
Understanding the Notch Signaling Pathway in Acute Myeloid Leukemia Stem Cells: From Hematopoiesis to Neoplasia



Abstract
:Simple Summary
Abstract
1. Introduction
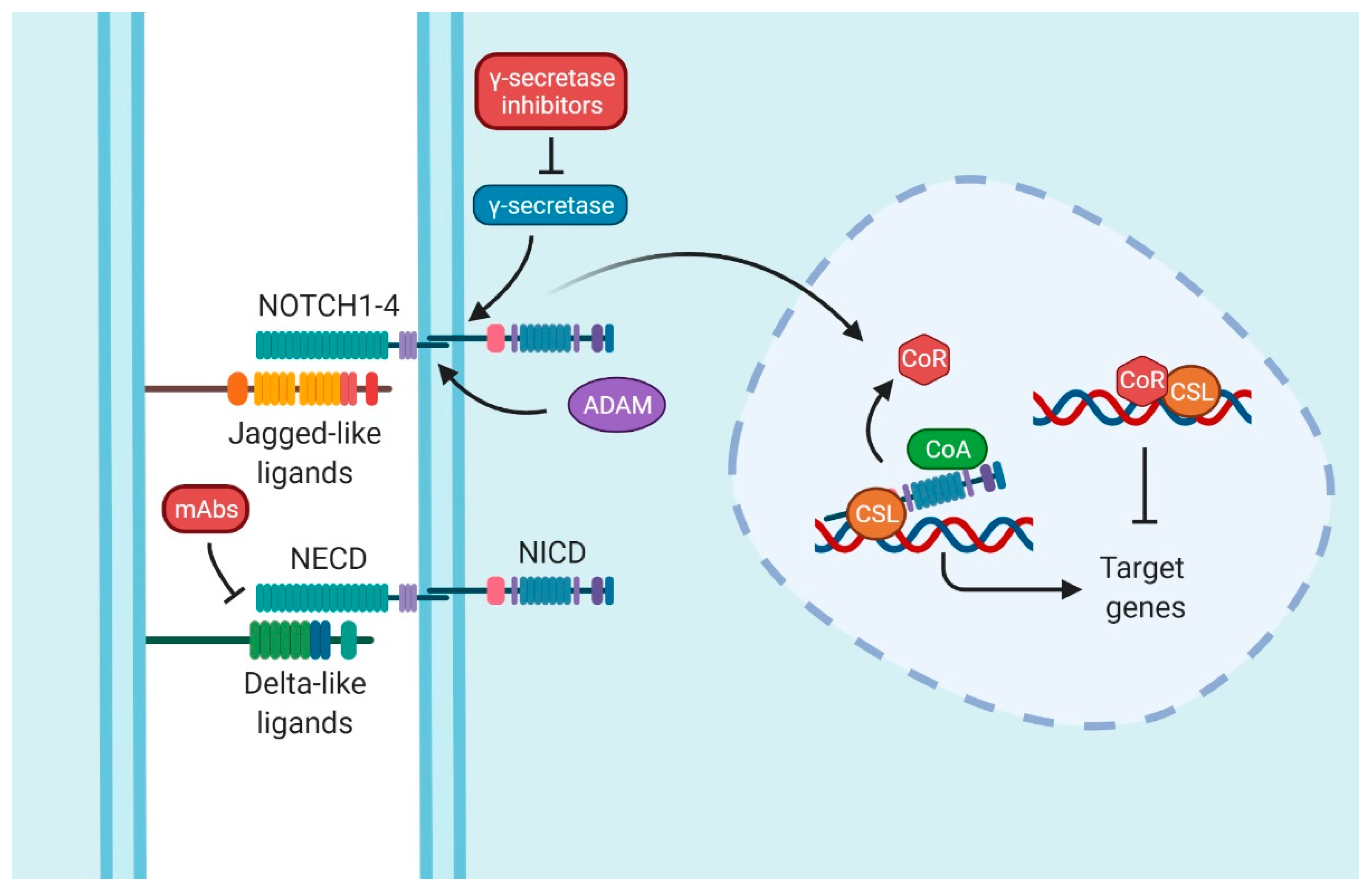
2. Notch Signaling Pathway
3. Notch Signaling in Hematopoiesis
4. The Role of Notch in Hematological Neoplasia
5. Crosstalk and Non-Canonical Activation
6. Discussion
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Maillard, I.; Adler, S.H.; Pear, W.S. Notch and the Immune System. Immunity 2003, 19, 781–791. [Google Scholar] [CrossRef] [Green Version]
- Abel, E.V.; Kim, E.J.; Wu, J.; Hynes, M.; Bednar, F.; Proctor, E.; Wang, L.; Dziubinski, M.L.; Simeone, D.M. The Notch Pathway Is Important in Maintaining the Cancer Stem Cell Population in Pancreatic Cancer. PLoS ONE 2014, 9, e91983. [Google Scholar] [CrossRef] [PubMed]
- Dexter, J.S. The Analysis of a Case of Continuous Variation in Drosophila by a Study of Its Linkage Relations. Am. Nat. 1914, 48, 712–758. [Google Scholar] [CrossRef]
- Morgan, T.H. The Theory of the Gene. Am. Nat. 1917, 51, 513–544. [Google Scholar] [CrossRef]
- Wharton, K.A.; Johansen, K.M.; Xu, T.; Artavanis-Tsakonas, S. Nucleotide Sequence from the Neurogenic Locus Notch Implies a Gene Product That Shares Homology with Proteins Containing EGF-like Repeats. Cell 1985, 43, 567–581. [Google Scholar] [CrossRef]
- Greenwald, I. LIN-12/Notch Signaling: Lessons from Worms and Flies. Genes Dev. 1998, 12, 1751–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, A.; Yoshino, M.; Hikosaka, M.; Okuyama, K.; Zhou, L.; Sakano, S.; Yagita, H.; Hayashi, S.-I. An Evolutionary-Conserved Function of Mammalian Notch Family Members as Cell Adhesion Molecules. PLoS ONE 2014, 9, e108535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Pompa, J.L.; Wakeham, A.; Correia, K.M.; Samper, E.; Brown, S.; Aguilera, R.J.; Nakano, T.; Honjo, T.; Mak, T.W.; Rossant, J.; et al. Conservation of the Notch Signalling Pathway in Mammalian Neurogenesis. Development 1997, 124, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Kopan, R.; Ilagan, M.X.G. The Canonical Notch Signaling Pathway: Unfolding the Activation Mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Souza, B.; Meloty-Kapella, L.; Weinmaster, G. Canonical and Non-Canonical Notch Ligands. Curr. Top. Dev. Biol. 2010, 92, 73–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radtke, F.; Raj, K. The Role of Notch in Tumorigenesis: Oncogene or Tumour Suppressor? Nat. Rev. Cancer 2003, 3, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Fortini, M.E.; Artavanis-Tsakonas, S. The Suppressor of Hairless Protein Participates in Notch Receptor Signaling. Cell 1994, 79, 273–282. [Google Scholar] [CrossRef]
