
Expression and Activity of the NF-κB Subunits in Chronic Lymphocytic Leukaemia: A Role for RelB and Non-Canonical Signalling


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Sample Collection and Information
2.2. NF-κB DNA Binding Activity and Expression
2.3. Western Blotting
2.4. Cytotoxicity Assays in CLL Cells
2.5. Co-Culture with CD40L-Expressing Cells
3. Results
3.1. NF-κB Subunit Quantification in CLL Patient Samples
3.2. Unmutated IGHV and High-Risk Cytogenetic Abnormalities Are Associated with Increased NF-κB Activation
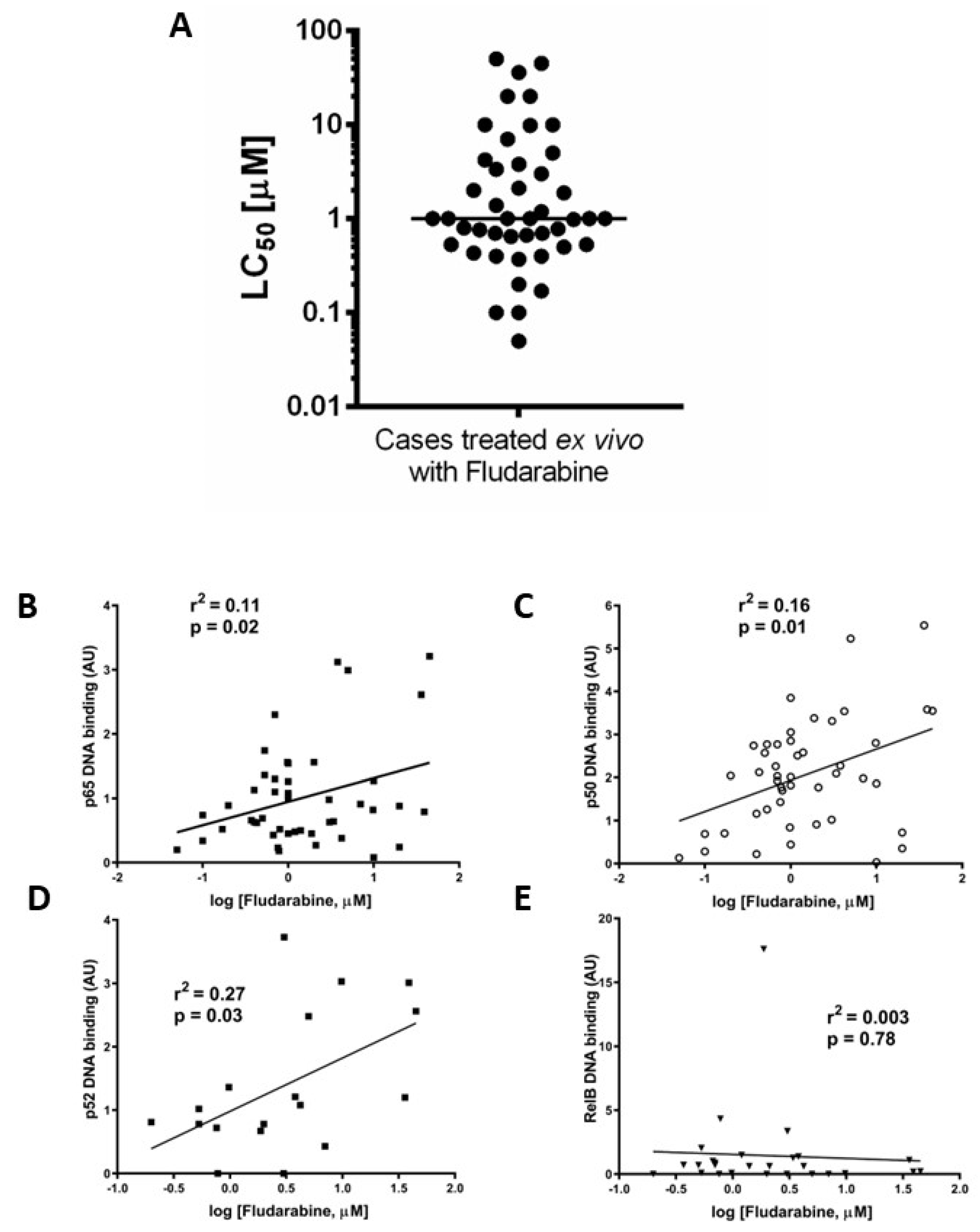
3.3. Ex Vivo Chemoresistance Is Significantly Associated with High p65 and p50 Levels but Not p52 or RelB
3.4. Increased Activity of RelB in ATM Dysfunctional Cases
3.5. RelB and p52 Expression Is Increased Following CD40L-Stimulated Proliferation in CLL
3.6. High Levels of RelB Activity Are Associated with Shorter Time to Treatment and Survival in CLL
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roeker, L.E.; Thompson, M.; Mato, A.R. Current Treatment of Chronic Lymphocytic Leukemia: The Diminishing Role of Chemoimmunotherapy. Drugs 2022, 82, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Fürstenau, M.; Eichhorst, B. Novel Agents in Chronic Lymphocytic Leukemia: New Combination Therapies and Strategies to Overcome Resistance. Cancers 2021, 13, 1336. [Google Scholar] [CrossRef] [PubMed]
- Zenz, T.; Eichhorst, B.; Busch, R.; Denzel, T.; Häbe, S.; Winkler, D.; Bühler, A.; Edelmann, J.; Bergmann, M.; Hopfinger, G.; et al. TP53 Mutation and survival in Chronic Lymphocytic Leukemia. J. Clin. Oncol. 2010, 29, 4473–4479. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, T.; Skowronska, A. The role of ATM mutations and 11q deletions in disease progression in chronic lymphocytic leukemia. Leuk. Lymphoma 2014, 55, 1227–1239. [Google Scholar] [CrossRef]
- Perkins, N.D. Achieving Transcriptional Specificity with NF-κB. Int. J. Biochem. Cell Biol. 1997, 29, 1433–1448. [Google Scholar] [CrossRef]
- Perkins, N.D. The diverse and complex roles of NF-κB subunits in cancer. Nat. Rev. Cancer 2012, 12, 121–132. [Google Scholar] [CrossRef]
- Veuger, S.J.; Durkacz, B.W. Persistence of unrepaired DNA double strand breaks caused by inhibition of ATM does not lead to radiosensitization in the absence of NF-κB activation. DNA Repair. 2010, 10, 235–244. [Google Scholar] [CrossRef]
- Gaudio, E.; Spizzo, R.; Paduano, F.; Luo, Z.; Efanov, A.; Palamarchuk, A.; Leber, A.S.; Kaou, M.; Zanesi, N.; Bottoni, A.; et al. Tcl1 interacts with Atm and enhances NF-κB activation in hematologic malignancies. Blood 2012, 119, 180–187. [Google Scholar] [CrossRef]
