
Advances in Dietary Phenolic Compounds to Improve Chemosensitivity of Anticancer Drugs

 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
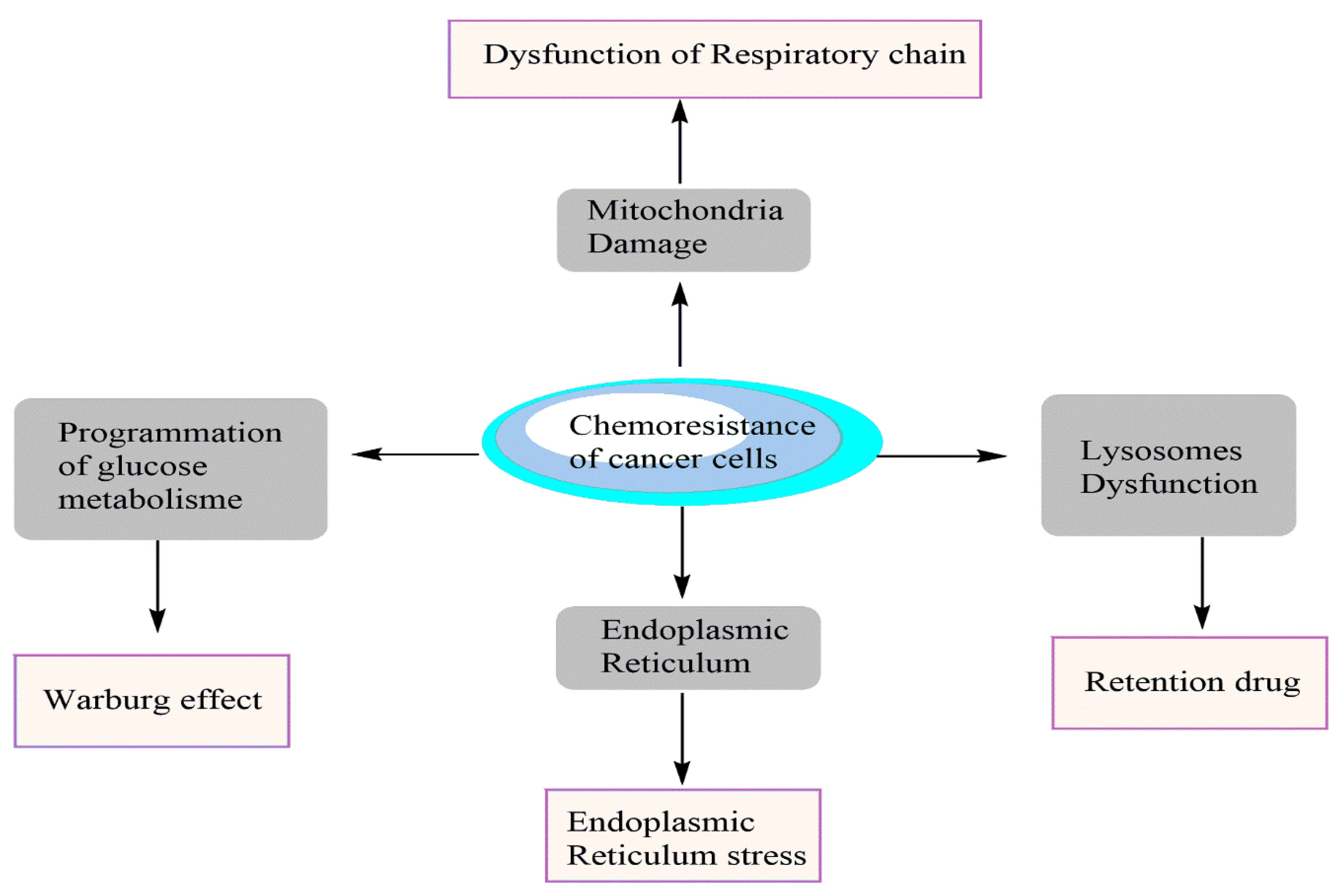
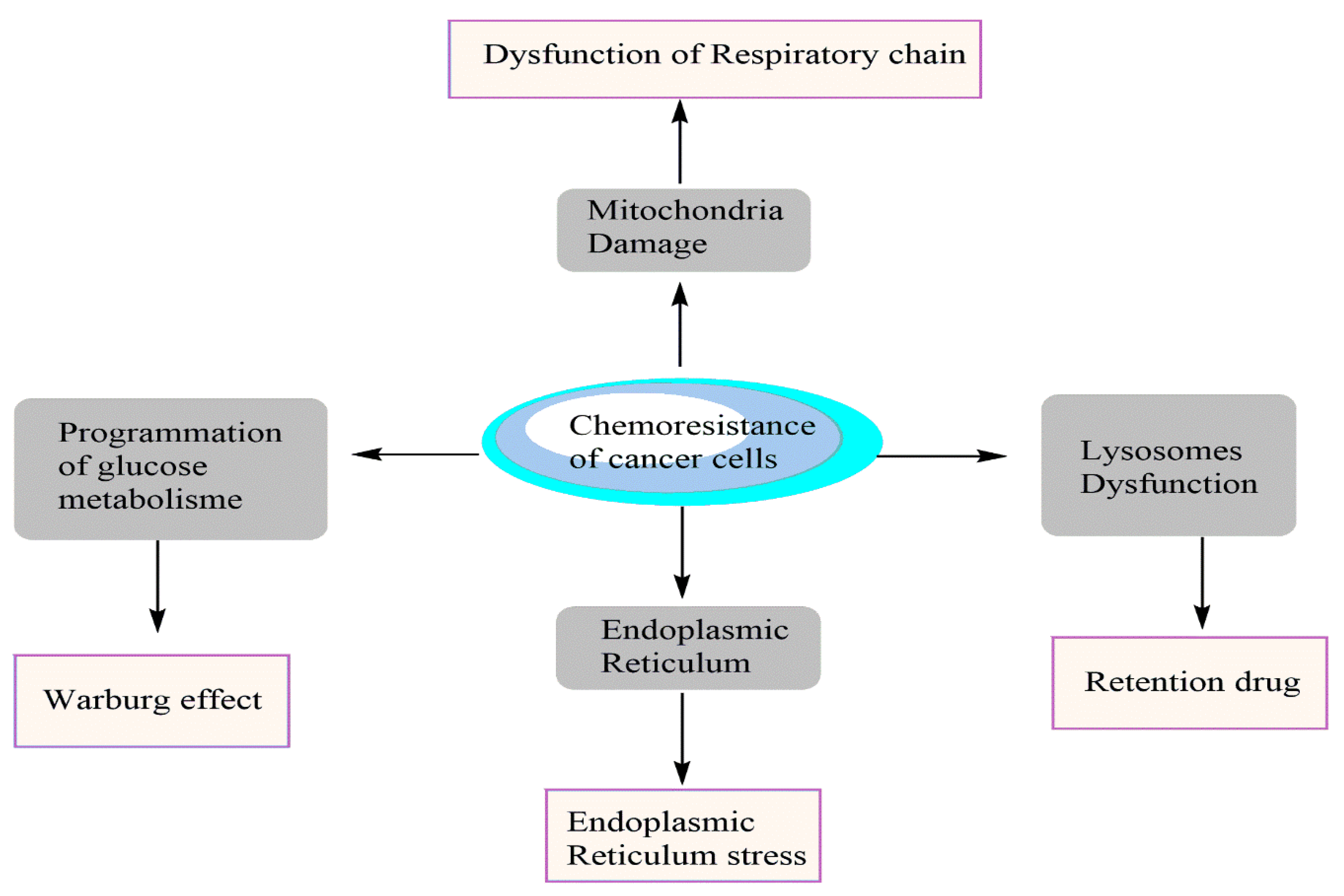
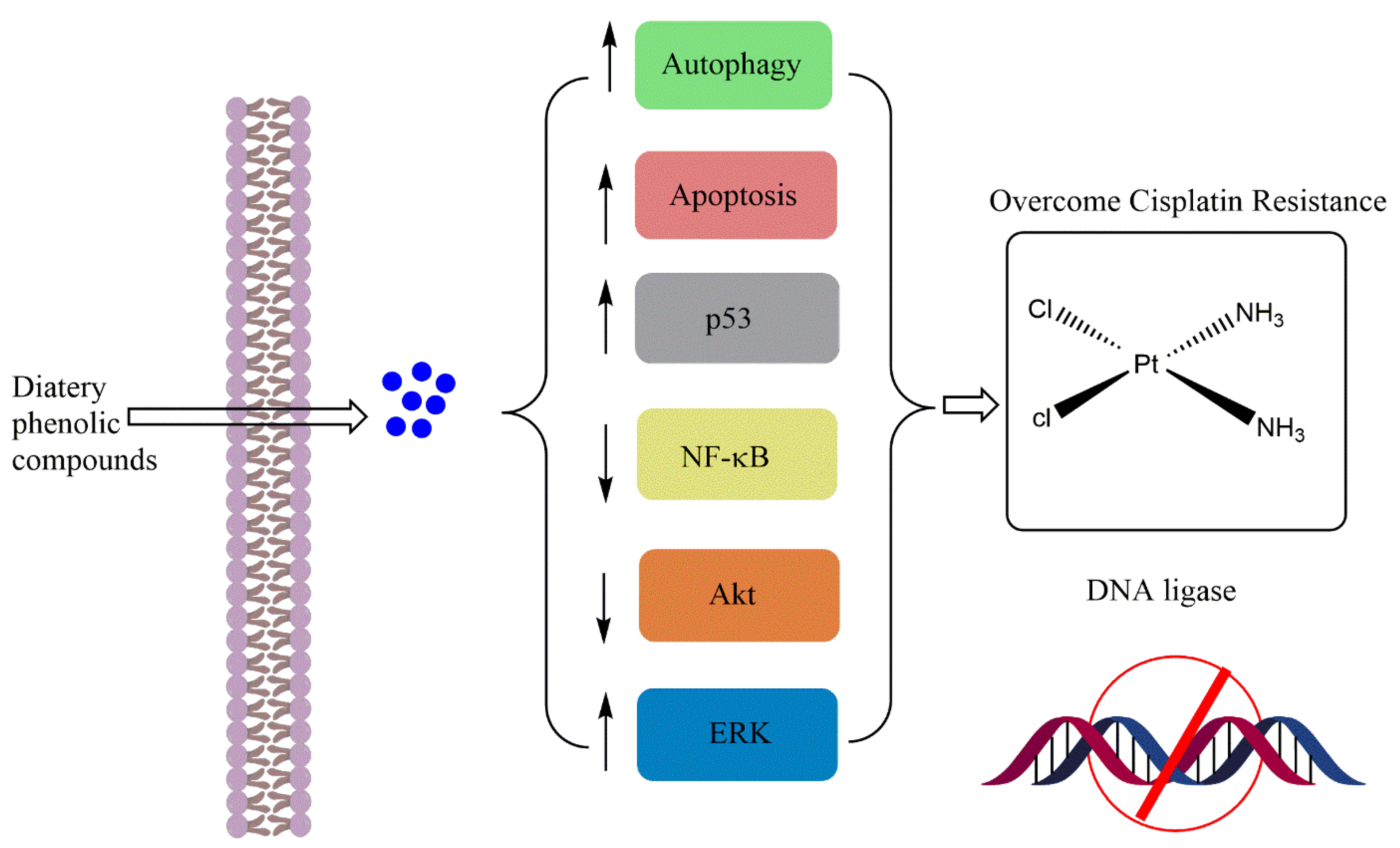
1. Introduction
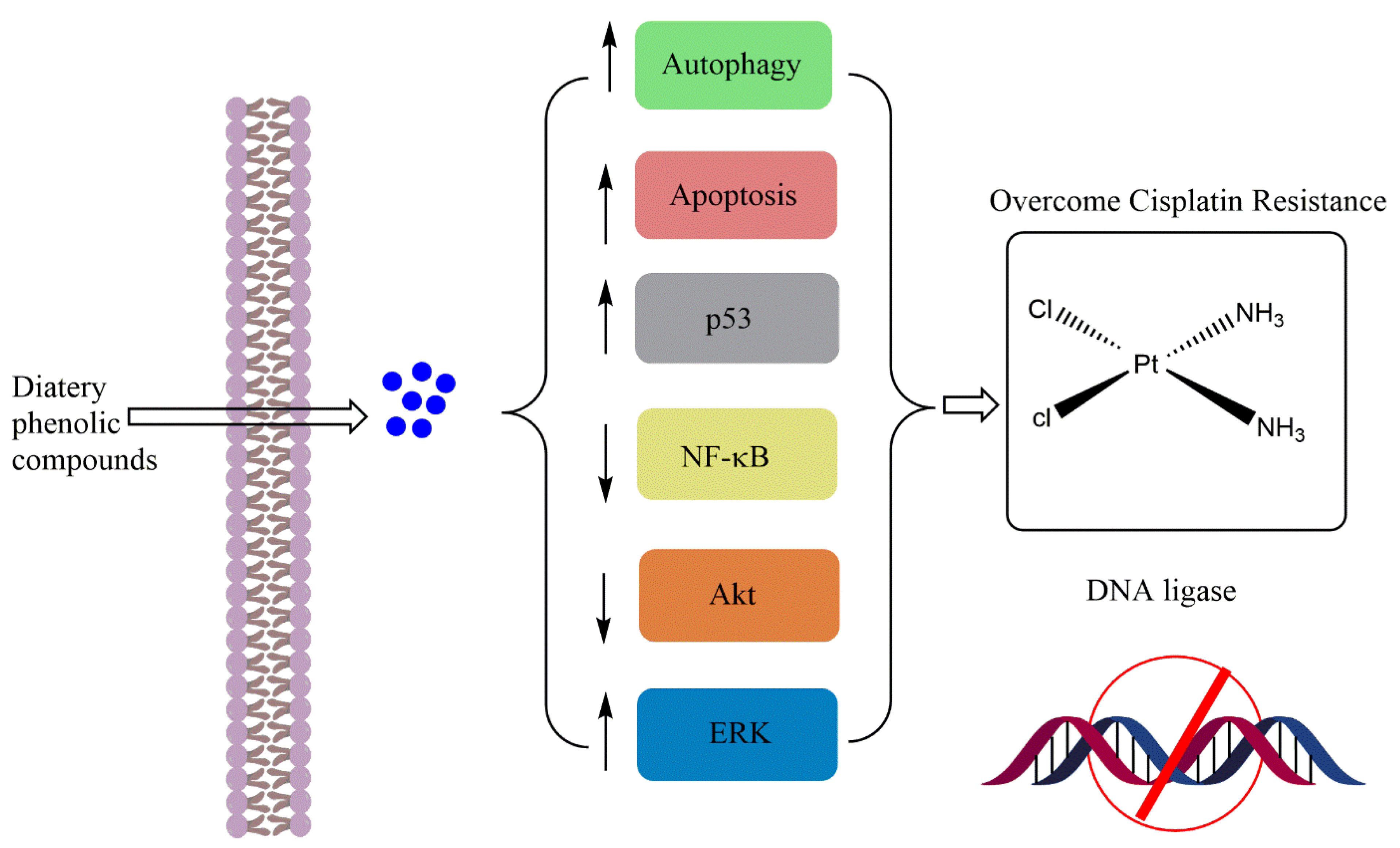
2. Dietary Phenolic Compounds Improving the Chemosensitivity of Anticancer Drugs
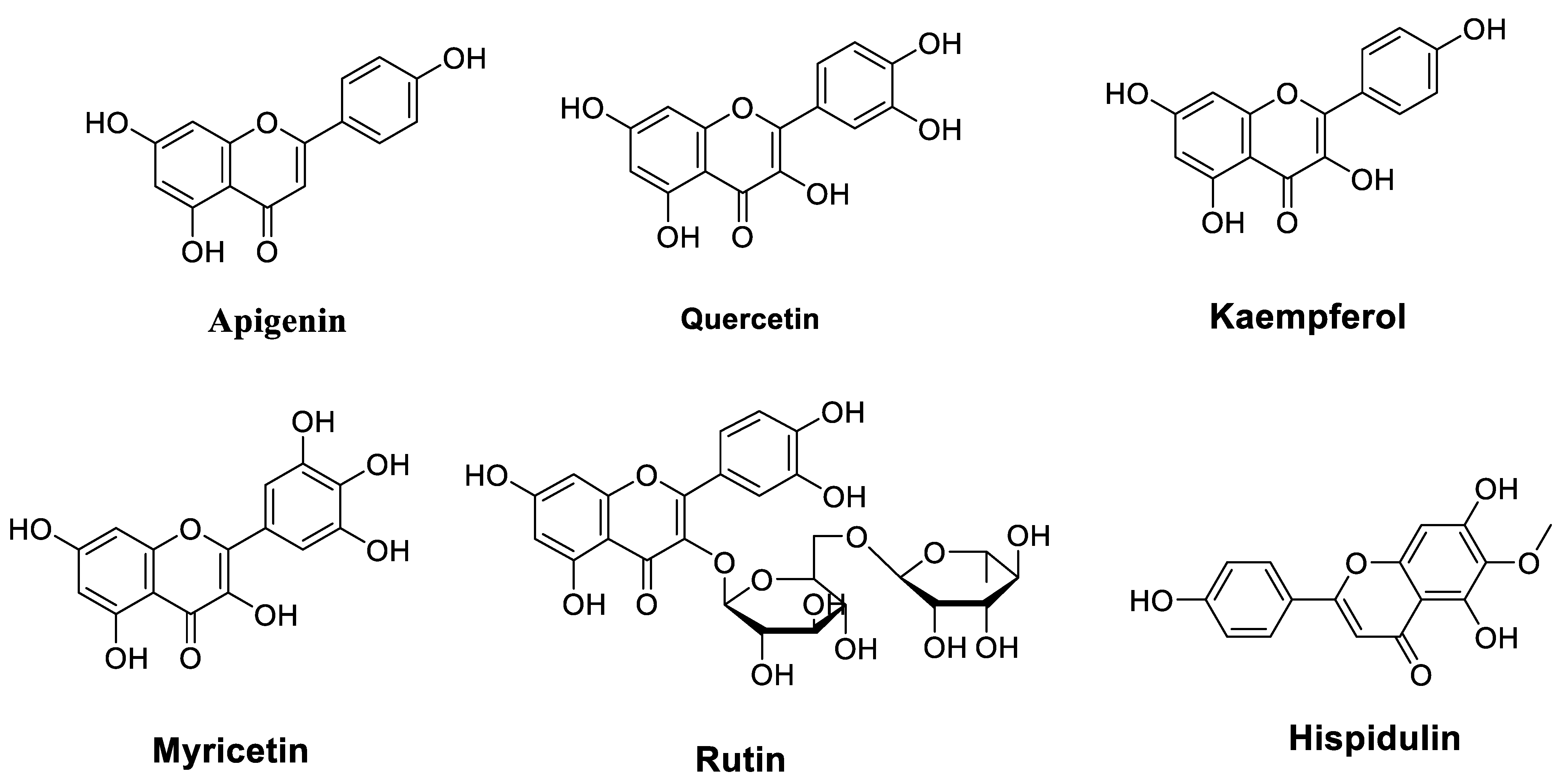
2.1. Flavonoids
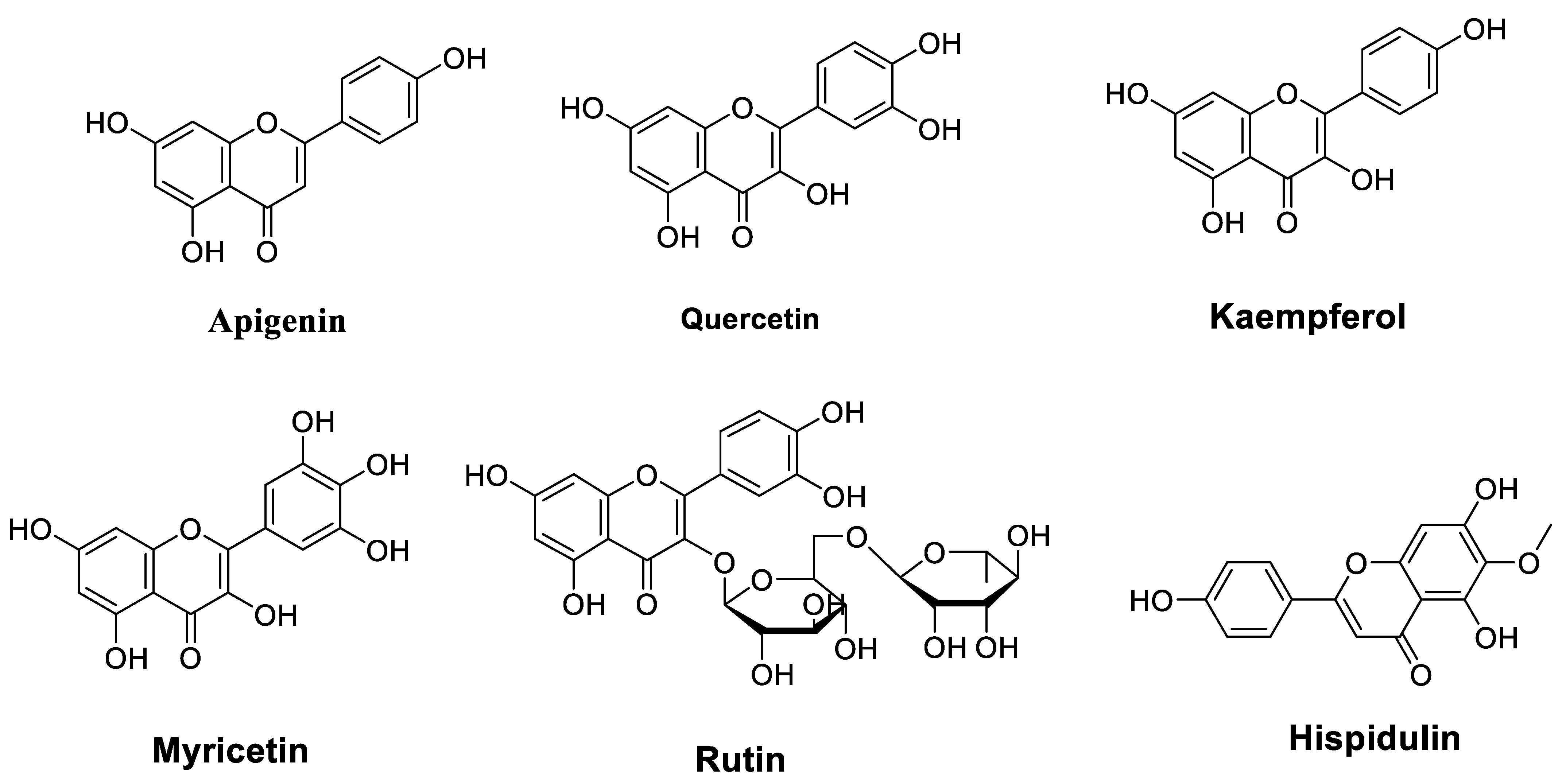
2.1.1. Flavones
2.1.2. Flavanols
2.1.3. Flavonols
2.1.4. Anthocyanidins
2.2. Non-Flavonoids
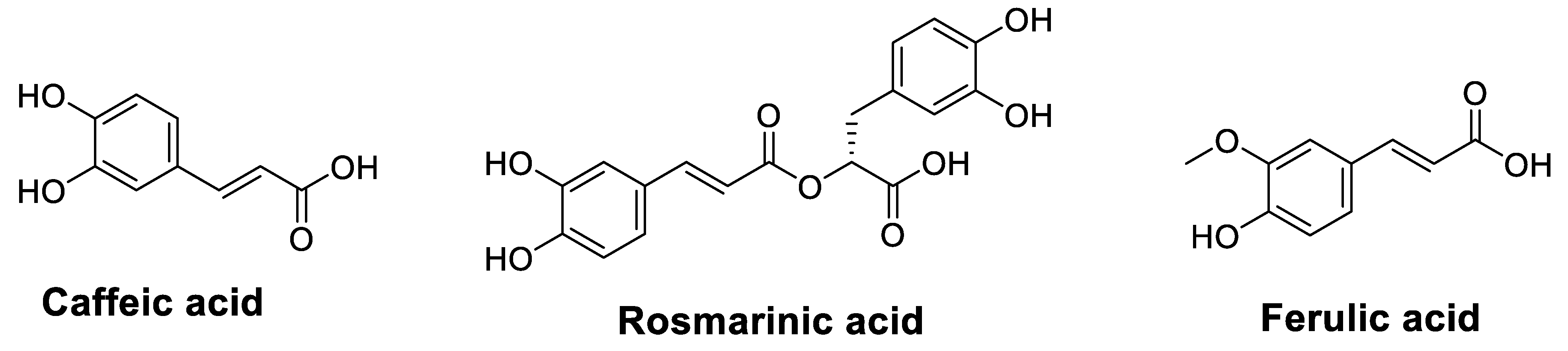
2.2.1. Phenolic Acids
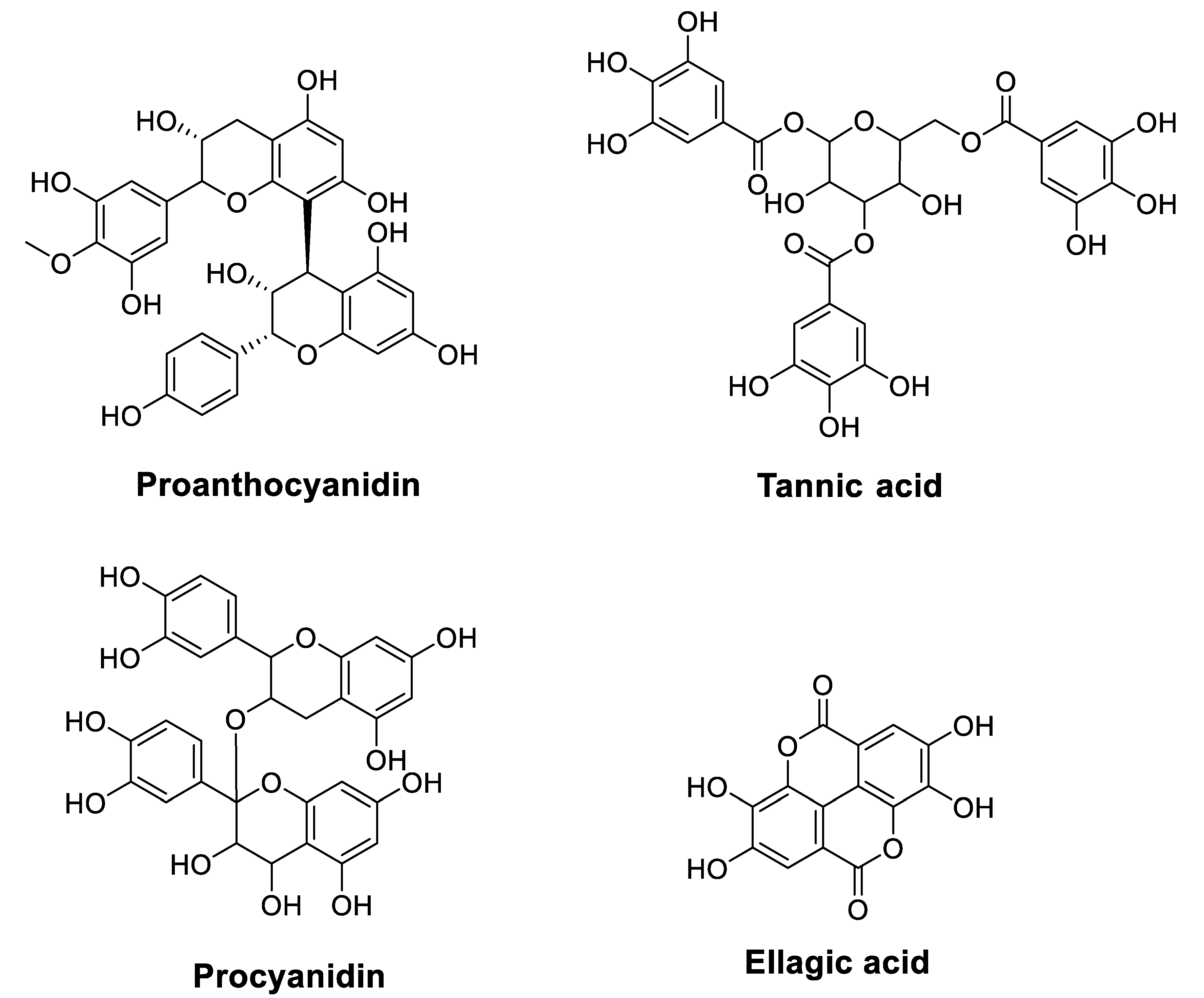
2.2.2. Tannins
3. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACNs | Anthocyanidins |
| Akt | Protein Kinase B |
| APG | Apigenin |
| CAPE | Caffeic Acid Phenethyl Ester |
| CLL | Chronic Lymphocytic Leukemia |
| COX-2 | Cyclooxygenase-2 |
| CRC | Colorectal Carcinoma |
| DM | Daunomycin |
| DOX | Doxorubicin |
| ELISA | Enzyme-linked immunosorbent assay |
| EMT | Epithelial-Mesenchymal Transition |
| ERK | Extracellular Signal-Regulated Kinase |
| FA | Ferulic acid |
| GLUT-1 | Glucose Transporter-1 |
| Gem | Gemcitabine |
| GR | Gamma Radiation |
| HCC | Hepatocellular Carcinoma |
| HIF-1α | Hypoxia-Inducible Factor-1α |
| HO | Heme Oxygenase |
| HPD | Hispidulin |
| HPDE | Human pancreatic ductal epithelium |
| IL | Interleukin |
| IR | Ionizing Radiation |
| JNK | C-Jun N-Terminal Kinase |
| KAE | Kaempferol |
| MAPK | Mitogen-Activated Protein Kinase |
| MDR1 | Multidrug resistance protein 1 |
| MM | Multiple Myeloma |
| mTOR | mammalian Target of Rapamycin |
| MYR | Myricetin |
| NF-ĸB | Nuclear Factor Kappa B |
| Nrf2 | Nuclear factor erythroid-related factor 2 |
| NSCLC | Non-Small Cell Lung Cancer |
| OC | Ovarian Cancer |
| OXP | Oxaliplatin |
| P-gp | P-glycoprotein |
| PTEN | Phosphatase and Tensin Homolog |
| PTX | Paclitaxel |
| Que | Quercetin |
| ROS | Reactive Oxygen Species |
| RTN | Rutin |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| TA | Tannic acid |
| TNF-α | Tumor Necrosis Factor-α |
| TMZ | Temozolomide |
| VBL | Vinblastine |
| VCR | Vincristine |
| 5-FU | 5-Fluorouracil |
References
- Hurson, A.N.; Ahearn, T.U.; Keeman, R.; Abubakar, M.; Jung, A.Y.; Kapoor, P.M.; Koka, H.; Yang, X.R.; Chang-Claude, J.; Martínez, E. Systematic Literature Review of Risk Factor Associations with Breast Cancer Subtypes in Women of African, Asian, Hispanic, and European Descents. Cancer Res. 2022, 82, 3670. [Google Scholar] [CrossRef]
- Stopsack, K.H.; Nandakumar, S.; Arora, K.; Nguyen, B.; Vasselman, S.E.; Nweji, B.; McBride, S.M.; Morris, M.J.; Rathkopf, D.E.; Slovin, S.F. Differences in Prostate Cancer Genomes by Self-Reported Race: Contributions of Genetic Ancestry, Modifiable Cancer Risk Factors, and Clinical FactorsRacial Differences in Prostate Cancer Genomes. Clin. Cancer Res. 2022, 28, 318–326. [Google Scholar] [CrossRef]
- Tan, D.A.; Dayu, A.R.B. Menopausal Hormone Therapy: Why We Should No Longer Be Afraid of the Breast Cancer Risk. Climacteric 2022, 25, 362–368. [Google Scholar] [CrossRef]
- Dobroslavić, E.; Repajić, M.; Dragović-Uzelac, V.; Elez Garofulić, I. Isolation of Laurus Nobilis Leaf Polyphenols: A Review on Current Techniques and Future Perspectives. Foods 2022, 11, 235. [Google Scholar] [CrossRef]
- Mitra, S.; Tareq, A.M.; Das, R.; Emran, T.B.; Nainu, F.; Chakraborty, A.J.; Ahmad, I.; Tallei, T.E.; Idris, A.M.; Simal-Gandara, J. Polyphenols: A First Evidence in the Synergism and Bioactivities. Food Rev. Int. 2022, 1–23. [Google Scholar] [CrossRef]
- Rosero, S.; Del Pozo, F.; Simbaña, W.; Álvarez, M.; Quinteros, M.F.; Carrillo, W.; Morales, D. Polyphenols and Flavonoids Composition, Anti-Inflammatory and Antioxidant Properties of Andean Baccharis Macrantha Extracts. Plants 2022, 11, 1555. [Google Scholar] [CrossRef]
- Islam, B.U.; Suhail, M.; Khan, M.K.; Zughaibi, T.A.; Alserihi, R.F.; Zaidi, S.K.; Tabrez, S. Polyphenols as Anticancer Agents: Toxicological Concern to Healthy Cells. Phytother. Res. 2021, 35, 6063–6079. [Google Scholar] [CrossRef]
- Rauf, A.; Shariati, M.A.; Imran, M.; Bashir, K.; Khan, S.A.; Mitra, S.; Emran, T.B.; Badalova, K.; Uddin, M.; Mubarak, M.S. Comprehensive Review on Naringenin and Naringin Polyphenols as a Potent Anticancer Agent. Environ. Sci. Pollut. Res. 2022, 29, 31025–31041. [Google Scholar] [CrossRef]
- Mottaghi, S.; Abbaszadeh, H. Natural Lignans Honokiol and Magnolol as Potential Anticarcinogenic and Anticancer Agents. A Comprehensive Mechanistic Review. Nutr. Cancer 2022, 74, 761–778. [Google Scholar] [CrossRef]
- Yoganathan, S.; Alagaratnam, A.; Acharekar, N.; Kong, J. Ellagic Acid and Schisandrins: Natural Biaryl Polyphenols with Therapeutic Potential to Overcome Multidrug Resistance in Cancer. Cells 2021, 10, 458. [Google Scholar] [CrossRef]
- Chan, K.-F.; Zhao, Y.; Burkett, B.A.; Wong, I.L.; Chow, L.M.; Chan, T.H. Flavonoid Dimers as Bivalent Modulators for P-Glycoprotein-Based Multidrug Resistance: Synthetic Apigenin Homodimers Linked with Defined-Length Poly (Ethylene Glycol) Spacers Increase Drug Retention and Enhance Chemosensitivity in Resistant Cancer Cells. J. Med. Chem. 2006, 49, 6742–6759. [Google Scholar] [CrossRef]