- Kopan, R. Notch Signaling. Cold Spring Harb. Perspect. Biol. 2012, 4, a011213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Cornejo, H.; Saucedo-Correa, G.; Oviedo-Boyso, J.; Valdez-Alarcón, J.J.; Baizabal-Aguirre, V.M.; Cajero-Juárez, M.; Bravo-Patiño, A. The CSL Proteins, Versatile Transcription Factors and Context Dependent Corepressors of the Notch Signaling Pathway. Cell Div. 2016, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Weng, A.P.; Millholland, J.M.; Yashiro-Ohtani, Y.; Arcangeli, M.L.; Lau, A.; Wai, C.; Del Bianco, C.; Rodriguez, C.G.; Sai, H.; Tobias, J.; et al. C-Myc Is an Important Direct Target of Notch1 in T-Cell Acute Lymphoblastic Leukemia/Lymphoma. Genes Dev. 2006, 20, 2096–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronchini, C.; Capobianco, A.J. Induction of Cyclin D1 Transcription and CDK2 Activity by Notch(Ic): Implication for Cell Cycle Disruption in Transformation by Notch(Ic). Mol. Cell. Biol. 2001, 21, 5925–5934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangarajan, A.; Talora, C.; Okuyama, R.; Nicolas, M.; Mammucari, C.; Oh, H.; Aster, J.C.; Krishna, S.; Metzger, D.; Chambon, P.; et al. Notch Signaling Is a Direct Determinant of Keratinocyte Growth Arrest and Entry into Differentiation. EMBO J. 2001, 20, 3427–3436. [Google Scholar] [CrossRef] [Green Version]
- Sionov, R.V.; Kfir-Erenfeld, S.; Spokoini, R.; Yefenof, E. A Role for Bcl-2 in Notch1-Dependent Transcription in Thymic Lymphoma Cells. Adv. Hematol. 2012, 2012, 435241. Available online: https://www.hindawi.com/journals/ah/2012/435241/ (accessed on 25 September 2020). [CrossRef] [PubMed]
- Wang, L.; Song, G.; Liu, M.; Chen, B.; Chen, Y.; Shen, Y.; Zhu, J.; Zhou, X. MicroRNA-375 Overexpression Influences P19 Cell Proliferation, Apoptosis and Differentiation through the Notch Signaling Pathway. Int. J. Mol. Med. 2016, 37, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, R.; Ohtsuka, T.; Kobayashi, T. The Hes Gene Family: Repressors and Oscillators That Orchestrate Embryogenesis. Development 2007, 134, 1243–1251. [Google Scholar] [CrossRef] [Green Version]
