
Epigenetic Rewiring of Metastatic Cancer to the Brain: Focus on Lung and Colon Cancers
 , , , ,
, , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. DNA Purification and Bisulfite Conversion
2.3. Infinium MethylationEPIC BeadChip Array
2.4. Data Analysis
3. Results
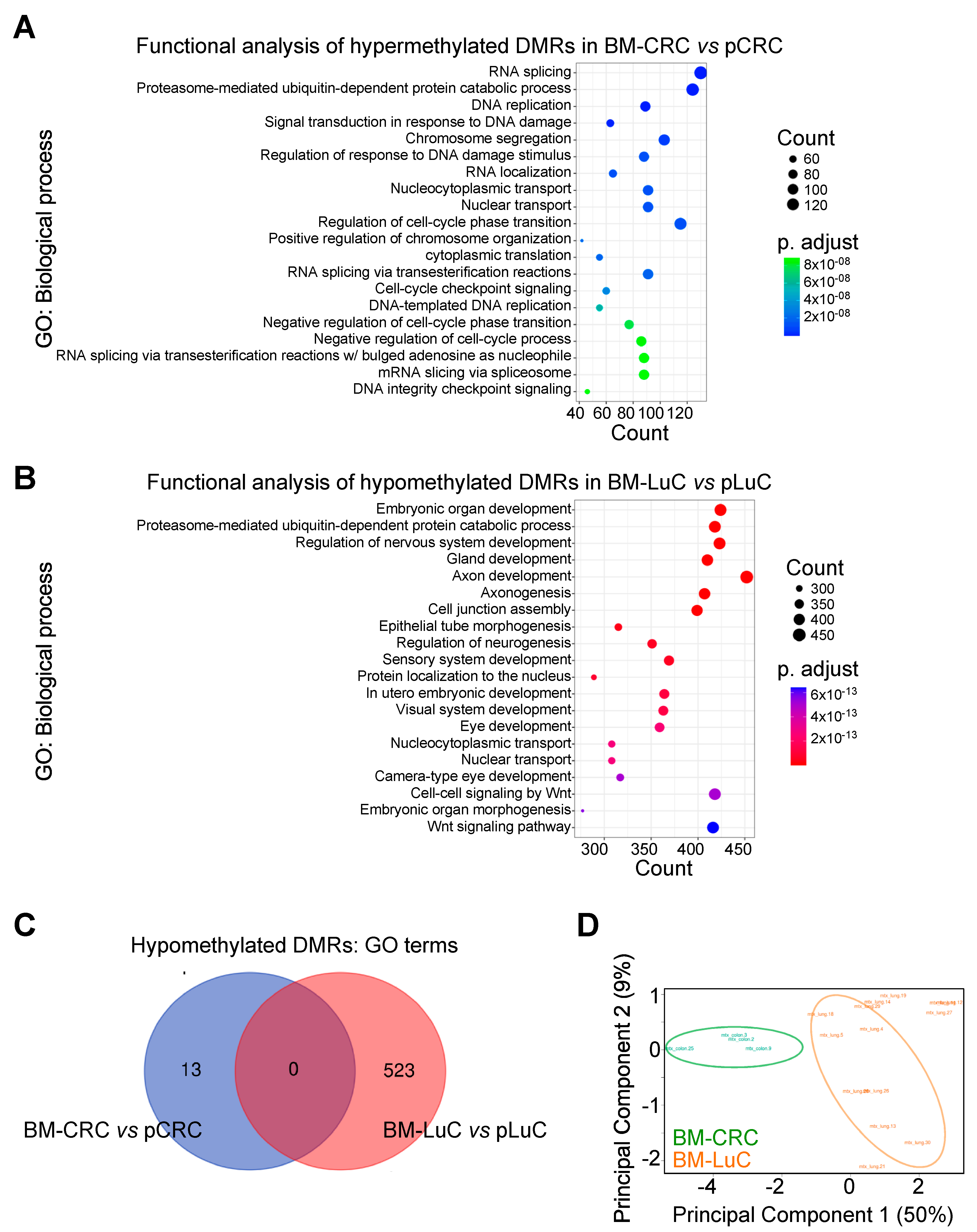
3.1. CpG Methylation Pattern in Brain Metastasis from CRC or Lung Cancers
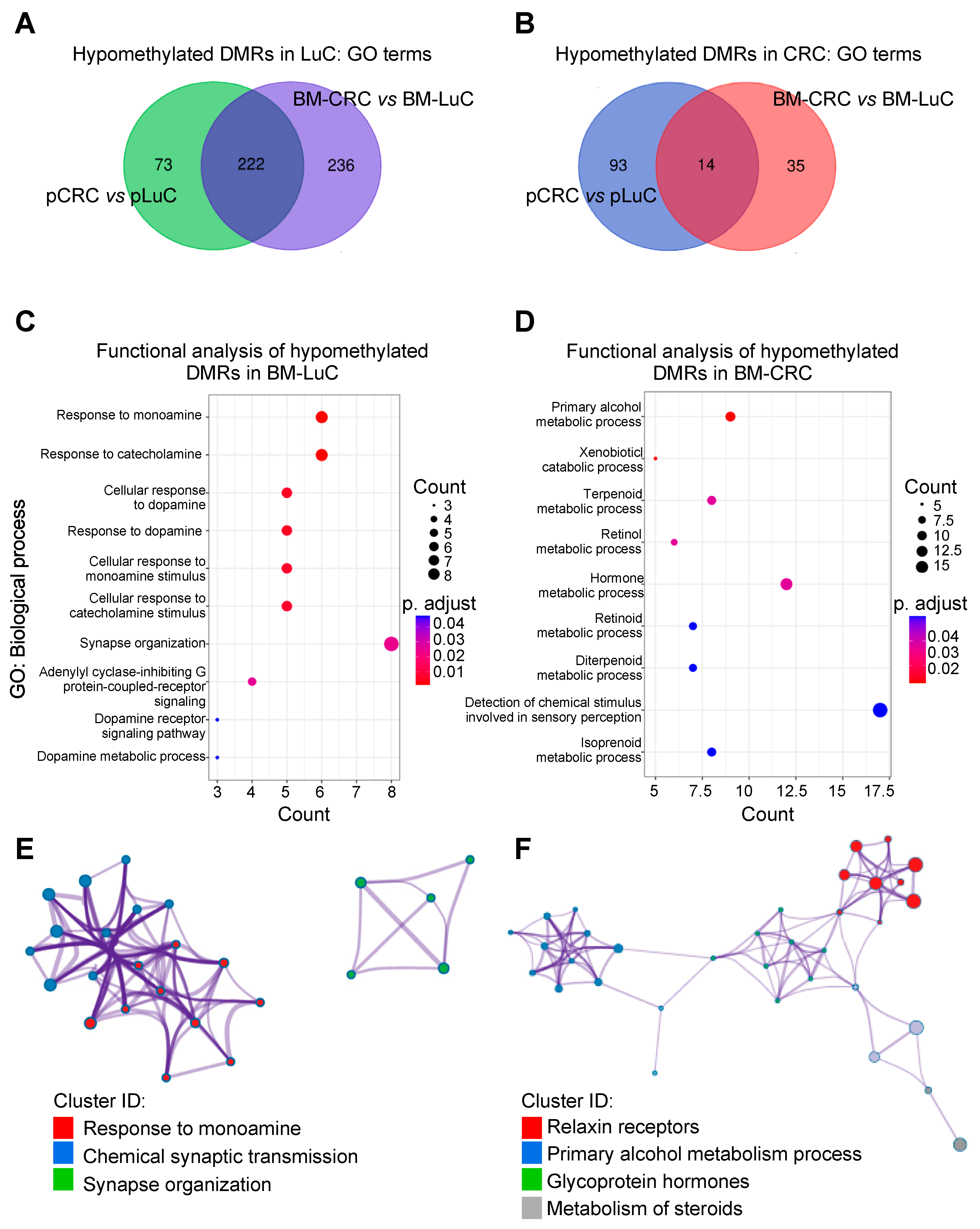
3.2. Differentially Methylated Regions Specific for CRC and LuC brain Metastasis
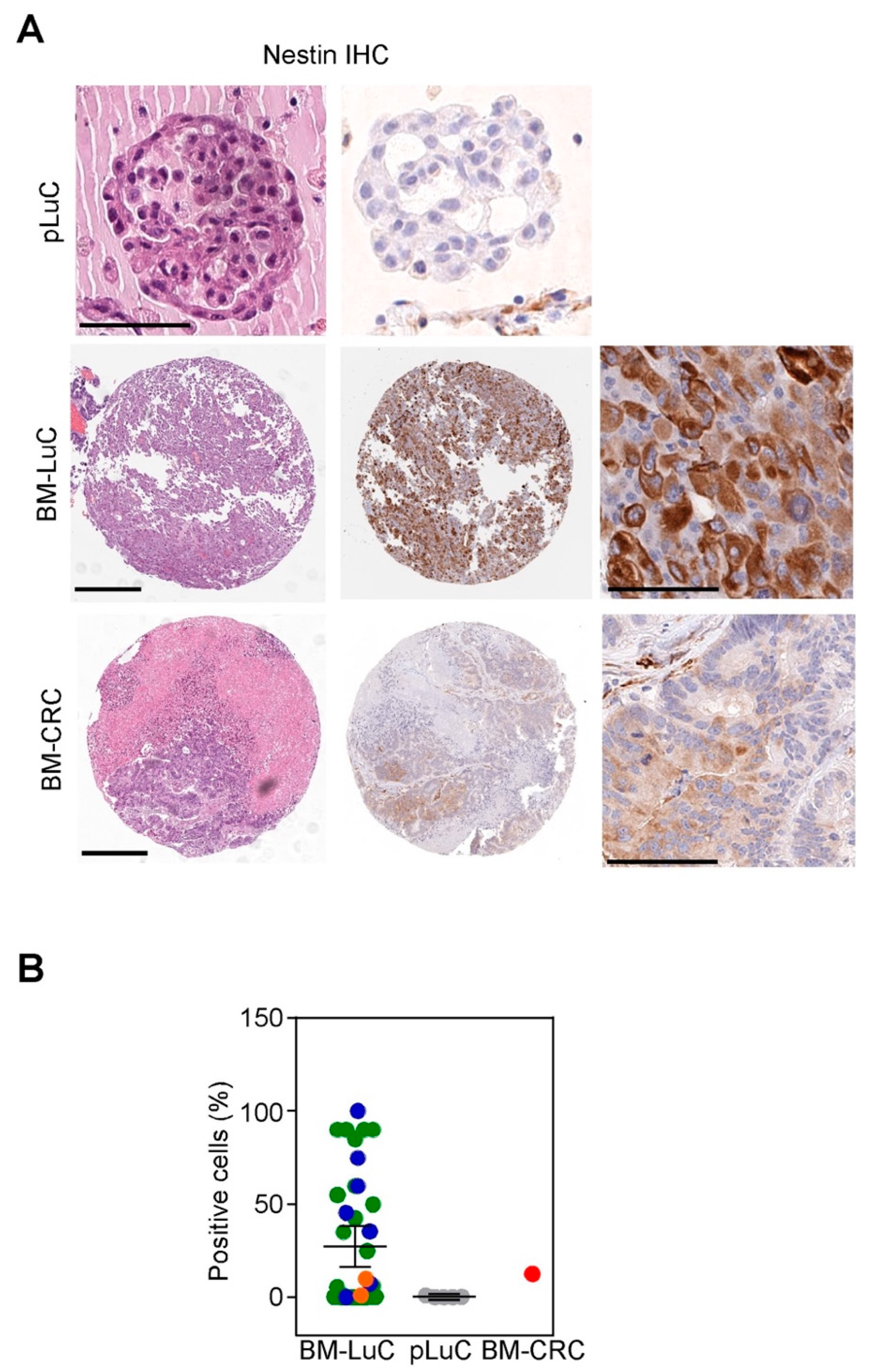
3.3. Brain Metastases from Lung Cancer Acquire Neuronal Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sacks, P.; Rahman, M. Epidemiology of Brain Metastases. Neurosurg. Clin. N. Am. 2020, 31, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Achrol, A.S.; Rennert, R.C.; Anders, C.; Soffietti, R.; Ahluwalia, M.S.; Nayak, L.; Peters, S.; Arvold, N.D.; Harsh, G.R.; Steeg, P.S.; et al. Brain metastases. Nat. Rev. Dis. Primers 2019, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Cagney, D.N.; Martin, A.M.; Catalano, P.J.; Redig, A.J.; Lin, N.U.; Lee, E.Q.; Wen, P.Y.; Dunn, I.F.; Bi, W.L.; Weiss, S.E.; et al. Incidence and prognosis of patients with brain metastases at diagnosis of systemic malignancy: A population-based study. Neuro-Oncology 2017, 19, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Caroli, M.; Di Cristofori, A.; Lucarella, F.; Raneri, F.A.; Portaluri, F.; Gaini, S.M. Surgical brain metastases: Management and outcome related to prognostic indexes: A critical review of a ten-year series. ISRN Surg. 2011, 2011, 207103. