Function and Mechanism of Antiviral Wasp Venom Peptide Protopolybia-MP III and Its Derivatives against HSV-1

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Protopolybia-MP III Strongly Inhibits HSV-1 Replication In Vitro
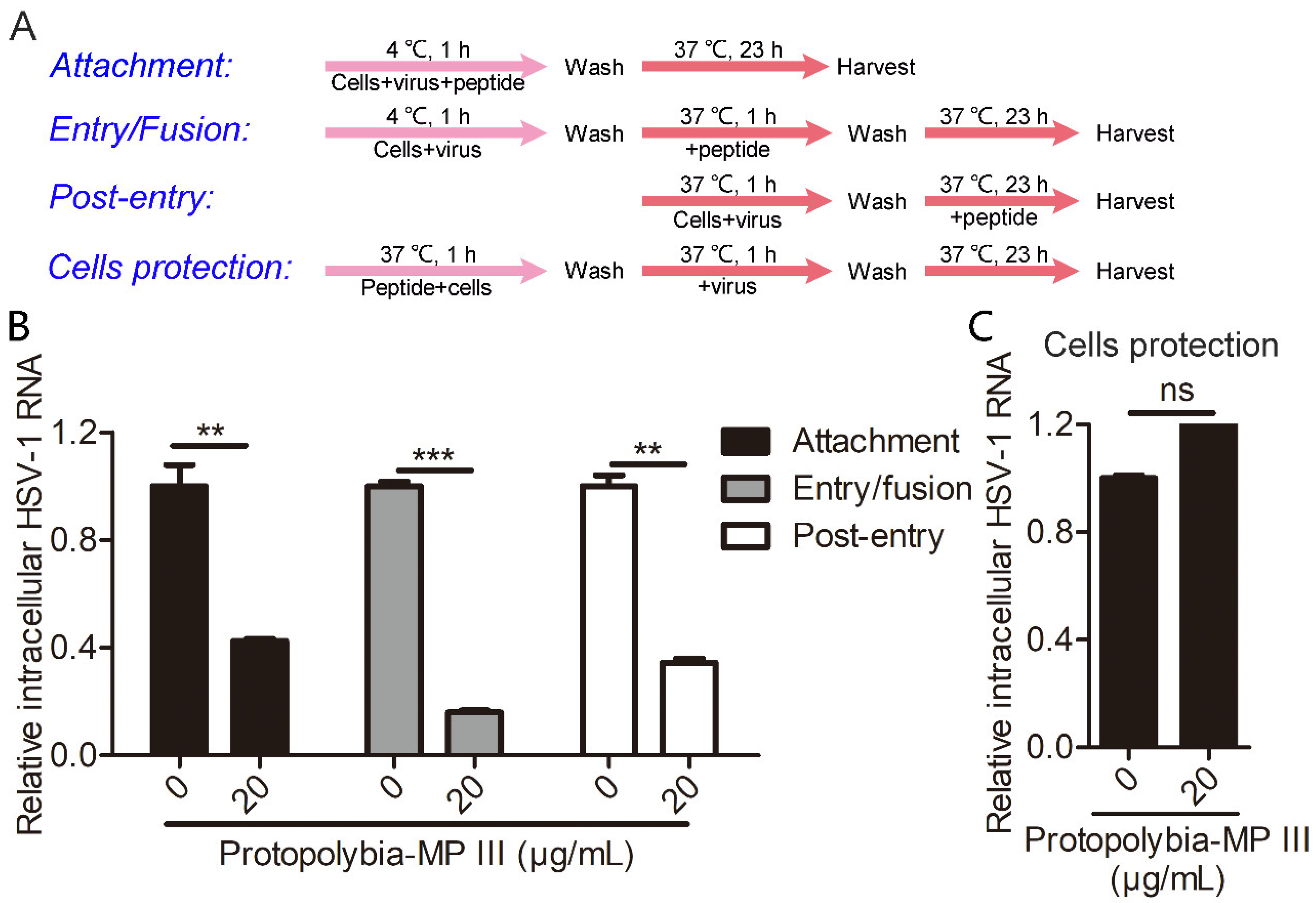
2.2. Protopolybia-MP III Blocks Multiple Steps of the HSV-1 Life Cycle
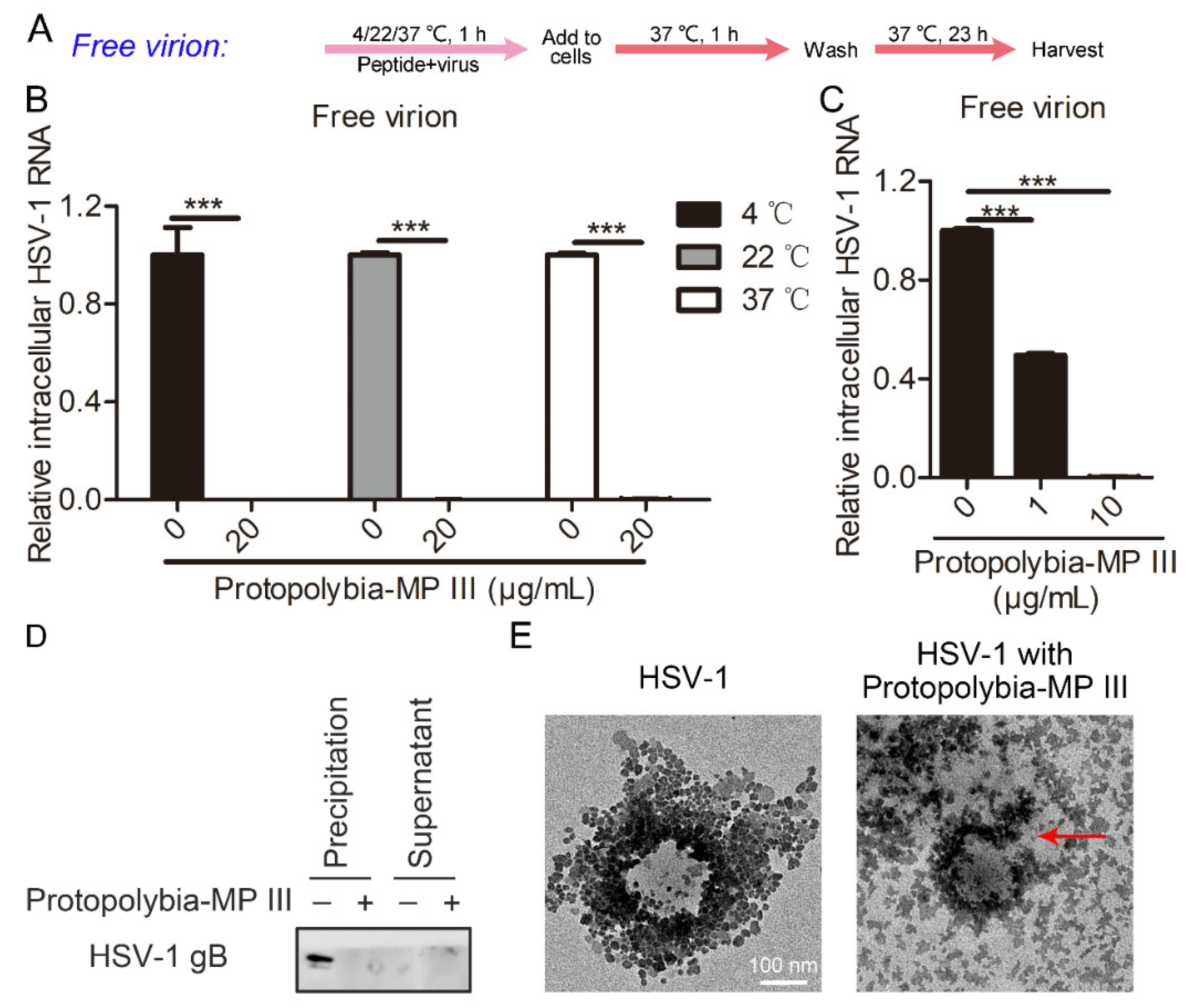
2.3. Protopolybia-MP III Destroys the Integrity of the HSV-1 Virion
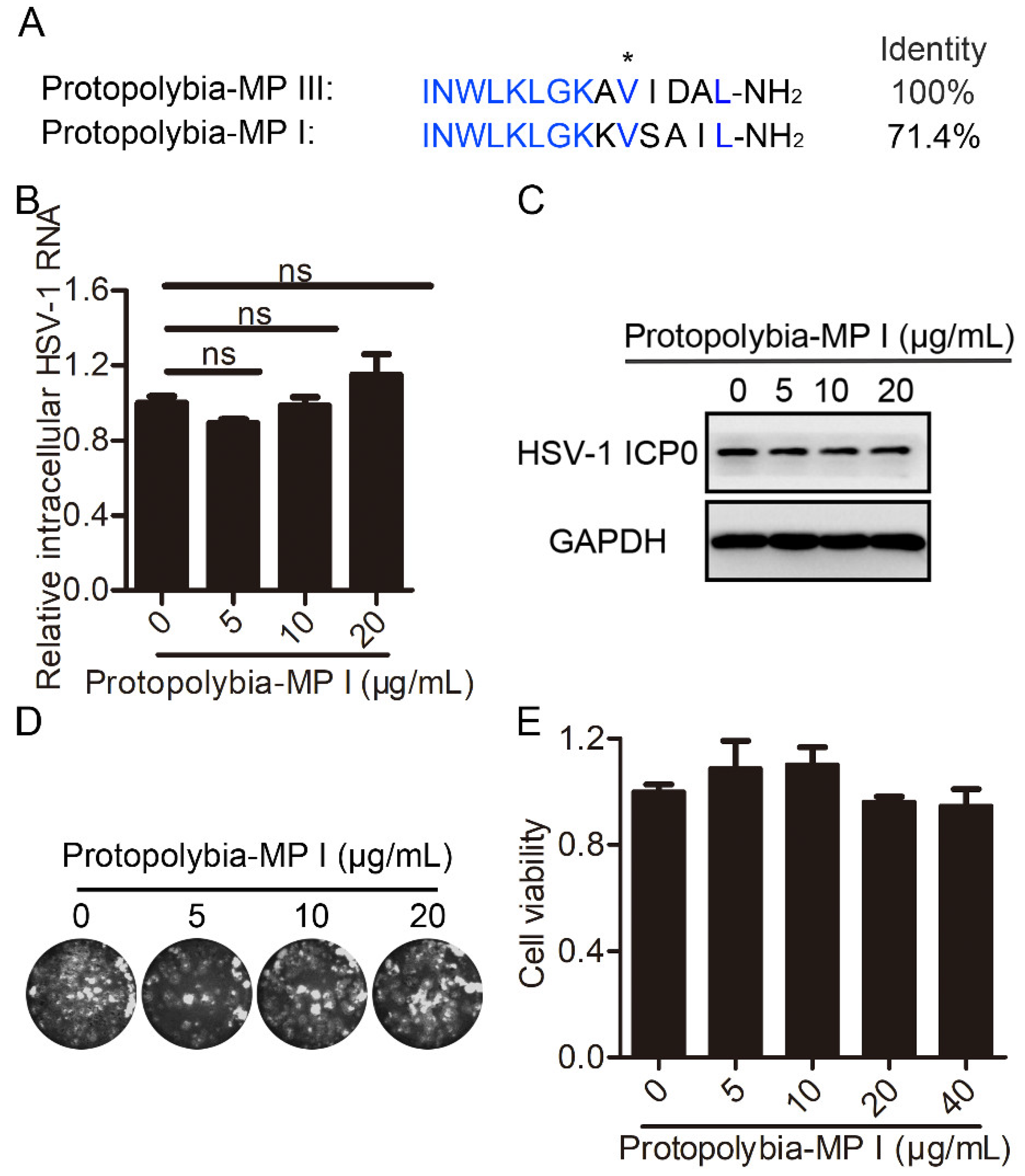
2.4. Protopolybia-MP I Is a Natural Homolog of Protopolybia-MP III with Weak Antiviral Activity against HSV-1
2.5. Mutant Peptide Design and Antiviral Activity Analysis Based on Two Natural Wasp Venom Peptides Protopolybia-MP III and Protopolybia-MP I
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis and Purification
4.2. Cell Culture and Virus Infection
4.3. CCK8 Assay
4.4. qPCR
4.5. Western Blotting
4.6. Plaque Formation Assay
4.7. Immunofluorescence Assay
4.8. Virion Integrity Assay
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rauch, S.; Jasny, E.; Schmidt, K.E.; Petsch, B. New Vaccine Technologies to Combat Outbreak Situations. Front. Immunol. 2018, 9, 1963. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Ahmad, S.; Anunnitipat, K.; Oksenych, V.; Zusinaite, E.; Tenson, T.; Bjoras, M.; Kainov, D.E. Seven classes of antiviral agents. Cell. Mol. Life Sci. 2022, 79, 605. [Google Scholar] [CrossRef] [PubMed]
- Tompa, D.R.; Immanuel, A.; Srikanth, S.; Kadhirvel, S. Trends and strategies to combat viral infections: A review on FDA approved antiviral drugs. Int. J. Biol. Macromol. 2021, 172, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.D.; Chu, J.J.H. Natural products as Zika antivirals. Med. Res. Rev. 2022, 42, 1739–1780. [Google Scholar] [CrossRef]