- Adler, S.H.; Chiffoleau, E.; Xu, L.; Dalton, N.M.; Burg, J.M.; Wells, A.D.; Wolfe, M.S.; Turka, L.A.; Pear, W.S. Notch Signaling Augments T Cell Responsiveness by Enhancing CD25 Expression. J. Immunol. 2003, 171, 2896–2903. [Google Scholar] [CrossRef] [PubMed]
- Robert-Moreno, À.; Espinosa, L.; de la Pompa, J.L.; Bigas, A. RBPjκ-Dependent Notch Function Regulates Gata2 and Is Essential for the Formation of Intra-Embryonic Hematopoietic Cells. Development 2005, 132, 1117–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feyerabend, T.B.; Terszowski, G.; Tietz, A.; Blum, C.; Luche, H.; Gossler, A.; Gale, N.W.; Radtke, F.; Fehling, H.J.; Rodewald, H.-R. Deletion of Notch1 Converts Pro-T Cells to Dendritic Cells and Promotes Thymic B Cells by Cell-Extrinsic and Cell-Intrinsic Mechanisms. Immunity 2009, 30, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stier, S.; Cheng, T.; Dombkowski, D.; Carlesso, N.; Scadden, D.T. Notch1 Activation Increases Hematopoietic Stem Cell Self-Renewal In Vivo and Favors Lymphoid over Myeloid Lineage Outcome. Blood 2002, 99, 2369–2378. [Google Scholar] [CrossRef] [Green Version]
- Pui, J.C.; Allman, D.; Xu, L.; DeRocco, S.; Karnell, F.G.; Bakkour, S.; Lee, J.Y.; Kadesch, T.; Hardy, R.R.; Aster, J.C.; et al. Notch1 Expression in Early Lymphopoiesis Influences B versus T Lineage Determination. Immunity 1999, 11, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Radtke, F.; Wilson, A.; Stark, G.; Bauer, M.; van Meerwijk, J.; MacDonald, H.R.; Aguet, M. Deficient T Cell Fate Specification in Mice with an Induced Inactivation of Notch1. Immunity 1999, 10, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; von Boehmer, H. Notch Signaling in T-Cell Development and T-ALL. ISRN Hematol 2011, 2011, 921706. [Google Scholar] [CrossRef] [Green Version]
- Jaleco, A.C.; Neves, H.; Hooijberg, E.; Gameiro, P.; Clode, N.; Haury, M.; Henrique, D.; Parreira, L. Differential Effects of Notch Ligands Delta-1 and Jagged-1 in Human Lymphoid Differentiation. J. Exp. Med. 2001, 194, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Garbe, A.I.; Krueger, A.; Gounari, F.; Zúñiga-Pflücker, J.C.; von Boehmer, H. Differential Synergy of Notch and T Cell Receptor Signaling Determines Alphabeta versus Gammadelta Lineage Fate. J. Exp. Med. 2006, 203, 1579–1590. [Google Scholar] [CrossRef] [Green Version]
- Fowlkes, B.J.; Robey, E.A. A Reassessment of the Effect of Activated Notch1 on CD4 and CD8 T Cell Development. J. Immunol. 2002, 169, 1817–1821. [Google Scholar] [CrossRef] [Green Version]
