Simple Summary
Regulatory CD4+ T cell (Treg) is one of the suppressive immune cells, but data on its clinical relevance in large human breast cancer cohort is limited. Abundance of Tregs in 5177 breast cancer patient samples from five independent cohorts was analyzed by the xCell algorithm using tumor transcriptomics. Treg abundance was lower in metastatic tumors when compared to matched primary tumors. Treg was associated with a high mutation rate of TP53 genes and copy number mutations as well as with increased tumor infiltration of M2 macrophages and decreased infiltration of T helper type 1 cells. Interestingly, low Treg abundance was significantly associated with pathological complete response (pCR) after neoadjuvant chemotherapy (NAC) in TNBC, but not in ER-positive/Her2-negative subtype. Abundance of Treg was also associated with high expression of multiple immune checkpoint molecules. In conclusion, Treg abundance may have a potential as a predictive biomarker of pCR after NAC in TNBC.
Abstract
Regulatory CD4+ T cell (Treg), a subset of tumor-infiltrating lymphocytes (TILs), are known to suppress anticancer immunity but its clinical relevance in human breast cancer remains unclear. In this study, we estimated the relative abundance of Tregs in breast cancer of multiple patient cohorts by using the xCell algorithm on bulk tumor gene expression data. In total, 5177 breast cancer patients from five independent cohorts (TCGA-BRCA, GSE96058, GSE25066, GSE20194, and GSE110590) were analyzed. Treg abundance was not associated with cancer aggressiveness, patient survival, or immune activity markers, but it was lower in metastatic tumors when compared to matched primary tumors. Treg was associated with a high mutation rate of TP53 genes and copy number mutations as well as with increased tumor infiltration of M2 macrophages and decreased infiltration of T helper type 1 (Th1) cells. Pathological complete response (pCR) after neoadjuvant chemotherapy (NAC) was significantly associated with low Treg abundance in triple negative breast cancer (TNBC) but not in ER-positive/Her2-negative subtype. High Treg abundance was significantly associated with high tumor expression of multiple immune checkpoint inhibitor genes. In conclusion, Treg abundance may have potential as a predictive biomarker of pCR after NAC in TNBC.
1. Introduction
Tumor-infiltrating lymphocytes (TILs) have gained growing attention because they indicate a better prognosis in several types of cancer [1,2,3]. Breast cancer typically has less immune cell infiltration compared to other cancers. Triple negative breast cancer (TNBC) is the most aggressive subtype because of its worst outcome [4]; however, TNBC is also the most immunogenic subtype that attracts TILs due to genomic instability and higher mutation rate [5,6]. Tumoral abundance of TILs in breast cancer predicts response to neoadjuvant chemotherapy (NAC) and correlates with better prognosis in TNBC [7,8].
NAC for locally advanced breast cancer not only facilitates breast conserving surgery, but it also tests the tumor’s sensitivity to anti-cancer treatment, thereby guiding adjuvant treatment [9]. However, there are also reports that contradict the TILs’ effect on the response to NAC or prognosis [10]. One of the reasons for this discrepancy is that TILs are a heterogeneous cell population containing both tolerogenic regulatory CD4+FoxP3+ T cells (Tregs) and tumor recognizing CD8+ effector T cells. Currently, TILs are identified by pathology in clinical practice that does not distinguish between these different subsets of T cells.
Regulatory CD4+ T cells (Tregs) are a type of TILs that has gained considerable attention [11,12,13]. Tregs play a critical role in immune tolerance and they suppress antitumor immune responses [14,15]. Tumor-infiltrating Tregs suppress the proliferation of effector T lymphocytes, which prohibits an adequate anti-cancerous immune response and promotes tumor growth [16]. However, the prognostic significance of Tregs in breast cancer remains ambiguous, and data from large patient cohort studies are awaited to fully understand their clinical relevance. Tregs are mostly identified by immunohistochemistry (IHC) of Forkhead box protein 3 (Foxp3) protein. However, because Foxp3+ T lymphocytes are heterogeneous cells, Foxp3 IHC staining alone is insufficient to identify these cells [17,18,19]. Although IHC is the gold standard to identify and quantify immune cells, it is examiner-dependent and therefore results may vary and be inaccurate. Our group and others have reported the clinical relevance of the tumor immune microenvironment (TIME) using computational algorithms from bulk tumor transcriptome of the patient cohorts [20,21,22,23,24]. Utilizing xCell algorithm on transcriptomic data, we found that adipocytes in TIME of breast cancer are associated with metastasis and inflammation-related pathways particularly in ER-positive/human epidermal growth factor receptor 2 (HER2)-negative breast cancer [25]. We also found that, in breast cancer, a biologically aggressive phenotype and anti-cancerous immunity is associated with high mutation rate [22]. An advantage of this approach is that it allows the analyses of actual patient cohorts that are linked with transcriptomic data but were collected for completely unrelated motives. In this study, we hypothesized that transcriptionally defined low levels of tumor infiltration of Tregs is associated with pathological complete response (pCR) after NAC and can serve as a predicative biomarker in breast cancer patients. In this study, we analyzed 5177 breast cancer patients from five completely independent cohorts—TCGA-BRCA (n = 1065) [26], GSE96058 (n = 3273) [27], GSE25066 (n = 508) [28] GSE20194 (n = 248) [29], and GSE110590 (n = 83)—to study the association of Treg abundance, pathological characteristics, and clinical outcomes of patients with various breast cancer types.
2. Results
2.1. Abundance of Regulatory T Cell (Treg) in Tumors Is Not Associated with Cancer Aggressiveness
We utilized the xCell algorithm [30] to estimate the relative abundance of Tregs from transcriptome of a bulk tumor. The xCell algorithm defined Treg by expression profile of 39 genes (Table S1). First, we were interested to find out whether stage, grade, and disease-free and disease specific survival are associated with the levels Treg infiltration in patients with different types of breast cancer. In the breast cancer cohort of The Cancer Genome Atlas (TCGA-BRCA; n = 1098), abundance of Tregs in primary tumors was not associated with cancer subtype, American Joint Committee on Cancer (AJCC) staging, or Nottingham pathological grade (Figure 1A; p = 0.640, p = 0.172, and p = 0.963, respectively).
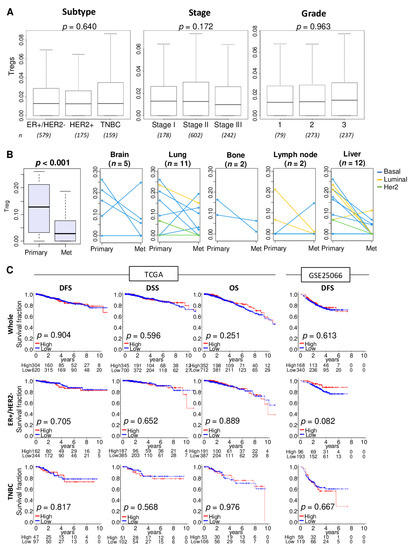
Figure 1.