- Gao, A.-M.; Ke, Z.-P.; Wang, J.-N.; Yang, J.-Y.; Chen, S.-Y.; Chen, H. Apigenin Sensitizes Doxorubicin-Resistant Hepatocellular Carcinoma BEL-7402/ADM Cells to Doxorubicin via Inhibiting PI3K/Akt/Nrf2 Pathway. Carcinogenesis 2013, 34, 1806–1814. [Google Scholar] [CrossRef]
- Johnson, J.L.; de Mejia, E.G. Interactions between Dietary Flavonoids Apigenin or Luteolin and Chemotherapeutic Drugs to Potentiate Anti-Proliferative Effect on Human Pancreatic Cancer Cells, in Vitro. Food Chem. Toxicol. 2013, 60, 83–91. [Google Scholar] [CrossRef]
- Wu, D.-G.; Yu, P.; Li, J.-W.; Jiang, P.; Sun, J.; Wang, H.-Z.; Zhang, L.-D.; Wen, M.-B.; Bie, P. Apigenin Potentiates the Growth Inhibitory Effects by IKK-β-Mediated NF-ΚB Activation in Pancreatic Cancer Cells. Toxicol. Lett. 2014, 224, 157–164. [Google Scholar] [CrossRef]
- Xu, Y.-Y.; Wu, T.-T.; Zhou, S.-H.; Bao, Y.-Y.; Wang, Q.-Y.; Fan, J.; Huang, Y.-P. Apigenin Suppresses GLUT-1 and p-AKT Expression to Enhance the Chemosensitivity to Cisplatin of Laryngeal Carcinoma Hep-2 Cells: An in Vitro Study. Int. J. Clin. Exp. Pathol. 2014, 7, 3938. [Google Scholar]
- Bao, Y.-Y.; Zhou, S.-H.; Lu, Z.-J.; Fan, J.; Huang, Y.-P. Inhibiting GLUT-1 Expression and PI3K/Akt Signaling Using Apigenin Improves the Radiosensitivity of Laryngeal Carcinoma in Vivo. Oncol. Rep. 2015, 34, 1805–1814. [Google Scholar] [CrossRef]
- Hu, X.-Y.; Liang, J.-Y.; Guo, X.-J.; Liu, L.; Guo, Y.-B. 5-Fluorouracil Combined with Apigenin Enhances Anticancer Activity through Mitochondrial Membrane Potential (ΔΨm)-Mediated Apoptosis in Hepatocellular Carcinoma. Clin. Exp. Pharmacol. Physiol. 2015, 42, 146–153. [Google Scholar] [CrossRef]
- Ju, S.M.; Kang, J.G.; Bae, J.S.; Pae, H.O.; Lyu, Y.S.; Jeon, B.H. The Flavonoid Apigenin Ameliorates Cisplatin-Induced Nephrotoxicity through Reduction of P53 Activation and Promotion of PI3K/Akt Pathway in Human Renal Proximal Tubular Epithelial Cells. Evid. -Based Complementary Altern. Med. 2015, 2015, 186436 . [Google Scholar] [CrossRef]
- Gaballah, H.H.; Gaber, R.A.; Mohamed, D.A. Apigenin Potentiates the Antitumor Activity of 5-FU on Solid Ehrlich Carcinoma: Crosstalk between Apoptotic and JNK-Mediated Autophagic Cell Death Platforms. Toxicol. Appl. Pharmacol. 2017, 316, 27–35. [Google Scholar] [CrossRef]
- Gao, A.-M.; Zhang, X.-Y.; Ke, Z.-P. Apigenin Sensitizes BEL-7402/ADM Cells to Doxorubicin through Inhibiting MiR-101/Nrf2 Pathway. Oncotarget 2017, 8, 82085. [Google Scholar] [CrossRef]
- Gao, A.-M.; Zhang, X.-Y.; Hu, J.-N.; Ke, Z.-P. Apigenin Sensitizes Hepatocellular Carcinoma Cells to Doxorubic through Regulating MiR-520b/ATG7 Axis. Chem.-Biol. Interact. 2018, 280, 45–50. [Google Scholar] [CrossRef]
- Li, Q.; Li, L.; Zhao, X.; Cheng, Z.; Ma, J. Apigenin Induces Apoptosis and Reverses the Drug Resistance of Ovarian Cancer Cells. Int. J. Clin. Exp. Med. 2020, 13, 1987–1994. [Google Scholar]
- Thangasamy, T.; Sittadjody, S.; Mitchell, G.C.; Mendoza, E.E.; Radhakrishnan, V.M.; Limesand, K.H.; Burd, R. Quercetin Abrogates Chemoresistance in Melanoma Cells by Modulating ΔNp73. BMC Cancer 2010, 10, 282. [Google Scholar] [CrossRef]
- Chuang-Xin, L.; Wen-Yu, W.; Yao, C.U.I.; Xiao-Yan, L.; Yun, Z. Quercetin Enhances the Effects of 5-Fluorouracil-Mediated Growth Inhibition and Apoptosis of Esophageal Cancer Cells by Inhibiting NF-ΚB. Oncol. Lett. 2012, 4, 775–778. [Google Scholar] [CrossRef]
- Li, S.; Li, K.; Zhang, J.; Dong, Z. The Effect of Quercetin on Doxorubicin Cytotoxicity in Human Breast Cancer Cells. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. -Anti-Cancer Agents) 2013, 13, 352–355. [Google Scholar] [CrossRef]
- Maciejczyk, A.; Surowiak, P. Quercetin Inhibits Proliferation and Increases Sensitivity of Ovarian Cancer Cells to Cisplatin and Paclitaxel. Ginekol. Pol. 2013, 84, 590–595. [Google Scholar] [CrossRef]
- Sang, D.; Li, R.; Lan, Q. Quercetin Sensitizes Human Glioblastoma Cells to Temozolomide in Vitro via Inhibition of Hsp27. Acta Pharmacol. Sin. 2014, 35, 832–838. [Google Scholar] [CrossRef]
- Li, S.; Qiao, S.; Zhang, J.; Li, K. Quercetin Increase the Chemosensitivity of Breast Cancer Cells to Doxorubicin via PTEN/Akt Pathway. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. -Anti-Cancer Agents) 2015, 15, 1185–1189. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, E.J.; Min, K.H.; Hur, G.Y.; Lee, S.H.; Lee, S.Y.; Kim, J.H.; Shin, C.; Shim, J.J.; In, K.H. Quercetin Enhances Chemosensitivity to Gemcitabine in Lung Cancer Cells by Inhibiting Heat Shock Protein 70 Expression. Clin. Lung Cancer 2015, 16, e235–e243. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, C.; Ma, T.; Jiang, L.; Tang, L.; Shi, T.; Zhang, S.; Zhang, L.; Zhu, P.; Li, J. Reversal Effect of Quercetin on Multidrug Resistance via FZD7/β-Catenin Pathway in Hepatocellular Carcinoma Cells. Phytomedicine 2018, 43, 37–45. [Google Scholar] [CrossRef]
- Shu, Y.; Xie, B.; Liang, Z.; Chen, J. Quercetin Reverses the Doxorubicin Resistance of Prostate Cancer Cells by Downregulating the Expression of C-Met. Oncol. Lett. 2018, 15, 2252–2258. [Google Scholar] [CrossRef]
- Lan, C.-Y.; Chen, S.-Y.; Kuo, C.-W.; Lu, C.-C.; Yen, G.-C. Quercetin Facilitates Cell Death and Chemosensitivity through RAGE/PI3K/AKT/MTOR Axis in Human Pancreatic Cancer Cells. J. Food Drug Anal. 2019, 27, 887–896. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, J.; Yu, C.; Xiang, L.; Li, L.; Shi, D.; Lin, F. Quercetin Enhanced Paclitaxel Therapeutic Effects towards PC-3 Prostate Cancer through ER Stress Induction and ROS Production. OncoTargets Ther. 2020, 13, 513. [Google Scholar] [CrossRef]
