
Copy Number Profiles of Prostate Cancer in Men of Middle Eastern Ancestry

 , , , and
, , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Fluorescence In Situ Hybridization Patient Cohort
2.2. OncoScan SNP Array Patient Cohort
2.3. SNP Microarray Data Generation and CNA Calling
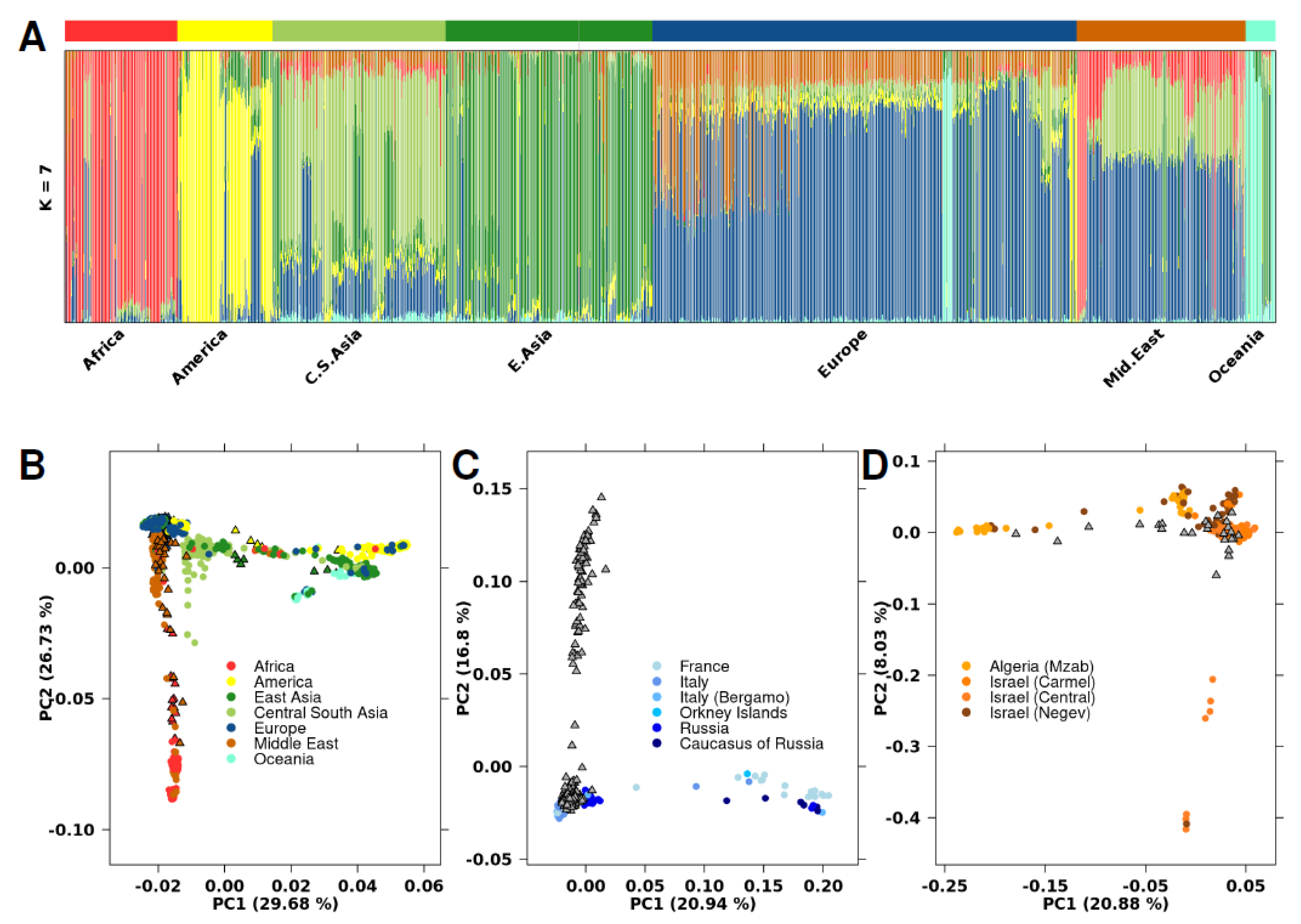
2.4. Inferring Ancestry
2.5. Statistical Analysis
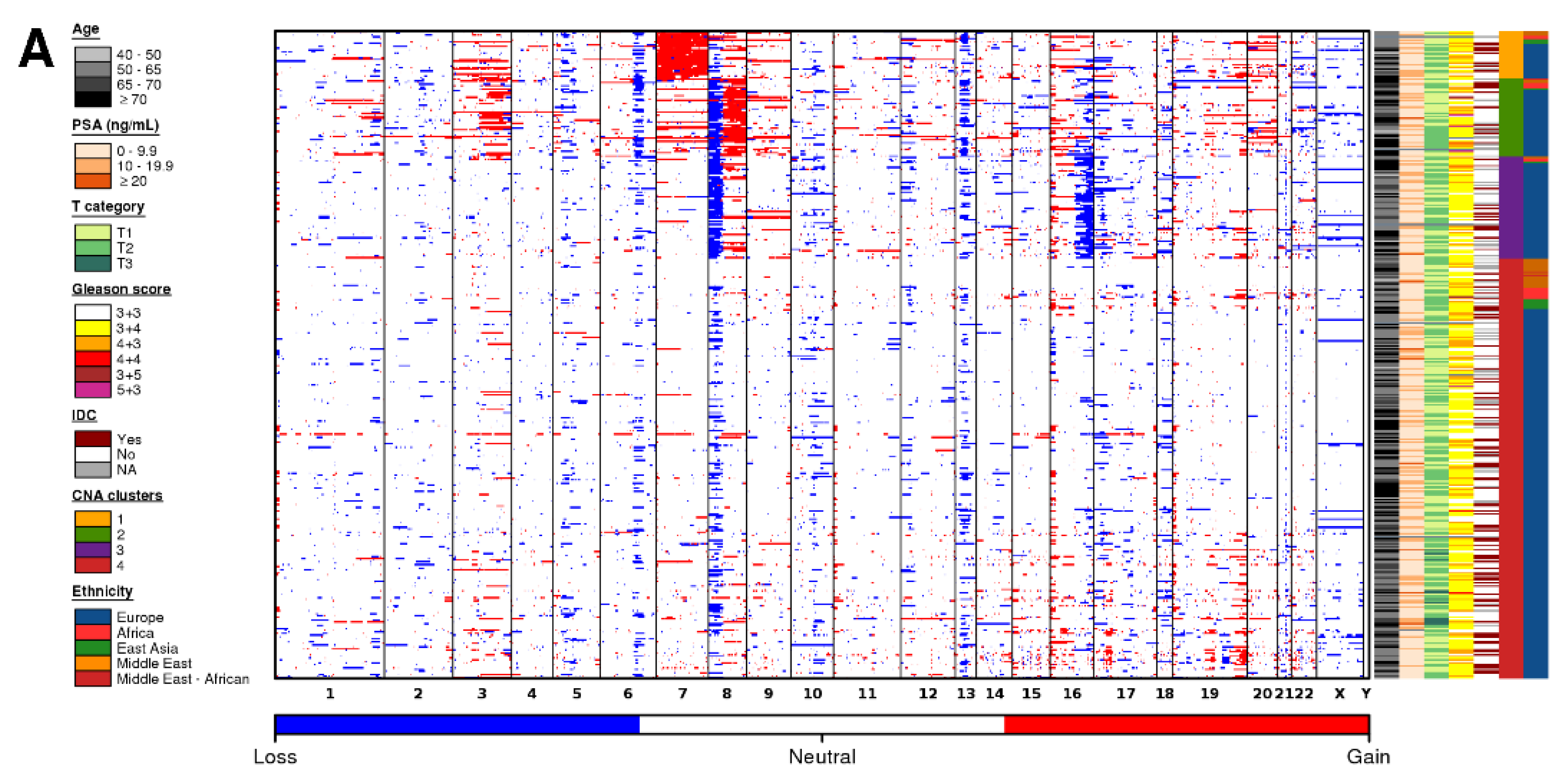
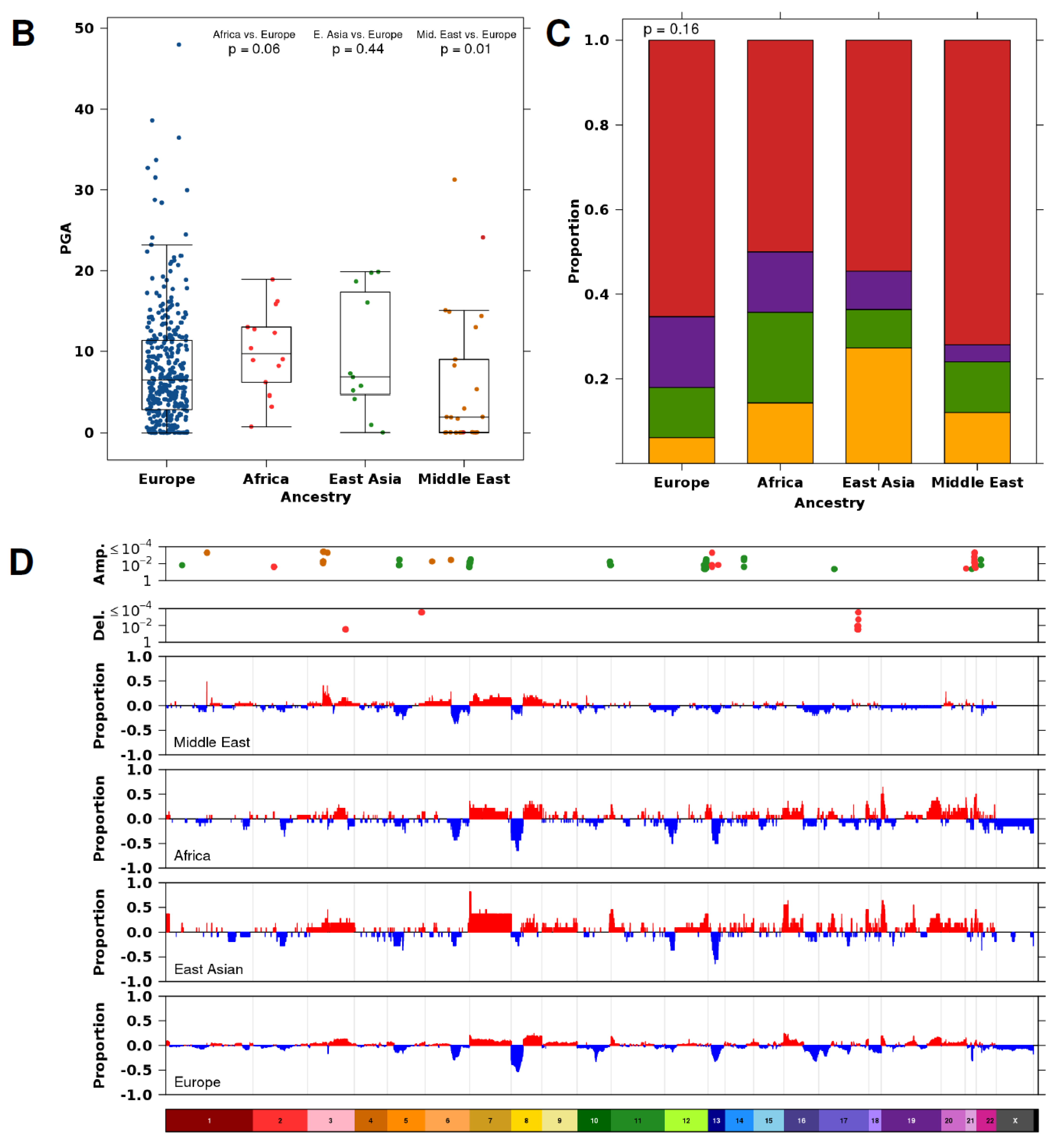
3. Results
3.1. Genetic Ancestry Inference
3.2. Ancestry-Associated CNA Mutation Density
3.3. Ancestry-Associated CNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weischenfeldt, J.; Simon, R.; Feuerbach, L.; Schlangen, K.; Weichenhan, D.; Minner, S.; Wuttig, D.; Warnatz, H.-J.; Stehr, H.; Rausch, T.; et al. Integrative Genomic Analyses Reveal an Androgen-Driven Somatic Alteration Landscape in Early-Onset Prostate Cancer. Cancer Cell 2013, 23, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.A.; Fraser, M.; Livingstone, J.; Espiritu, S.M.G.; Thorne, H.; Huang, V.; Lo, W.; Shiah, Y.-J.; Yamaguchi, T.N.; Sliwinski, A.; et al. Germline BRCA2 Mutations Drive Prostate Cancers with Distinct Evolutionary Trajectories. Nat. Commun. 2017, 8, 13671. [Google Scholar] [CrossRef] [PubMed]
- SEER Cancer Statistics Review, 1975–2016. Available online: https://seer.cancer.gov/csr/1975_2016/index.html (accessed on 24 October 2019).