- Singer, A. New Perspectives on a Developmental Dilemma: The Kinetic Signaling Model and the Importance of Signal Duration for the CD4/CD8 Lineage Decision. Curr. Opin. Immunol. 2002, 14, 207–215. [Google Scholar] [CrossRef]
- Kumano, K.; Chiba, S.; Kunisato, A.; Sata, M.; Saito, T.; Nakagami-Yamaguchi, E.; Yamaguchi, T.; Masuda, S.; Shimizu, K.; Takahashi, T.; et al. Notch1 but Not Notch2 Is Essential for Generating Hematopoietic Stem Cells from Endothelial Cells. Immunity 2003, 18, 699–711. [Google Scholar] [CrossRef]
- Maillard, I.; Koch, U.; Dumortier, A.; Shestova, O.; Xu, L.; Sai, H.; Pross, S.E.; Aster, J.C.; Bhandoola, A.; Radtke, F.; et al. Canonical Notch Signaling Is Dispensable for the Maintenance of Adult Hematopoietic Stem Cells. Cell Stem Cell 2008, 2, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Woll, P.S.; Buza-Vidas, N.; Chin, D.W.L.; Boukarabila, H.; Luís, T.C.; Stenson, L.; Bouriez-Jones, T.; Ferry, H.; Mead, A.J.; et al. Canonical Notch Signaling Is Dispensable for Adult Steady-State and Stress Myelo-Erythropoiesis. Blood 2018, 131, 1712–1719. [Google Scholar] [CrossRef] [Green Version]
- Weng, A.P.; Ferrando, A.A.; Lee, W.; Morris, J.P.; Silverman, L.B.; Sanchez-Irizarry, C.; Blacklow, S.C.; Look, A.T.; Aster, J.C. Activating Mutations of NOTCH1 in Human T Cell Acute Lymphoblastic Leukemia. Science 2004, 306, 269–271. [Google Scholar] [CrossRef] [Green Version]
- Zheng, R.-Y.; Wang, S.-J.; Wang, C.; Li, T.; Liao, L.-X.; Li, M.-L.; Chen, S.-M.; Guo, R.; Wang, W.-Q.; Zhang, Y.; et al. Gene Mutation in Acute Lymphoblastic Leukemia by DNA Sequencing. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2020, 28, 1791–1795. [Google Scholar] [CrossRef]
- Aref, S.; El Agdar, M.; Salama, O.; Zeid, T.A.; Sabry, M. Significance of NOTCH1 Mutations Détections in T-Acute Lymphoblastic Leukemia Patients. Cancer Biomark. Sect. A Dis. Markers 2020, 27, 157–162. [Google Scholar] [CrossRef]
- Taj, M.M.; Moorman, A.V.; Hamadeh, L.; Petit, A.; Asnafi, V.; Alby-Laurent, F.; Vora, A.; Mansour, M.R.; Gale, R.; Chevret, S.; et al. Prognostic Value of Oncogenetic Mutations in Pediatric T Acute Lymphoblastic Leukemia: A Comparison of UKALL2003 and FRALLE2000T Protocols. Leukemia 2022, 36, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Pelullo, M.; Quaranta, R.; Talora, C.; Checquolo, S.; Cialfi, S.; Felli, M.P.; te Kronnie, G.; Borga, C.; Besharat, Z.M.; Palermo, R.; et al. Notch3/Jagged1 Circuitry Reinforces Notch