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.H.; Kotecha, R.; Chao, S.T.; Ahluwalia, M.S.; Sahgal, A.; Chang, E.L. Current approaches to the management of brain metastases. Nat. Rev. Clin. Oncol. 2020, 17, 279–299. [Google Scholar] [CrossRef]
- Soffietti, R.; Trevisan, E.; Rudà, R. Targeted therapy in brain metastasis. Curr. Opin. Oncol. 2012, 24, 679–686. [Google Scholar] [CrossRef]
- Zoni, E.; van der Pluijm, G.; Gray, P.C.; Kruithof-de Julio, M. Epithelial Plasticity in Cancer: Unmasking a MicroRNA Network for TGF-β-, Notch-, and Wnt-Mediated EMT. J. Oncol. 2015, 2015, 198967. [Google Scholar] [CrossRef]
- Kim, B.N.; Ahn, D.H.; Kang, N.; Yeo, C.D.; Kim, Y.K.; Lee, K.Y.; Kim, T.J.; Lee, S.H.; Park, M.S.; Yim, H.W.; et al. TGF-β induced EMT and stemness characteristics are associated with epigenetic regulation in lung cancer. Sci. Rep. 2020, 10, 10597. [Google Scholar] [CrossRef]
- Su, H.T.; Weng, C.C.; Hsiao, P.J.; Chen, L.H.; Kuo, T.L.; Chen, Y.W.; Kuo, K.K.; Cheng, K.H. Stem cell marker nestin is critical for TGF-β1-mediated tumor progression in pancreatic cancer. Mol. Cancer Res. 2013, 11, 768–779. [Google Scholar] [CrossRef]
