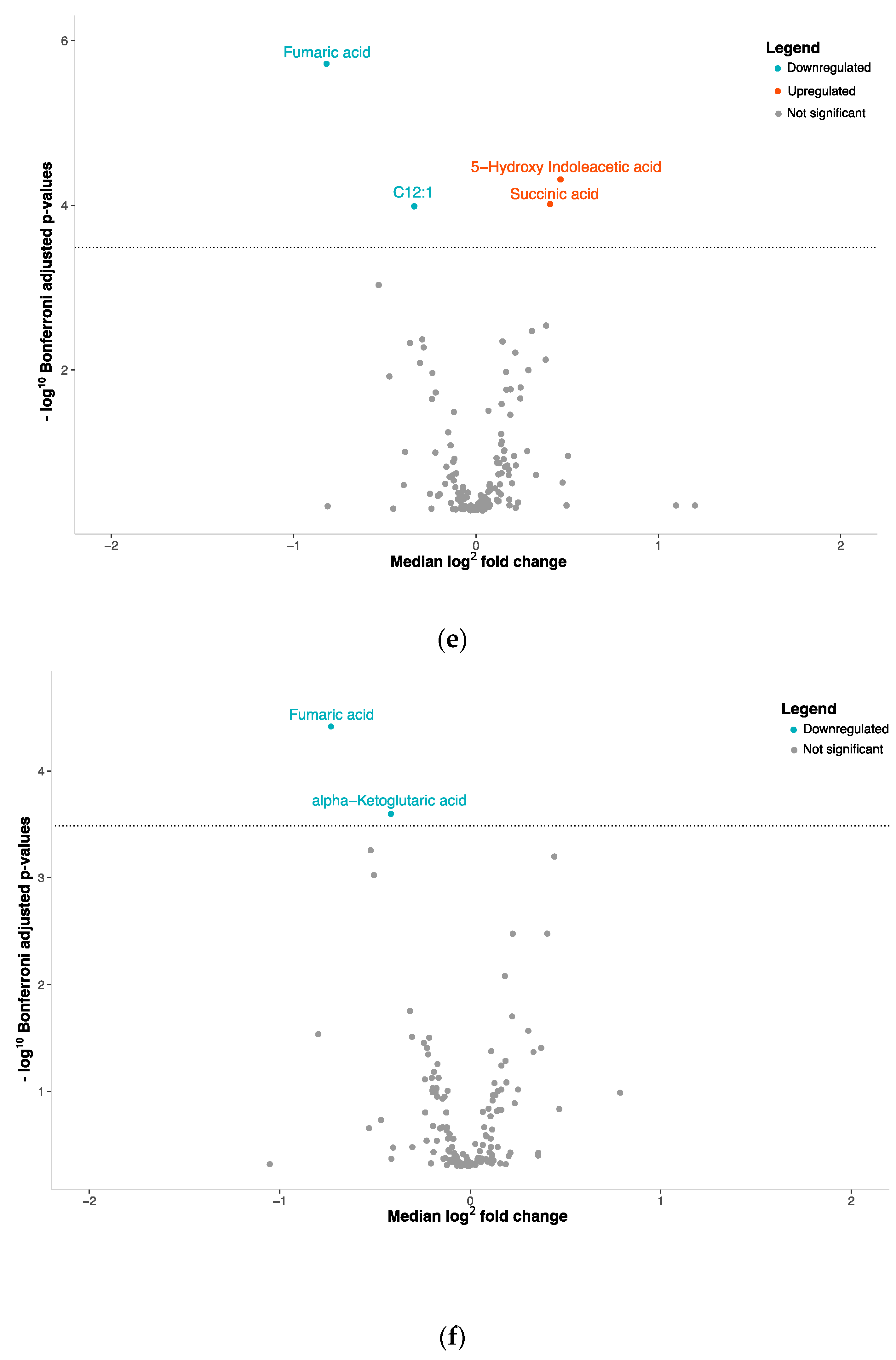
3.2. NENs Have a Distinct Plasmatic Profile
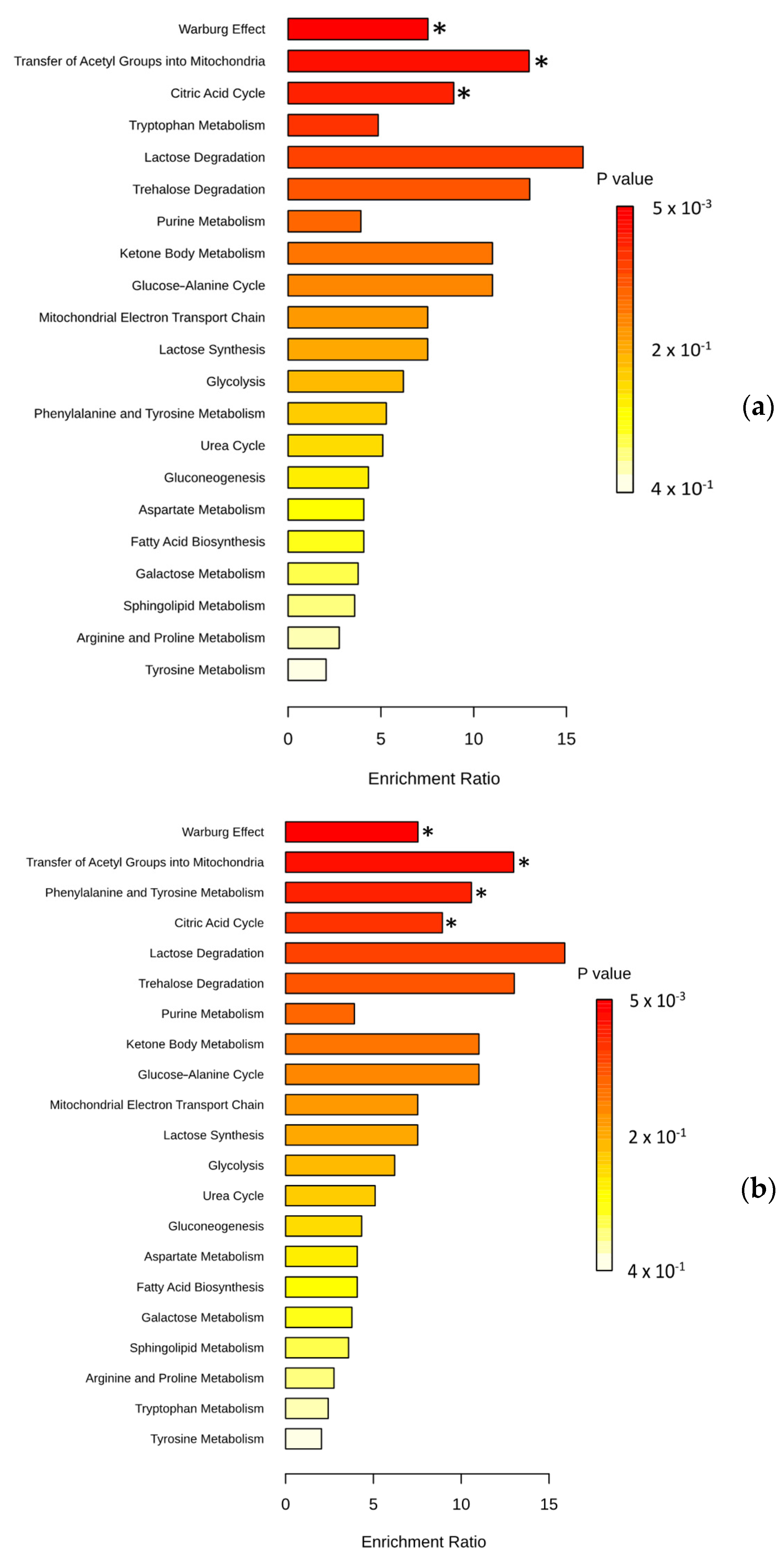
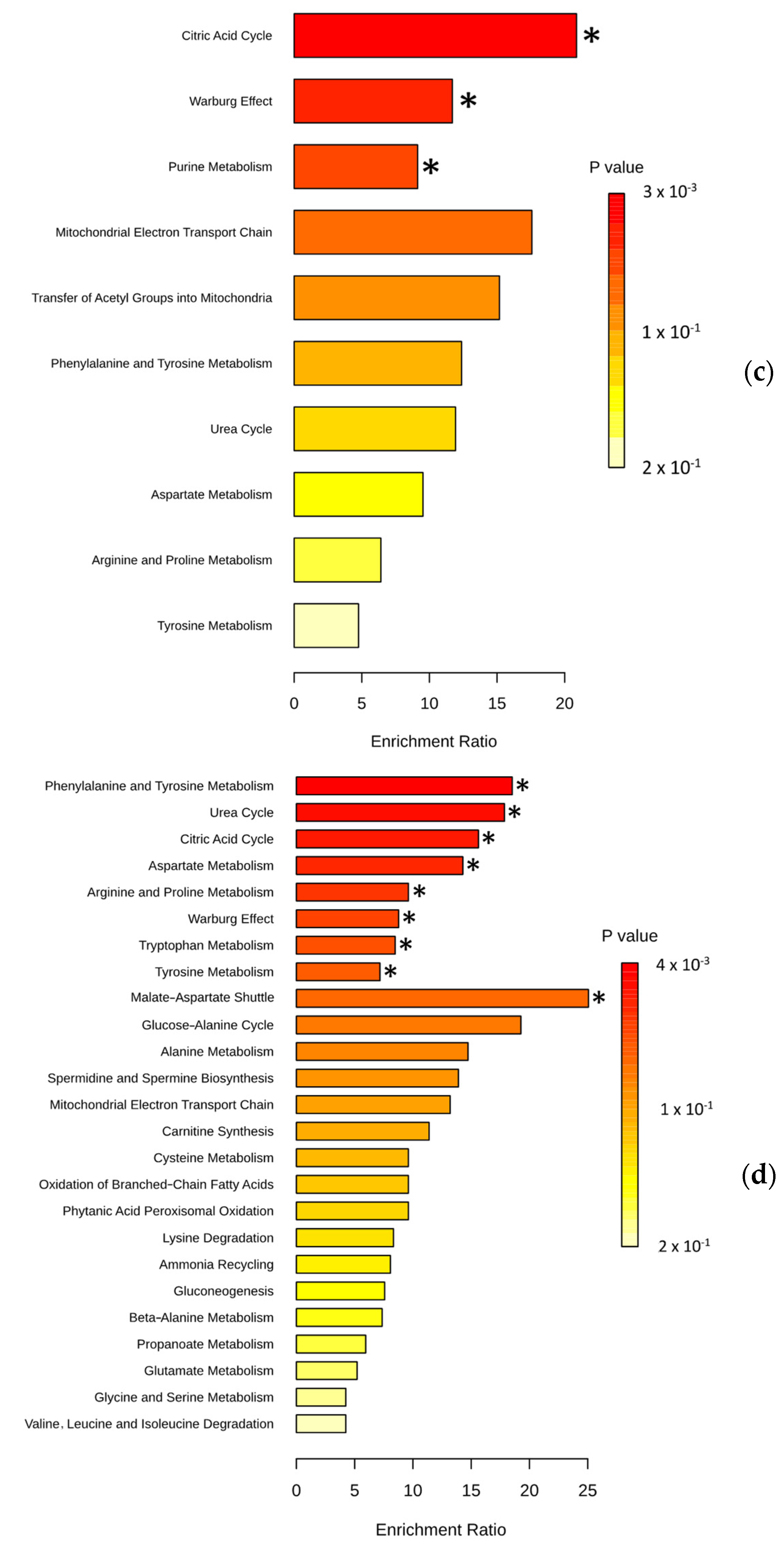
3.3. Pathway Analysis
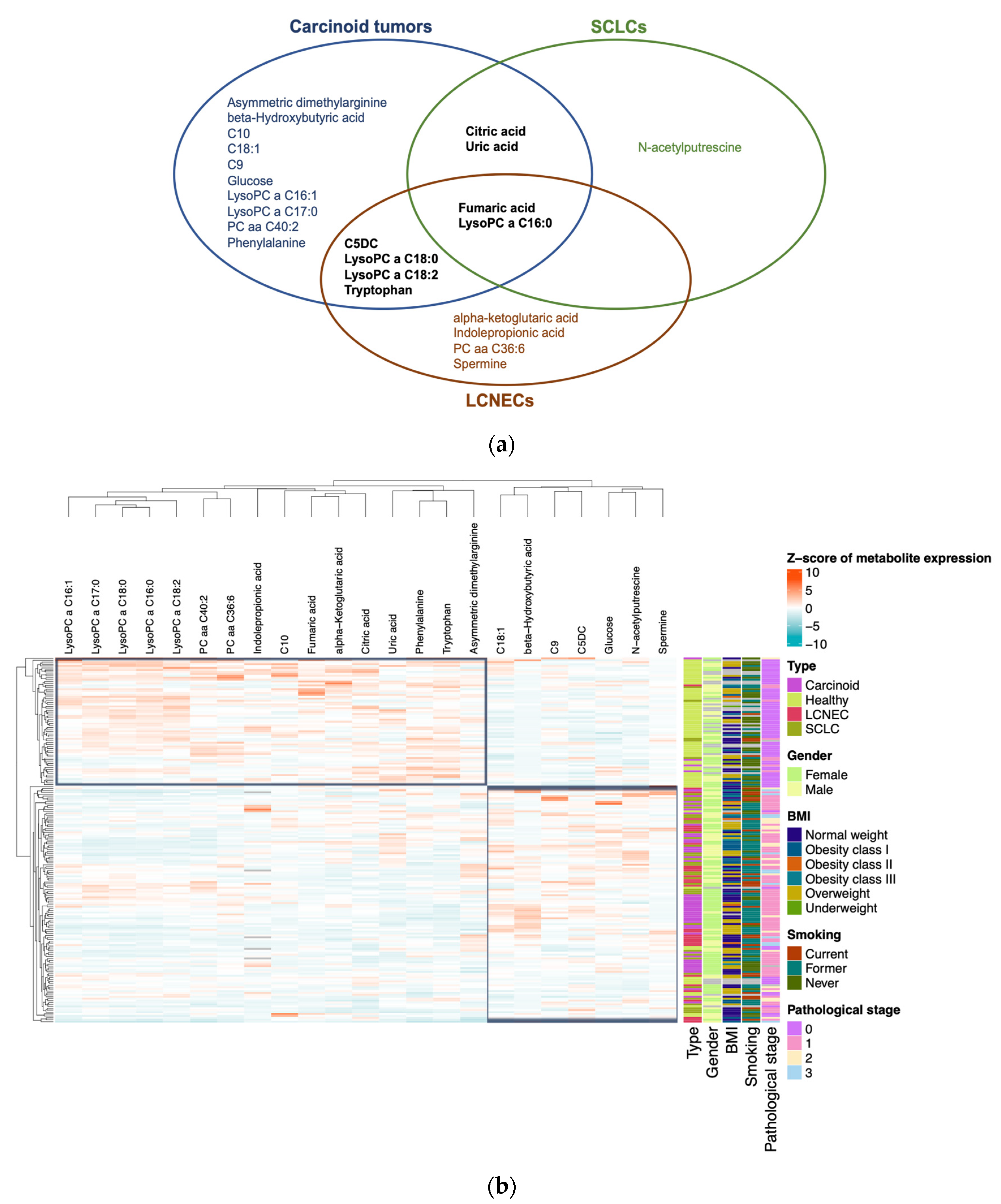
3.4. Plasma Metabolite Profile Can Predict Cancer Subtypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Classification of Tumours Editorial Board. Thoracic Tumours; WHO Classification of Tumours Editorial Board: Lyon, France, 2021; ISBN 978-92-832-4506-3.
- Travis, W.D.; Brambilla, E.; Nicholson, A.G.; Yatabe, Y.; Austin, J.H.M.; Beasley, M.B.; Chirieac, L.R.; Dacic, S.; Duhig, E.; Flieder, D.B.; et al. The 2015 World Health Organization Classification of Lung Tumors: Impact of Genetic, Clinical and Radiologic Advances Since the 2004 Classification. J. Thorac. Oncol. 2015, 10, 1243–1260. [Google Scholar] [CrossRef] [PubMed]
- Rekhtman, N. Lung Neuroendocrine Neoplasms: Recent Progress and Persistent Challenges. Mod. Pathol. 2022, 35, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Klöppel, G. Neuroendocrine Neoplasms: Dichotomy, Origin and Classifications. Visc. Med. 2017, 33, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Metovic, J.; Barella, M.; Bianchi, F.; Hofman, P.; Hofman, V.; Remmelink, M.; Kern, I.; Carvalho, L.; Pattini, L.; Sonzogni, A.; et al. Morphologic and Molecular Classification of Lung Neuroendocrine Neoplasms. Virchows Arch. 2021, 478, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Man, D.; Wu, J.; Shen, Z.; Zhu, X. Prognosis of Patients with Neuroendocrine Tumor: A SEER Database Analysis. Cancer Manag. Res. 2018, 10, 5629–5638. [Google Scholar] [CrossRef] [PubMed]