Association of Tregs abundance with clinical characteristics, metastasis, and survival outcomes. (A) Boxplots of Tregs level distribution by subtype, American Joint Committee on Cancer stages, and Nottingham pathological grade in the TCGA cohort. Kruskal-Wallis test was used for the analysis. Group sizes are shown underneath the plots. (B) Boxplots of the Tregs levels in the primary breast cancer and different metastatic tumors. Matched pair comparison of Treg levels between the primary and each metastatic tumor; brain, lung, bone, lymph node, and liver in the GSE110590 cohort. Mann-Whitney U test was used to the analysis. Blue line; Basal type, yellow line; Luminal type, and green line; Her2 type. (C) Disease-free survival (DFS), disease-specific survival (DSS), and overall survival (OS) stratified by Tregs levels (low (blue line) and high (red line)) within whole tumors, estrogen receptor (ER)-positive/human epidermal growth factor receptor 2(HER2)-negative, and triple negative breast cancer (TNBC) in the TCGA and GSE25066 cohorts. The top tertile percent was used as a cut-off between high and low Tregs groups within each cohort. Log-rank test with Kaplan-Meier survival curves was used for the survival analysis.
Abundance of Tregs in various metastatic breast cancer (in brain, lung, bone, lymph node, and liver) were found to be significantly less compared with primary tumors in the GSE110590 cohort (Figure 1B; n = 83, p < 0.001). The reduction of Treg abundance defined by tumor transcriptome analysis was roughly the same for all breast cancer subtypes. Treg abundance in the metastases did not vary by site of metastasis.
To examine the association of the amount of Tregs in primary tumors and disease outcome, patients were divided into high and low Tregs groups using the top tertile as a cut-off within a cohort. Distribution of Treg was shown by histograms in the TCGA and GSE25066 cohorts (Figure S1). Abundance of Tregs in primary tumors was not associated with disease outcome as measured by disease-free survival (DFS), disease-specific survival (DSS), or overall survival (OS) in the TCGA and GSE25066 cohort (n = 508) (Figure 1C).
2.2. High Abundance of Treg Was Not Associated with Tumor Mutational Load or Enrichment of Immune Response, but Associated with Increased Infiltration of M2 Macrophages and T Helper Type 2 (Th2) Cells and Decreased Infiltration of T Helper 1 (Th1) Cells
Tumor mutational burden across several tumor types correlates with TIL infiltration and favorable response to treatment with immune check point inhibitors [31,32]. Next, we were interested to find out whether Treg infiltration is associated with tumor mutational load. To test this notion, we first examined the association of Treg infiltration and several mutation-related scores in TCGA cohort as described by Thorsson et al. [33]. High Treg infiltration was significantly associated with a high score of copy number alteration (CNA), but not silent- or nonsilent-mutation score (Figure 2A, p = 0.023, 0.155, and 0.271, respectively). Next, we examined the association of Treg infiltration and mutation rate of TP53 and PIK3CAM, which are the top two most mutated genes in breast cancer in the TCGA cohort. High Treg infiltration was associated with a high TP53 mutation rate, but not with the PIK3CAM mutation rate (Figure 2B, p = 0.037 and 0.945, respectively).
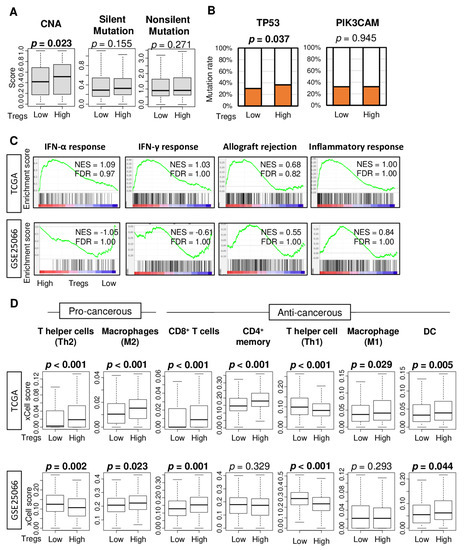
Figure 2.
Mutation, immune-related gene sets, and tumor infiltrating immune cell compositions by low and high abundance of Treg in the TCGA and GSE25066 cohorts. (A) Boxplots of comparison between low and high Treg groups with mutation-related score; copy number alteration (CNA), silent- and nonsilent-mutation score in TCGA cohort. Mann-Whitney U test was used for the analysis. (B) Bar plots comparing low and high Treg groups with TP53 and PIK3CAM genes mutation rates in the TCGA cohort. Fisher’s exact test was used for the analysis. (C) Enrichment plots for Hallmark gene sets for which highly enriched in the Treg high tumors compared to low tumors in the TCGA and GSE25066 cohorts, along with normalized enrichment score (NES) and false discovery rate (FDR). The statistical significance of GSEA was determined by FDR < 0.25. (D) Boxplots comparing low and high Treg groups with infiltrating immune cell compositions using xCell algorithm in breast cancer. Pro-cancerous immune cells included T helper type 2 cells (Th2), and M2 macrophages. Anti-cancerous immune cells included CD8+ T cells, CD4+ memory T cells, T helper type 1 cells (Th1), and M1 macrophages. The top tertile percent was used as a cut-off between low and high Treg group. Mann-Whitney U test was used for the analysis.
Given that Tregs are immunosuppressive and inhibit effector T cells proliferation [34], we also expected Treg infiltration to be associated with suppressed immune response and with less infiltration of anti-cancer immune cells in tumor immune microenvironment (TIME). To this end, gene set enrichment analysis (GSEA) of the Hallmark gene sets was performed on the TCGA and GSE25066 cohorts. Treg high abundance tumors did not enrich any of the immune-related Hallmark gene sets, such as interferon (IFN)-α response, IFN-γ response, allograft rejection, and inflammatory response, in either of the two cohorts (Figure 2C).
Next, we investigated if Treg abundance in the cohorts was associated with relative infiltration of various immune cells, which were estimated using the xCell deconvolution method on the bulk tumor transcriptome. We found that high abundance of Tregs was associated with high infiltration of M2 macrophages, which are generally considered to be immunosuppressive cells, CD8+ T cells, and dendritic cells (DC) and with less infiltration of T helper type 1 (Th1) cells in both cohorts (Figure 2D). Infiltrations of T helper type 2 (Th2) cells, CD4+ memory T cells, and M1 macrophages were also significantly higher in Treg-high tumors in either TCGA or GSE25066 cohort, but not both.