- Feng, M.; Fei, S.; Xia, J.; Labropoulou, V.; Swevers, L.; Sun, J. Antimicrobial Peptides as Potential Antiviral Factors in Insect Antiviral Immune Response. Front. Immunol. 2020, 11, 2030. [Google Scholar] [CrossRef]
- Qureshi, A.; Thakur, N.; Tandon, H.; Kumar, M. AVPdb: A database of experimentally validated antiviral peptides targeting medically important viruses. Nucleic Acids Res. 2014, 42, D1147–D1153. [Google Scholar] [CrossRef]
- Sureram, S.A.I.; Ueoka, R.; Rittà, M.; Francese, R.; Srivibool, R.; Darshana, D.; Piel, J.; Ruchirawat, S.; Muratori, L.; Lembo, D.; et al. The Peptide A-3302-B Isolated from a Marine Bacterium Micromonospora sp. Inhibits HSV-2 Infection by Preventing the Viral Egress from Host Cells. Int. J. Mol. Sci. 2022, 23, 947. [Google Scholar] [CrossRef]
- Zannella, C.; Chianese, A.; Palomba, L.; Marcocci, M.E.; Bellavita, R.; Merlino, F.; Grieco, P.; Folliero, V.; De Filippis, A.; Mangoni, M.; et al. Broad-Spectrum Antiviral Activity of the Amphibian Antimicrobial Peptide Temporin L and Its Analogs. Int. J. Mol. Sci. 2022, 23, 2060. [Google Scholar] [CrossRef]
- Li, F.; Lang, Y.; Ji, Z.; Xia, Z.; Han, Y.; Cheng, Y.; Liu, G.; Sun, F.; Zhao, Y.; Gao, M.; et al. A scorpion venom peptide Ev37 restricts viral late entry by alkalizing acidic organelles. J. Biol. Chem. 2019, 294, 182–194. [Google Scholar] [CrossRef]
- Yu, J.; Dai, Y.; Fu, Y.; Wang, K.; Yang, Y.; Li, M.; Xu, W.; Wei, L. Cathelicidin antimicrobial peptides suppress EV71 infection via regulating antiviral response and inhibiting viral binding. Antivir. Res. 2021, 187, 105021. [Google Scholar] [CrossRef]
- Agarwal, G.; Gabrani, R. Antiviral Peptides: Identification and Validation. Int. J. Pept. Res. Ther. 2021, 27, 149–168. [Google Scholar] [CrossRef] [PubMed]
