
IGF-I and Hyaluronic Acid Mitigate the Negative Effect of Irradiation on Human Skin Keratinocytes


 , ,
, ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Isolation of Primary Keratinocytes
2.3. Adipose-Derived Stem Cell Conditioned Medium
2.4. Production of Platelet Lysate
2.5. Cell X-ray Irradiation
2.6. Irradiation of Keratinocyte 2D Cell Cultures
2.7. RNA Isolation, Reverse Transcription, and Quantitative Real-Time PCR
2.8. Irradiation of Keratinocytes on a 3D Collagen-Elastin Matrix
2.9. Statistical Analysis
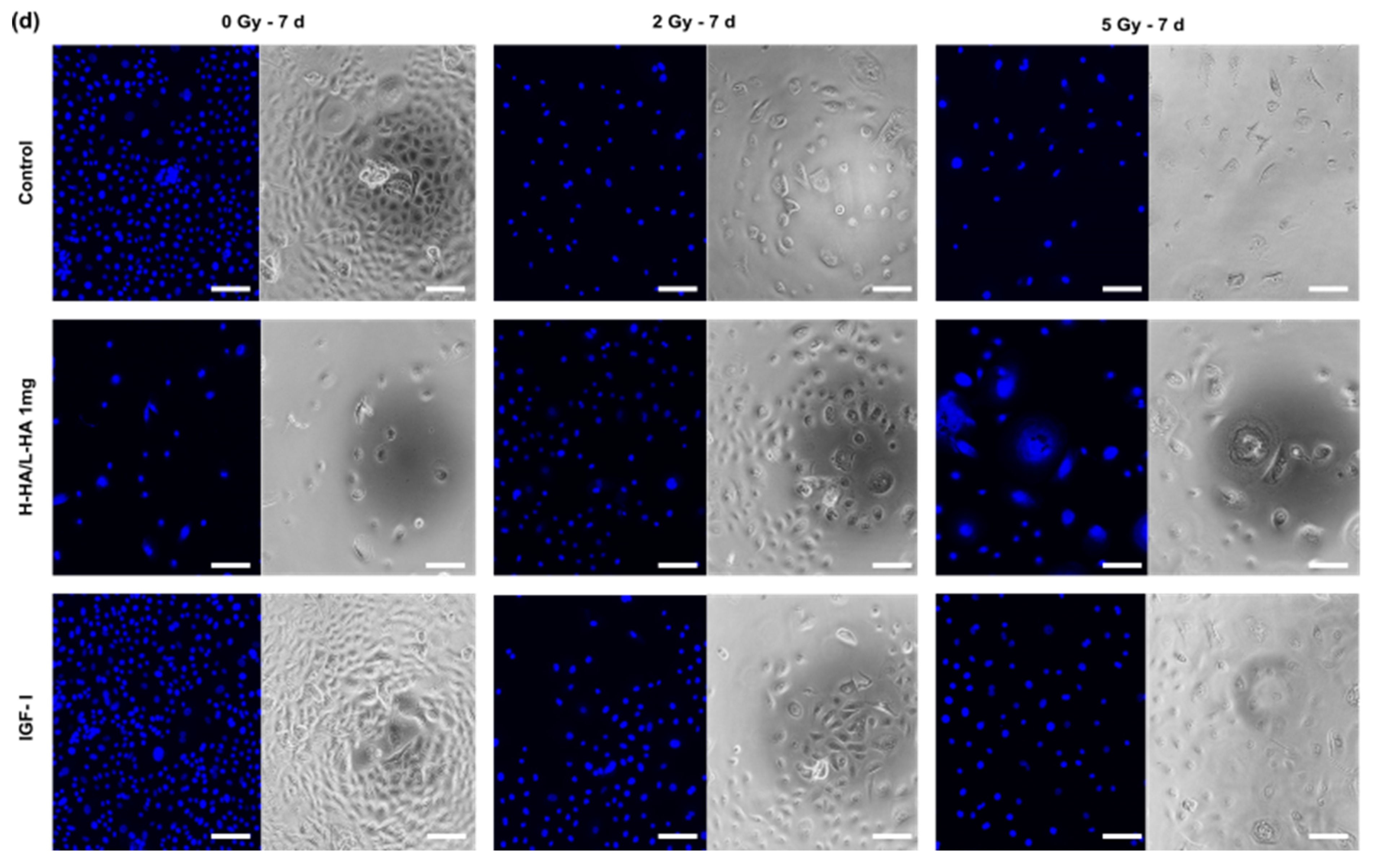
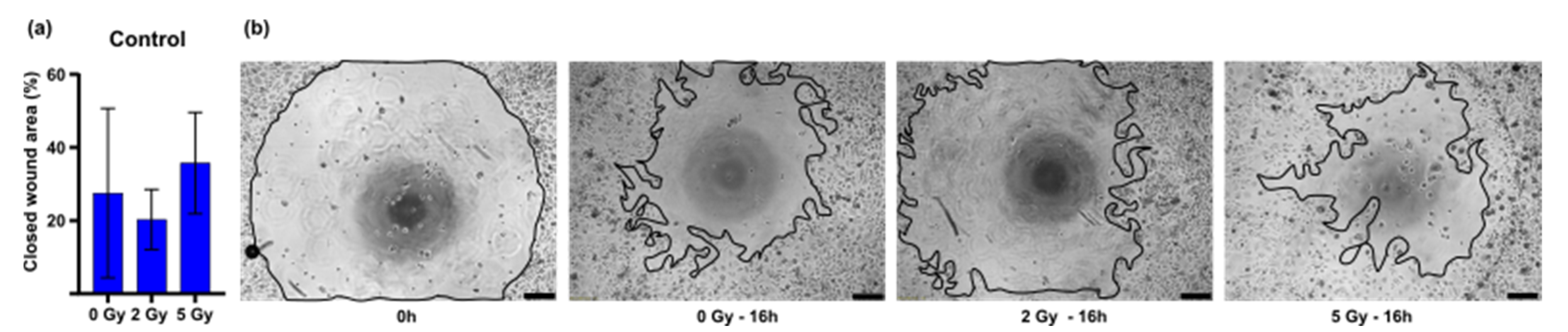
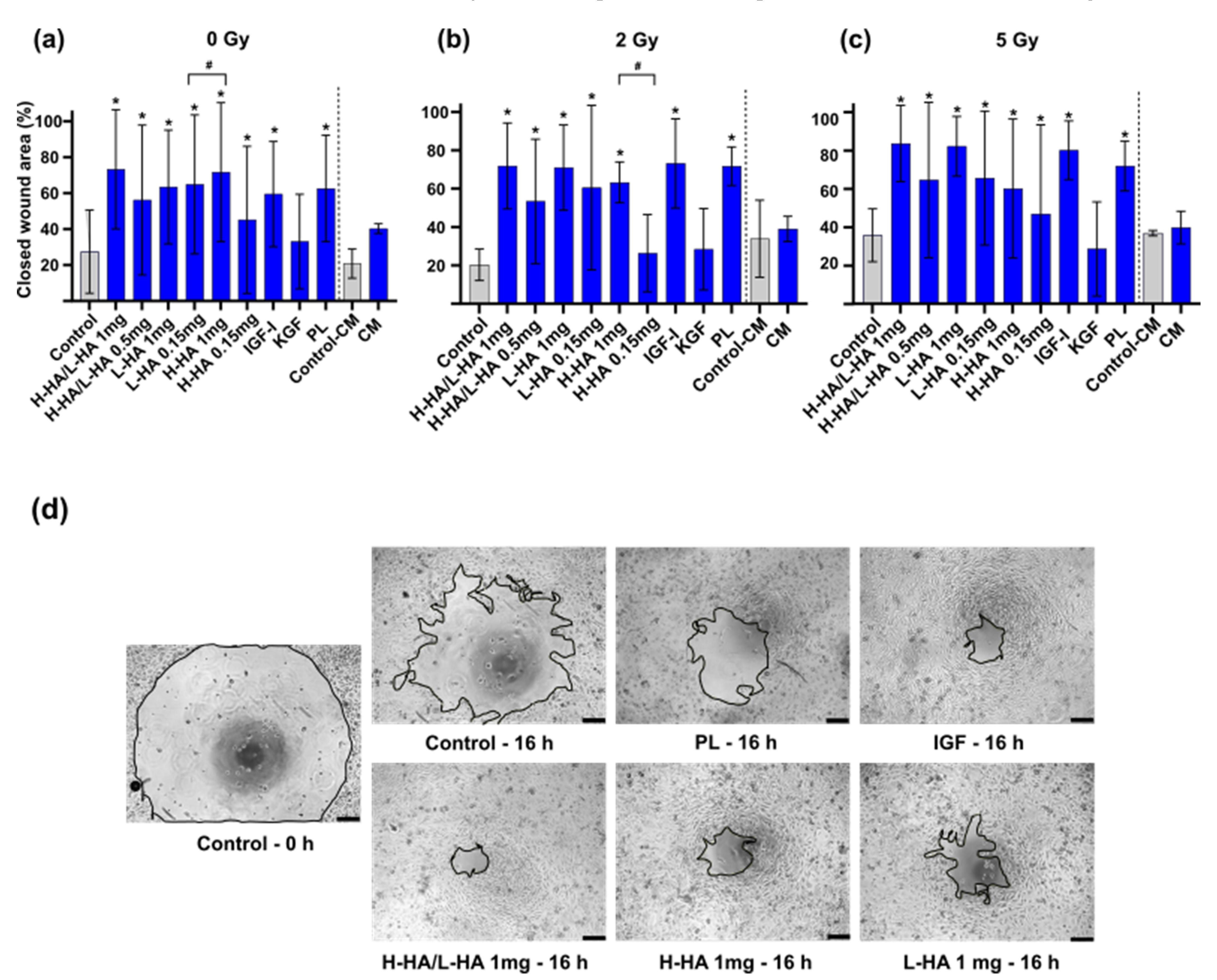
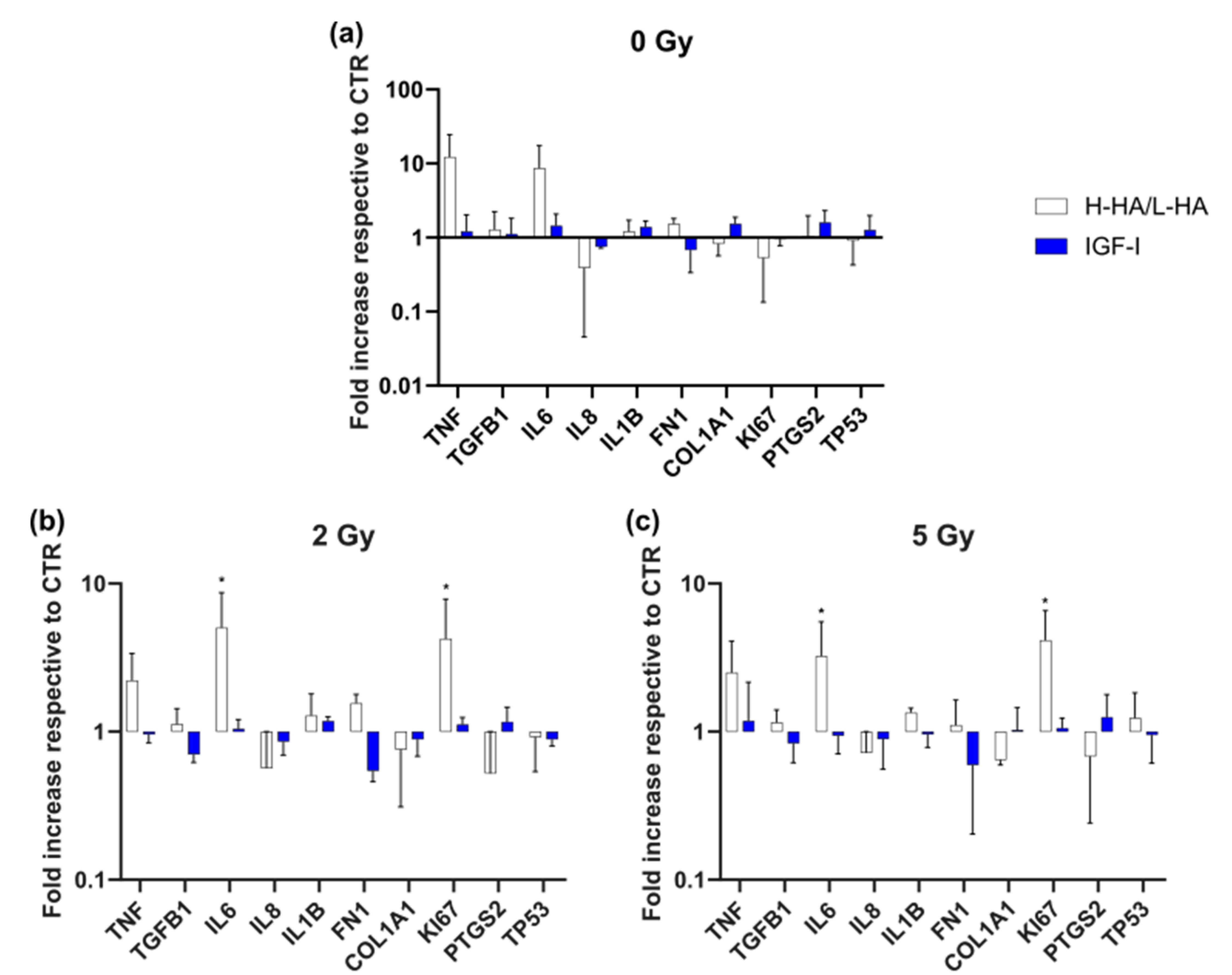
3. Results
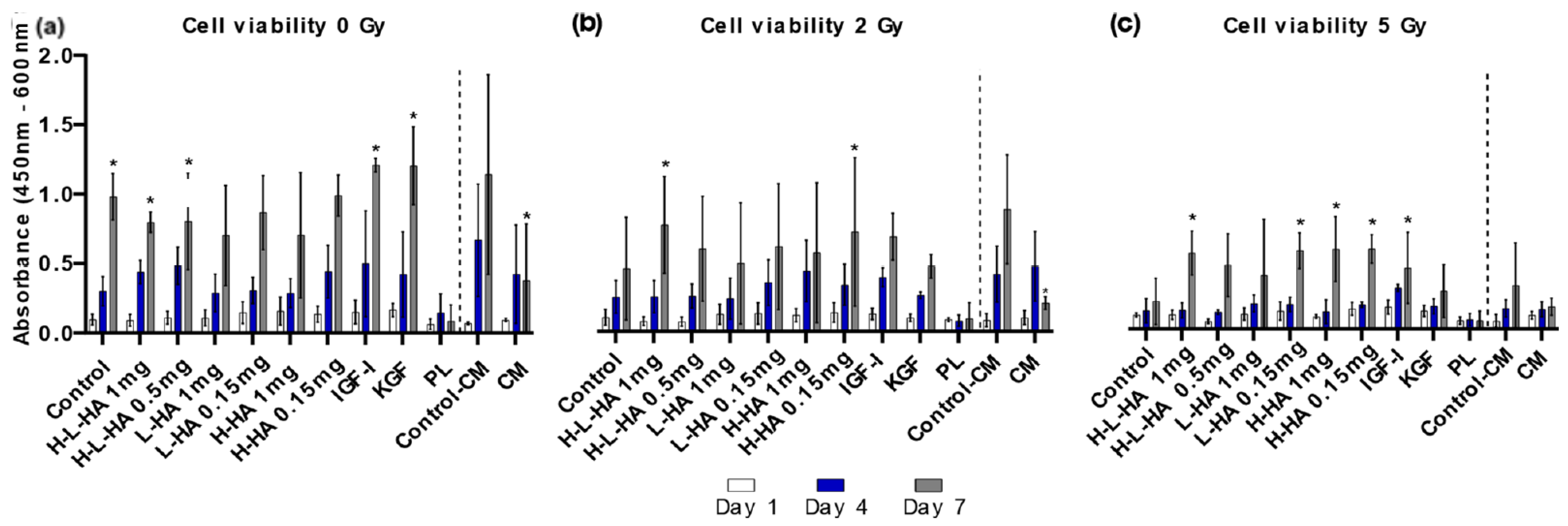
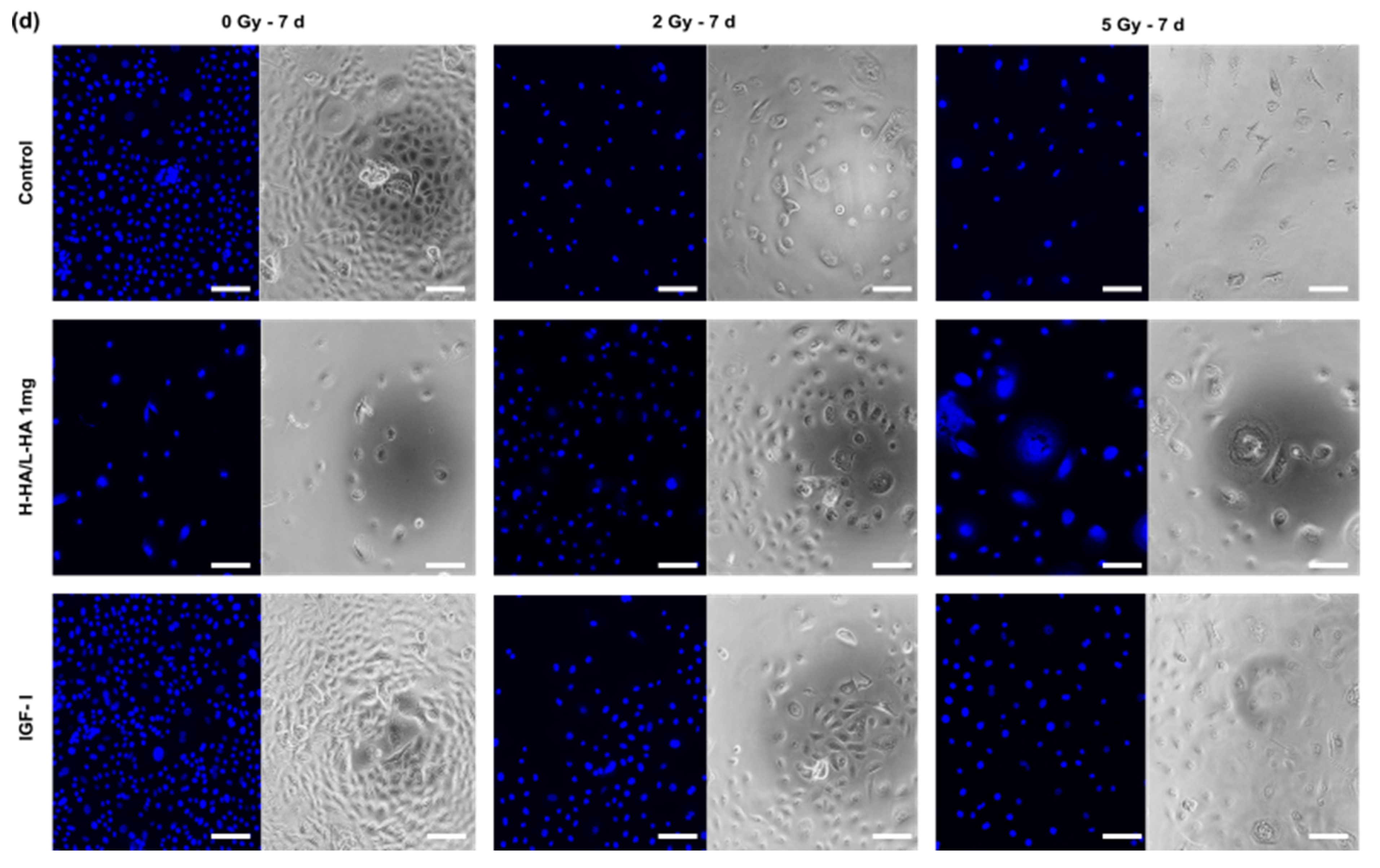
3.1. Cell Viability
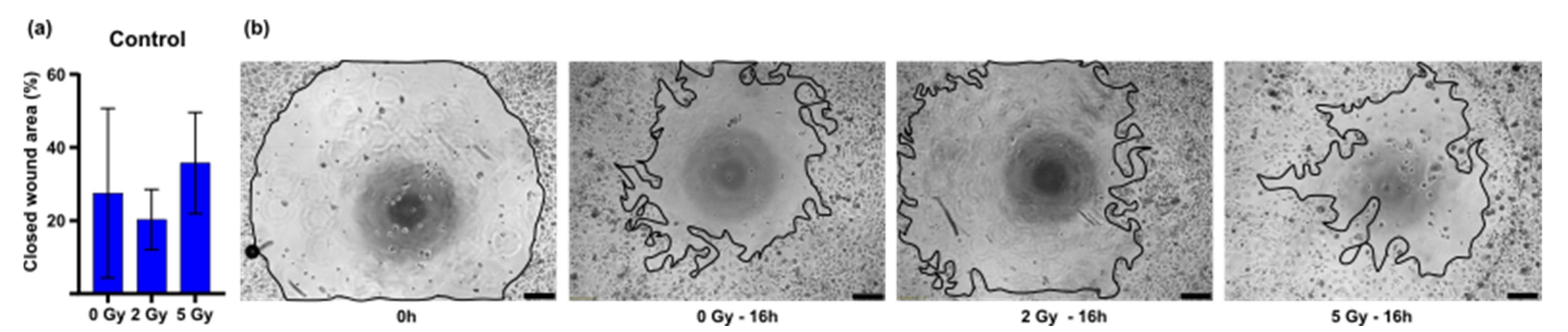
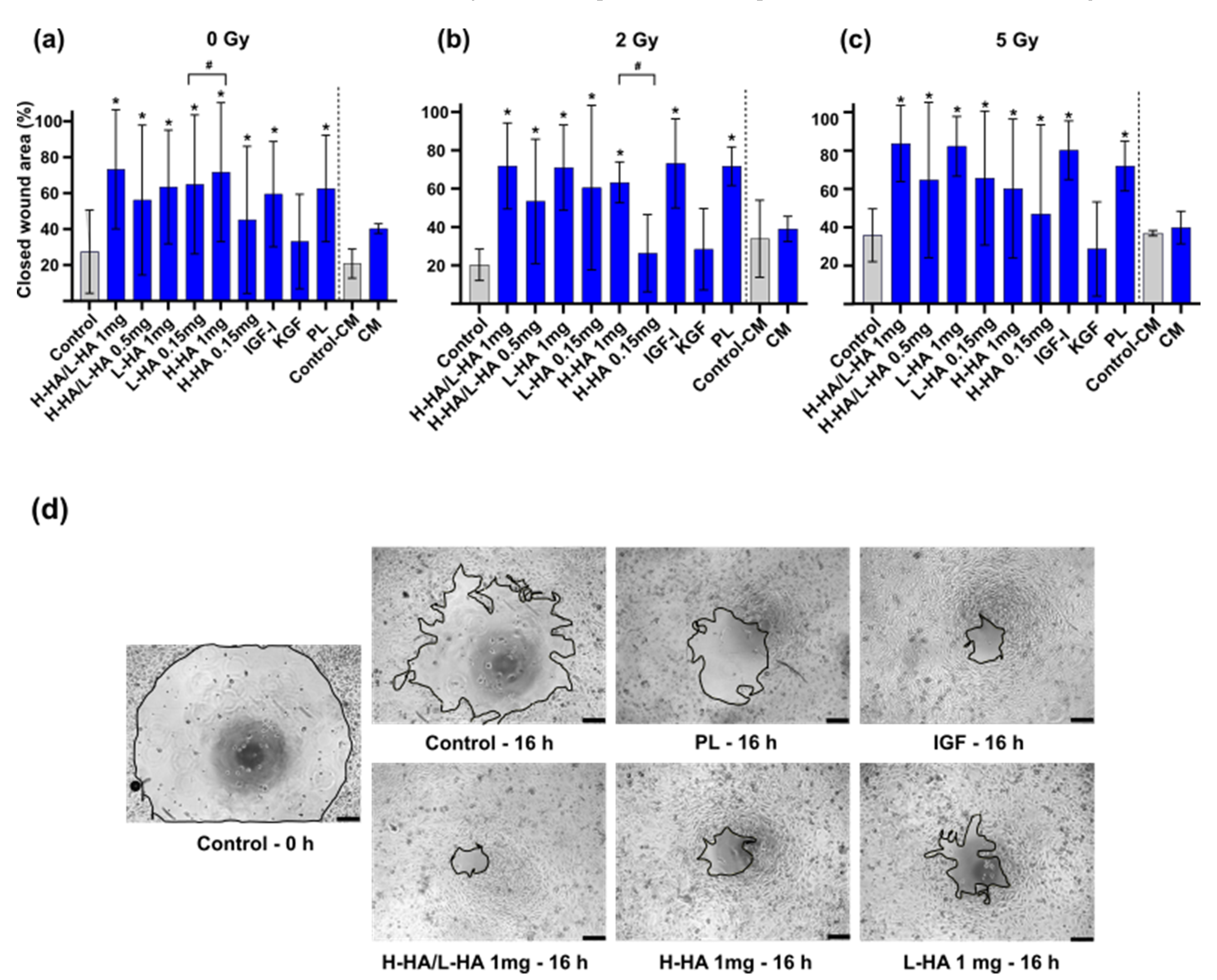
3.2. In Vitro Wound Closure Time
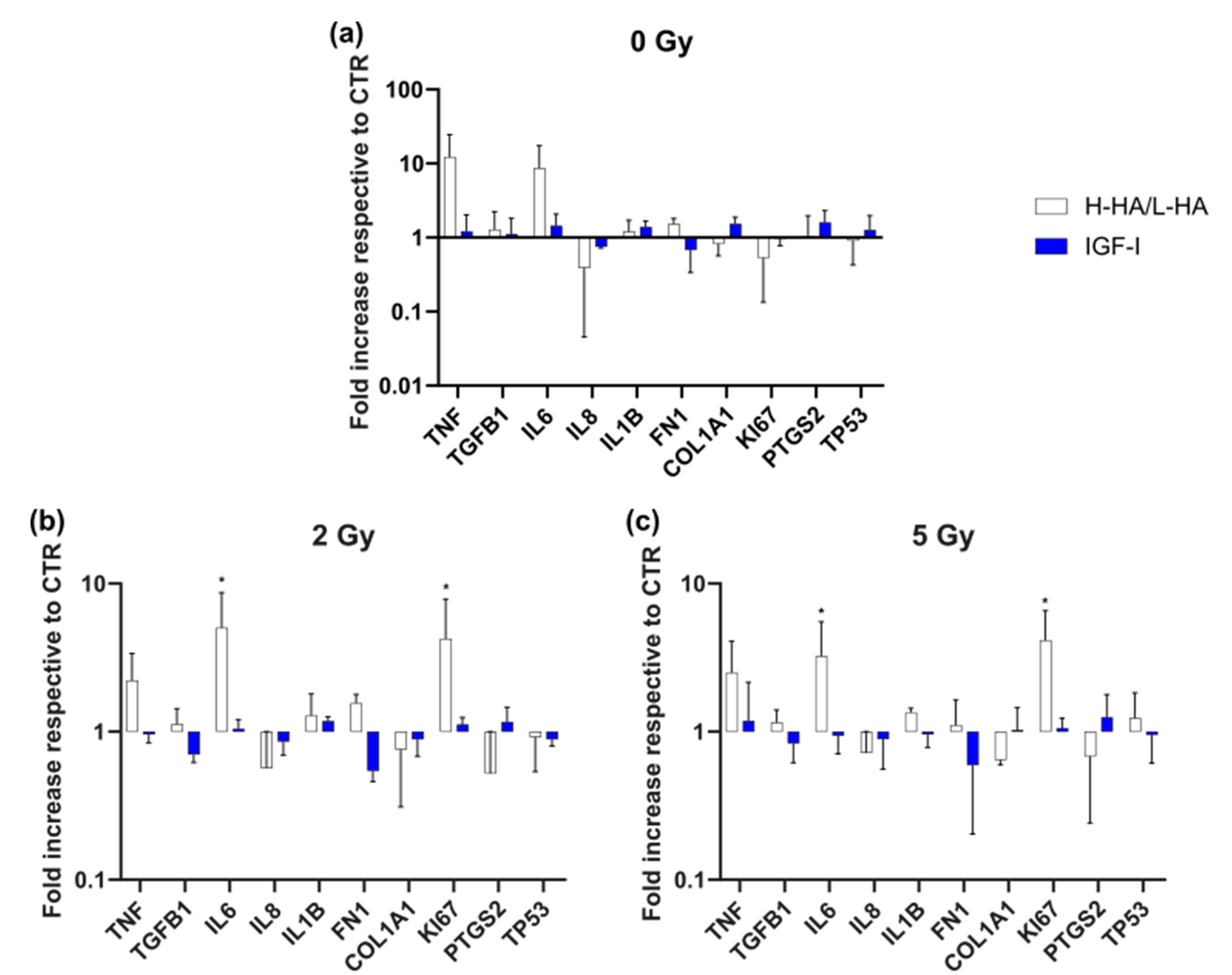
3.3. Gene Expression (qPCR)
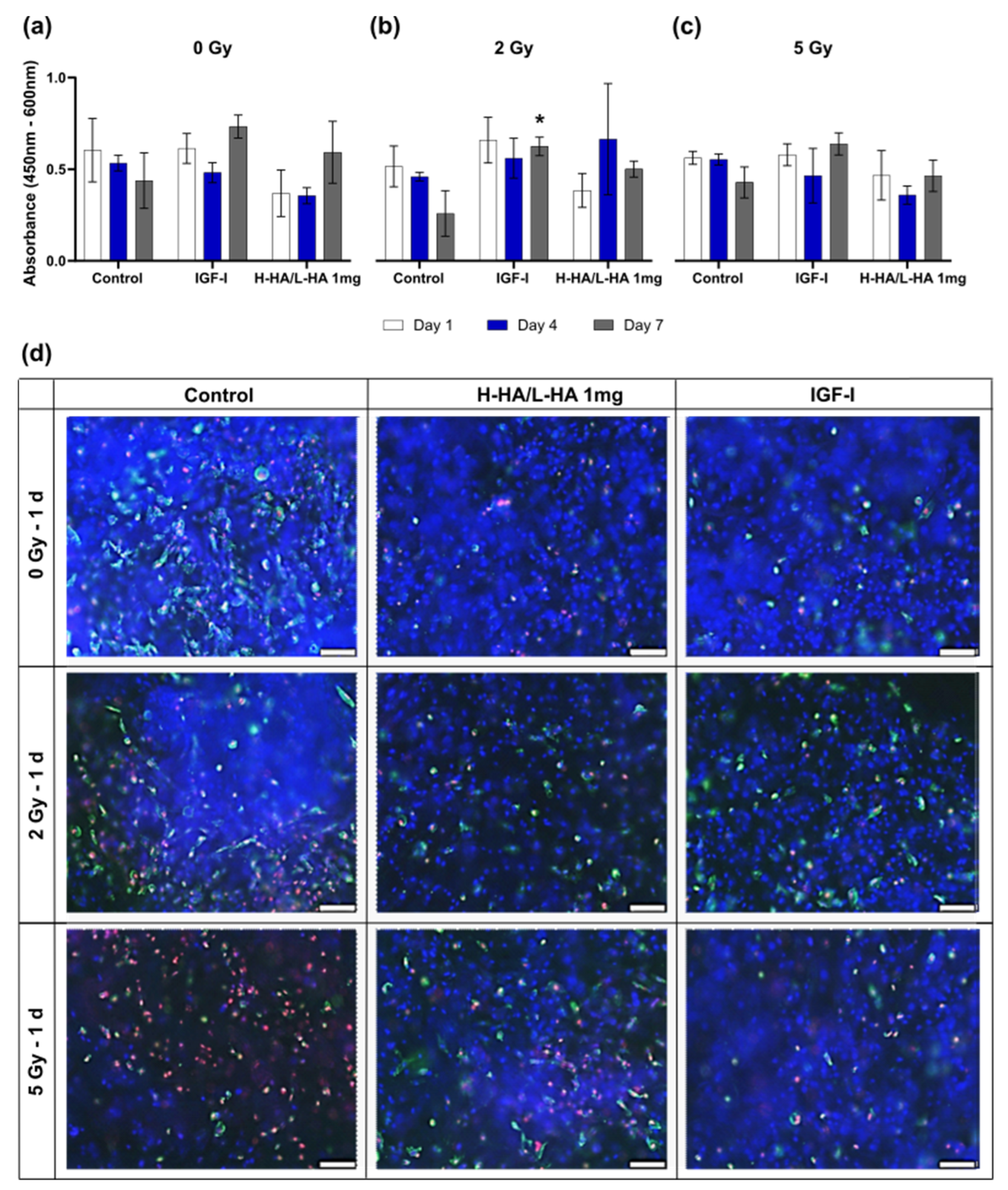
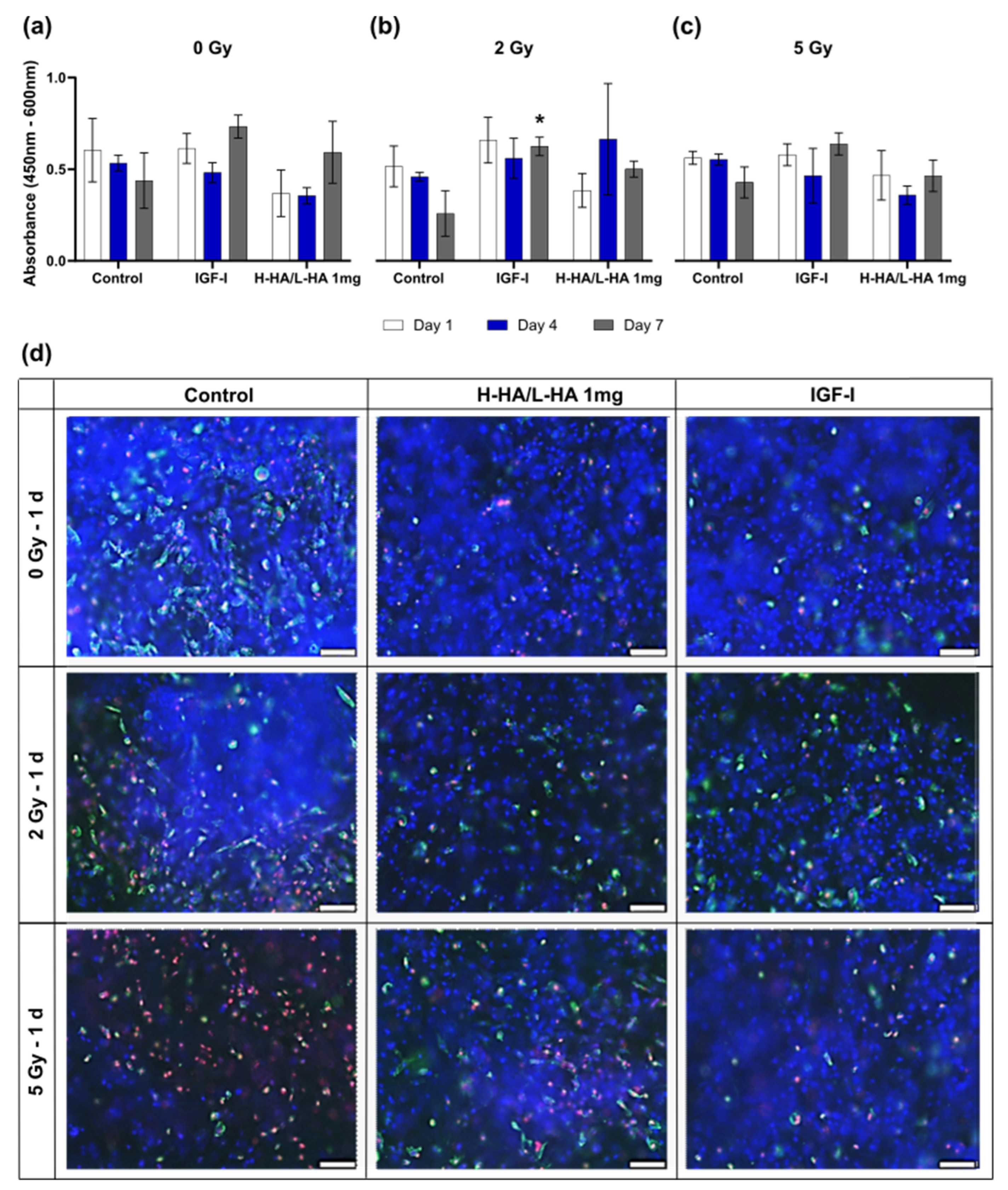
3.4. Cell Viability on the 3D Collagen-Elastin Scaffold
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Porock, D. Factors influencing the severity of radiation skin and oral mucosal reactions: Development of a conceptual framework. Eur. J. Cancer Care 2002, 11, 33–43. [Google Scholar]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horch, R.E.; Ludolph, I.; Arkudas, A. Reconstruction of oncological defects of the perianal region. Chirurg 2021, 92, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Horch, R.E.; Ludolph, I.; Cai, A.; Weber, K.; Grützmann, R.; Arkudas, A. Interdisciplinary Surgical Approaches in Vaginal and Perineal Reconstruction of Advanced Rectal and Anal Female Cancer Patients. Front. Oncol. 2020, 10, 719. [Google Scholar] [CrossRef] [PubMed]
- Götzl, R.; Sterzinger, S.; Arkudas, A.; Boos, A.M.; Semrau, S.; Vassos, N.; Grützmann, R.; Agaimy, A.; Hohenberger, W.; Horch, R.E.; et al. The role of plastic reconstructive surgery in surgical therapy of soft tissue sarcomas. Cancers 2020, 12, 3534. [Google Scholar] [CrossRef] [PubMed]