2.3. Pathological Complete Response (pCR) to Neoadjuvant Chemotherapy (NAC) Is Associated with Less Treg Abundance in Triple Negative Breast Cancer (TNBC) but Not in ER-Positive/Her2-Negative Breast Cancer
TILs have recently been reported to not only act as a prognostic indicator but also as a predictive biomarker for pCR to NAC, particularly in patients with TNBC [35]. Therefore, it was of interest to study the association of pCR to NAC with levels of immune cells in the primary tumor. The xCell algorithm was used to estimate infiltration of pro-cancerous, as well as anti-cancerous immune cells by bulk tumor transcriptome profiles from the GSE25066 (n = 508) and GSE20194 (n = 248) cohorts. pCR was not associated with high infiltration of either CD4+ or CD8+ T cells, which constitute the majority of TILs, in either ER-positive/Her2-negative or TNBC subtypes. pCR was significantly associated with high levels of Th1 and Th2 cells, and M1 macrophages in ER-positive/HER2-negative breast cancer in the GSE25066 cohort, but not the GSE20194 cohort (Figure 3A). For the TNBC subtype, the lack of Treg infiltration alone had significant association with pCR consistently in both patient cohorts (Figure 3B, both p = 0.014). These findings suggest that less abundance of Treg alone demonstrated consistent and significant association with pCR to NAC in TNBC.

Figure 3.
Association of pathological complete response (pCR) with fraction of multiple tumor infiltrating immune cells in the GSE25066 and GSE20194 cohorts. These cohorts were chosen because of their response to neoadjuvant chemotherapy data. Boxplots of levels of immune cells with residual disease (RD) and pCR after NAC in (A) ER-positive/HER2-negative tumors and (B) TNBCs in both patient cohorts. Mann–Whitney U test was used for the analysis. Pro-cancerous immune cells included Tregs, T helper type 2 cells (Th2), and M2 macrophages. Anti-cancerous immune cells included CD8 T cells, CD4 memory T cells, T helper type 1 cells (Th1), M1 macrophages, and Natural Killer T cells (NK).
2.4. High Abundance of Treg Was Associated with Significantly Worse pCR Rate after NAC in TNBC, but Not in ER-Positive/Her2-Negative Breast Cancer
Given that pCR after NAC was significantly associated with less infiltration of Tregs in TNBC, we investigated whether high top tertile amount of Treg could be a predictive biomarker for NAC. We found that high abundance of Treg prior to the treatment was associated with significantly lower pCR rate in TNBC patients consistently in both the GSE25066 and GSE20194 cohorts (Figure 4. p = 0.009, and 0.034, respectively). No association was observed between Treg and pCR rate in ER-positive/HER2-negative patients in both cohorts. These results suggest that high abundance of Treg in TNBC may be a predictive biomarker of pCR after NAC.
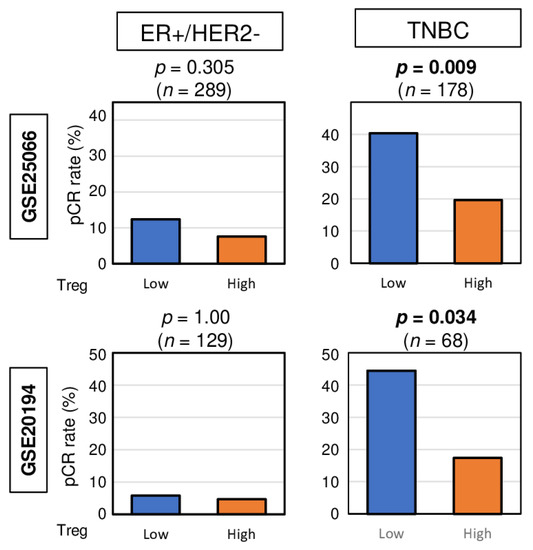
Figure 4.
Bar plots depicting the pCR rates between low and high abundance of Treg groups among patients with ER-positive/HER2-negative tumors and TNBCs in the GSE25066 (n = 508) and GSE20194 (n = 248) cohorts. These cohorts were chosen because of their response to neoadjuvant chemotherapy data. Fisher’s exact test was used for the analysis.
2.5. Tumors with High Abundance of Treg Are Associated with High Expressions of Immune Checkpoint Molecules
Given the recent approval of immune checkpoint inhibitors (ICIs) as treatment for some breast cancer patients [36], it was of interest to examine the association of abundance of Tregs with expression of major immune checkpoint molecules. We found that high abundance of Treg had significantly elevated gene expression of programmed death ligand 1 (PD-L1) and cytotoxic T-lymphocyte-associated protein 4 (CTLA4) in ER-positive/HER2-negative tumors, but it had significantly elevated levels of PD-L1, PD-L2, and CTLA4 in TNBC in the TCGA cohort (Figure 5). In the GSE96058 cohort, tumors of either subtype with high abundance of Tregs had significantly elevated expression of all the immune checkpoint genes examined. These findings suggest that abundance of Tregs is associated with expression of immune checkpoint molecules.
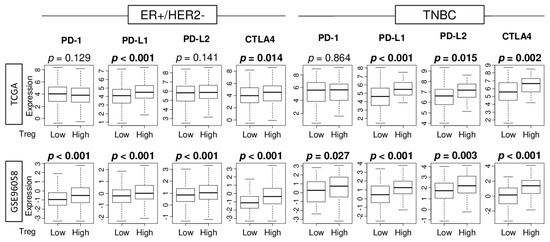
Figure 5.
Association of Treg infiltration with expression of immune checkpoint molecules. Boxplots comparing low and high infiltration of Tregs in gene expression of immune checkpoint molecules in ER-positive/HER2-negative and TNBC patients in the TCGA and GSE96058 cohorts which have the PD-L1 (CD274) expression data. Mann-Whitney U test was used for the analysis.
3. Discussion
In this study, we investigated the clinical relevance of Treg abundance in breast cancer by estimating Treg levels using the xCell algorithm on tumor transcriptomes. There was no association of Treg abundance with cancer aggressiveness, patient survival, or immune-related pathways within tumors. Abundance of Treg in metastatic tumor was lower than that in primary tumors. High abundance of Tregs was significantly associated with high mutation rate of TP53 gene and copy number mutation score. Treg abundance was significantly associated with increased tumor infiltration of M2 macrophages, CD8+ T cells, and DC and decreased infiltration of Th1 cells. Interestingly, pCR after NAC was significantly associated with low abundance of Tregs in TNBC, but not in ER-positive/HER2-negative breast cancer. No other TIL subset besides Treg was found to be associated with pCR after NAC. High abundance of Tregs significantly associated with low pCR rate consistently in two independent patient cohorts. In addition, high Treg abundance was significantly associated with high expression of multiple immune checkpoint molecules in the bulk tumors.
TILs constitute the tumor immune surveillance system. Although breast cancer is immunologically less active compared to other solid tumors, there are data suggesting TILs can predicate response to therapy and survival in breast cancer [35,37]. Recently it has been shown that not only does the quantity of TILs determine the clinical outcome, but also the types of lymphocytes [38,39,40]. For instance, the role tumoral CD4+ T cells, a major component of TILs, is multifaceted. CD4+ T cells that differentiated into Th1 cells have been shown to sustain the cancer cytotoxic functions of CD8+ T cells. On the other hand, Treg subset of CD4+ T cells have been shown to inhibit cytotoxic functions of CD8+T cells, support B cell growth, and promote cancer progression [41].