- O’Donnell, A.; Pepper, C.; Mitchell, S.; Pepper, A. NF-kB and the CLL microenvironment. Front. Oncol. 2023, 13, 1169397. [Google Scholar] [CrossRef]
- Gilmore, T.D. Multiple myeloma: Lusting for NF-kappaB. Cancer Cell 2007, 12, 95–97. [Google Scholar] [CrossRef]
- Rahal, R.; Frick, M.; Romero, R.; Korn, J.M.; Kridel, R.; Chan, F.C.; Meissner, B.; Bhang, H.E.; Ruddy, D.; Kauffmann, A.; et al. Pharmacological and genomic profiling identifies NF-κB-targeted treatment strategies for mantle cell lymphoma. Nat. Med. 2014, 20, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Diop, F.; Moia, R.; Favini, C.; Spaccarotella, E.; De Paoli, L.; Bruscaggin, A.; Spina, V.; Terzi-di-Bergamo, L.; Arruga, F.; Tarantelli, C.; et al. Biological and clinical implications of BIRC3 mutations in chronic lymphocytic leukemia. Haematologica 2019, 105, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Puente, X.S.; Pinyol, M.; Quesada, V.; Conde, L.; Ordóñez, G.R.; Villamor, N.; Escaramis, G.; Jares, P.; Beà, S.; González-Díaz, M.; et al. Whole-genome sequencing identifies recurrent mutations in chronic lymphocytic leukaemia. Nature 2011, 475, 101–105. [Google Scholar] [CrossRef]
- Puente, X.S.; Beà, S.; Valdés-Mas, R.; Villamor, N.; Gutiérrez-Abril, J.; Martín-Subero, J.; Munar, M.; Rubio-Pérez, C.; Jares, P.; Aymerich, M.; et al. Non-coding recurrent mutations in chronic lymphocytic leukaemia. Nature 2015, 526, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Furman, R.R.; Asgary, Z.; Mascarenhas, J.O.; Liou, H.C.; Schattner, E.J. Modulation of NF-κB activity and apoptosis in chronic lymphocytic leukaemia B cells. J. Immun. 2000, 164, 2200–2206. [Google Scholar] [CrossRef]
- Hewamana, S.; Alghazal, S.; Lin, T.T.; Clement, M.; Jenkins, C.; Guzman, M.L.; Jordan, C.T.; Neelakantan, S.; Crooks, P.A.; Burnett, A.K.; et al. The NF-κB subunit Rel A is associated with in vitro survival and clinical disease progression in chronic lymphocytic leukemia and represents a promising therapeutic target. Blood 2008, 111, 4681–4689. [Google Scholar] [CrossRef]
- Steele, A.J.; Jones, D.T.; Ganeshaguru, K.; Duke, V.M.; Yogashangary, B.C.; North, J.M.; Lowdell, M.W.; Kottaridis, P.D.; Mehta, A.B.; Prentice, A.G.; et al. The sesquiterpene lactone parthenolide induces selective apoptosis of B-chronic lymphocytic leukemia cells in vitro. Leukemia 2006, 20, 1073–1079. [Google Scholar] [CrossRef]