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; de Carvalho Neves, N.; Franco, O.L. Antiviral peptides as promising therapeutic drugs. Cell. Mol. Life Sci. 2019, 76, 3525–3542. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Wahed, A.; Yosri, N.; Sakr, H.H.; Du, M.; Algethami, A.F.M.; Zhao, C.; Abdelazeem, A.H.; Tahir, H.E.; Masry, S.H.D.; Abdel-Daim, M.M.; et al. Wasp Venom Biochemical Components and Their Potential in Biological Applications and Nanotechnological Interventions. Toxins 2021, 13, 206. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, M.; Huang, X.; Wu, X.; Xiao, H.; Jin, F.; Lv, J.; Cheng, J.; Zhao, Y.; Zhang, C. Wasp venom from Vespa magnifica acts as a neuroprotective agent to alleviate neuronal damage after stroke in rats. Pharm. Biol. 2022, 60, 334–346. [Google Scholar] [CrossRef]
- Herrera, C.; Leza, M.; Martínez-López, E. Diversity of compounds in Vespa spp. venom and the epidemiology of its sting: A global appraisal. Arch. Toxicol. 2020, 94, 3609–3627. [Google Scholar] [CrossRef]
- de Santana, C.J.C.; Pires Junior, O.R.; Fontes, W.; Palma, M.S.; Castro, M.S. Mastoparans: A Group of Multifunctional alpha-Helical Peptides With Promising Therapeutic Properties. Front. Mol. Biosci. 2022, 9, 824989. [Google Scholar] [CrossRef]
- Alhakamy, N.A.; Ahmed, O.A.A.; Md, S.; Fahmy, U.A. Mastoparan, a Peptide Toxin from Wasp Venom Conjugated Fluvastatin Nanocomplex for Suppression of Lung Cancer Cell Growth. Polymers 2021, 13, 4225. [Google Scholar] [CrossRef]
- Wang, K.R.; Yan, J.-X.; Zhang, B.Z.; Song, J.J.; Jia, P.F.; Wang, R. Novel mode of action of polybia-MPI, a novel antimicrobial peptide, in multi-drug resistant leukemic cells. Cancer Lett. 2009, 278, 65–72. [Google Scholar] [CrossRef]
- Hilterbrand, A.T.; Daly, R.E.; Heldwein, E.E. Contributions of the Four Essential Entry Glycoproteins to HSV-1 Tropism and the Selection of Entry Routes. mBio 2021, 12, e00143-21. [Google Scholar] [CrossRef]