Due to the ability to inhibit anti-cancerous immunity, a high abundance of tumor-infiltrating Tregs were associated with worse prognosis [42,43,44,45,46]. Wang et al. have shown that high infiltration of Treg correlated with worse DFS, suggesting that Treg abundance may be a prognostic biomarker for breast cancer [18]. However, others have challenged this notion by either showing a lack of correlation of Treg abundance with survival [47] or by showing that abundance of Treg was associated with better survival in some cancers [48], including TNBC [49,50,51]. For instance, Treg identified by IHC was associated with the efficacy of NAC in breast cancer patients [52]. Furthermore, our group and others have found that abundance of Treg in metastatic tumor was lower than that in primary tumors [53,54], which appears paradoxical considering metastatic tumors are far more aggressive than the primary cancer. One of the reasons for these results may be due to the method of identifying Tregs. The most common method to identify Tregs is to stain Forkhead box protein 3 (Foxp3) by IHC [55,56]. The issue is that Foxp3-positive T cells are heterogeneous population of cells, and Foxp3 alone is not sufficient to capture Tregs. For instance, when Tregs were identified by Foxp3 alone, there was no association of Tregs and survival outcome of colorectal cancer. On the other hand, a positive association with patient outcome was found when Foxp3 and B lymphocyte-induced maturation protein-1 (Blimp-1) were both used to identify Tregs [17,57]. Another example is that Tregs were associated with good outcome in B-cell lymphoma when they were identified by Foxp3 alone, but Tregs were associated with poor patient outcome when they were identified by both CTLA-4 and Foxp3 [57]. Zhang et al. reported that staining with Foxp3 together with CD25 is better than Foxp3 alone to evaluate Tregs [58]. These examples eloquently demonstrate that multiple markers are necessary to identify biologically active Tregs in human cancer [17,18,19]. Our group has previously reported the clinical relevance of specific cells in the TIME by using multiple gene expression profiles with algorithms [59,60,61,62]. This approach was feasible and efficient in analyzing large patient cohorts because we utilized transcriptome of publicly available large cohorts that are generated for different research objectives. The gold standard to analyze immune cells is IHC or flow cytometry. Nonetheless, IHC is limited by its ability to only quantify cells and user dependent variation of results. Flow cytometry requires fresh sample, which is usually very challenging to obtain. Moreover, both methods are highly costly and labor intensive compared to our in silico approach. We believe that current study can be one of the breakthroughs of the field since we estimated the relative abundance of Tregs in breast cancer by using the xCell algorithm to analyze gene expression data of bulk tumors of multiple large patient cohorts. In addition to Tregs, the xCell algorithm can be used to infer the relative abundance of 63 other types of immune and stromal cells using the known gene signatures of each cell types [30]. We further found that, although the amount of Tregs was not associated with survival outcome, pCR after NAC was significantly associated with Tregs and not with any other immune cells in TNBC. Furthermore, the Treg abundance showed a potential as a predictive biomarker for NAC response in TNBC. No other immune cells showed similar results in the two independent cohorts. One of the reasons may be due to differences in NAC treatment regimens between cohorts. Taxane and anthracycline regimen was used in the GSE25066 and paclitaxel, 5-fluorouracil, cyclophosphamide and doxorubicin regimen was used in the GSE20194 cohort. Further verification is required to conclude the association between immune cells and treatment response.
Because each breast cancer subtypes is biologically different, the impact of TILs on clinical outcome should be evaluated separately in each subtype [63]. TNBC is known to be both the most aggressive breast cancer subtype as well as the most “immunomodulatory subtype” because it has elevated immune cell infiltration. Although ICIs are approved for TNBC, it is effective to very limited population and the patient selection remains a major challenge. Given that our results show that the majority of immune checkpoint molecules were associated with abundance of Tregs not only in TNBC but also in ER-positive/HER2-negative breast cancer, we cannot help but speculate that patients whose tumors show Treg abundance could be the population who would respond to ICIs.
This study has some limitations. The biggest limitation is that our results are based on analyses of tumor gene expression alone without any direct quantification of tumoral Tregs using gold standard such as flow cytometry and/or immunohistochemistry, which are analyses in protein level. Our approach allowed us to analyze very large sample number cohorts; however, we were unable to validate our results at protein level because we do not have physical access to the samples of these cohorts. To this end, further studies using protein levels are needed to investigate the clinical applicability of our findings in the future. This is a retrospective study that used publicly available cohorts of multiple previous studies. The cohorts we examined vary between each other by clinical characteristics. Additionally, data regarding co-morbidities and therapeutic information were lacking. Furthermore, our analyses are limited to exact spatial location of where the sample was taken by the original authors. For breast cancer, TILs can be divided into intratumoral and stromal subtypes according to their localization in the tumor tissue. The International TILs Working Group recommended the use of stromal TILs to assess the significance of TILs in breast cancer [64], whereas our samples are more likely to be taken from inside the tumor. Recently, some claim that effector T cell/Treg ratio is important for survival and drug response; however, we were unable to conduct this analysis due to technical difficulty. Finally, this study did not assess biological mechanisms that underlie its clinical findings.
4. Materials and Methods
4.1. Data of Breast Cancer Cohorts
Data for tumor gene expression and corresponding clinical information for the TCGA-BRCA (n = 1065) [26] cohort were obtained from the cBio Cancer Genomic portal [65]. Mutation data (TP53 and PIK3CAM) of the TCGA cohort were also obtained from the cBio Cancer Genomic portal, as we previously reported [66]. TCGA was chosen as the main cohort for our analyses because it is one of the largest breast cancer cohorts with robust tumor transcriptome and clinical data. However, one of the weaknesses of the TCGA is that the treatment data are limited, and it is assumed that all the patient underwent “standard of care”. The Gene Expression Omnibus (GEO) repository was used to access the tumor gene expression and clinical data from the Brueffer et al. (GSE96058; n = 3273) [27], Symmans et al. (GSE25066; n = 508) [28], Shi et al. (GSE20194; n = 248) [29], and Siegel et al. (GSE110590; n = 83) [67] cohorts. GSE25066 was chosen as the validating cohort of TCGA cohort; however, it needs to be noted that the sample size of GSE25066 is about a half of TCGA. GSE25066 and GSE20194 cohorts were chosen because these two cohorts were among the largest cohorts with patient NAC response data that we identified. GSE110590 cohort was used to investigate the association of Treg infiltration with metastases because it has transcriptomic data for metastatic breast cancer, of which was not available in any other cohorts. Finally, we used the GSE110590 cohort to investigate the association of Treg infiltration with expression of multiple immune checkpoint molecules because in-depth coverage of transcriptome was available in this cohort. This information is shown in Table S3. We believe that having multiple completely independently reported large cohorts support our hypothesis, making our argument even stronger. The average of the signals from the multiple probes was used to analyze genes with multiple hybridization probes in microarray-based gene expression data. Gene expression data were transformed for log2 in all analyses. The top tertile was defined as the high Treg infiltration within cohorts.