- Righi, L.; Volante, M.; Papotti, M. Small-Cell Carcinoma of the Lung: What We Learned about It? Acta Cytol. 2022, 66, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, F.G.; Battafarano, R.J. Large-Cell Neuroendocrine Carcinoma of the Lung. Cancer Control 2006, 13, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Pyun, W.Y.; Park, H.W. Cancer Metabolism: Phenotype, Signaling and Therapeutic Targets. Cells 2020, 9, 2308. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Shu, X.; Xiang, Y.-B.; Rothman, N.; Yu, D.; Li, H.-L.; Yang, G.; Cai, H.; Ma, X.; Lan, Q.; Gao, Y.-T.; et al. Prospective Study of Blood Metabolites Associated with Colorectal Cancer Risk. Int. J. Cancer 2018, 143, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Pakiet, A.; Kobiela, J.; Stepnowski, P.; Sledzinski, T.; Mika, A. Changes in Lipids Composition and Metabolism in Colorectal Cancer: A Review. Lipids Health Dis. 2019, 18, 29. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-P.; Lei, Q.-Y. Perspectives of Reprogramming Breast Cancer Metabolism. Adv. Exp. Med. Biol. 2017, 1026, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zheng, J.; Ahmed, R.; Huang, G.; Reid, J.; Mandal, R.; Maksymuik, A.; Sitar, D.S.; Tappia, P.S.; Ramjiawan, B.; et al. A High-Performing Plasma Metabolite Panel for Early-Stage Lung Cancer Detection. Cancers 2020, 12, 622. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. MSEA: A Web-Based Tool to Identify Biologically Meaningful Patterns in Quantitative Metabolomic Data. Nucleic Acids Res. 2010, 38, W71–W77. [Google Scholar] [CrossRef]