- Mawalizadeh, F.; Mohammadzadeh, G.; Khedri, A.; Rashidi, M. Quercetin Potentiates the Chemosensitivity of MCF-7 Breast Cancer Cells to 5-Fluorouracil. Mol. Biol. Rep. 2021, 48, 7733–7742. [Google Scholar] [CrossRef]
- Safi, A.; Heidarian, E.; Ahmadi, R. Quercetin Synergistically Enhances the Anticancer Efficacy of Docetaxel through Induction of Apoptosis and Modulation of PI3K/AKT, MAPK/ERK, and JAK/STAT3 Signaling Pathways in MDA-MB-231 Breast Cancer Cell Line. Int. J. Mol. Cell. Med. 2021, 10, 11. [Google Scholar]
- Yanqiu, H.; Linjuan, C.; Jin, W.; Hongjun, H.; Yongjin, S.; Guobin, X.; Hanyun, R. The Effects of Quercetin and Kaempferol on Multidrug Resistance and the Expression of Related Genes in Human Erythroleukemic K562/A Cells. Afr. J. Biotechnol. 2011, 10, 13399–13406. [Google Scholar] [CrossRef]
- Riahi-Chebbi, I.; Souid, S.; Othman, H.; Haoues, M.; Karoui, H.; Morel, A.; Srairi-Abid, N.; Essafi, M.; Essafi-Benkhadir, K. The Phenolic Compound Kaempferol Overcomes 5-Fluorouracil Resistance in Human Resistant LS174 Colon Cancer Cells. Sci. Rep. 2019, 9, 195. [Google Scholar] [CrossRef]
- Wu, H.; Du, J.; Li, C.; Li, H.; Guo, H.; Li, Z. Kaempferol Can Reverse the 5-Fu Resistance of Colorectal Cancer Cells by Inhibiting PKM2-Mediated Glycolysis. Int. J. Mol. Sci. 2022, 23, 3544. [Google Scholar] [CrossRef]
- Wang, L.; Feng, J.; Chen, X.; Guo, W.; Du, Y.; Wang, Y.; Zang, W.; Zhang, S.; Zhao, G. Myricetin Enhance Chemosensitivity of 5-Fluorouracil on Esophageal Carcinoma in Vitro and in Vivo. Cancer Cell Int. 2014, 14, 71. [Google Scholar] [CrossRef]
- Zheng, A.-W.; Chen, Y.-Q.; Zhao, L.-Q.; Feng, J.-G. Myricetin Induces Apoptosis and Enhances Chemosensitivity in Ovarian Cancer Cells. Oncol. Lett. 2017, 13, 4974–4978. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; Arnold Apostolides, N.; El Beyrouthy, M. Chemical Composition and Antiradical Capacity of Essential Oils from Lebanese Medicinal Plants. J. Essent. Oil Res. 2014, 26, 466–472. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, G.; Hu, J.; Zhu, Y.; Lan, H.; Shen, X.; Lv, Y.; Huang, L. Rutin Attenuates Sorafenib-Induced Chemoresistance and Autophagy in Hepatocellular Carcinoma by Regulating BANCR/MiRNA-590-5P/OLR1 Axis. Int. J. Biol. Sci. 2021, 17, 3595. [Google Scholar] [CrossRef]
- Gao, H.; Xie, J.; Peng, J.; Han, Y.; Jiang, Q.; Han, M.; Wang, C. Hispidulin Inhibits Proliferation and Enhances Chemosensitivity of Gallbladder Cancer Cells by Targeting HIF-1α. Exp. Cell Res. 2015, 332, 236–246. [Google Scholar] [CrossRef]
- Li, X.; Chen, L.; Gao, Y.; Zhang, Q.; Chang, A.K.; Yang, Z.; Bi, X. Black Raspberry Anthocyanins Increased the Antiproliferative Effects of 5-Fluorouracil and Celecoxib in Colorectal Cancer Cells and Mouse Model. J. Funct. Foods 2021, 87, 104801. [Google Scholar] [CrossRef]
- Kang, S.H.; Bak, D.-H.; Chung, B.Y.; Bai, H.-W.; Kang, B.S. Delphinidin Enhances Radio-Therapeutic Effects via Autophagy Induction and JNK/MAPK Pathway Activation in Non-Small Cell Lung Cancer. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2020, 24, 413–422. [Google Scholar] [CrossRef]
- Eguchi, H.; Matsunaga, H.; Onuma, S.; Yoshino, Y.; Matsunaga, T.; Ikari, A. Down-Regulation of Claudin-2 Expression by Cyanidin-3-Glucoside Enhances Sensitivity to Anticancer Drugs in the Spheroid of Human Lung Adenocarcinoma A549 Cells. Int. J. Mol. Sci. 2021, 22, 499. [Google Scholar] [CrossRef]
- Kao, T.-Y.; Chung, Y.-C.; Hou, Y.-C.; Tsai, Y.-W.; Chen, C.-H.; Chang, H.-P.; Chou, J.-L.; Hsu, C.-P. Effects of Ellagic Acid on Chemosensitivity to 5-Fluorouracil in Colorectal Carcinoma Cells. Anticancer Res. 2012, 32, 4413–4418. [Google Scholar]
- Engelke, L.H.; Hamacher, A.; Proksch, P.; Kassack, M.U. Ellagic Acid and Resveratrol Prevent the Development of Cisplatin Resistance in the Epithelial Ovarian Cancer Cell Line A2780. J. Cancer 2016, 7, 353. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Liao, H.-F.; Tsai, T.-H.; Wang, S.-Y.; Shiao, M.-S. Caffeic Acid Phenethyl Ester Preferentially Sensitizes CT26 Colorectal Adenocarcinoma to Ionizing Radiation without Affecting Bone Marrow Radioresponse. Int. J. Radiat. Oncol. Biol. Phys. 2005, 63, 1252–1261. [Google Scholar] [CrossRef]
- Lin, Y.-H.; Chiu, J.-H.; Tseng, W.-S.; Wong, T.-T.; Chiou, S.-H.; Yen, S.-H. Antiproliferation and Radiosensitization of Caffeic Acid Phenethyl Ester on Human Medulloblastoma Cells. Cancer Chemother. Pharmacol. 2006, 57, 525–532. [Google Scholar] [CrossRef]
- Matsunaga, T.; Tsuchimura, S.; Azuma, N.; Endo, S.; Ichihara, K.; Ikari, A. Caffeic Acid Phenethyl Ester Potentiates Gastric Cancer Cell Sensitivity to Doxorubicin and Cisplatin by Decreasing Proteasome Function. Anti-Cancer Drugs 2019, 30, 251–259. [Google Scholar] [CrossRef]
- Sonoki, H.; Tanimae, A.; Furuta, T.; Endo, S.; Matsunaga, T.; Ichihara, K.; Ikari, A. Caffeic Acid Phenethyl Ester Down-Regulates Claudin-2 Expression at the Transcriptional and Post-Translational Levels and Enhances Chemosensitivity to Doxorubicin in Lung Adenocarcinoma A549 Cells. J. Nutr. Biochem. 2018, 56, 205–214. [Google Scholar] [CrossRef]
- Anjaly, K.; Tiku, A.B. Caffeic Acid Phenethyl Ester Induces Radiosensitization via Inhibition of DNA Damage Repair in Androgen-Independent Prostate Cancer Cells. Environ. Toxicol. 2022, 37, 995–1006. [Google Scholar] [CrossRef]