- Marcocci, M.E.; Napoletani, G.; Protto, V.; Kolesova, O.; Piacentini, R.; Li Puma, D.D.; Lomonte, P.; Grassi, C.; Palamara, A.T.; De Chiara, G. Herpes Simplex Virus-1 in the Brain: The Dark Side of a Sneaky Infection. Trends Microbiol. 2020, 28, 808–820. [Google Scholar] [CrossRef]
- Rowe, A.M.; St Leger, A.J.; Jeon, S.; Dhaliwal, D.K.; Knickelbein, J.E.; Hendricks, R.L. Herpes keratitis. Prog. Retin. Eye Res. 2013, 32, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Farooq, A.V.; Shukla, D. Herpes simplex epithelial and stromal keratitis: An epidemiologic update. Surv. Ophthalmol. 2012, 57, 15. [Google Scholar] [CrossRef] [PubMed]
- Frobert, E.; Cortay, J.-C.; Ooka, T.; Najioullah, F.; Thouvenot, D.; Lina, B.; Morfin, F. Genotypic detection of acyclovir-resistant HSV-1: Characterization of 67 ACV-sensitive and 14 ACV-resistant viruses. Antivir. Res. 2008, 79, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Ling, S.; Wang, D.; Dai, Y.; Jiang, H.; Zhou, X.; Paludan, S.R.; Hong, J.; Cai, Y. Targeting herpes simplex virus with CRISPR–Cas9 cures herpetic stromal keratitis in mice. Nat. Biotechnol. 2021, 39, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-J.; Buznyk, O.; Kuffova, L.; Rajendran, V.; Forrester, J.V.; Phopase, J.; Islam, M.M.; Skog, M.; Ahlqvist, J.; Griffith, M. Cathelicidin LL-37 and HSV-1 Corneal Infection: Peptide Versus Gene Therapy. Transl. Vision. Sci. Technol. 2014, 3, 4. [Google Scholar] [CrossRef]
- Rodriguez-Izquierdo, I.; Serramia, M.J.; Gomez, R.; De La Mata, F.J.; Bullido, M.J.; Munoz-Fernandez, M.A. Gold Nanoparticles Crossing Blood-Brain Barrier Prevent HSV-1 Infection and Reduce Herpes Associated Amyloid-βsecretion. J. Clin. Med. 2020, 9, 155. [Google Scholar]