- Narita, K.; Matsuda, Y.; Seike, M.; Naito, Z.; Gemma, A.; Ishiwata, T. Nestin regulates proliferation, migration, invasion and stemness of lung adenocarcinoma. Int. J. Oncol. 2014, 44, 1118–1130. [Google Scholar] [CrossRef]
- Termini, J.; Neman, J.; Jandial, R. Role of the neural niche in brain metastatic cancer. Cancer Res. 2014, 74, 4011–4015. [Google Scholar] [CrossRef] [PubMed]
- Ramaswamy, S.; Tamayo, P.; Rifkin, R.; Mukherjee, S.; Yeang, C.H.; Angelo, M.; Ladd, C.; Reich, M.; Latulippe, E.; Mesirov, J.P.; et al. Multiclass cancer diagnosis using tumor gene expression signatures. Proc. Natl. Acad. Sci. USA 2001, 98, 15149–15154. [Google Scholar] [CrossRef] [PubMed]
- Fidler, I.J.; Kripke, M.L. Metastasis results from preexisting variant cells within a malignant tumor. Science 1977, 197, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Yuzhalin, A.E.; Yu, D. Brain Metastasis Organotropism. Cold Spring Harb. Perspect. Med. 2020, 10, a037242. [Google Scholar] [CrossRef]
- Hirata, E.; Ishibashi, K.; Kohsaka, S.; Shinjo, K.; Kojima, S.; Kondo, Y.; Mano, H.; Yano, S.; Kiyokawa, E.; Sahai, E. The Brain Microenvironment Induces DNMT1 Suppression and Indolence of Metastatic Cancer Cells. iScience 2020, 23, 101480. [Google Scholar] [CrossRef]
- Ramos, E.A.; Grochoski, M.; Braun-Prado, K.; Seniski, G.G.; Cavalli, I.J.; Ribeiro, E.M.; Camargo, A.A.; Costa, F.F.; Klassen, G. Epigenetic changes of CXCR4 and its ligand CXCL12 as prognostic factors for sporadic breast cancer. PLoS ONE 2011, 6, e29461. [Google Scholar] [CrossRef]