- Muthusamy, G.; Balupillai, A.; Ramasamy, K.; Shanmugam, M.; Gunaseelan, S.; Mary, B.; Prasad, N.R. Ferulic Acid Reverses ABCB1-Mediated Paclitaxel Resistance in MDR Cell Lines. Eur. J. Pharmacol. 2016, 786, 194–203. [Google Scholar] [CrossRef]
- Muthusamy, G.; Gunaseelan, S.; Prasad, N.R. Ferulic Acid Reverses P-Glycoprotein-Mediated Multidrug Resistance via Inhibition of PI3K/Akt/NF-ΚB Signaling Pathway. J. Nutr. Biochem. 2019, 63, 62–71. [Google Scholar] [CrossRef]
- Heo, S.-K.; Noh, E.-K.; Yoon, D.-J.; Jo, J.-C.; Koh, S.; Baek, J.H.; Park, J.-H.; Min, Y.J.; Kim, H. Rosmarinic Acid Potentiates ATRA-Induced Macrophage Differentiation in Acute Promyelocytic Leukemia NB4 Cells. Eur. J. Pharmacol. 2015, 747, 36–44. [Google Scholar] [CrossRef]
- Yu, C.; Chen, D.; Liu, H.; Li, W.; Lu, J.; Feng, J. Rosmarinic Acid Reduces the Resistance of Gastric Carcinoma Cells to 5-Fluorouracil by Downregulating FOXO4-Targeting MiR-6785-5p. Biomed. Pharmacother. 2019, 109, 2327–2334. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.; Wei, C.; Rankin, G.O.; Rojanasakul, Y.; Ren, N.; Ye, X.; Chen, Y.C. Dietary Compound Proanthocyanidins from Chinese Bayberry (Myrica Rubra Sieb. et Zucc.) Leaves Inhibit Angiogenesis and Regulate Cell Cycle of Cisplatin-Resistant Ovarian Cancer Cells via Targeting Akt Pathway. J. Funct. Foods 2018, 40, 573–581. [Google Scholar] [CrossRef]
- Ravindranathan, P.; Pasham, D.; Goel, A. Oligomeric Proanthocyanidins (OPCs) from Grape Seed Extract Suppress the Activity of ABC Transporters in Overcoming Chemoresistance in Colorectal Cancer Cells. Carcinogenesis 2019, 40, 412–421. [Google Scholar] [CrossRef]
- Naus, P.J.; Henson, R.; Bleeker, G.; Wehbe, H.; Meng, F.; Patel, T. Tannic Acid Synergizes the Cytotoxicity of Chemotherapeutic Drugs in Human Cholangiocarcinoma by Modulating Drug Efflux Pathways. J. Hepatol. 2007, 46, 222–229. [Google Scholar] [CrossRef]
- Geng, N.; Zheng, X.; Wu, M.; Yang, L.; Li, X.; Chen, J. Tannic Acid Synergistically Enhances the Anticancer Efficacy of Cisplatin on Liver Cancer Cells through Mitochondria-Mediated Apoptosis. Oncol. Rep. 2019, 42, 2108–2116. [Google Scholar] [CrossRef]
- Yu, F.; Liu, W.; Gong, X.R.; Zhou, Y.B.; Lin, Y. Procyanidins Enhance the Chemotherapeutic Sensitivity of Laryngeal Carcinoma Cells to Cisplatin through Autophagy Pathway. Lin Chuang Er Bi Yan Hou Tou Jing Wai Ke Za Zhi J. Clin. Otorhinolaryngol. Head Neck Surg. 2018, 32, 447–456. [Google Scholar]
- Mady, F.M.; Shaker, M.A. Enhanced Anticancer Activity and Oral Bioavailability of Ellagic Acid through Encapsulation in Biodegradable Polymeric Nanoparticles. Int. J. Nanomed. 2017, 12, 7405. [Google Scholar] [CrossRef]
- Cetin, A.; Biltekin, B.; Degirmencioglu, S. Ellagic Acid Enhances the Antitumor Efficacy of Bevacizumab in an in Vitro Glioblastoma Model. World Neurosurg. 2019, 132, e59–e65. [Google Scholar] [CrossRef]
- Cetin, A.; Biltekin, B. Ellagic Acid Enhances Antitumor Efficacy of Temozolomide in an in Vitro Glioblastoma Model. Turk Neurosurg 2020, 30, 813–821. [Google Scholar] [CrossRef]
- Wu, Y.-S.; Ho, J.-Y.; Yu, C.-P.; Cho, C.-J.; Wu, C.-L.; Huang, C.-S.; Gao, H.-W.; Yu, D.-S. Ellagic Acid Resensitizes Gemcitabine-Resistant Bladder Cancer Cells by Inhibiting Epithelial-Mesenchymal Transition and Gemcitabine Transporters. Cancers 2021, 13, 2032. [Google Scholar] [CrossRef]
- Moon, J.Y.; Ediriweera, M.K.; Ryu, J.Y.; Kim, H.Y.; Cho, S.K. Catechol Enhances Chemo-and Radio-Sensitivity by Targeting AMPK/Hippo Signaling in Pancreatic Cancer Cells. Oncol. Rep. 2021, 45, 1133–1141. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecules | Origins | Experimental Approaches | Key Results | References |
|---|---|---|---|---|
| Apigenin | Synthetized | MDA435/LCC6 and P388 cells Cell proliferation assay ATPase assay | Enhanced the cytotoxicity of paclitaxel (PTX), doxorubicin (Dox), daunomycin (DM), vincristine (VCR), and vinblastine, resulting in a reduction of IC50 by 5–50 times | [11] |
| Purchased | Parental human HCC cells (BEL-7402) and BEL-7402/ADM cells MTT assay Cell cycle analysis Real-time quantitative PCR Western blot analysis In vivo xenograft studies | Sensitized Dox-resistant BEL-7402 (BEL- 7402/ADM) cells to Dox Increased intracellular concentration of Dox Reduced Nrf2 expression APG + Dox (in vivo) inhibited tumor growth, reduced cell proliferation, and induced apoptosis more substantially when compared with Dox treatment alone | [12] | |
| Purchased | Human pancreatic cancer cell line BxPC-3 Human pancreatic ductal epithelium (HPDE) cells Western blot analysis MTS cell proliferation assay | APG (13 μM) + gemcitabine (Gem) (13 μM) inhibited cell proliferation APG (11–19 μM) + Gem (10 μM) inhibited growth by 59–73% Enhanced the anti-proliferative activity of chemotherapeutic drugs | [13] | |
| Purchased | Human pancreatic cancer cell lines AsPc-1, Panc-1, and MiaPaCa-2 MTT assay Cell apoptosis assay Western blot analysis In vitro IKK-β kinase activity assay Xenograft model | Reduced cell growth Induced cell apoptosis Down-regulated the TNF-α-induced NF-κB DNA binding activity Suppressed pancreatic cancer growth and IKK-β activation in nude mice xenograft | [14] | |
| Purchased | Laryngeal carcinoma Hep-2 cell line RT-PCR Cell counting Kit-8 (CCK-8) system Western blot analysis | Enhanced the cisplatin (CP)-induced suppression of Hep-2 cell growth in a concentration- and time-dependent manner Reduced the levels of GLUT-1 mRNA and GLUT-1 and p-Akt proteins in CP-treated Hep-2 cells in a concentration and time-dependent manner | [15] | |