- Grewal, A.S.; Jones, J.; Lin, A. Palliative Radiation Therapy for Head and Neck Cancers. Int. J. Radiat. Oncol. 2019, 105, 254–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neri, M.; Fabbri, M.; D’Errico, S.; Di Paolo, M.; Frati, P.; Gaudio, R.M.; La Russa, R.; Maiese, A.; Marti, M.; Pinchi, E.; et al. Regulation of miRNAs as new tool for cutaneous vitality lesions demonstration in ligature marks in deaths by hanging. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Rudner, J.; Belka, C.; Marini, P.; Wagner, J.; Faltin, H.; Lepple-Wienhues, A.; Bamberg, M.; Budach, W. Radiation sensitivity and apoptosis in human lymphoma cells. Int. J. Radiat. Biol. 2001, 77, 1–11. [Google Scholar] [CrossRef]
- Turesson, I.; Simonsson, M.; Hermansson, I.; Book, M.; Sigurdadottir, S.; Thunberg, U.; Qvarnström, F.; Johansson, K.-A.; Fessé, P.; Nyman, J. Epidermal Keratinocyte Depletion during Five Weeks of Radiotherapy is Associated with DNA Double-Strand Break Foci, Cell Growth Arrest and Apoptosis: Evidence of Increasing Radioresponsiveness and Lack of Repopulation; The Number of Melanocytes Remains Un. Radiat. Res. 2020, 193, 481–496. [Google Scholar] [CrossRef]
- Rottensteiner-Brandl, U.; Bertram, U.; Lingens, L.F.; Köhn, K.; Distel, L.; Fey, T.; Körner, C.; Horch, R.E.; Arkudas, A. Free transplantation of a tissue engineered bone graft into an irradiated, critical-size femoral defect in rats. Cells 2021, 10, 2256. [Google Scholar] [CrossRef]
- Brown, K.R.; Rzucidlo, E. Acute and chronic radiation injury. J. Vasc. Surg. 2011, 53, 15S–21S. [Google Scholar] [CrossRef] [PubMed]
- Orth, M.; Lauber, K.; Niyazi, M.; Friedl, A.; Li, M.; Maihöfer, C.; Schüttrumpf, L.; Ernst, A.; Niemöller, O.M.; Belka, C. Current concepts in clinical radiation oncology. Radiat. Environ. Biophys. 2014, 53, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlik, T.M.; Keyomarsi, K. Role of cell cycle in mediating sensitivity to radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2004, 59, 928–942. [Google Scholar] [CrossRef] [PubMed]
- Soare, A.; Györfi, H.A.; Matei, A.E.; Dees, C.; Rauber, S.; Wohlfahrt, T.; Chen, C.-W.; Ludolph, I.; Horch, R.E.; Baeuerle, T.; et al. Dipeptidylpeptidase 4 as a Marker of Activated Fibroblasts and a Potential Target for the Treatment of Fibrosis in Systemic Sclerosis. Arthritis Rheumatol. 2020, 72, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Spałek, M. Chronic radiation-induced dermatitis: Challenges and solutions. Clin. Cosmet. Investig. Dermatol. 2016, 9, 473–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, M.; Alavi, A.; Wong, R.; Akita, S. Radiodermatitis: A Review of Our Current Understanding. Am. J. Clin. Dermatol. 2016, 17, 277–292. [Google Scholar] [CrossRef] [PubMed]