Signaling and Sustains T-ALL. Neoplasia 2014, 16, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, G.; Rasi, S.; Rossi, D.; Trifonov, V.; Khiabanian, H.; Ma, J.; Grunn, A.; Fangazio, M.; Capello, D.; Monti, S.; et al. Analysis of the Chronic Lymphocytic Leukemia Coding Genome: Role of NOTCH1 Mutational Activation. J. Exp. Med. 2011, 208, 1389–1401. [Google Scholar] [CrossRef] [Green Version]
- Rossi, D.; Rasi, S.; Fabbri, G.; Spina, V.; Fangazio, M.; Forconi, F.; Marasca, R.; Laurenti, L.; Bruscaggin, A.; Cerri, M.; et al. Mutations of NOTCH1 Are an Independent Predictor of Survival in Chronic Lymphocytic Leukemia. Blood 2012, 119, 521–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeu, F.; Delgado, J.; Royo, C.; Baumann, T.; Stankovic, T.; Pinyol, M.; Jares, P.; Navarro, A.; Martín-García, D.; Beà, S.; et al. Clinical Impact of Clonal and Subclonal TP53, SF3B1, BIRC3, NOTCH1, and ATM Mutations in Chronic Lymphocytic Leukemia. Blood 2016, 127, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Klinakis, A.; Lobry, C.; Abdel-Wahab, O.; Oh, P.; Haeno, H.; Buonamici, S.; van De Walle, I.; Cathelin, S.; Trimarchi, T.; Araldi, E.; et al. A Novel Tumour-Suppressor Function for the Notch Pathway in Myeloid Leukaemia. Nature 2011, 473, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Aref, S.; Rizk, R.; El Agder, M.; Fakhry, W.; El Zafarany, M.; Sabry, M. NOTCH-1 Gene Mutations Influence Survival in Acute Myeloid Leukemia Patients. Asian Pac. J. Cancer Prev. 2020, 21, 1987–1992. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhao, Y.; Xu, M.; Dai, Q.; Meng, W.; Yang, J.; Qin, R. Activation of Notch Signal Pathway Is Associated with a Poorer Prognosis in Acute Myeloid Leukemia. Med. Oncol. 2011, 28 (Suppl. 1), 483–489. [Google Scholar] [CrossRef] [PubMed]
- Kannan, S.; Sutphin, R.M.; Hall, M.G.; Golfman, L.S.; Fang, W.; Nolo, R.M.; Akers, L.J.; Hammitt, R.A.; McMurray, J.S.; Kornblau, S.M.; et al. Notch Activation Inhibits AML Growth and Survival: A Potential Therapeutic Approach. J. Exp. Med. 2013, 210, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-A.; Pietras, E.M.; Passegué, E. Deregulated Notch and Wnt Signaling Activates Early-Stage Myeloid Regeneration Pathways in Leukemia. J. Exp. Med. 2020, 217, e20190787. [Google Scholar] [CrossRef] [PubMed]