| Purchased | Laryngeal hep-2 carcinoma cell line Nude mouse model of laryngeal carcinoma Western blot analysis | Improved xenograft radio-sensitivity Reduced the expression of PI3K mRNA, Akt, and GLUT-1 after X-ray radiation | [16] | |
| Purchased | Tumor xenografts in nude mice SK-Hep-1 and BEL-7402 cells MTT assay Annexin V/PI assay Western blotting analysis Cellular ROS detection | Enhanced the cytotoxicity of 5-FU in HCC cells APG + 5-fluorouracil (5-FU) (in vivo) inhibited HCC xenograft tumor growth APG + 5-FU increased the levels of reactive oxygen species (ROS) APG + 5-FU decreased the mitochondrial membrane potential (ΔΨm) APG + 5-FU decreased Bcl-2 expression | [17] | |
| Purchased | Human renal proximal tubular epithelial (HK-2) cells MTT assay Analysis of cell morphology and cell cycleCaspase-3 activity assay Western blot analysis ROS production assay | Inhibited the CP-induced apoptosis of HK-2 cells Induced cell cycle arrest Inhibited caspase-3 activity and PARP cleavage Reduced CP-induced phosphorylation and expression of p53 Promoted the CP-induced Akt phosphorylation | [18] | |
| Purchased | 80 Swiss albino male mice ELISA Quantitative real-time RT-PCR Histopathological and immunohistochemical analysis | APG alone or combined with 5-FU Increased Beclin-1 levels, caspase-3 and -9, and JNK activities, decreased tumor volume, Mcl-1expression, and total antioxidant capacity, alleviated histopathological changes, and decreased Ki-67 proliferation index | [19] | |
| Not reported | BEL-7402 and BEL-7402/ADM cells TUNEL assay qRT-PCR Annexin V-FITC/PI apoptosis assay Western blot analysis | Reversed Dox sensitivity Induced the caspase-dependent apoptosis in BEL-7402/ADM cells Induced the miR-101 expression | [20] | |
| Not reported | Human hepatocellular carcinoma (HCC) and adjacent normal tissue specimens qRT-PCR MTT assay Western blot analysis In vivo xenograft studies | Enhanced Dox sensitivity Induced miR-520b expression Inhibited ATG7-dependent autophagy in BEL-7402/ADM cells Inhibited hepatocellular carcinoma xenograft growth | [21] | |
| Purchased | Ovarian cancer-sensitive cell line SKOV3 Ovarian cancer drug-resistant cell line SKOV3/DDP MTT assay PCR test Western blot test Apoptosis test | Enhanced the chemosensitivity of ovarian cancer-sensitive cells and drug-resistant cells Induced the apoptosis of ovarian cancer cells by down-regulating the Mcl-1 gene | [22] | |
| Quercetin | Purchased | DB-1 melanoma and SK Mel 28 cell lines Western blot analysis Annexin V-FITC staining RNA isolation and RT-PCR Immunocytochemistry siRNA transfection | Induced a redistribution of ΔNp73 in the cytoplasm and nucleus Que + temozolomide (TMZ) abolished drug insensitivity and caused a more than additive induction of apoptosis | [23] |
| Not reported | Human esophageal cancer cells (EC9706 and Eca109) MTT assay Annexin V-FITC/propidium iodide (PI)-stained fluorescence-activated cell sorting (FACS) Western blot analysis | Que + 5-FU inhibited growth and stimulated apoptosis in EC9706 and Eca109 esophageal cancer cells compared to Que | [24] | |
| Purchased | MCF-7 and MCF-7/Dox cells MTT assay Flow cytometry Matrigel invasion assay Western blot analysis | Increased intracellular concentration of Dox Improved Dox cytotoxicity Que + Dox inhibited cell proliferation and invasion and suppressed HIF-1α and P-gp expression | [25] | |
| Not reported | Human ovarian cancer cell lines, SKOV-3, EFO27, OVCAR-3, and A2780P Evaluation of quercetin toxicity SRB staining | Inhibited proliferation and increased sensitivity of ovarian cancer cells to CP and PTX | [26] | |
| Purchased | U251 and U87 human glioblastoma cells MTT assay Flow cytometry Western blot analysis | Que (30 μmol/L) + TMZ (100 μmol/L) inhibited cell viability and enhanced TMZ inhibition Que did not affect the caspase-3 activity and cell apoptosis, whereas combined with TMZ, it increased the caspase-3 activity and induced cell apoptosis. | [27] | |
| Purchased | MCF-7 cells and MCF-7/Dox cells MTT assay Flow cytometry | Que + Dox inhibited cell proliferation and invasion Que + Dox increased cell apoptosis Que + Dox up-regulated PTEN expression Que + Dox down-regulated p-Akt expression | [28] | |
| Purchased | Lung cancer cells (A549 and H460 cells) Western blot analysis | Reduced cell viability Suppressed HSP70 expression Improved Gem-induced cell death linked to increased caspase-3 and caspase-9 activities Que + Gem down-regulated HSP70 expression more significantly than treatment with Que or Gem alone | [29] | |
| Not reported | BEL-7402 and multidrug-resistant cell line BEL/5-FU MTT assay Flow cytometry Real-time PCR Western blot analysis | Increased intracellular accumulation of Dox Increased sensitivity of BEL/5-FU cells to chemotherapeutic drugs Down-regulated the expressions of ABCB1, ABCC1, and ABCC2 Inhibited the functions and expressions of ABCB1, ABCC1, and ABCC2 efflux pump | [30] | |
| Purchased | Human prostate cancer cell line PC3 MTT assay Western blot analysis Flow cytometry | Inhibited c-met expression and the downstream PI3K/AKT pathway Que + Dox promoted the Dox-induced cell apoptosis through the mitochondrial/ROS pathway | [31] | |