- Washburn, J.G.; Wojno, K.J.; Dey, J.; Powell, I.J.; Macoska, J.A. 8pter-P23 Deletion Is Associated with Racial Differences in Prostate Cancer Outcome. Clin. Cancer Res. 2000, 6, 4647–4652. [Google Scholar] [PubMed]
- Shan, J.; Al-Rumaihi, K.; Rabah, D.; Al-Bozom, I.; Kizhakayil, D.; Farhat, K.; Al-Said, S.; Kfoury, H.; Dsouza, S.P.; Rowe, J.; et al. Genome Scan Study of Prostate Cancer in Arabs: Identification of Three Genomic Regions with Multiple Prostate Cancer Susceptibility Loci in Tunisians. J. Transl. Med. 2013, 11, 121. [Google Scholar] [CrossRef]
- Al Olama, A.A.; Kote-Jarai, Z.; Berndt, S.I.; Conti, D.V.; Schumacher, F.; Han, Y.; Benlloch, S.; Hazelett, D.J.; Wang, Z.; Saunders, E.; et al. A Meta-Analysis of 87,040 Individuals Identifies 23 New Susceptibility Loci for Prostate Cancer. Nat. Genet. 2014, 46, 1103–1109. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.-P.; Chen, R.; Zhang, C.-L.; Zhang, W.; Xiao, G.-A.; Wang, F.-B.; Ta, N.; Gao, X.; Sun, Y.-H. Prevalence and Clinical Application of TMPRSS2-ERG Fusion in Asian Prostate Cancer Patients: A Large-Sample Study in Chinese People and a Systematic Review. Asian J. Androl. 2019. [Google Scholar] [CrossRef]
- Tonon, L.; Fromont, G.; Boyault, S.; Thomas, E.; Ferrari, A.; Sertier, A.-S.; Kielbassa, J.; Le Texier, V.; Kamoun, A.; Elarouci, N.; et al. Mutational Profile of Aggressive, Localised Prostate Cancer from African Caribbean Men Versus European Ancestry Men. Eur. Urol. 2019, 75, 11–15. [Google Scholar] [CrossRef]
- Abdelsalam, R.A.; Khalifeh, I.; Box, A.; Kalantarian, M.; Ghosh, S.; Abou-Ouf, H.; Lotfi, T.; Shahait, M.; Palanisamy, N.; Bismar, T.A. Molecular Characterization of Prostate Cancer in Middle Eastern Population Highlights Differences with Western Populations with Prognostic Implication. J. Cancer Res. Clin. Oncol. 2020, 146, 1701–1709. [Google Scholar] [CrossRef]
- Espiritu, S.M.G.; Liu, L.Y.; Rubanova, Y.; Bhandari, V.; Holgersen, E.M.; Szyca, L.M.; Fox, N.S.; Chua, M.L.K.; Yamaguchi, T.N.; Heisler, L.E.; et al. The Evolutionary Landscape of Localized Prostate Cancers Drives Clinical Aggression. Cell 2018, 173, 1003–1013.e15. [Google Scholar] [CrossRef]
- Lalonde, E.; Ishkanian, A.S.; Sykes, J.; Fraser, M.; Ross-Adams, H.; Erho, N.; Dunning, M.J.; Halim, S.; Lamb, A.D.; Moon, N.C.; et al. Tumour Genomic and Microenvironmental Heterogeneity for Integrated Prediction of 5-Year Biochemical Recurrence of Prostate Cancer: A Retrospective Cohort Study. Lancet Oncol. 2014, 15, 1521–1532. [Google Scholar] [CrossRef]
- Fraser, M.; Sabelnykova, V.Y.; Yamaguchi, T.N.; Heisler, L.E.; Livingstone, J.; Huang, V.; Shiah, Y.-J.; Yousif, F.; Lin, X.; Masella, A.P.; et al. Genomic Hallmarks of Localized, Non-Indolent Prostate Cancer. Nature 2017, 541, 359–364. [Google Scholar] [CrossRef]
- Sinha, A.; Huang, V.; Livingstone, J.; Wang, J.; Fox, N.S.; Kurganovs, N.; Ignatchenko, V.; Fritsch, K.; Donmez, N.; Heisler, L.E.; et al. The Proteogenomic Landscape of Curable Prostate Cancer. Cancer Cell 2019, 35, 414–427.e6. [Google Scholar] [CrossRef] [PubMed]