- Lobry, C.; Ntziachristos, P.; Ndiaye-Lobry, D.; Oh, P.; Cimmino, L.; Zhu, N.; Araldi, E.; Hu, W.; Freund, J.; Abdel-Wahab, O.; et al. Notch Pathway Activation Targets AML-Initiating Cell Homeostasis and Differentiation. J. Exp. Med. 2013, 210, 301–319. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Li, T.; Shang, Z.; Zhao, L.; Xu, Q.; Tan, J.; Qin, Y.; Zhang, Y.; Cao, Y.; Wang, N.; et al. Combined Inhibition of Notch and FLT3 Produces Synergistic Cytotoxic Effects in FLT3/ITD+ Acute Myeloid Leukemia. Signal Transduct. Target. Ther. 2020, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Li, G.-H.; Fan, Y.-Z.; Liu, X.-W.; Zhang, B.-F.; Yin, D.-D.; He, F.; Huang, S.-Y.; Kang, Z.-J.; Xu, H.; Liu, Q.; et al. Notch Signaling Maintains Proliferation and Survival of the HL60 Human Promyelocytic Leukemia Cell Line and Promotes the Phosphorylation of the Rb Protein. Mol. Cell. Biochem. 2010, 340, 7–14. [Google Scholar] [CrossRef]
- Takam Kamga, P.; Collo, G.D.; Resci, F.; Bazzoni, R.; Mercuri, A.; Quaglia, F.M.; Tanasi, I.; Delfino, P.; Visco, C.; Bonifacio, M.; et al. Notch Signaling Molecules as Prognostic Biomarkers for Acute Myeloid Leukemia. Cancers 2019, 11, 1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Varol, M.; Tuorkey, M.; Sak, K.; Parashar, N.C.; Barwal, T.S.; Sharma, U.; Iqubal, A.; Parashar, G.; et al. NOTCH Signaling: Journey of an Evolutionarily Conserved Pathway in Driving Tumor Progression and Its Modulation as a Therapeutic Target. Crit. Rev. Oncol./Hematol. 2021, 164, 103403. [Google Scholar] [CrossRef] [PubMed]
- Deangelo, D.J.; Stone, R.M.; Silverman, L.B.; Stock, W.; Attar, E.C.; Fearen, I.; Dallob, A.; Matthews, C.; Stone, J.; Freedman, S.J.; et al. A Phase I Clinical Trial of the Notch Inhibitor MK-0752 in Patients with T-Cell Acute Lymphoblastic Leukemia/Lymphoma (T-ALL) and Other Leukemias. J. Clin. Oncol. 2006, 24, 6585. [Google Scholar] [CrossRef]
- Zweidler-McKay, P.A.; DeAngelo, D.J.; Douer, D.; Dombret, H.; Ottmann, O.G.; Vey, N.; Thomas, D.A.; Zhu, L.; Huang, F.; Bajaj, G.; et al. The Safety and Activity of BMS-906024, a Gamma Secretase Inhibitor (GSI) with Anti-Notch Activity, in Patients with Relapsed T-Cell Acute Lymphoblastic Leukemia (T-ALL): Initial Results of a Phase 1 Trial. Blood 2014, 124, 968. [Google Scholar] [CrossRef]
- SpringWorks Therapeutics, Inc. A Randomized, Double-Blind, Placebo-Controlled, Phase 3 Trial of Nirogacestat Versus Placebo in Adult Patients With Progressing Desmoid Tumors/Aggressive Fibromatosis (DT/AF). 2021. Available online: https://www.clinicaltrials.gov/ (accessed on 30 November 2021).