| Purchased | Human pancreatic cancer cell lines Transfection of small interfering RNA MTT assay Western blot analysis Cell cycle measurement | Attenuated RAGE expression to facilitate cell cycle arrest, autophagy, apoptosis, and GEM chemosensitivity in MIA Paca-2 GEMR cells | [32] | |
| Purchased | Human prostate cancer (PC-3) cell lines Nude male BALB/c mice MTT assay Intracellular ROS content assays RNA extraction and qRT-PCR Western blot analysis Immunohistochemistry | Que + PTX inhibited cell proliferation, increased apoptosis, arrested cell cycle at the G2/M phase, inhibited cell migration, induced ER stress, and increased ROS generation Que + PTX exerted the most beneficial therapeutic effects (in vivo) Increased the cancer cell-killing effects of PTX (in vivo) | [33] | |
| Not reported | MCF 7 cells MTT assay Flow cytometry qRT-PCR ELISA | Quer + 5-FU improved apoptosis by increasing the gene expression of Bax and p53 and caspase-9 activity and decreasing Bcl2 gene expression Quer + 5-FU decreased colony formation | [34] | |
| Purchased | MDA-MB-231 human breast cancer cell line MTT assay Flow cytometry qRT-PCR Western blot analysis | Decreased cell viability Que (95 μM) + docetaxel (7 nM) up-regulated p53, increased BAX levels, and decreased levels of BCL2, pERK1/2, AKT, and STAT3 proteins | [35] | |
| Kaempferol | Purchased | Human myelogenous leukemia K562 cells and the adriamycin-resistant variant K562/A cells MTT assay Annexin V/PI analysis PCR array | Kae + Que inhibited the growth of both cells Kae + Que increased the sensitivity of both cells Kae + Que induced apoptosis Kae + Que influenced the expression of drug transporter genes | [36] |
| Purchased | LS174 colon cancer cells MTT assay Colony formation assay Spheroid generation Sensitization assay Measurement of ROS Western blot analysis qRT-PCR | Chemo-sensitized 5-FU-resistant LS174-R cells Blocked the production of ROS and modulated the expression of JAK/STAT3, MAPK, PI3K/AKT, and NF-κBKae + 5-FU exerted a synergistic inhibitory effect on cell viability Kae + 5-FU enhanced apoptosis and induced cell cycle arrest in chemo-resistant and sensitive cells | [37] | |
| Purchased | Human colorectal cancer cell line HCT8 5-FU-resistant cell line HCT8-R CCK-8 assay qPCR assay Western blot analysis Clonogenic assay | Reversed the drug resistance of HCT8-R cells to 5-FU Reduced glucose uptake and lactic acid production in drug-resistant colorectal cancer cells Promoted the expression of microRNA-326 in colon cancer cells Reversed the resistance of colorectal cancer cells to 5-FU | [38] | |
| Myricetin | Purchased | Esophageal carcinoma EC9706 cells Colony formation assays Flow cytometry Western blot analysis Nude mouse tumor xenograft model | MYR + 5-FU suppressed cell survival fraction and proliferation, and increased cell apoptosis MYR + 5-FU decreased survivin, cyclin D, and Bcl-2, and increased the expression level of caspase-3 and p53 MYR + 5-FU reduced the growth rate of tumor xenografts in mice | [39] |
| Purchased | A2780 and OVCAR3 ovarian cancer cells MTT assay Apoptosis assay Boyden chamber assay Western blot analysis | Induced cytotoxicity, with an IC50 value of 25 μM Induced cell apoptosis, accompanied by the modulation of certain pro- and anti-apoptotic markers Increased paclitaxel cytotoxicity | [40] | |
| Rutin | Purchased | Human breast cancer MDA- MB-231 cellsCalcein acetoxymethyl accumulation assayRhodamine-123 uptake assay Annexin V and 7-aminoactinomycin D Propidium iodide staining | Increased the anticancer activity of both chemotherapeutic agents Decreased the activity of adenosine triphosphate binding cassette transporters RTN (20 μM) enhanced cytotoxicity related to cyclophosphamide and methotrexate RTN (20 and 50 μM) arrested the cell cycle at the G2/M and G0/G1 phases, respectively, thus promoting cell apoptosis | [41] |
| Purchased | Human HCC cell lines qRT-PCR Luciferase reporter assay Cell viability assay Flow cytometry In vivo tumor xenograft | Attenuated autophagy and BANCR expression in SO-resistant cells Decreased the number of autophagosomes in HepG2/SO and HCCLM3/SO cells Enhanced the efficacy of SO in a xenograft model of HCC in nude mice | [42] | |
| Hispidulin | Not reported | Human gallbladder carcinoma cell line GBC-SD MTT assay Western blot analysis Flow cytometry Caspase-3 activity assay qRT-PCR In vivo xenograft experiments | Inhibited the growth of GBC cells Promoted apoptosis in GBC cells Induced cell arrest at the G0/G1 phase Exerted antitumor effect mediated through HIF-1α inhibition Repressed the transactivation activity and expression of HIF-1α Suppressed the HIF-1α expression via AMPK signaling | [43] |
| Molecules | Origins | Experimental Approaches | Key Results | References |
|---|---|---|---|---|
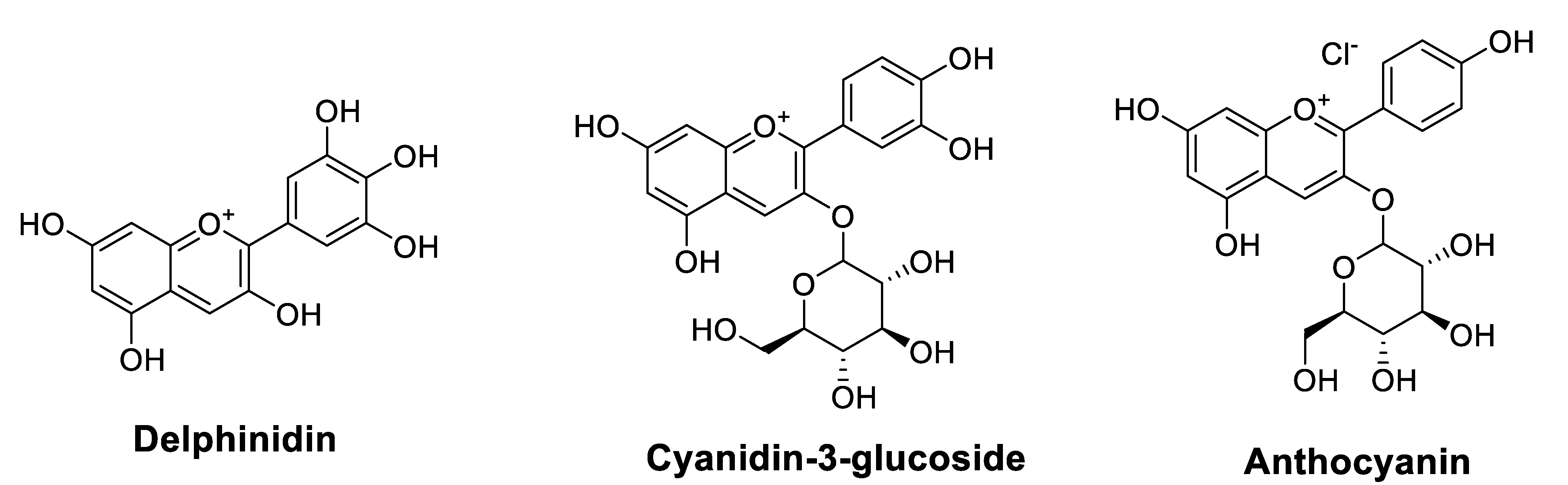
| Delphinidin | Purchased | A549 cell line (human, lung, and carcinoma) MTT assay Immunofluorescence staining Western blot analysis qRT-PCR | Induced apoptosis in A549 cells Promoted apoptosis in the radiation-exposed A549 cells Induced autophagy in radiation-exposed A549 cellsActivated autophagic cell death and the JNK/MAPK signaling pathway in radiation-exposed A549 cells | [45] |