- Rebbeck, T.R. Prostate Cancer Genetics: Variation by Race, Ethnicity, and Geography. Semin. Radiat. Oncol. 2017, 27, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Cann, H.M.; de Toma, C.; Cazes, L.; Legrand, M.-F.; Morel, V.; Piouffre, L.; Bodmer, J.; Bodmer, W.F.; Bonne-Tamir, B.; Cambon-Thomsen, A.; et al. A Human Genome Diversity Cell Line Panel. Science 2002, 296, 261–262. [Google Scholar] [CrossRef]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The Variant Call Format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast Model-Based Estimation of Ancestry in Unrelated Individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef]
- P’ng, C.; Green, J.; Chong, L.C.; Waggott, D.; Prokopec, S.D.; Shamsi, M.; Nguyen, F.; Mak, D.Y.F.; Lam, F.; Albuquerque, M.A.; et al. BPG: Seamless, Automated and Interactive Visualization of Scientific Data. BMC Bioinform. 2019, 20, 42. [Google Scholar] [CrossRef]
- Van Loo, P.; Nilsen, G.; Nordgard, S.H.; Vollan, H.K.M.; Børresen-Dale, A.-L.; Kristensen, V.N.; Lingjærde, O.C. Analyzing Cancer Samples with SNP Arrays. Methods Mol. Biol. 2012, 802, 57–72. [Google Scholar] [CrossRef]
- Phillips, C.; Parson, W.; Lundsberg, B.; Santos, C.; Freire-Aradas, A.; Torres, M.; Eduardoff, M.; Børsting, C.; Johansen, P.; Fondevila, M.; et al. Building a Forensic Ancestry Panel from the Ground up: The EUROFORGEN Global AIM-SNP Set. Forensic Sci. Int. Genet. 2014, 11, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Z.; Absher, D.M.; Tang, H.; Southwick, A.M.; Casto, A.M.; Ramachandran, S.; Cann, H.M.; Barsh, G.S.; Feldman, M.; Cavalli-Sforza, L.L.; et al. Worldwide Human Relationships Inferred from Genome-Wide Patterns of Variation. Science 2008, 319, 1100–1104. [Google Scholar] [CrossRef] [PubMed]
- Houlahan, K.E.; Shiah, Y.-J.; Gusev, A.; Yuan, J.; Ahmed, M.; Shetty, A.; Ramanand, S.G.; Yao, C.Q.; Bell, C.; O’Connor, E.; et al. Genome-Wide Germline Correlates of the Epigenetic Landscape of Prostate Cancer. Nat. Med. 2019, 25, 1615–1626. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Wei, G.-H.; Liu, D.; Wang, L.; Hou, Y.; Zhu, S.; Peng, L.; Zhang, Q.; Cheng, Y.; Su, H.; et al. Whole-Genome and Transcriptome Sequencing of Prostate Cancer Identify New Genetic Alterations Driving Disease Progression. Eur. Urol. 2018, 73, 322–339. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Mosquera, J.M.; Garofalo, A.; Oh, C.; Baco, M.; Amin-Mansour, A.; Rabasha, B.; Bahl, S.; Mullane, S.A.; Robinson, B.D.; et al. Exome Sequencing of African-American Prostate Cancer Reveals Loss-of-Function ERF Mutations. Cancer Discov. 2017, 7, 973–983. [Google Scholar] [CrossRef]
- Jaratlerdsiri, W.; Chan, E.K.F.; Gong, T.; Petersen, D.C.; Kalsbeek, A.M.F.; Venter, P.A.; Stricker, P.D.; Bornman, M.S.R.; Hayes, V.M. Whole-Genome Sequencing Reveals Elevated Tumor Mutational Burden and Initiating Driver Mutations in African Men with Treatment-Naïve, High-Risk Prostate Cancer. Cancer Res. 2018, 78, 6736–6746. [Google Scholar] [CrossRef]