- Kwon, C.; Cheng, P.; King, I.N.; Andersen, P.; Shenje, L.; Nigam, V.; Srivastava, D. Notch Post-Translationally Regulates β-Catenin Protein in Stem and Progenitor Cells. Nat. Cell Biol. 2011, 13, 1244–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangolini, M.; Götte, F.; Moore, A.; Ammon, T.; Oelsner, M.; Lutzny-Geier, G.; Klein-Hitpass, L.; Williamson, J.C.; Lehner, P.J.; Dürig, J.; et al. Notch2 Controls Non-Autonomous Wnt-Signalling in Chronic Lymphocytic Leukaemia. Nat. Commun. 2018, 9, 3839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Wu, J.; Yang, J.; Sun, S.; Chai, R.; Chen, Z.-Y.; Li, H. Notch Inhibition Induces Mitotically Generated Hair Cells in Mammalian Cochleae via Activating the Wnt Pathway. Proc. Natl. Acad. Sci. USA 2015, 112, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, W.; Guo, L.; Zhao, L.; Sun, S.; Li, H. The Crosstalk between the Notch, Wnt, and SHH Signaling Pathways in Regulating the Proliferation and Regeneration of Sensory Progenitor Cells in the Mouse Cochlea. Cell Tissue Res. 2021, 386, 281–296. [Google Scholar] [CrossRef]
- Lainez-González, D.; Serrano-López, J.; Alonso-Domínguez, J.M. Understanding the Hedgehog Signaling Pathway in Acute Myeloid Leukemia Stem Cells: A Necessary Step toward a Cure. Biology (Basel) 2021, 10, 255. [Google Scholar] [CrossRef]
- Nicolas, M.; Wolfer, A.; Raj, K.; Kummer, J.A.; Mill, P.; van Noort, M.; Hui, C.; Clevers, H.; Dotto, G.P.; Radtke, F. Notch1 Functions as a Tumor Suppressor in Mouse Skin. Nat. Genet. 2003, 33, 416–421. [Google Scholar] [CrossRef]
- Androutsellis-Theotokis, A.; Leker, R.R.; Soldner, F.; Hoeppner, D.J.; Ravin, R.; Poser, S.W.; Rueger, M.A.; Bae, S.-K.; Kittappa, R.; McKay, R.D.G. Notch Signalling Regulates Stem Cell Numbers in Vitro and in Vivo. Nature 2006, 442, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Ingram, W.J.; McCue, K.I.; Tran, T.H.; Hallahan, A.R.; Wainwright, B.J. Sonic Hedgehog Regulates Hes1 through a Novel Mechanism That Is Independent of Canonical Notch Pathway Signalling. Oncogene 2008, 27, 1489–1500. [Google Scholar] [CrossRef] [PubMed]
- Domingo-Domenech, J.; Vidal, S.J.; Rodriguez-Bravo, V.; Castillo-Martin, M.; Quinn, S.A.; Rodriguez-Barrueco, R.; Bonal, D.M.; Charytonowicz, E.; Gladoun, N.; de la Iglesia-Vicente, J.; et al. Suppression of Acquired Docetaxel Resistance in Prostate Cancer through Depletion of Notch- and Hedgehog-Dependent Tumor-Initiating Cells. Cancer Cell 2012, 22, 373–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doody, R.S.; Raman, R.; Farlow, M.; Iwatsubo, T.; Vellas, B.; Joffe, S.; Kieburtz, K.; He, F.; Sun, X.; Thomas, R.G.; et al. A Phase 3 Trial of Semagacestat for Treatment of Alzheimer’s Disease. N. Engl. J. Med. 2013, 369, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Okuhashi, Y.; Itoh, M.; Tohda, S. GLI1 and CTNNB1 Knockdown Activates NOTCH and MTOR Signalling in NB4 Myeloid Leukaemia Cells. Anticancer Res. 2018, 38, 6329–6332. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, A.; Banerjee, D.; Chandra, S.; Banerji, S.K.; Ghosh, R.; Roy, R.; Banerjee, S. Deregulation and Cross Talk among Sonic Hedgehog, Wnt, Hox and Notch Signaling in Chronic Myeloid Leukemia Progression. Leukemia 2007, 21, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Liu, M.; Gonzalez-Perez, R.R. Role of Notch and Its Oncogenic Signaling Crosstalk in Breast Cancer. Biochim. Biophys. Acta 2011, 1815, 197–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamakura, S.; Oishi, K.; Yoshimatsu, T.; Nakafuku, M.; Masuyama, N.; Gotoh, Y. Hes Binding to STAT3 Mediates Crosstalk between Notch and JAK-STAT Signalling. Nat. Cell Biol. 2004, 6, 547–554. [Google Scholar] [CrossRef]