- Bian, X.; Liu, R.; Meng, Y.; Xing, D.; Xu, D.; Lu, Z. Lipid Metabolism and Cancer. J. Exp. Med. 2020, 218, e20201606. [Google Scholar] [CrossRef]
- Currie, E.; Schulze, A.; Zechner, R.; Walther, T.C.; Farese, R.V. Cellular Fatty Acid Metabolism and Cancer. Cell Metab. 2013, 18, 153–161. [Google Scholar] [CrossRef]
- Kim, K.S.; Moon, S.W.; Moon, M.H.; Hyun, K.Y.; Kim, S.J.; Kim, Y.K.; Kim, K.Y.; Jekarl, D.W.; Oh, E.-J.; Kim, Y. Metabolic Profiles of Lung Adenocarcinoma via Peripheral Blood and Diagnostic Model Construction. Sci. Rep. 2023, 13, 7304. [Google Scholar] [CrossRef]
- Bose, S.; Le, A. Glucose Metabolism in Cancer. Adv. Exp. Med. Biol. 2018, 1063, 3–12. [Google Scholar] [CrossRef]
- Eniafe, J.; Jiang, S. The Functional Roles of TCA Cycle Metabolites in Cancer. Oncogene 2021, 40, 3351–3363. [Google Scholar] [CrossRef] [PubMed]
- Crotti, S.; D’Angelo, E.; Bedin, C.; Fassan, M.; Pucciarelli, S.; Nitti, D.; Bertazzo, A.; Agostini, M. Tryptophan Metabolism along the Kynurenine and Serotonin Pathways Reveals Substantial Differences in Colon and Rectal Cancer. Metabolomics 2017, 13, 148. [Google Scholar] [CrossRef]
- Sarlinova, M.; Baranovicova, E.; Skalicanova, M.; Dzian, A.; Petras, M.; Lehotsky, J.; Kalenska, D.; Racay, P.; Matakova, T.; Halasova, E. Metabolomic Profiling of Blood Plasma of Patients with Lung Cancer and Malignant Tumors with Metastasis in the Lungs Showed Similar Features and Promising Statistical Discrimination against Controls. Neoplasma 2021, 68, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Shen, X.; Lei, J.; Xu, Q.; Yu, Y.; Li, R.; Wu, E.; Ma, Q. Hyperglycemia, a Neglected Factor during Cancer Progression. BioMed Res. Int. 2014, 2014, e461917. [Google Scholar] [CrossRef] [PubMed]