- Lewis, T.; Corcoran, D.B.; Thurston, D.E.; Giles, P.J.; Ashelford, K.; Walsby, E.J.; Fegan, C.D.; Pepper, A.G.S.; Miraz Rahman, K.; Pepper, C. Novel pyrrolobenzodiazepine benzofused hybrid molecules inhibit NF-κB activity and synergise with bortezomib and ibrutinib in hematological cancers. Haematologica 2021, 106, 958–967. [Google Scholar] [CrossRef]
- Willmore, E.; Elliott, S.L.; Mainou-Fowler, T.; Summerfield, G.P.; Jackson, G.H.; O’Neill, F.; Lowe, C.; Carter, A.; Harris, R.; Pettitt, A.R.; et al. DNA-dependent protein kinase is a therapeutic target and an indicator of poor prognosis in B-cell chronic lymphocytic leukemia. Clin. Cancer Res 2009, 14, 3984–3992. [Google Scholar] [CrossRef]
- Hunter, J.E.; Willmore, E.; Irving, J.A.E.; Hostomsky, Z.; Veuger, S.J.; Durkacz, B.W. NF-κB mediates radio-sensitization by the PARP-1 inhibitor, AG-014699. Oncogene 2012, 31, 251–264. [Google Scholar] [CrossRef]
- Bhattacharya, N.; Sarno, A.; Idler, I.S.; Führer, M.; Zenz, T.; Döhner, H.; Stilgenbauer, S.; Mertens, D. High-throughput detection of nuclear factor-kappaB activity using a sensitive oligo-based chemiluminescent enzyme-linked immunosorbent assay. Int. J. Cancer 2010, 127, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Pepper, C.; Mahdi, J.G.; Buggins, A.G.; Hewamana, S.; Walsby, E.; Mahdi, E.; Al-Haza’a, A.; Mahdi, A.J.; Lin, T.T.; Pearce, L.; et al. Two novel aspirin analogues show selective cytotoxicity in primary chronic lymphocytic leukaemia cells that is associated with dual inhibition of Rel A and COX-2. Cell Prolif. 2011, 44, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhou, P.; Wang, W.; Sun, A.; Guo, F. RelB, together with RelA, sustains cell survival and confers proteasome inhibitor sensitivity of chronic lymphocytic leukemia cells from bone marrow. J. Mol. Med. 2014, 92, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Grosjean-Raillard, J.; Tailler, M.; Adès, L.; Perfettini, J.L.; Fabre, C.; Braun, T.; De Botton, S.; Fenaux, P.; Kroemer, G. ATM mediates constitutive NF-κB activation in high-risk myelodysplastic syndrome and acute myeloid leukemia. Oncogene 2009, 28, 1099–1109. [Google Scholar] [CrossRef]
- Tromp, J.M.; Tonino, S.H.; Elias, J.A.; Jaspers, A.; Luijks, D.M.; Kater, A.P.; van Lier, R.A.; van Oers, M.H.; Eldering, E. Dichotomy in NF-kappaB signaling and chemoresistance in immunoglobulin variable heavy-chain-mutated versus unmutated CLL cells upon CD40/TLR9 triggering. Oncogene 2010, 29, 5071–5082. [Google Scholar] [CrossRef]
- Coscia, M.; Pantaleoni, F.; Riganti, C.; Vitale, C.; Rigoni, M.; Peola, S.; Castella, B.; Foglietta, M.; Griggio, V.; Drandi, D.; et al. IGHV unmutated CLL B cells are more prone to spontaneous apoptosis and subject to environmental prosurvival signals than mutated CLL B cells. Leukemia 2011, 25, 828–837. [Google Scholar] [CrossRef]