- Hymes, S.R.; Strom, E.A.; Fife, C. Radiation dermatitis: Clinical presentation, pathophysiology, and treatment 2006. J. Am. Acad. Dermatol. 2006, 54, 28–46. [Google Scholar] [CrossRef]
- Chan, R.J.; Webster, J.; Chung, B.; Marquart, L.; Ahmed, M.; Garantziotis, S. Prevention and treatment of acute radiation-induced skin reactions: A systematic review and meta-analysis of randomized controlled trials. BMC Cancer 2014, 14, 53. [Google Scholar] [CrossRef] [Green Version]
- Pignol, J.-P.; Olivotto, I.; Rakovitch, E.; Gardner, S.; Sixel, K.; Beckham, W.; Vu, T.T.T.; Truong, P.; Ackerman, I.; Paszat, L. A multicenter randomized trial of breast intensity-modulated radiation therapy to reduce acute radiation dermatitis. J. Clin. Oncol. 2008, 26, 2085–2092. [Google Scholar] [CrossRef] [Green Version]
- Schmuth, M.; Wimmer, M.; Hofer, S.; Sztankay, A.; Weinlich, G.; Linder, D.; Elias, P.; Fritsch, P.; Fritsch, E. Topical corticosteroid therapy for acute radiation dermatitis: A prospective, randomized, double-blind study. Br. J. Dermatol. 2002, 146, 983–991. [Google Scholar] [CrossRef]
- Niazi, T.M.; Vuong, T.; Azoulay, L.; Marijnen, C.; Bujko, K.; Nasr, E.; Lambert, C.; Duclos, M.; Faria, S.; David, M.; et al. Silver clear nylon dressing is effective in preventing radiation-induced dermatitis in patients with lower gastrointestinal cancer: Results from a phase III study. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, e305–e310. [Google Scholar] [CrossRef] [PubMed]
- Isoir, M.; Roque, T.; Squiban, C.; Milliat, F.; Mondon, P.; Mas-Chamberlin, C.; Benderitter, M.; Guipaud, O.; Tamarat, R. Protective effect of geranylgeranylacetone against radiation-induced delayed effects on human keratinocytes. Radiat. Res. 2013, 179, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Salvo, N.; Barnes, E.; van Draanen, J.; Stacey, E.; Mitera, G.; Breen, D.; Giotis, A.; Czarnota, G.; Pang, J.; De Angelis, C. Prophylaxis and management of acute radiation-induced skin reactions: A systematic review of the literature. Curr. Oncol. 