- Bucheit, A.D.; Chen, G.; Siroy, A.; Tetzlaff, M.; Broaddus, R.; Milton, D.; Fox, P.; Bassett, R.; Hwu, P.; Gershenwald, J.E.; et al. Complete loss of PTEN protein expression correlates with shorter time to brain metastasis and survival in stage IIIB/C melanoma patients with BRAFV600 mutations. Clin. Cancer Res. 2014, 20, 5527–5536. [Google Scholar] [CrossRef]
- Miranda Furtado, C.L.; Dos Santos Luciano, M.C.; Silva Santos, R.D.; Furtado, G.P.; Moraes, M.O.; Pessoa, C. Epidrugs: Targeting epigenetic marks in cancer treatment. Epigenetics 2019, 14, 1164–1176. [Google Scholar] [CrossRef]
- Lu, Z.; Zou, J.; Li, S.; Topper, M.J.; Tao, Y.; Zhang, H.; Jiao, X.; Xie, W.; Kong, X.; Vaz, M.; et al. Epigenetic therapy inhibits metastases by disrupting premetastatic niches. Nature 2020, 579, 284–290. [Google Scholar] [CrossRef]
- Chianese, A.; Santella, B.; Ambrosino, A.; Stelitano, D.; Rinaldi, L.; Galdiero, M.; Zannella, C.; Franci, G. Oncolytic Viruses in Combination Therapeutic Approaches with Epigenetic Modulators: Past, Present, and Future Perspectives. Cancers 2021, 13, 2761. [Google Scholar] [CrossRef]
- Bartlett, D.L.; Liu, Z.; Sathaiah, M.; Ravindranathan, R.; Guo, Z.; He, Y.; Guo, Z.S. Oncolytic viruses as therapeutic cancer vaccines. Mol. Cancer 2013, 12, 103. [Google Scholar] [CrossRef] [PubMed]