- Burley, T.A.; Kennedy, E.; Broad, G.; Boyd, M.; Li, D.; Woo, T.; West, C.; Ladikou, E.E.; Ashworth, I.; Fegan, C.; et al. Targeting the Non-Canonical NF-κB Pathway in Chronic Lymphocytic Leukemia and Multiple Myeloma. Cancers 2022, 14, 1489. [Google Scholar] [CrossRef]
- Haselager, M.; Thijssen, R.; West, C.; Young, L.; Van Kampen, R.; Willmore, E.; Mackay, S.; Kater, A.; Eldering, E. Regulation of Bcl-XL by non-canonical NF-κB in the context of CD40-induced drug resistance in CLL. Cell Death Differ. 2021, 28, 1658–1668. [Google Scholar] [CrossRef]
- Eluard, B.; Nuan-Aliman, S.; Faumont, N.; Collares, D.; Bordereaux, D.; Montagne, A.; Martins, I.; Cagnard, N.; Caly, M.; Taoui, O.; et al. The alternative RelB NF-κB subunit is a novel critical player in diffuse large B-cell lymphoma. Blood 2022, 139, 384–398. [Google Scholar] [CrossRef]
- Ranuncolo, S.M.; Pittaluga, S.; Evbuomwan, M.O.; Jaffe, E.S.; Lewis, B.A. Hodgkin lymphoma requires stabilized NIK and constitutive RelB expression for survival. Blood 2012, 120, 3756–3763. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulligan, E.A.; Tudhope, S.J.; Hunter, J.E.; Clift, A.E.G.; Elliott, S.L.; Summerfield, G.P.; Wallis, J.; Pepper, C.J.; Durkacz, B.; Veuger, S.; et al. Expression and Activity of the NF-κB Subunits in Chronic Lymphocytic Leukaemia: A Role for RelB and Non-Canonical Signalling. Cancers 2023, 15, 4736. https://doi.org/10.3390/cancers15194736
Mulligan EA, Tudhope SJ, Hunter JE, Clift AEG, Elliott SL, Summerfield GP, Wallis J, Pepper CJ, Durkacz B, Veuger S, et al. Expression and Activity of the NF-κB Subunits in Chronic Lymphocytic Leukaemia: A Role for RelB and Non-Canonical Signalling. Cancers. 2023; 15(19):4736. https://doi.org/10.3390/cancers15194736
Chicago/Turabian StyleMulligan, Evan A., Susan J. Tudhope, Jill E. Hunter, Arabella E. G. Clift, Sarah L. Elliott, Geoffrey P. Summerfield, Jonathan Wallis, Chris J. Pepper, Barabara Durkacz, Stephany Veuger, and et al. 2023. "Expression and Activity of the NF-κB Subunits in Chronic Lymphocytic Leukaemia: A Role for RelB and Non-Canonical Signalling" Cancers 15, no. 19: 4736. https://doi.org/10.3390/cancers15194736
APA StyleMulligan, E. A., Tudhope, S. J., Hunter, J. E., Clift, A. E. G., Elliott, S. L., Summerfield, G. P., Wallis, J., Pepper, C. J., Durkacz, B., Veuger, S., & Willmore, E. (2023). Expression and Activity of the NF-κB Subunits in Chronic Lymphocytic Leukaemia: A Role for RelB and Non-Canonical Signalling. Cancers, 15(19), 4736. https://doi.org/10.3390/cancers15194736