2010, 17, 94–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mak, S.; Zee, C.; Molassiotis, A.; Chan, S.; Leung, S.; Mo, K.; Johnson, P. A comparison of wound treatments in nasopharyngeal cancer patients receiving radiation therapy. Cancer Nurs. 2006, 28, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Hickok, J.T.; Morrow, G.R.; Roscoe, J.A.; Mustian, K.; Okunieff, P. Occurrence, severity, and longitudinal course of twelve common symptoms in 1129 consecutive patients during radiotherapy for cancer. J. Pain. Symptom Manag. 2005, 30, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Turesson, I.; Bernefors, R.; Book, M.; Flogegård, M.; Hermansson, I.; Johansson, K.A.; Lindh, A.; Sigurdardottir, S.; Thunberg, U.; Nyman, J. Normal tissue response to low doses of radiotherapy assessed by molecular markers: A study of skin in patients treated for prostate cancer. Acta Oncol. 2001, 40, 941–951. [Google Scholar]
- Bernier, J.; Bonner, J.; Vermorken, J.B.; Bensadoun, R.J.; Dummer, R.; Giralt, J.; Kornek, G.; Hartley, A.; Mesia, R.; Robert, C.; et al. Consensus guidelines for the management of radiation dermatitis and coexisting acne-like rash in patients receiving radiotherapy plus EGFR inhibitors for the treatment of squamous cell carcinoma of the head and neck. Ann. Oncol. 2008, 19, 142–149. [Google Scholar] [CrossRef]
- Sadagurski, M.; Yakar, S.; Weingarten, G.; Holzenberger, M.; Rhodes, C.J.; Breitkreutz, D.; LeRoith, D.; Wertheimer, E. Insulin-Like Growth Factor 1 Receptor Signaling Regulates Skin Development and Inhibits Skin Keratinocyte Differentiation. Mol. Cell. Biol. 2006, 26, 2675–2687. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, C.; Hurwitz, S.A.; Kumar, M.G.; Cotton, J.; Spandau, D.F. Activation of the insulin-like growth factor-1 receptor promotes the survival of human keratinocytes following ultraviolet B irradiation. Int. J. Cancer 1999, 80, 431–438. [Google Scholar] [CrossRef]
- Hašová, M.; Crhák, T.; Šafránková, B.; Dvořáková, J.; Muthný, T.; Velebný, V.; Kubala, L. Hyaluronan minimizes effects of UV irradiation on human keratinocytes. Arch. Dermatol. Res. 2011, 303, 277–284. [Google Scholar] [CrossRef]
- Emmerson, E.; Campbell, L.; Davies, F.C.J.; Ross, N.L.; Ashcroft, G.S.; Krust, A.; Matthew, P.C.; Hardman, J. Insulin-like growth factor-1 promotes wound healing in estrogen-deprived mice: New insights into cutaneous IGF-1R/ERα cross talk. J. Investig. Dermatol. 2012, 132, 2838–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-Q.; Lee, S.; Wilson, H.; Seeger, M.; Iordanov, H.; Gatla, N.; Whittington, A.; Bach, D.; Lu, J.