- Wieczór, A.M.; Wieczór, R.; Kulwas, A.; Rość, D. Asymmetric Dimethylarginine and Angiogenesis: Biological Significance. Int. Angiol. 2018, 37, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Wolin, E.M. Challenges in the Diagnosis and Management of Well-Differentiated Neuroendocrine Tumors of the Lung (Typical and Atypical Carcinoid): Current Status and Future Considerations. Oncologist 2015, 20, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hong, S.; Yang, J.; Zhang, X.; Wang, Y.; Wang, H.; Peng, J.; Hong, L. Targeting Purine Metabolism in Ovarian Cancer. J. Ovarian Res. 2022, 15, 93. [Google Scholar] [CrossRef]
- Losman, J.-A.; Koivunen, P.; Kaelin, W.G. 2-Oxoglutarate-Dependent Dioxygenases in Cancer. Nat. Rev. Cancer 2020, 20, 710–726. [Google Scholar] [CrossRef]
- Sári, Z.; Mikó, E.; Kovács, T.; Jankó, L.; Csonka, T.; Lente, G.; Sebő, É.; Tóth, J.; Tóth, D.; Árkosy, P.; et al. Indolepropionic Acid, a Metabolite of the Microbiome, Has Cytostatic Properties in Breast Cancer by Activating AHR and PXR Receptors and Inducing Oxidative Stress. Cancers 2020, 12, 2411. [Google Scholar] [CrossRef]
- Holbert, C.E.; Cullen, M.T.; Casero, R.A.; Stewart, T.M. Polyamines in Cancer: Integrating Organismal Metabolism and Antitumour Immunity. Nat. Rev. Cancer 2022, 22, 467–480. [Google Scholar] [CrossRef]