- Takata, R.; Takahashi, A.; Fujita, M.; Momozawa, Y.; Saunders, E.J.; Yamada, H.; Maejima, K.; Nakano, K.; Nishida, Y.; Hishida, A.; et al. 12 New Susceptibility Loci for Prostate Cancer Identified by Genome-Wide Association Study in Japanese Population. Nat. Commun. 2019, 10, 4422. [Google Scholar] [CrossRef]
- Emeville, E.; Broquère, C.; Brureau, L.; Ferdinand, S.; Blanchet, P.; Multigner, L.; Romana, M. Copy Number Variation of GSTT1 and GSTM1 and the Risk of Prostate Cancer in a Caribbean Population of African Descent. PLoS ONE 2014, 9, e107275. [Google Scholar] [CrossRef]
- Wong, M.C.S.; Goggins, W.B.; Wang, H.H.X.; Fung, F.D.H.; Leung, C.; Wong, S.Y.S.; Ng, C.F.; Sung, J.J.Y. Global Incidence and Mortality for Prostate Cancer: Analysis of Temporal Patterns and Trends in 36 Countries. Eur. Urol. 2016, 70, 862–874. [Google Scholar] [CrossRef]
- Hussain, A.M.A.; Lafta, R.K. Cancer Trends in Iraq 2000–2016. Oman. Med. J. 2021, 36, e219. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Dataset | Europe | Africa | E. Asia | N. American | Mid. East |
|---|---|---|---|---|---|
| ISUP Grade Group | |||||
| 1 | 60 | 1 | 3 | 1 | 9 |
| 2 | 200 | 9 | 6 | 3 | 12 |
| 3 | 75 | 4 | 2 | 1 | 3 |
| 4 | 7 | 0 | 0 | 0 | 1 |
| 5 | 2 | 0 | 0 | 0 | 0 |
| NA | 2 | 0 | 0 | 0 | 0 |
| Age at Treatment | |||||
| 40–50 | 7 | 1 | 0 | 1 | 0 |
| 50–65 | 160 | 4 | 5 | 3 | 8 |
| 65–70 | 74 | 2 | 1 | 0 | 12 |
| ≥70 | 105 | 7 | 5 | 1 | 5 |
| Pre-treatment PSA | |||||
| ≤9.9 ng/mL | 4 | 1 | 11 | 3 | 0 |
| 10–19.9 ng/mL | 93 | 3 | 0 | 1 | 8 |
| ≥20 ng/mL | 249 | 10 | 0 | 1 | 17 |
| T category | |||||
| T1 | 165 | 7 | 4 | 4 | 1 |
| T2 | 181 | 7 | 7 | 1 | 13 |
| T3 | 0 | 0 | 0 | 0 | 11 |
| Intraductal carcinoma or cribriform architecture | |||||
| Yes | 196 | 3 | 0 | 0 | 21 |
| No | 100 | 10 | 9 | 4 | 4 |
| NA | 50 | 1 | 2 | 1 | 0 |
| Total | 346 | 14 | 11 | 5 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albawardi, A.; Livingstone, J.; Almarzooqi, S.; Palanisamy, N.; Houlahan, K.E.; Awwad, A.A.A.; Abdelsalam, R.A.; Boutros, P.C.; Bismar, T.A. Copy Number Profiles of Prostate Cancer in Men of Middle Eastern Ancestry. Cancers 2021, 13, 2363. https://doi.org/10.3390/cancers13102363
Albawardi A, Livingstone J, Almarzooqi S, Palanisamy N, Houlahan KE, Awwad AAA, Abdelsalam RA, Boutros PC, Bismar TA. Copy Number Profiles of Prostate Cancer in Men of Middle Eastern Ancestry. Cancers. 2021; 13(10):2363. https://doi.org/10.3390/cancers13102363
Chicago/Turabian StyleAlbawardi, Alia, Julie Livingstone, Saeeda Almarzooqi, Nallasivam Palanisamy, Kathleen E. Houlahan, Aktham Adnan Ahmad Awwad, Ramy A. Abdelsalam, Paul C. Boutros, and Tarek A. Bismar. 2021. "Copy Number Profiles of Prostate Cancer in Men of Middle Eastern Ancestry" Cancers 13, no. 10: 2363. https://doi.org/10.3390/cancers13102363
APA StyleAlbawardi, A., Livingstone, J., Almarzooqi, S., Palanisamy, N., Houlahan, K. E., Awwad, A. A. A., Abdelsalam, R. A., Boutros, P. C., & Bismar, T. A. (2021). Copy Number Profiles of Prostate Cancer in Men of Middle Eastern Ancestry. Cancers, 13(10), 2363. https://doi.org/10.3390/cancers13102363