-Y.; Paller, A.S. Ganglioside GM3 depletion reverses impaired wound healing in diabetic mice by activating IGF-1 and insulin receptors. J. Investig. Dermatol. 2014, 134, 1446–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Hou, J.; Yuan, Q.; Xin, P.; Cheng, H.; Gu, Z.; Wu, J. Arginine derivatives assist dopamine-hyaluronic acid hybrid hydrogels to have enhanced antioxidant activity for wound healing. Chem. Eng. J. 2020, 392, 123775. [Google Scholar] [CrossRef]
- Yang, W.; Xu, H.; Lan, Y.; Zhu, Q.; Liu, Y.; Huang, S.; Shi, S.; Hancharou, A.; Tang, B.; Guo, R. Preparation and characterisation of a novel silk fibroin/hyaluronic acid/sodium alginate scaffold for skin repair. Int. J. Biol. Macromol. 2019, 130, 58–67. [Google Scholar] [CrossRef]
- Khurana, A.; Banothu, A.K.; Thanusha, A.V.; Nayal, A.; Dinda, A.K.; Singhal, M.; Bharani, K.K.; Koul, V. Preclinical efficacy study of a porous biopolymeric scaffold based on gelatin-hyaluronic acid-chondroitin sulfate in a porcine burn injury model: Role of critical molecular markers, (.V.E.G.F.A.; N-cadherin, C.O.X.-2.).; gamma sterilization efficacy a comparison of healing potential to, I.n.t.e.g.r.a.T.M. Biomed. Mater. 2021, 16, 055020. Available online: https://iopscience.iop.org/article/10.1088/1748-605X/ac1d3e (accessed on 8 January 2021).
- D’agostino, A.; Stellavato, A.; Busico, T.; Papa, A.; Tirino, V.; Papaccio, G.; La Gatta, A.; De Rosa, M.; Schiraldi, C. In vitro analysis of the effects on wound healing of high- and low-molecular weight chains of hyaluronan and their hybrid H-HA/L-HA complexes. BMC Cell Biol. 2015, 16, 19. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ngo, H.T.; Hwang, E.; Wei, X.; Liu, Y.; Liu, J.; Yi, T.-H. Conditioned medium from human adipose-derived mesenchymal stem cell culture prevents uvb-induced skin aging in human keratinocytes and dermal fibroblasts. Int. J. Mol. Sci. 2020, 21, 49. [Google Scholar] [CrossRef] [Green Version]
- Ong, H.T.; Redmond, S.L.; Marano, R.J.; Atlas, M.D.; Von Unge, M.; Aabel, P.; Dilley, R.J. Paracrine Activity from Adipose-Derived Stem Cells on in Vitro Wound Healing in Human Tympanic Membrane Keratinocytes. Stem Cells Dev. 2017, 26, 405–418. [Google Scholar] [CrossRef]
- Varghese, J.; Griffin, M.; Mosahebi, A.; Butler, P. Systematic review of patient factors affecting adipose stem cell viability and function: Implications for regenerative therapy. Stem Cell Res. Ther. 2017, 8, 45. [Google Scholar] [CrossRef] [Green Version]
