Fusarium Species and Mycotoxins Associated with Sorghum Grains in Uruguay

 , and
, and
Abstract
1. Introduction
2. Results
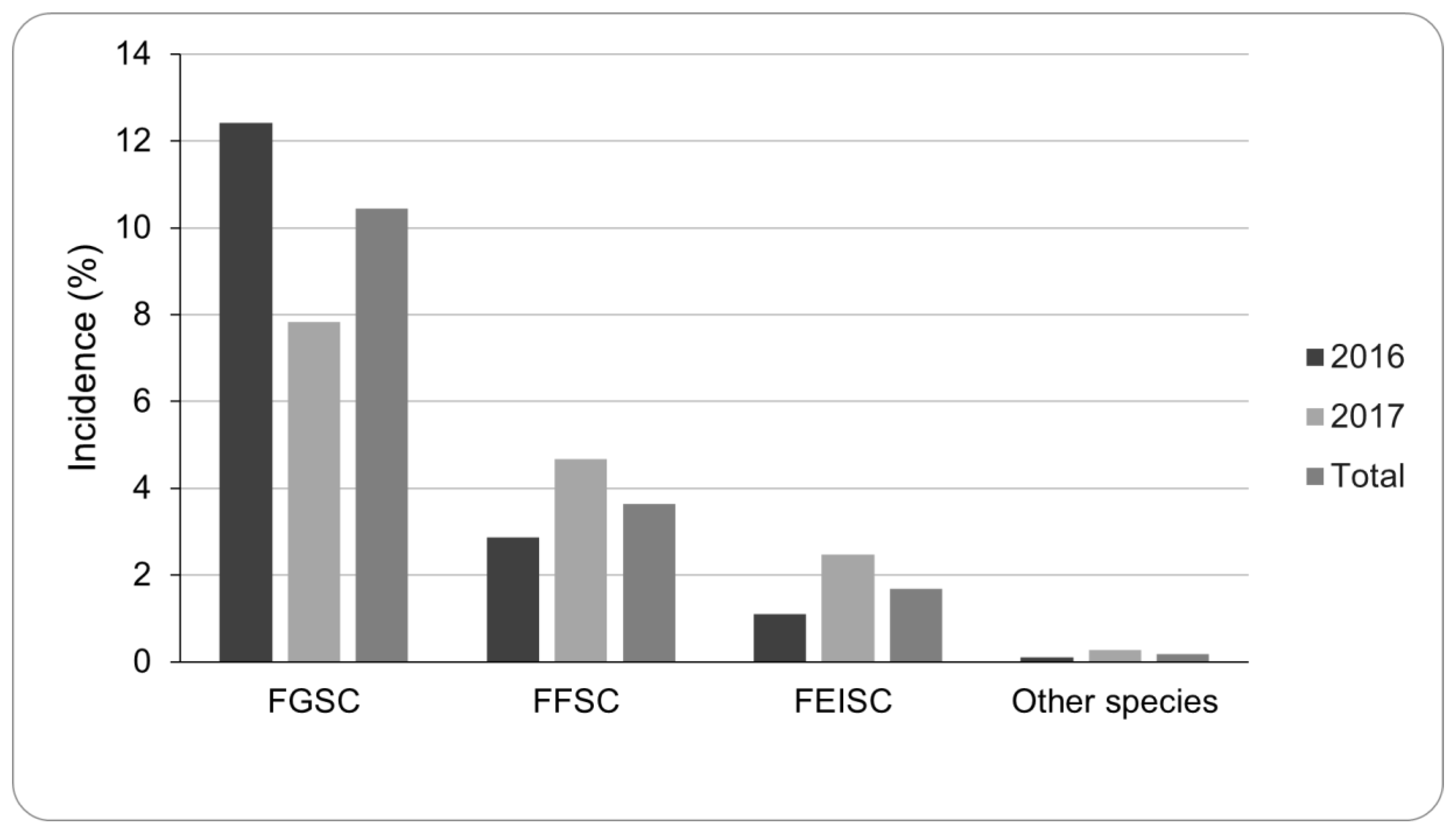