4.2. Tumor Immune Cell Scoring Using xCell
The tumor Tregs score was determined with the xCell algorithm [30], using bulk tumor gene expression profiles. The xCell algorithm was used for tumor composition analysis of infiltrated immune cells, as we previously reported [68,69,70]. Genes used to calculate each immune cell are shown in Tables S1 and S2.
4.3. Gene Set Enrichment Analysis
Gene set enrichment analysis were performed with Gene Set Enrichment Analyses (GSEA) software (Java version 4.0) [71,72] with MSigDB Hallmark gene sets [73]. Statistical significance was set with a false discovery rate (FDR) of 0.25, as recommended by the GSEA software and as we previously reported [66,74,75,76,77,78,79,80].
4.4. Other
R software (version 4.0.1, R Project for Statistical Computing) and Excel (version 16 for Windows; Microsoft, Redmond, WA, USA) were used for data analyses. Within patient cohorts, those in the top tertile for tumor Tregs score were considered to be in the high Tregs group, while the other were grouped into the low Tregs group. Gene expression data were transformed for log2 in all analyses. Kruskal-Wallis test was used to compare multiple groups. Mann-Whitney U test and Fisher’s exact tests were used to compare two groups. Survival analyses were performed using the Kaplan–Meier method with log-rank test.
5. Conclusions
pCR after NAC was significantly associated with low Treg abundance in TNBC, but not in ER-positive/HER2-negative subtype. Thus, we conclude that Treg abundance may have a potential as a predictive biomarker of pCR after NAC in TNBC.
Supplementary Materials
The following are available online at https://www.mdpi.com/2072-6694/12/10/3038/s1, Table S1: Genes used to calculate the regulatory T cells (Tregs) score in the xCell method. Table S2: Genes used to calculate each immune cell, including CD8+ T cell, Cd4+ memory T cell, T helper type 1 and type 2 cell, M1 and M2 macrophages, dendritic cell (DC), and NK cell. Table S3: Available data of each cohort. Figure S1: Histogram of Treg in TCGA and GSE25066 cohorts.
Author Contributions
Conceptualization, I.E., K.T., M.O., R.M. and T.I.; Methodology, K.T., M.A., M.O. and Y.T.; Formal Analysis, M.O.; Writing—Original Draft Preparation, M.O.; Writing—Review and Editing, F.A.A., K.T., L.Y. and E.Z.; Supervision, K.T.; and Project Administration, K.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institutes of Health, USA grant R01CA160688; a grant from the Edward K. Duch Foundation to K.T.; Grant-in-Aid for Scientific Research Grant Numbers 19H03714 and 18K19576 to M.N.; and National Cancer Institute, USA cancer center support grant P30-CA016056 to Roswell Park Comprehensive Cancer Center (RPCCC).
Acknowledgments
We thank Santosh Patnaik of RPCCC for feedback.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AJCC | American Joint Committee on Cancer |
| DFS | disease-free survival |
| DSS | disease-specific survival |
| ER | estrogen receptor |
| FDR | false discovery rate |
| HER2 | human epidermal growth factor receptor 2 |
| NES | normalized enrichment score |
| OS | overall survival |
| pCR | pathological complete response |
| TCGA | The Cancer Genome Atlas |
| TNBC | triple negative breast cancer |
References
- Pagès, F.; Galon, J.; Dieu-Nosjean, M.C.; Tartour, E.; Sautès-Fridman, C.; Fridman, W.H. Immune infiltration in human tumors: A prognostic factor that should not be ignored. Oncogene 2010, 29, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Kawaguchi, T.; Yan, L.; Peng, X.; Qi, Q.; Takabe, K. Cytolytic Activity Score to Assess Anticancer Immunity in Colorectal Cancer. Ann. Surg. Oncol. 2018, 25, 2323–2331. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Kawaguchi, T.; Peng, X.; Qi, Q.; Liu, S.; Yan, L.; Takabe, K. Tumor Infiltrating Lymphocytes and Macrophages Improve Survival in Microsatellite Unstable Colorectal Cancer. Sci. Rep. 2019, 9, 13455. [Google Scholar] [CrossRef]
- Foulkes, W.D.; Smith, I.E.; Reis-Filho, J.S. Triple-negative breast cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef]
- Kwa, M.J.; Adams, S. Checkpoint inhibitors in triple-negative breast cancer (TNBC): Where to go from here. Cancer 2018, 124, 2086–2103. [Google Scholar] [CrossRef] [PubMed]
- Disis, M.L.; Stanton, S.E. Triple-negative breast cancer: Immune modulation as the new treatment paradigm. Am. Soc. Clin. Oncol. Educ. Book Am. Soc. Clin. Oncol. Annu. Meet. 2015, 35, e25–e30. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Tsuda, H.; Shimizu, C.; Yamamoto, S.; Shibata, T.; Yamamoto, H.; Hirata, T.; Yonemori, K.; Ando, M.; Tamura, K.; et al. Tumor-infiltrating lymphocytes are correlated with response to neoadjuvant chemotherapy in triple-negative breast cancer. Breast Cancer Res. Treat. 2012, 132, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Denkert, C.; von Minckwitz, G.; Brase, J.C.; Sinn, B.V.; Gade, S.; Kronenwett, R.; Pfitzner, B.M.; Salat, C.; Loi, S.; Schmitt, W.D.; et al. Tumor-infiltrating lymphocytes and response to neoadjuvant chemotherapy with or without carboplatin in human epidermal growth factor receptor 2-positive and triple-negative primary breast cancers. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2015, 33, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Hamy, A.S.; Pierga, J.Y.; Sabaila, A.; Laas, E.; Bonsang-Kitzis, H.; Laurent, C.; Vincent-Salomon, A.; Cottu, P.; Lerebours, F.; Rouzier, R.; et al. Stromal lymphocyte infiltration after neoadjuvant chemotherapy is associated with aggressive residual disease and lower disease-free survival in HER2-positive breast cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2017, 28, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Lang, R.; Zhao, J.; Zhang, X.; Pringle, G.A.; Fan, Y.; Yin, D.; Gu, F.; Yao, Z.; Fu, L. CD8+ cytotoxic T cell and FOXP3+ regulatory T cell infiltration in relation to breast cancer survival and molecular subtypes. Breast Cancer Res. Treat. 2011, 130, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Miyara, M.; Sakaguchi, S. Natural regulatory T cells: Mechanisms of suppression. Trends Mol. Med. 2007, 13, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Tanchot, C.; Terme, M.; Pere, H.; Tran, T.; Benhamouda, N.; Strioga, M.; Banissi, C.; Galluzzi, L.; Kroemer, G.; Tartour, E. Tumor-infiltrating regulatory T cells: Phenotype, role, mechanism of expansion in situ and clinical significance. Cancer Microenviron. Off. J. Int. Cancer Microenviron. Soc. 2013, 6, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.A.; Oda, J.M.; Amarante, M.K.; Cesar Voltarelli, J. Regulatory T cells and breast cancer: Implications for immunopathogenesis. Cancer Metastasis Rev. 2010, 29, 569–579. [Google Scholar] [CrossRef]