- Kengelbach-Weigand, A.; Thielen, C.; Bäuerle, T.; Götzl, R.; Gerber, T.; Körner, C.; Beier, J.P.; Horch, R.E.; Boos, A.M. Personalized medicine for reconstruction of critical-size bone defects—A translational approach with customizable vascularized bone tissue. Npj Regen. Med. 2021, 6, 1–13. [Google Scholar] [CrossRef]
- Steiner, D.; Winkler, S.; Heltmann-Meyer, S.; Trossmann, V.T.; Fey, T.; Scheibel, T.; Horch, R.E.; Arkudas, A. Enhanced vascularization and de novo tissue formation in hydrogels made of engineered RGD-tagged spider silk proteins in the arteriovenous loop model. Biofabrication 2021, 13, 045003. [Google Scholar] [CrossRef] [PubMed]
- Steiner, D.; Mutschall, H.; Winkler, S.; Horch, R.E.; Arkudas, A. The adipose-derived stem cell and endothelial cell coculture system—Role of growth factors? Cells 2021, 10, 2074. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.E.; Patel, N.A.; Carter, G.; Moor, A.; Patel, R.; Ghansah, T.; Mathur, A.; Murr, M.M.; Bickford, P.; Gould, L.J.; et al. Comparison of Markers and Functional Attributes of Human Adipose-Derived Stem Cells and Dedifferentiated Adipocyte Cells from Subcutaneous Fat of an Obese Diabetic Donor. Adv. Wound Care 2014, 3, 219–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michurina, S.; Stafeev, I.; Podkuychenko, N.; Sklyanik, I.; Shestakova, E.; Yah’Yaev, K.; Yurasov, A.; Ratner, E.; Menshikov, M.; Parfyonova, Y.; et al. Decreased UCP-1 expression in beige adipocytes from adipose-derived stem cells of type 2 diabetes patients associates with mitochondrial ROS accumulation during obesity. Diabetes Res. Clin. Pract. 2020, 169, 169. [Google Scholar] [CrossRef]
- Bähr, C.; Groner, B. The IGF-1 receptor and its contributions to metastatic tumor growth—Novel approaches to the inhibition of IGF-1R function. Growth Factors 2005, 23, 1–14. Available online: https://www.tandfonline.com/doi/abs/10.1080/08977190400020229 (accessed on 8 January 2020). [CrossRef] [PubMed]
- Economou, M.A.; All-Ericsson, C.; Bykov, V.; Girnita, L.; Bartolazzi, A.; Larsson, O.; Seregard, S. Receptors for the liver synthesized growth factors IGF-1 and HGF/SF in uveal melanoma: Intercorrelation and prognostic implications. Investig. Opthalmol. Vis. Sci. 2005, 46, 4372–4375. [Google Scholar] [CrossRef]