- Cheng, Y.; Sun, F.; Wang, L.; Gao, M.; Xie, Y.; Sun, Y.; Liu, H.; Yuan, Y.; Yi, W.; Huang, Z.; et al. Virus-induced p38 MAPK activation facilitates viral infection. Theranostics 2020, 10, 12223–12240. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Liu, X.; Luo, X.; Sun, F.; Qin, C.; Ding, L.; Zhu, W.; Zhang, H.; Zhou, H.; Chen, Z. Characterization of the Molecular Diversity and Degranulation Activity of Mastoparan Family Peptides from Wasp Venoms. Toxins 2023, 15, 331. [Google Scholar] [CrossRef]
- Muttenthaler, M.; King, G.F.; Adams, D.J.; Alewood, P.F. Trends in peptide drug discovery. Nat. Rev. Drug Discov. 2021, 20, 309–325. [Google Scholar] [CrossRef]
- Kuroki, A.; Tay, J.; Lee, G.H.; Yang, Y.Y. Broad-Spectrum Antiviral Peptides and Polymers. Adv. Healthc. Mater. 2021, 10, e2101113. [Google Scholar] [CrossRef]
- Wachinger, M.K.A.; Winder, D.; von Pechmann, N.; Ludvigsen, A.; Neumann, M.; Holle, R.; Salmons, B.; Erfle, V.; Brack-Werner, R. Antimicrobial peptides melittin and cecropin inhibit replication of human immunodeficiency virus 1 by suppressing viral gene expression. J. Gen. Virol. 1998, 79, 10. [Google Scholar] [CrossRef] [PubMed]
- Nawae, W.; Hannongbua, S.; Ruengjitchatchawalya, M. Molecular dynamics exploration of poration and leaking caused by Kalata B1 in HIV-infected cell membrane compared to host and HIV membranes. Sci. Rep. 2017, 7, 3638. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Li, T.; Song, Y.; Zhang, R.; Zeng, Z.; Han, S.; Zhang, X.; Wu, Y.; Li, W.; Cao, Z. Inhibitory activity and mechanism of two scorpion venom peptides against herpes simplex virus type 1. Antivir. Res. 2014, 102, 1–10. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Casciaro, B.; Genovese, A.; Brancaccio, D.; Marcocci, M.E.; Novellino, E.; Carotenuto, A.; Palamara, A.T.; Mangoni, M.L.; Nencioni, L. Temporin G, an amphibian antimicrobial peptide against influenza and parainfluenza respiratory viruses: Insights into biological activity and mechanism of action. FASEB J. 2021, 35, e21358. [Google Scholar] [CrossRef] [PubMed]