- Kryczek, I.; Liu, R.; Wang, G.; Wu, K.; Shu, X.; Szeliga, W.; Vatan, L.; Finlayson, E.; Huang, E.; Simeone, D.; et al. FOXP3 defines regulatory T cells in human tumor and autoimmune disease. Cancer Res. 2009, 69, 3995–4000. [Google Scholar] [CrossRef] [PubMed]
- DiPaolo, R.J.; Glass, D.D.; Bijwaard, K.E.; Shevach, E.M. CD4+CD25+ T cells prevent the development of organ-specific autoimmune disease by inhibiting the differentiation of autoreactive effector T cells. J. Immunol. 2005, 175, 7135–7142. [Google Scholar] [CrossRef]
- Vignali, D.A.; Collison, L.W.; Workman, C.J. How regulatory T cells work. Nat. Rev. Immunol. 2008, 8, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Ward-Hartstonge, K.A.; McCall, J.L.; McCulloch, T.R.; Kamps, A.K.; Girardin, A.; Cretney, E.; Munro, F.M.; Kemp, R.A. Inclusion of BLIMP-1(+) effector regulatory T cells improves the Immunoscore in a cohort of New Zealand colorectal cancer patients: A pilot study. Cancer Immunol. Immunother. 2017, 66, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Ward-Hartstonge, K.A.; Kemp, R.A. Regulatory T-cell heterogeneity and the cancer immune response. Clin. Transl. Immunol. 2017, 6, e154. [Google Scholar] [CrossRef]
- Liu, X.; Wang, X.; Ding, J.; Gao, Y.; Zhao, Y.; Zhao, R.; Sun, Q.; Zhang, S. FOXP3 and CD25 double staining antibody cocktails identify regulatory T cells in different types of tumor tissues using tissue microarrays. Ann. Diagn. Pathol. 2019, 38, 67–70. [Google Scholar] [CrossRef]
- Katsuta, E.; Maawy, A.A.; Yan, L.; Takabe, K. High expression of bone morphogenetic protein (BMP) 6 and BMP7 are associated with higher immune cell infiltration and better survival in estrogen receptor-positive breast cancer. Oncol. Rep. 2019, 42, 1413–1421. [Google Scholar] [CrossRef]
- Asaoka, M.; Ishikawa, T.; Takabe, K.; Patnaik, S.K. APOBEC3-Mediated RNA Editing in Breast Cancer is Associated with Heightened Immune Activity and Improved Survival. Int. J. Mol. Sci. 2019, 20, 5621. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Asaoka, M.; Yan, L.; Rashid, O.M.; Oshi, M.; Ishikawa, T.; Nagahashi, M.; Takabe, K. Biologically Aggressive Phenotype and Anti-cancer Immunity Counterbalance in Breast Cancer with High Mutation Rate. Sci. Rep. 2020, 10, 1852. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, T.; Yan, L.; Asaoka, M.; Rashid, O.; Takabe, K. Late recurrence of breast cancer is associated with pro-cancerous immune microenvironment in the primary tumor. Sci. Rep. 2019, 9, 16942. [Google Scholar] [CrossRef]
- Takahashi, H.; Kawaguchi, T.; Yan, L.; Peng, X.; Qi, Q.; Morris, L.G.T.; Chan, T.A.; Tsung, A.; Otsuji, E.; Takabe, K. Immune Cytolytic Activity for Comprehensive Understanding of Immune Landscape in Hepatocellular Carcinoma. Cancers 2020, 12, 1221. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Oshi, M.; Katsuta, E.; Yan, L.; Huang, J.L.; Nagahashi, M.; Matsuhashi, N.; Futamura, M.; Yoshida, K.; Takabe, K. Intratumoral Adipocyte-High Breast Cancer Enrich for Metastatic and Inflammation-Related Pathways but Associated with Less Cancer Cell Proliferation. Int. J. Mol. Sci. 2020, 21, 5744. [Google Scholar] [CrossRef]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416. [Google Scholar] [CrossRef] [PubMed]
- Brueffer, C.; Vallon-Christersson, J.; Grabau, D.; Ehinger, A.; Häkkinen, J.; Hegardt, C.; Malina, J.; Chen, Y.; Bendahl, P.O.; Manjer, J.; et al. Clinical Value of RNA Sequencing-Based Classifiers for Prediction of the Five Conventional Breast Cancer Biomarkers: A Report from the Population-Based Multicenter Sweden Cancerome Analysis Network—Breast Initiative. JCO Precis. Oncol. 2018, 2, 1–18. [Google Scholar] [CrossRef]
- Itoh, M.; Iwamoto, T.; Matsuoka, J.; Nogami, T.; Motoki, T.; Shien, T.; Taira, N.; Niikura, N.; Hayashi, N.; Ohtani, S.; et al. Estrogen receptor (ER) mRNA expression and molecular subtype distribution in ER-negative/progesterone receptor-positive breast cancers. Breast Cancer Res. Treat. 2014, 143, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Campbell, G.; Jones, W.D.; Campagne, F.; Wen, Z.; Walker, S.J.; Su, Z.; Chu, T.M.; Goodsaid, F.M.; Pusztai, L.; et al. The MicroArray Quality Control (MAQC)-II study of common practices for the development and validation of microarray-based predictive models. Nat. Biotechnol. 2010, 28, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Aran, D.; Hu, Z.; Butte, A.J. xCell: Digitally portraying the tissue cellular heterogeneity landscape. Genome Biol. 2017, 18, 220. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.A.; Yarchoan, M.; Jaffee, E.; Swanton, C.; Quezada, S.A.; Stenzinger, A.; Peters, S. Development of tumor mutation burden as an immunotherapy biomarker: Utility for the oncology clinic. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2019, 30, 44–56. [Google Scholar] [CrossRef]
- Samstein, R.M.; Lee, C.H.; Shoushtari, A.N.; Hellmann, M.D.; Shen, R.; Janjigian, Y.Y.; Barron, D.A.; Zehir, A.; Jordan, E.J.; Omuro, A.; et al. Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat. Genet. 2019, 51, 202–206. [Google Scholar] [CrossRef]
- Thorsson, V.; Gibbs, D.L.; Brown, S.D.; Wolf, D.; Bortone, D.S.; Ou Yang, T.H.; Porta-Pardo, E.; Gao, G.F.; Plaisier, C.L.; Eddy, J.A.; et al. The Immune Landscape of Cancer. Immunity 2019, 51, 411–412. [Google Scholar] [CrossRef]