- Matsui, Y.; Inomata, M.; Izumi, K.; Sonoda, K.; Shiraishi, N.; Kitano, S. Hyaluronic acid stimulates tumor-cell proliferation at wound sites. Gastrointest. Endosc. 2004, 60, 539–543. [Google Scholar] [CrossRef]
- Resnicoff, M.; Coppola, D.; Sell, C.; Rubin, R.; Ferrone, S.; Baserga, R. Growth Inhibition of Human Melanoma Cells in Nude Mice by Antisense Strategies to the Type 1 Insulin-like Growth Factor Receptor. Cancer Res. 1994, 54, 4848–4850. [Google Scholar]
- Baserga, R.; Peruzzi, F.; Reiss, K. The IGF-1 receptor in cancer biology. Int. J. Cancer 2003, 107, 873–877. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/ijc.11487 (accessed on 8 January 2020). [CrossRef]
- Coppola, D.; Baserga, R.; Rubini, M.; Ambrose, D.; Rubin, R. Rat Glioblastoma Cells Expressing an Antisense RNA to the Insulin-like Growth Factor-1 (IGF-1) Receptor Are Nontumorigenic and Induce Regression of Wild-Type Tumors. Cancer Res. 1994, 54, 2218–2222. [Google Scholar]
- Reiss, K.; D’Ambrosio, C.; Tu, X.; Tu, C.; Baserga, R. Inhibition of tumor growth by a dominant negative mutant of the insulin- like growth factor I receptor with a bystander effect. Clin. Cancer Res. 1998, 4, 2647–2655. [Google Scholar] [PubMed]
- Haas, A.F.; Isseroff, R.R.; Wheeland, R.G.; Rood, P.A.; Graves, P.J. Low-energy helium-neon laser irradiation increases the motility of cultured human keratinocytes. J. Investig. Dermatol. 1990, 94, 822–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, D.H.; Abrahamse, H. The role of laser fluence in cell viability, proliferation, and membrane integrity of wounded human skin fibroblasts following Helium-Neon laser irradiation. Lasers Surg. Med. 2006, 38, 74–83. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/lsm.20271 (accessed on 6 January 2021). [CrossRef] [PubMed]
- Mvula, B.; Mathope, T.; Moore, T.; Abrahamse, H. The effect of low level laser irradiation on adult human adipose derived stem cells. Lasers Med. Sci. 2008, 23, 277–282. Available online: https://link.springer.com/article/10.1007/s10103-007-0479-1 (accessed on 6 January 2021). [CrossRef]
- Goetze, K.; Scholz, M.; Taucher-Scholz, G.; Mueller-Klieser, W. The impact of conventional and heavy ion irradiation on tumor cell migration in vitro. Int. J. Radiat. Biol. 2007, 83, 889–896. Available online: https://www.tandfonline.com/doi/abs/10.1080/09553000701753826 (accessed on 6 January 2021). [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Supplement | Concentration |