- Graci, J.D.; Cameron, C.E. Mechanisms of action of ribavirin against distinct viruses. Rev. Med. Virol. 2005, 16, 37–48. [Google Scholar] [CrossRef]
- Huang, D.; Taha, M.S.; Nocera, A.L.; Workman, A.D.; Amiji, M.M.; Bleier, B.S. Cold exposure impairs extracellular vesicle swarm–mediated nasal antiviral immunity. J. Allergy Clin. Immunol. 2023, 151, 509–525.e508. [Google Scholar] [CrossRef]
- Sasaki-Tanaka, R.S.T.; Moriyama, M.; Okamoto, H.; Kogure, H.; Kanda, T. Amantadine and Rimantadine Inhibit Hepatitis A Virus Replication through the Induction of Autophagy. J. Virol. 2022, 96, e0064622. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, F.; Ye, X.; Han, T.; Xia, J.; Wu, L.; Zhu, W.; Ding, L.; Luo, X.; Qin, C.; Chen, Z. Function and Mechanism of Antiviral Wasp Venom Peptide Protopolybia-MP III and Its Derivatives against HSV-1. Toxins 2024, 16, 132. https://doi.org/10.3390/toxins16030132
Sun F, Ye X, Han T, Xia J, Wu L, Zhu W, Ding L, Luo X, Qin C, Chen Z. Function and Mechanism of Antiviral Wasp Venom Peptide Protopolybia-MP III and Its Derivatives against HSV-1. Toxins. 2024; 16(3):132. https://doi.org/10.3390/toxins16030132
Chicago/Turabian StyleSun, Fang, Xiangdong Ye, Tanran Han, Jingwen Xia, Lili Wu, Wen Zhu, Li Ding, Xudong Luo, Chenhu Qin, and Zongyun Chen. 2024. "Function and Mechanism of Antiviral Wasp Venom Peptide Protopolybia-MP III and Its Derivatives against HSV-1" Toxins 16, no. 3: 132. https://doi.org/10.3390/toxins16030132
APA StyleSun, F., Ye, X., Han, T., Xia, J., Wu, L., Zhu, W., Ding, L., Luo, X., Qin, C., & Chen, Z. (2024). Function and Mechanism of Antiviral Wasp Venom Peptide Protopolybia-MP III and Its Derivatives against HSV-1. Toxins, 16(3), 132. https://doi.org/10.3390/toxins16030132