- Zeng, Q.; Li, S.; Chepeha, D.B.; Giordano, T.J.; Li, J.; Zhang, H.; Polverini, P.J.; Nor, J.; Kitajewski, J.; Wang, C.-Y. Crosstalk between Tumor and Endothelial Cells Promotes Tumor Angiogenesis by MAPK Activation of Notch Signaling. Cancer Cell 2005, 8, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Masiero, M.; Minuzzo, S.; Pusceddu, I.; Moserle, L.; Persano, L.; Agnusdei, V.; Tosello, V.; Basso, G.; Amadori, A.; Indraccolo, S. Notch3-Mediated Regulation of MKP-1 Levels Promotes Survival of T Acute Lymphoblastic Leukemia Cells. Leukemia 2011, 25, 588–598. [Google Scholar] [CrossRef] [Green Version]
- Mittal, S.; Subramanyam, D.; Dey, D.; Kumar, R.V.; Rangarajan, A. Cooperation of Notch and Ras/MAPK Signaling Pathways in Human Breast Carcinogenesis. Mol. Cancer 2009, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornejo, M.G.; Mabialah, V.; Sykes, S.M.; Khandan, T.; Lo Celso, C.; Lopez, C.K.; Rivera-Muñoz, P.; Rameau, P.; Tothova, Z.; Aster, J.C.; et al. Crosstalk between NOTCH and AKT Signaling during Murine Megakaryocyte Lineage Specification. Blood 2011, 118, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Laws, A.M.; Osborne, B.A. P53 Regulates Thymic Notch1 Activation. Eur. J. Immunol. 2004, 34, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Mutvei, A.P.; Chivukula, I.V.; Andersson, E.R.; Ramsköld, D.; Sandberg, R.; Lee, K.L.; Kronqvist, P.; Mamaeva, V.; Ostling, P.; et al. Non-Canonical Notch Signaling Activates IL-6/JAK/STAT Signaling in Breast Tumor Cells and Is Controlled by P53 and IKKα/IKKβ. Oncogene 2013, 32, 4892–4902. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Drug | Mechanism of Action | Clinical Trial | Phase | Status |
|---|---|---|---|---|
| RO4929097 | γ-secretase inhibitor | NCT01158274 | I | C |
| NCT01088763 | I | T | ||
| NCT01236586 | I | W | ||
| LY3039478 | γ-secretase inhibitor | NCT02518113 | I/II | C |
| MK0752 | γ-secretase inhibitor | NCT00100152 | I | T |
| BMS-906024 | γ-secretase inhibitor | NCT01363817 | I | C |
| Nirogacestat PF-03084014 | γ-secretase inhibitor | NCT00878189 | I | C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Láinez-González, D.; Serrano-López, J.; Alonso-Dominguez, J.M. Understanding the Notch Signaling Pathway in Acute Myeloid Leukemia Stem Cells: From Hematopoiesis to Neoplasia. Cancers 2022, 14, 1459. https://doi.org/10.3390/cancers14061459
Láinez-González D, Serrano-López J, Alonso-Dominguez JM. Understanding the Notch Signaling Pathway in Acute Myeloid Leukemia Stem Cells: From Hematopoiesis to Neoplasia. Cancers. 2022; 14(6):1459. https://doi.org/10.3390/cancers14061459
Chicago/Turabian StyleLáinez-González, Daniel, Juana Serrano-López, and Juan Manuel Alonso-Dominguez. 2022. "Understanding the Notch Signaling Pathway in Acute Myeloid Leukemia Stem Cells: From Hematopoiesis to Neoplasia" Cancers 14, no. 6: 1459. https://doi.org/10.3390/cancers14061459
APA StyleLáinez-González, D., Serrano-López, J., & Alonso-Dominguez, J. M. (2022). Understanding the Notch Signaling Pathway in Acute Myeloid Leukemia Stem Cells: From Hematopoiesis to Neoplasia. Cancers, 14(6), 1459. https://doi.org/10.3390/cancers14061459