- MacTavish, H.; Diallo, J.S.; Huang, B.; Stanford, M.; Le Boeuf, F.; De Silva, N.; Cox, J.; Simmons, J.G.; Guimond, T.; Falls, T.; et al. Enhancement of vaccinia virus based oncolysis with histone deacetylase inhibitors. PLoS ONE 2010, 5, e14462. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, C.; Marchini, A. H-1 Parvovirus as a Cancer-Killing Agent: Past, Present, and Future. Viruses 2019, 11, 562. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Rivadeneira, D.B.; Caino, M.C.; Chae, Y.C.; Speicher, D.W.; Tang, H.Y.; Vaira, V.; Bosari, S.; Palleschi, A.; Rampini, P.; et al. The Mitochondrial Unfoldase-Peptidase Complex ClpXP Controls Bioenergetics Stress and Metastasis. PLoS Biol. 2016, 14, e1002507. [Google Scholar] [CrossRef] [PubMed]
- Maksimovic, J.; Phipson, B.; Oshlack, A. A cross-package Bioconductor workflow for analysing methylation array data. F1000Res 2016, 5, 1281. [Google Scholar] [CrossRef] [PubMed]
- Terrasi, A.; Bertolini, I.; Martelli, C.; Gaudioso, G.; Di Cristofori, A.; Storaci, A.M.; Formica, M.; Bosari, S.; Caroli, M.; Ottobrini, L.; et al. Specific V-ATPase expression sub-classifies IDHwt lower-grade gliomas and impacts glioma growth in vivo. EBioMedicine 2019, 41, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://bioinformatics.psb.ugent.be/cgi-bin/liste/Venn/calculate_venn.htpl (accessed on 7 March 2023).