- Bettelli, E.; Carrier, Y.; Gao, W.; Korn, T.; Strom, T.B.; Oukka, M.; Weiner, H.L.; Kuchroo, V.K. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 2006, 441, 235–238. [Google Scholar] [CrossRef]
- Loi, S.; Michiels, S.; Salgado, R.; Sirtaine, N.; Jose, V.; Fumagalli, D.; Kellokumpu-Lehtinen, P.L.; Bono, P.; Kataja, V.; Desmedt, C.; et al. Tumor infiltrating lymphocytes are prognostic in triple negative breast cancer and predictive for trastuzumab benefit in early breast cancer: Results from the FinHER trial. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2014, 25, 1544–1550. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Joyce, D.; Takabe, K. Current status and limitations of immunotherapy for breast cancer. Surgery 2020, 167, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, E.M.; Al-Foheidi, M.E.; Al-Mansour, M.M.; Kazkaz, G.A. The prognostic value of tumor-infiltrating lymphocytes in triple-negative breast cancer: A meta-analysis. Breast Cancer Res. Treat. 2014, 148, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Bos, R.; Sherman, L.A. CD4+ T-cell help in the tumor milieu is required for recruitment and cytolytic function of CD8+ T lymphocytes. Cancer Res. 2010, 70, 8368–8377. [Google Scholar] [CrossRef] [PubMed]
- Bos, R.; Marquardt, K.L.; Cheung, J.; Sherman, L.A. Functional differences between low- and high-affinity CD8(+) T cells in the tumor environment. Oncoimmunology 2012, 1, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.J.; Tonlaar, N.Y.; Garwood, E.R.; Huo, D.; Moore, D.H.; Khramtsov, A.I.; Au, A.; Baehner, F.; Chen, Y.; Malaka, D.O.; et al. Proliferating macrophages associated with high grade, hormone receptor negative breast cancer and poor clinical outcome. Breast Cancer Res. Treat. 2011, 128, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.H.; Goh, S.Y.; Wong, S.C.; Lam, K.P. T helper cell-specific regulation of inducible costimulator expression via distinct mechanisms mediated by T-bet and GATA-3. J. Biol. Chem. 2008, 283, 128–136. [Google Scholar] [CrossRef]
- deLeeuw, R.J.; Kost, S.E.; Kakal, J.A.; Nelson, B.H. The prognostic value of FoxP3+ tumor-infiltrating lymphocytes in cancer: A critical review of the literature. Clin. Cancer Res. 2012, 18, 3022–3029. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, M.; Sasano, H.; Tamaki, K.; Hirakawa, H.; Takahashi, Y.; Nakagawa, S.; Watanabe, G.; Tada, H.; Suzuki, A.; Ohuchi, N.; et al. Prognostic significance of tumor-infiltrating CD8+ and FOXP3+ lymphocytes in residual tumors and alterations in these parameters after neoadjuvant chemotherapy in triple-negative breast cancer: A retrospective multicenter study. Breast Cancer Res. 2015, 17, 124. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.; Seki, N.; Toh, U.; Hattori, S.; Kawahara, A.; Yamaguchi, T.; Koura, K.; Takahashi, R.; Otsuka, H.; Takahashi, H.; et al. FOXP3 expression in tumor cells and tumor-infiltrating lymphocytes is associated with breast cancer prognosis. Mol. Clin. Oncol. 2013, 1, 625–632. [Google Scholar] [CrossRef]
- Kim, S.; Lee, A.; Lim, W.; Park, S.; Cho, M.S.; Koo, H.; Moon, B.I.; Sung, S.H. Zonal difference and prognostic significance of foxp3 regulatory T cell infiltration in breast cancer. J. Breast Cancer 2014, 17, 8–17. [Google Scholar] [CrossRef]
- Merlo, A.; Casalini, P.; Carcangiu, M.L.; Malventano, C.; Triulzi, T.; Mènard, S.; Tagliabue, E.; Balsari, A. FOXP3 expression and overall survival in breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2009, 27, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.M.; Paish, E.C.; Powe, D.G.; Macmillan, R.D.; Lee, A.H.; Ellis, I.O.; Green, A.R. An evaluation of the clinical significance of FOXP3+ infiltrating cells in human breast cancer. Breast Cancer Res. Treat. 2011, 127, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Correale, P.; Rotundo, M.S.; Del Vecchio, M.T.; Remondo, C.; Migali, C.; Ginanneschi, C.; Tsang, K.Y.; Licchetta, A.; Mannucci, S.; Loiacono, L.; et al. Regulatory (FoxP3+) T-cell tumor infiltration is a favorable prognostic factor in advanced colon cancer patients undergoing chemo or chemoimmunotherapy. J. Immunother. 2010, 33, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Yeong, J.; Thike, A.A.; Lim, J.C.; Lee, B.; Li, H.; Wong, S.C.; Hue, S.S.; Tan, P.H.; Iqbal, J. Higher densities of Foxp3(+) regulatory T cells are associated with better prognosis in triple-negative breast cancer. Breast Cancer Res. Treat. 2017, 163, 21–35. [Google Scholar] [CrossRef]
- West, N.R.; Kost, S.E.; Martin, S.D.; Milne, K.; Deleeuw, R.J.; Nelson, B.H.; Watson, P.H. Tumour-infiltrating FOXP3(+) lymphocytes are associated with cytotoxic immune responses and good clinical outcome in oestrogen receptor-negative breast cancer. Br. J. Cancer 2013, 108, 155–162. [Google Scholar] [CrossRef]
- Lee, S.; Cho, E.Y.; Park, Y.H.; Ahn, J.S.; Im, Y.H. Prognostic impact of FOXP3 expression in triple-negative breast cancer. Acta Oncol. 2013, 52, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiang, Q.; Yu, L.; Hu, T.; Chen, Y.; Wang, J.; Nie, X.; Cheng, J. Changes in Tumor-Infiltrating Lymphocytes and Vascular Normalization in Breast Cancer Patients after Neoadjuvant Chemotherapy and Their Correlations with DFS. Front. Oncol. 2019, 9, 1545. [Google Scholar] [CrossRef] [PubMed]
- Ogiya, R.; Niikura, N.; Kumaki, N.; Bianchini, G.; Kitano, S.; Iwamoto, T.; Hayashi, N.; Yokoyama, K.; Oshitanai, R.; Terao, M.; et al. Comparison of tumor-infiltrating lymphocytes between primary and metastatic tumors in breast cancer patients. Cancer Sci. 2016, 107, 1730–1735. [Google Scholar] [CrossRef]
- Szekely, B.; Bossuyt, V.; Li, X.; Wali, V.B.; Patwardhan, G.A.; Frederick, C.; Silber, A.; Park, T.; Harigopal, M.; Pelekanou, V.; et al. Immunological differences between primary and metastatic breast cancer. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2018, 29, 2232–2239. [Google Scholar] [CrossRef] [PubMed]