- Jones, M.H.; Virtanen, C.; Honjoh, D.; Miyoshi, T.; Satoh, Y.; Okumura, S.; Nakagawa, K.; Nomura, H.; Ishikawa, Y. Two Prognostically Significant Subtypes of High-Grade Lung Neuroendocrine Tumours Independent of Small-Cell and Large-Cell Neuroendocrine Carcinomas Identified by Gene Expression Profiles. Lancet 2004, 363, 775–781. [Google Scholar] [CrossRef]
- Onuki, N.; Wistuba, I.I.; Travis, W.D.; Virmani, A.K.; Yashima, K.; Brambilla, E.; Hasleton, P.; Gazdar, A.F. Genetic Changes in the Spectrum of Neuroendocrine Lung Tumors. Cancer 1999, 85, 600–607. [Google Scholar] [CrossRef]
- Yang, L.; Fan, Y.; Lu, H. Pulmonary Large Cell Neuroendocrine Carcinoma. Pathol. Oncol. Res. 2022, 28, 1610730. [Google Scholar] [CrossRef] [PubMed]
- Andrini, E.; Marchese, P.V.; De Biase, D.; Mosconi, C.; Siepe, G.; Panzuto, F.; Ardizzoni, A.; Campana, D.; Lamberti, G. Large Cell Neuroendocrine Carcinoma of the Lung: Current Understanding and Challenges. J. Clin. Med. 2022, 11, 1461. [Google Scholar] [CrossRef] [PubMed]
- Kuang, A.; Kouznetsova, V.L.; Kesari, S.; Tsigelny, I.F. Diagnostics of Thyroid Cancer Using Machine Learning and Metabolomics. Metabolites 2023, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Johansen, S.U.; Hansen, T.; Nordborg, A.; Meyer, R.; Goll, R.; Florholmen, J.; Jensen, E. Plasma Tryptophan Pathway Metabolites Quantified by Liquid Chromatography-Tandem Mass Spectrometry as Biomarkers in Neuroendocrine Tumor Patients. J. Neuroendocrinol. 2024, 36, e13372. [Google Scholar] [CrossRef] [PubMed]
- Eisenhofer, G.; Goldstein, D.S.; Walther, M.M.; Friberg, P.; Lenders, J.W.M.; Keiser, H.R.; Pacak, K. Biochemical Diagnosis of Pheochromocytoma: How to Distinguish True- from False-Positive Test Results. J. Clin. Endocrinol. Metab. 2003, 88, 2656–2666. [Google Scholar] [CrossRef]