- McInnes, T.; Zou, D.; Rao, D.S.; Munro, F.M.; Phillips, V.L.; McCall, J.L.; Black, M.A.; Reeve, A.E.; Guilford, P.J. Genome-wide methylation analysis identifies a core set of hypermethylated genes in CIMP-H colorectal cancer. BMC Cancer 2017, 17, 228. [Google Scholar] [CrossRef]
- Heide, T.; Househam, J.; Cresswell, G.D.; Spiteri, I.; Lynn, C.; Mossner, M.; Kimberley, C.; Fernandez-Mateos, J.; Chen, B.; Zapata, L.; et al. The co-evolution of the genome and epigenome in colorectal cancer. Nature 2022, 611, 733–743. [Google Scholar] [CrossRef]
- Parmar, S.; Easwaran, H. Genetic and epigenetic dependencies in colorectal cancer development. Gastroenterol. Rep. 2022, 10, goac035. [Google Scholar] [CrossRef]
- Orozco, J.I.J.; Knijnenburg, T.A.; Manughian-Peter, A.O.; Salomon, M.P.; Barkhoudarian, G.; Jalas, J.R.; Wilmott, J.S.; Hothi, P.; Wang, X.; Takasumi, Y.; et al. Epigenetic profiling for the molecular classification of metastatic brain tumors. Nat. Commun. 2018, 9, 4627. [Google Scholar] [CrossRef]
- Salomon, M.P.; Orozco, J.I.J.; Wilmott, J.S.; Hothi, P.; Manughian-Peter, A.O.; Cobbs, C.S.; Scolyer, R.A.; Hoon, D.S.B.; Marzese, D.M. Brain metastasis DNA methylomes, a novel resource for the identification of biological and clinical features. Sci. Data 2018, 5, 180245. [Google Scholar] [CrossRef] [PubMed]
- Karlow, J.A.; Devarakonda, S.; Xing, X.; Jang, H.S.; Govindan, R.; Watson, M.; Wang, T. Developmental Pathways Are Epigenetically Reprogrammed during Lung Cancer Brain Metastasis. Cancer Res. 2022, 82, 2692–2703. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, C.; Zhang, Y.; Xu, L.; Fang, W.; Zhu, Y.; Zheng, Y.; Chen, X.; Xie, X.; Hu, X.; et al. Genomic signatures reveal DNA damage response deficiency in colorectal cancer brain metastases. Nat. Commun. 2019, 10, 3190. [Google Scholar] [CrossRef] [PubMed]
- Poturnajova, M.; Furielova, T.; Balintova, S.; Schmidtova, S.; Kucerova, L.; Matuskova, M. Molecular features and gene expression signature of metastatic colorectal cancer (Review). Oncol. Rep. 2021, 45, 10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Mutation | Immunephenotype | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ID | Gender | Age at Dx | Histology | ALK-R | BRAF | EGFR | ERBB2 | KRAS | ROS1-R | CK7 | CKAE1/3 | NAPSIN-A | p40 | p63 | TTF1 | CDX2 | CK20 |
| Lung | |||||||||||||||||
| BM-LuC1 | M | 78 | NSCLC, NOS | WT | + | + | - | - | - | - | |||||||
| BM-LuC2 | M | 73 | ADCA | WT | + | - | + | ||||||||||