|---|---|---|
| 1 (Control) | ||
| 2 | H-L-HA (50/50) | 1 mg/mL |
| 3 | H-L-HA (50/50) | 0.15 mg/mL |
| 4 | L-HA | 1 mg/mL |
| 5 | L-HA | 0.15 mg/mL |
| 6 | H-HA | 1 mg/mL |
| 7 | H-HA | 0.15 mg/mL |
| 8 | IGF-I | 100 ng/mL |
| 9 | KGF | 100 ng/mL |
| 10 | PL | 50 µL/mL |
| 11 | ADSC Control CM | 3-fold concentrated |
| 12 | ADSC CM | 3-fold concentrated |
| Gene | 5′-3′ Primer Sequence |
|---|---|
| GAPDH | For: ACATCAAGAAGGTGGTGAAGCAGG Rev: ACAAAGTGGTCGTTGAGGGCAA |
| P53 | For: GAAAACCTACCAGGGCAGCT Rev: GGGAGTACGTGCAAGTCACA |
| TNF | For: TGGGATCATTGCCCTGTGAG Rev: GGTGTCTGAAGGAGGGGGTA |
| TGFb1 | For: CATGGAGGACCTGGATGCC Rev: TCCTGAAGACTCCCCAGACC |
| IL6 | For: AAAGAGGCACTGGCAGAAAA Rev: TTTCACCAGGCAAGTCTCCT |
| IL8 | For: GTTCCACTGTGCCTTGGTTT Rev: GCTTCCACATGTCCTCACAA |
| IL1B | For: GCTCGCCAGTGAAATGATGG Rev: GGTGGTCGGAGATTCGTAGC |
| COL1A1 | For: GCTCTTGCAACATCTCCCCT Rev: CCTTCCTGACTCTCCTCCGA |
| MKI67 | For: TCGACCCTACAGAGTGCTCA Rev: GTGGGGAGCAGAGGTTCTTC |
| PTGS-2/COX-2 | For: ATGGAAACAGAGAAGTTGGCAG Rev: GATACAGCTCCACAGCATCG |
| FN1 | For: GAGAAGTATGTGCATGGTGTCAG Rev: AATACTTCGACAGGACCACTTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sörgel, C.A.; Schmid, R.; Stadelmann, N.; Weisbach, V.; Distel, L.; Horch, R.E.; Kengelbach-Weigand, A. IGF-I and Hyaluronic Acid Mitigate the Negative Effect of Irradiation on Human Skin Keratinocytes. Cancers 2022, 14, 588. https://doi.org/10.3390/cancers14030588
Sörgel CA, Schmid R, Stadelmann N, Weisbach V, Distel L, Horch RE, Kengelbach-Weigand A. IGF-I and Hyaluronic Acid Mitigate the Negative Effect of Irradiation on Human Skin Keratinocytes. Cancers. 2022; 14(3):588. https://doi.org/10.3390/cancers14030588
Chicago/Turabian StyleSörgel, Celena A., Rafael Schmid, Nina Stadelmann, Volker Weisbach, Luitpold Distel, Raymund E. Horch, and Annika Kengelbach-Weigand. 2022. "IGF-I and Hyaluronic Acid Mitigate the Negative Effect of Irradiation on Human Skin Keratinocytes" Cancers 14, no. 3: 588. https://doi.org/10.3390/cancers14030588
APA StyleSörgel, C. A., Schmid, R., Stadelmann, N., Weisbach, V., Distel, L., Horch, R. E., & Kengelbach-Weigand, A. (2022). IGF-I and Hyaluronic Acid Mitigate the Negative Effect of Irradiation on Human Skin Keratinocytes. Cancers, 14(3), 588. https://doi.org/10.3390/cancers14030588