- Casadonte, R.; Kriegsmann, M.; Perren, A.; Baretton, G.; Deininger, S.-O.; Kriegsmann, K.; Welsch, T.; Pilarsky, C.; Kriegsmann, J. Development of a Class Prediction Model to Discriminate Pancreatic Ductal Adenocarcinoma from Pancreatic Neuroendocrine Tumor by MALDI Mass Spectrometry Imaging. Proteom. Clin. Appl. 2019, 13, e1800046. [Google Scholar] [CrossRef]
- Stine, Z.E.; Schug, Z.T.; Salvino, J.M.; Dang, C.V. Targeting Cancer Metabolism in the Era of Precision Oncology. Nat. Rev. Drug Discov. 2022, 21, 141–162. [Google Scholar] [CrossRef]
- Seijo, L.M.; Peled, N.; Ajona, D.; Boeri, M.; Field, J.K.; Sozzi, G.; Pio, R.; Zulueta, J.J.; Spira, A.; Massion, P.P.; et al. Biomarkers in Lung Cancer Screening: Achievements, Promises, and Challenges. J. Thorac. Oncol. 2019, 14, 343–357. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Cases (n = 120) | Controls (n = 71) | p-Value |
|---|---|---|---|
| Age (years), mean ± SD | 61.9 ± 9.8 | 56.7 ± 10.7 | 0.16 |
| Carcinoids (n = 50) | 59.3 ± 11.2 | 0.89 | |
| SCLC (n = 40) | 63.3 ± 8.1 | 4.73 × 10−2 | |
| LCNEC (n = 30) | 64.1 ± 8.9 | 0.03 | |
| Sex (%) | 0.29 | ||
| Male | 36.7 | 45.1 | |
| Female | 63.3 | 54.9 | |
| Smoking status (%) | 1.09 × 10−7 | ||
| Current smokers | 23.3 | 8.4 | |
| Ex-smokers | 58.3 | 35.2 | |
| Non-smokers | 18.3 | 56.3 | |
| BMI (kg/m2), mean ± SD | 27.8 ± 4.7 | 26.7 ± 6.0 | 0.19 |
| Characteristics | Cases (n = 120) | NSCLC (n = 466) | p-Value |
|---|---|---|---|
| Age (years), mean ± SD | 61.9 ± 9.8 | 65.2 ± 8.1 | 6.50 × 10−4 |