| BM-LuC3 | M | 60 | SCC | - | + | - | |||||||||||
| BM-LuC4 | M | 57 | ADCA | WT | WT | WT | + | + | - | + | - | ||||||
| BM-LuC5 | F | 70 | ADCA | WT | WT | Mut | + | + | + | - | |||||||
| BM-LuC6 | M | 55 | ADCA | WT | WT | WT | WT | + | - | + | - | - | |||||
| BM-LuC7 | M | 54 | ADCA | WT | WT | WT | + | + | + | - | |||||||
| BM-LuC8 | F | 65 | ADCA | + | + | - | + | - | |||||||||
| BM-LuC9 | M | 50 | NSCLC, NOS | WT | WT | ||||||||||||
| BM-LuC10 | M | 43 | NSCLC, NOS | WT | WT | WT | Mut | WT | WT | + | - | - | - | - | - | ||
| BM-LuC11 | F | 59 | SCC | + | |||||||||||||
| BM-LuC12 | M | 75 | SCC | - | + | ||||||||||||
| BM-LuC13 | F | 62 | ADCA | Mut | WT | + | - | - | - | - | |||||||
| BM-LuC14 | M | 69 | SCC | WT | WT | WT | - | + | + | - | - | ||||||
| Colon | |||||||||||||||||
| BM-CRC1 | M | 62 | ADCA | + | + | ||||||||||||
| BM-CRC2 | M | 65 | ADCA | - | + | + | |||||||||||
| BM-CRC3 | F | 71 | ADCA | WT | Mut | - | + | + | |||||||||
| BM-CRC4 | F | 80 | ADCA | + | + | ||||||||||||
| Primary | Histology | N |
|---|---|---|
| Metastases | ||
| Lung | ADCA | 32 |
| SCC | 2 | |
| NSCLC-NOS | 7 | |
| Colon | ADCA | 1 |
| Primary cancer | ||
| Lung | ADCA | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morotti, A.; Gentile, F.; Lopez, G.; Passignani, G.; Valenti, L.; Locatelli, M.; Caroli, M.; Fanizzi, C.; Ferrero, S.; Vaira, V. Epigenetic Rewiring of Metastatic Cancer to the Brain: Focus on Lung and Colon Cancers. Cancers 2023, 15, 2145. https://doi.org/10.3390/cancers15072145
Morotti A, Gentile F, Lopez G, Passignani G, Valenti L, Locatelli M, Caroli M, Fanizzi C, Ferrero S, Vaira V. Epigenetic Rewiring of Metastatic Cancer to the Brain: Focus on Lung and Colon Cancers. Cancers. 2023; 15(7):2145. https://doi.org/10.3390/cancers15072145
Chicago/Turabian StyleMorotti, Annamaria, Francesco Gentile, Gianluca Lopez, Giulia Passignani, Luca Valenti, Marco Locatelli, Manuela Caroli, Claudia Fanizzi, Stefano Ferrero, and Valentina Vaira. 2023. "Epigenetic Rewiring of Metastatic Cancer to the Brain: Focus on Lung and Colon Cancers" Cancers 15, no. 7: 2145. https://doi.org/10.3390/cancers15072145
APA StyleMorotti, A., Gentile, F., Lopez, G., Passignani, G., Valenti, L., Locatelli, M., Caroli, M., Fanizzi, C., Ferrero, S., & Vaira, V. (2023). Epigenetic Rewiring of Metastatic Cancer to the Brain: Focus on Lung and Colon Cancers. Cancers, 15(7), 2145. https://doi.org/10.3390/cancers15072145