- Beyer, M.; Schultze, J.L. Regulatory T cells in cancer. Blood 2006, 108, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef]
- Nakayama, S.; Yokote, T.; Akioka, T.; Hiraoka, N.; Nishiwaki, U.; Miyoshi, T.; Iwaki, K.; Takayama, A.; Masuda, Y.; Hatooka, J.; et al. Infiltration of effector regulatory T cells predicts poor prognosis of diffuse large B-cell lymphoma, not otherwise specified. Blood Adv. 2017, 1, 486–493. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.I.; Ding, J.; Sun, Q.; Zhang, S. The predictive and prognostic value of Foxp3+/CD25+ regulatory T cells and PD-L1 expression in triple negative breast cancer. Ann. Diagn. Pathol. 2019, 40, 143–151. [Google Scholar] [CrossRef]
- Oshi, M.; Takahashi, H.; Tokumaru, Y.; Yan, L.; Rashid, O.M.; Matsuyama, R.; Endo, I.; Takabe, K. G2M Cell Cycle Pathway Score as a Prognostic Biomarker of Metastasis in Estrogen Receptor (ER)-Positive Breast Cancer. Int. J. Mol. Sci. 2020, 21, 2921. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Oshi, M.; Katsuta, E.; Yan, L.; Satyananda, V.; Matsuhashi, N.; Futamura, M.; Akao, Y.; Yoshida, K.; Takabe, K. KRAS signaling enriched triple negative breast cancer is associated with favorable tumor immune microenvironment and better survival. Am. J. Cancer Res. 2020, 10, 897–907. [Google Scholar]
- Oshi, M.; Katsuta, E.; Yan, L.; Ebos, J.M.L.; Rashid, O.M.; Matsuyama, R.; Endo, I.; Takabe, K. A Novel 4-Gene Score to Predict Survival, Distant Metastasis and Response to Neoadjuvant Therapy in Breast Cancer. Cancers 2020, 12, 1148. [Google Scholar] [CrossRef] [PubMed]
- Oshi, M.; Takahashi, H.; Tokumaru, Y.; Yan, L.; Rashid, O.M.; Nagahashi, M.; Matsuyama, R.; Endo, I.; Takabe, K. The E2F Pathway Score as a Predictive Biomarker of Response to Neoadjuvant Therapy in ER+/HER2- Breast Cancer. Cells 2020, 9, 1643. [Google Scholar] [CrossRef]
- Stanton, S.E.; Disis, M.L. Clinical significance of tumor-infiltrating lymphocytes in breast cancer. J. Immunother. Cancer 2016, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Van den Eynden, G.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Takeshita, T.; Asaoka, M.; Katsuta, E.; Photiadis, S.J.; Narayanan, S.; Yan, L.; Takabe, K. High expression of polo-like kinase 1 is associated with TP53 inactivation, DNA repair deficiency, and worse prognosis in ER positive Her2 negative breast cancer. Am. J. Transl. Res. 2019, 11, 6507–6521. [Google Scholar]
- Siegel, M.B.; He, X.; Hoadley, K.A.; Hoyle, A.; Pearce, J.B.; Garrett, A.L.; Kumar, S.; Moylan, V.J.; Brady, C.M.; Van Swearingen, A.E.; et al. Integrated RNA and DNA sequencing reveals early drivers of metastatic breast cancer. J. Clin. Investig. 2018, 128, 1371–1383. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Katsuta, E.; Oshi, M.; Sporn, J.C.; Yan, L.; Le, L.; Matsuhashi, N.; Futamura, M.; Akao, Y.; Yoshida, K.; et al. High Expression of miR-34a Associated with Less Aggressive Cancer Biology but Not with Survival in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 3045. [Google Scholar] [CrossRef]
- Tokumaru, Y.; Asaoka, M.; Oshi, M.; Katsuta, E.; Yan, L.; Narayanan, S.; Sugito, N.; Matsuhashi, N.; Futamura, M.; Akao, Y.; et al. High Expression of microRNA-143 is Associated with Favorable Tumor Immune Microenvironment and Better Survival in Estrogen Receptor Positive Breast Cancer. Int. J. Mol. Sci. 2020, 21, 3213. [Google Scholar] [CrossRef]
- Takahashi, H.; Oshi, M.; Asaoka, M.; Yan, L.; Endo, I.; Takabe, K. Molecular Biological Features of Nottingham Histological Grade 3 Breast Cancers. Ann. Surg. Oncol. 2020, 27, 4475–4485. [Google Scholar] [CrossRef]
- Bild, A.; Febbo, P.G. Application of a priori established gene sets to discover biologically important differential expression in microarray data. Proc. Natl. Acad. Sci. USA 2005, 102, 15278–15279. [Google Scholar] [CrossRef] [PubMed]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Oshi, M.; Butash, A.L.; Asaoka, M.; Katsuta, E.; Peng, X.; Qi, Q.; Yan, L.; Takabe, K. Estrogen Receptor Positive Breast Cancer with High Expression of Androgen Receptor has Less Cytolytic Activity and Worse Response to Neoadjuvant Chemotherapy but Better Survival. Int. J. Mol. Sci. 2019, 20, 2655. [Google Scholar] [CrossRef]
- Okano, M.; Oshi, M.; Butash, A.L.; Katsuta, E.; Tachibana, K.; Saito, K.; Okayama, H.; Peng, X.; Yan, L.; Kono, K.; et al. Triple-Negative Breast Cancer with High Levels of Annexin A1 Expression Is Associated with Mast Cell Infiltration, Inflammation, and Angiogenesis. Int. J. Mol. Sci. 2019, 20, 4197. [Google Scholar] [CrossRef] [PubMed]
- Katsuta, E.; Yan, L.; Takeshita, T.; McDonald, K.A.; Dasgupta, S.; Opyrchal, M.; Takabe, K. High MYC mRNA expression is more clinically relevant than MYC DNA amplification in triple-negative breast cancer. Int. J. Mol. Sci. 2019, 21, 217. [Google Scholar] [CrossRef]
- Asaoka, M.; Patnaik, S.K.; Zhang, F.; Ishikawa, T.; Takabe, K. Lymphovascular invasion in breast cancer is associated with gene expression signatures of cell proliferation but not lymphangiogenesis or immune response. Breast Cancer Res. Treat. 2020, 181, 309–322. [Google Scholar] [CrossRef]
- Gandhi, S.; Elkhanany, A.; Oshi, M.; Dai, T.; Opyrchal, M.; Mohammadpour, H.; Repasky, E.A.; Takabe, K. Contribution of Immune Cells to Glucocorticoid Receptor Expression in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 4635. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Oshi, M.; Asaoka, M.; Ishikawa, T.; Endo, I.; Takabe, K. ASO Author Reflections: Transitioning from Morphology to Transcriptomics in Capturing Tumor Biology. Ann. Surg. Oncol. 2020, 27, 4486–4487. [Google Scholar] [CrossRef]
- Takahashi, H.; Katsuta, E.; Yan, L.; Tokumaru, Y.; Katz, M.H.G.; Takabe, K. Transcriptomic Profile of Lymphovascular Invasion, a Known Risk Factor of Pancreatic Ductal Adenocarcinoma Metastasis. Cancers 2020, 12, 2033. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).