| Cyanidin-3-glucoside (C3G) | Purchased | Human lung adenocarcinoma A549 cells Immunoblotting RNA isolation and qRT-PCR Immunofluorescence measurement Luciferase reporter assay | Reduced protein level of CLDN2 in A549 cells Inhibited Akt phosphorylation Increased p38 phosphorylation Reduced CLDN2 expression at transcriptional and post-translational steps mediated by Akt inhibition and p38 activation, respectively Improved Dox accumulation and cytotoxicity in spheroid models Increased the percentages of apoptotic and necrotic cells induced by Dox | [46] |
| Anthocyanins (ACNs) | Black raspberry | Colon cancer cell lines, SW480 and Caco2MTT assay Colony formation assays Western blot analysis Establishment of colitis-induced colon cancer mice model | Improved the chemotherapy efficacy of 5-FU and celecoxib ACNs + (5-FU or celecoxib) inhibited CRC cell proliferation (in vitro) and decreased the number of tumors in AOM-induced CRC mice (in vivo) | [44] |
| Molecules | Origins | Experimental Approaches | Key Results | References |
|---|---|---|---|---|
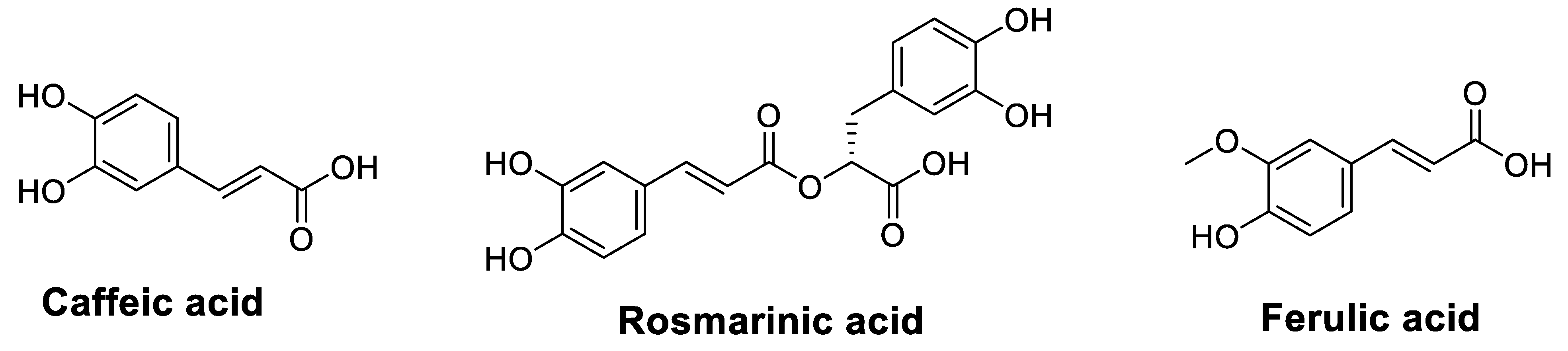
| Caffeic acid phenethyl ester (CAPE) | Not reported | Mouse CT26 colorectal adenocarcinoma cells BALB/c mouse with CT26 cells implantation Colony formation assay RT-PCR Flow cytometry | Depleted intracellular GSH in CT26 cells, but not in bone marrow cells Enhanced cell killing by IR Increased glutathione peroxidase, decreased glutathione reductase in CT26 cells Reversed radiation-activated NF-κB Induced a significant inhibition of tumor growth and prolongation of survival compared to IR alone (in vivo) | [49] |
| Purchased | Human medulloblastoma Daoy cell line and Human astroglia SVGp12 MTT and trypan blue exclusion assays ELISATUNEL assay Flow cytometry Western blot analysis | Inhibited Daoy cell growth in a time- and concentration-dependent manner Decreased G2/M fraction and increased S phase fractionDown-regulated expression of cyclin B1 protein Reduced the viability of irradiated Daoy cells No chemosensitizing effect on Dox or CP | [50] | |
| Purchased | Parental and the drug-resistant cells of stomach (MKN45) and colon (LoVo) cancers | Potentiated the apoptotic effects of Dox and CP against parental cells Reduced the production of Dox-induced ROS Reduced 26S proteasome-based proteolytic activities in parental MKN45 cells Up-regulated and significantly decreased chymotrypsin-like activity in Dox- or CP-resistant cells | [51] | |
| Not reported | Human lung adenocarcinoma A549 and RERF-LC-MS cell lines Immunoblotting RNA isolation and PCR Luciferase reporter assay Immunocytochemistry | Decreased claudin-2 protein level in a concentration-dependent manner Decreased (at 50 µM) mRNA level and promoter activity Decreased (at 50 µM) the level of p-NF−κB, and increased that of IκB Increased the expression and activity of protein phosphatase (PP) 1 and 2A Suppressed cell proliferation Enhanced Dox toxicity and accumulation in 3D spheroid cells | [52] | |
| Not reported | Prostate cancer (PCa) cells, DU145 and PC3 Evaluated the radiomodulatory potential of CAPE | CAPE + gamma radiation (GR) sensitized PCa cells to radiation in a concentration-dependent manner Improved the level of ionizing radiation (IR)-induced gamma H2AX foci and cell death by apoptosis CAPE + GR decreased the migration potential of PCa cells Sensitized PCa cells to radiation in vitro and induced apoptosis, increased Akt/mTOR phosphorylation and hampered cell migration CAPE + IR inhibited cell growth by decreasing RAD50 and RAD51 proteins | [53] | |
| Ferulic acid (FA) | Purchased | Multidrug resistance (MDR) cell lines MTT assay Colony formation assay Fluorescence microscopic analysis Cell cycle analysis Tryptophan fluorescence quenching PCR array Western blot analysis | Inhibited P-glycoprotein transport function in drug-resistant KB ChR8-5 cell lines Down-regulated ABCB1 expression in a concentration-dependent manner Decreased paclitaxel resistance in KBChR8-5 and HEK293/ABCB1 cells Enhanced paclitaxel-mediated cell cycle arrest and up-regulated paclitaxel-induced apoptotic signaling in KB-resistant cells | [54] |
| Purchased | Parental KB cells and P−gp overexpressing KB ChR8-5 cell lines MTT assay γH2AX assay Western blot analysis Immunocytochemistry Animals and tumor xenograft experiments | Increased the cytotoxicity of Dox and VCR in the P-gp overexpressing KB ChR8-5 cells Enhanced the formation of Dox-induced γH2AX foci and synergistically increased Dox-induced apoptotic signaling in drug-resistant cells FA + Dox reduced KB ChR8−5 tumor xenograft size three-fold compared to the group treated with Dox alone Reversed MDR by suppressing P-gp expression via inhibition of PI3K/Akt/NF−κB signaling pathway | [55] | |
| Rosmarinic acid (RA) | Purchased | Human acute promyelocytic leukemia NB4 cells Flow cytometry analysis Phagocytosis assay qRT-PCR | Potentiated ATRA-induced macrophage differentiation in APL cells | [56] |
| Not reported | Human gastric carcinoma cell line SGC7901 Apoptosis assay CCK8 assay Apoptosis assay RNA isolation and microarray qRT-PCR Luciferase reporter assay Western blot analysis | Increased the chemosensitivity of SGC7901 cells to 5-FUReduced IC50 of 5-FU (70.43 ± 1.06 μg/mL) compared to untreated SGC7901/5-FU cells (208.6 ± 1.09 μg/mL) RA + 5-FU increased apoptosis rate Reduced the expression levels of two miRNAs (miR-642a−3p and miR−6785-5p) Reduced P-gp expression and increased Bax expression in SGC7901/5-FU and SGC7901/5-FU -Si cells | [57] |
| Molecules | Origins | Experimental Approaches | Key Results | References |
|---|---|---|---|---|