| Carcinoids (n = 50) | 59.3 ± 11.2 | ||
| SCLC (n = 40) | 63.3 ± 8.1 | ||
| LCNEC (n = 30) | 64.1 ± 8.9 | ||
| Sex (%) | 7.70 × 10−2 | ||
| Male | 36.7 | 50.1 | |
| Female | 63.3 | 49.9 | |
| Smoking status (%) | 1.80 × 10−2 | ||
| Current smokers | 23.3 | 22.8 | |
| Ex-smokers | 58.3 | 72.2 | |
| Non-smokers | 18.3 | 4.6 | |
| Passive | 0 | 0.4 | |
| BMI (kg/m2), mean ± SD | 27.8 ± 4.7 | 27.1 ± 5.1 | 0.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boullier, C.; Lamaze, F.C.; Haince, J.-F.; Bux, R.A.; Orain, M.; Zheng, J.; Zhang, L.; Wishart, D.S.; Bossé, Y.; Manem, V.S.K.; et al. Metabolomic Profiling of Pulmonary Neuroendocrine Neoplasms. Cancers 2024, 16, 3179. https://doi.org/10.3390/cancers16183179
Boullier C, Lamaze FC, Haince J-F, Bux RA, Orain M, Zheng J, Zhang L, Wishart DS, Bossé Y, Manem VSK, et al. Metabolomic Profiling of Pulmonary Neuroendocrine Neoplasms. Cancers. 2024; 16(18):3179. https://doi.org/10.3390/cancers16183179
Chicago/Turabian StyleBoullier, Clémence, Fabien C. Lamaze, Jean-François Haince, Rashid Ahmed Bux, Michèle Orain, Jiamin Zheng, Lun Zhang, David S. Wishart, Yohan Bossé, Venkata S. K. Manem, and et al. 2024. "Metabolomic Profiling of Pulmonary Neuroendocrine Neoplasms" Cancers 16, no. 18: 3179. https://doi.org/10.3390/cancers16183179
APA StyleBoullier, C., Lamaze, F. C., Haince, J.-F., Bux, R. A., Orain, M., Zheng, J., Zhang, L., Wishart, D. S., Bossé, Y., Manem, V. S. K., & Joubert, P. (2024). Metabolomic Profiling of Pulmonary Neuroendocrine Neoplasms. Cancers, 16(18), 3179. https://doi.org/10.3390/cancers16183179