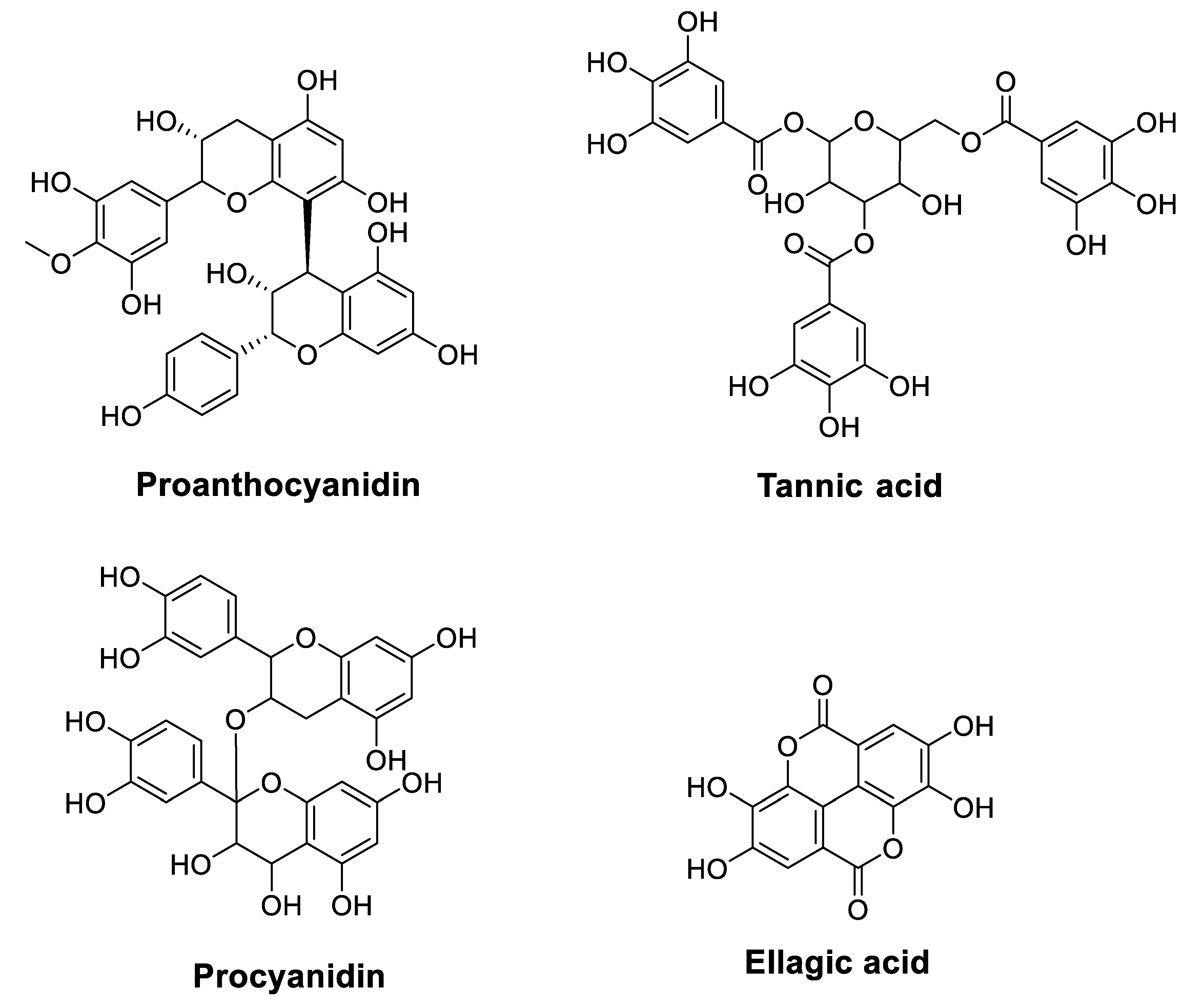
| Proanthocyanidins | Chinese bayberry leaves | Platinum-resistant human ovarian cancer cell line OVCAR-3 Flow cytometry MTT assay Colony formation assay Western blot assay | Induced inhibitory effects on the growth and CSC characteristics of OVCAR−3 SP cells Reduced the expression of β-catenin, cyclin D1, and c-Myc and inhibited the self-renewal capacity of cells Induced G1 cell cycle arrest in OVCAR−3 SP cells | [58] |
| Grape seed extract | Colorectal cancer cell lines, HCT116 and H716 Cell cycle and apoptosis analysis Cell viability and proliferation mRNA expression analysis Genome-wide RNA-sequencing analysisXenograft animal experiments | Sensitized acquired (HCT116-FOr cells) and innately chemoresistant (H716 cells) cancer cells to 5-FU and oxaliplatin (OXP) PCs + (5-FU and OXP) inhibited the growth of chemoresistant cells and decreased the expression of several key adenosine triphosphate-binding cassette (ABC) transporters Sensitized chemoresistant cells to 5-FU and OXPPCs + (5-FU and OXP) reduced chemoresistant xenograft tumor growth in mice | [59] | |
| Tannic acid | Purchased | Malignant human cholangiocytes Calcein retention assays Western blot analysis RT-PCR | Decreased malignant cholangiocyte growth Exhibited a synergistic effect with mitomycin C and 5-FU but not with Gem Decreased calcein efflux and expression of PGP, MRP1, and MRP2 membrane efflux pumps | [60] |
| Purchased | Liver cancer cell line HepG2 MTT assay Mitochondrial transmembrane potential qRT-PCR Western blot analysis | Inhibited HepG2 cell growth TA + CP induced mitochondria-mediated apoptosis in HepG2 cells and enhanced growth inhibitory effect compared to treatment alone | [61] | |
| Procyanidins | Not reported | Laryngeal cancer cell line TU686 Flow cytometry Cell immunofluorescence staining Western blot analysis | Inhibited TU686 cells in a concentration-dependent manner for 24 h Induced apoptosis of TU686 cells Increased expression of LC3−Ⅱ and Caspase-3 | [62] |
| Ellagic acid | Purchased | Colorectal carcinoma HT−29, Colo 320DM, SW480, and LoVo cells Trypan blue exclusion Annexin−V labeling Mitochondrial membrane potential (Δψm) Immunoblotting | EA + 5-FU inhibited cell proliferation of HT-29, Colo 320DM and SW480 cells EA + 5-FU increased apoptotic cell death of HT−29 and Colo 320DM cells EA potentiated 5-FU chemosensitivity in at least three colorectal cancer cell lines | [47] |
| Purchased | Epithelial ovarian cancer cell line A2780MTT assay Immunoblot analysis Signal pathway analysis Cell cycle analysis | Enhanced CP cytotoxicity in A2780CisR cells Prevented the development of CP resistance | [48] | |
| Purchased | Caco-2 and HTC-116 cells MTT assay In vitro drug release Male New Zealand white rabbits | Induced higher cell viability than EA-NP treated HCT−116 cells Oral administration of EA-NPs caused a 3.6-fold increase in the area under the curve compared to that of EA (in vivo) | [63] | |
| Purchased | Rat C6 glioma cells Immunohistochemistry RT-PCR | Reduced MGMT expression Affected the apoptotic proteins of p53 and caspase-3 at the protein level, but not at the gene level EA + bevacizumab (BEV) reduced cell viability EA + BEV reduced MDR1 expression only at 72 h | [64] | |
| Purchased | Rat C6 glioma cells Immunocytochemistry RT-PCR | EA + TMZ reduced cell viability Down-regulated MGMT expression independent of the presence of TMZ EA + TMZ reduced MDR1 expression only over 48 h compared to TMZ alone Up-regulated caspase-3 at 48 h, but up-regulated p53 at 48 and 72 h EA + TMZ enhanced immunoreactivities of p53 and caspase-3 proteins, but not of the genes | [65] | |
| Purchased | Four human bladder cancer cell lines, TSGH−8301, TSGH-9202, T24, and J82 MTT assay Flow cytometry Cell migration and invasion assays Western blot analysis qRT−PCR Xenograft model | Induced high cytotoxicity of Gem in GEM−resistant cells EA + Gem increased apoptosis and reduced cell motility in GCB-resistant cells Resensitized bladder cancer cells to Gem by reducing the epithelial–mesenchymal transition Reduced EMT by inhibiting the TGFβ−SMAD2/3 upward signaling pathway Inhibited the growth of bladder cancer tumors and increased the in vivo inhibitory effects of Gen on tumors | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouyahya, A.; Omari, N.E.; Bakrim, S.; Hachlafi, N.E.; Balahbib, A.; Wilairatana, P.; Mubarak, M.S. Advances in Dietary Phenolic Compounds to Improve Chemosensitivity of Anticancer Drugs. Cancers 2022, 14, 4573. https://doi.org/10.3390/cancers14194573
Bouyahya A, Omari NE, Bakrim S, Hachlafi NE, Balahbib A, Wilairatana P, Mubarak MS. Advances in Dietary Phenolic Compounds to Improve Chemosensitivity of Anticancer Drugs. Cancers. 2022; 14(19):4573. https://doi.org/10.3390/cancers14194573
Chicago/Turabian StyleBouyahya, Abdelhakim, Nasreddine El Omari, Saad Bakrim, Naoufal El Hachlafi, Abdelaali Balahbib, Polrat Wilairatana, and Mohammad S. Mubarak. 2022. "Advances in Dietary Phenolic Compounds to Improve Chemosensitivity of Anticancer Drugs" Cancers 14, no. 19: 4573. https://doi.org/10.3390/cancers14194573
APA StyleBouyahya, A., Omari, N. E., Bakrim, S., Hachlafi, N. E., Balahbib, A., Wilairatana, P., & Mubarak, M. S. (2022). Advances in Dietary Phenolic Compounds to Improve Chemosensitivity of Anticancer Drugs. Cancers, 14(19), 4573. https://doi.org/10.3390/cancers14194573