2.1. Species Identification
2.2. Trichothecene Genotype Determination
2.3. Fusarium Mycotoxin Production
2.4. Determination of Mycotoxins in Sorghum
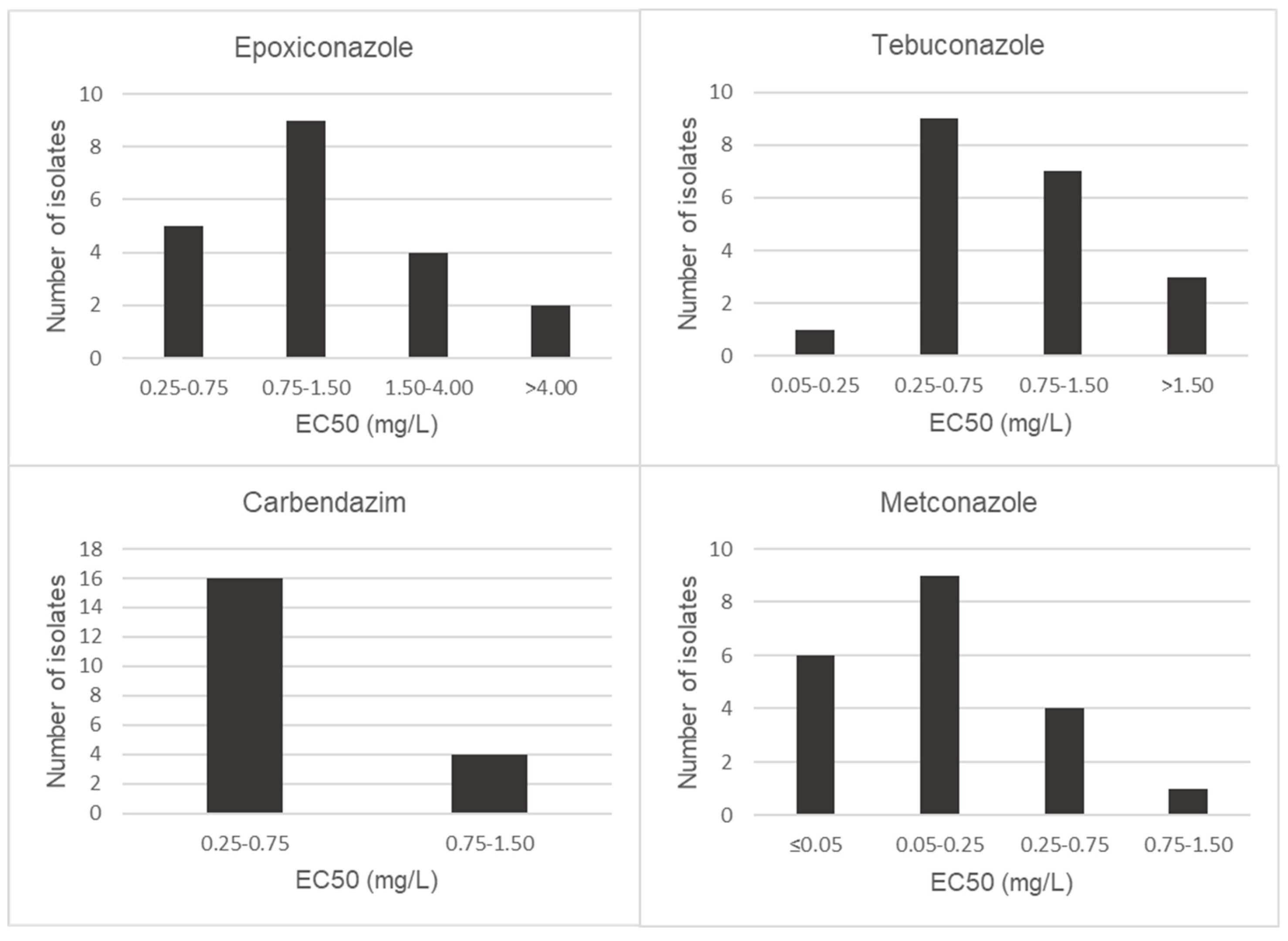
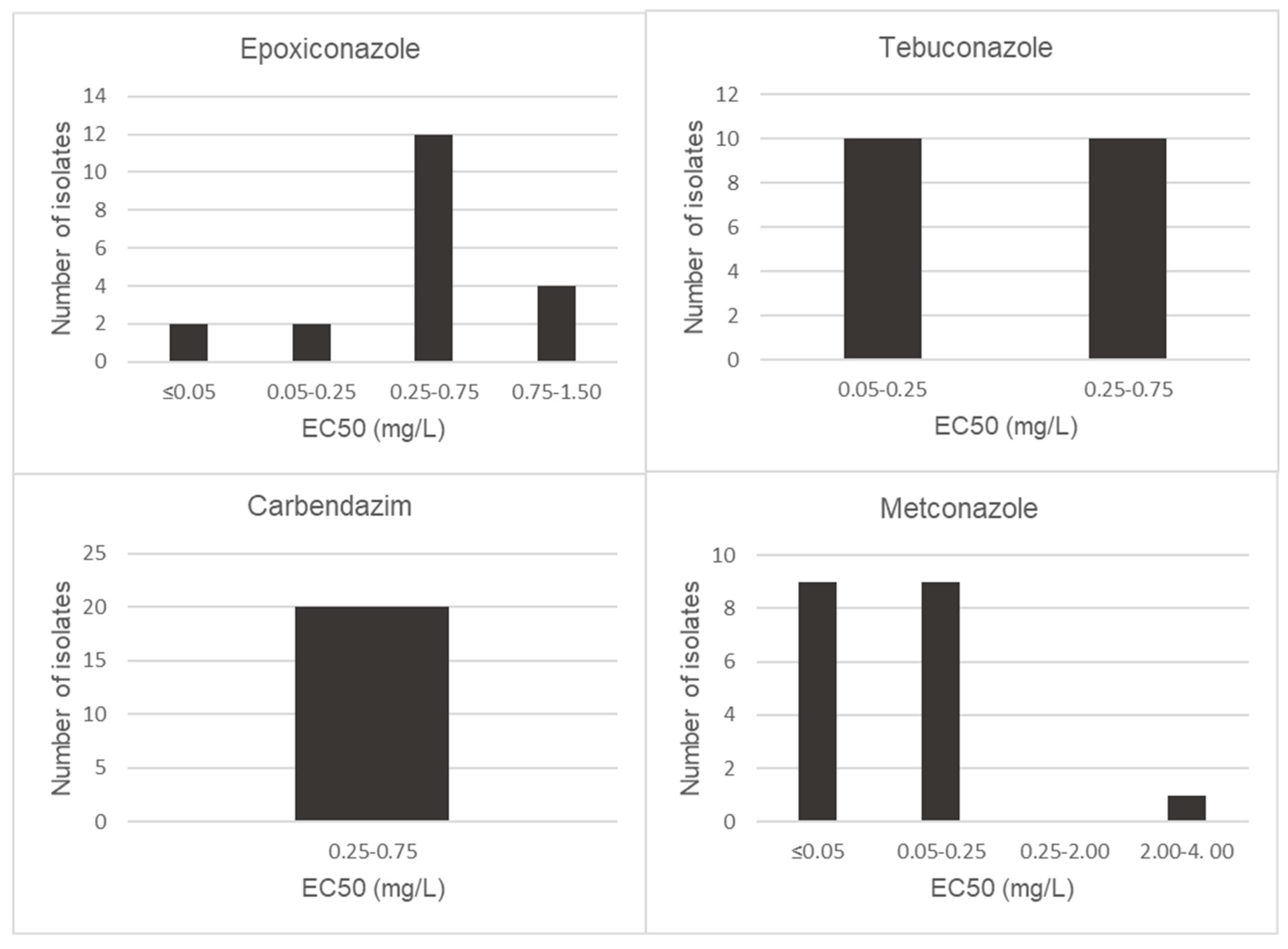
2.5. Fungicide Sensitivity Assay
3. Discussion
4. Conclusions
5. Materials and Methods
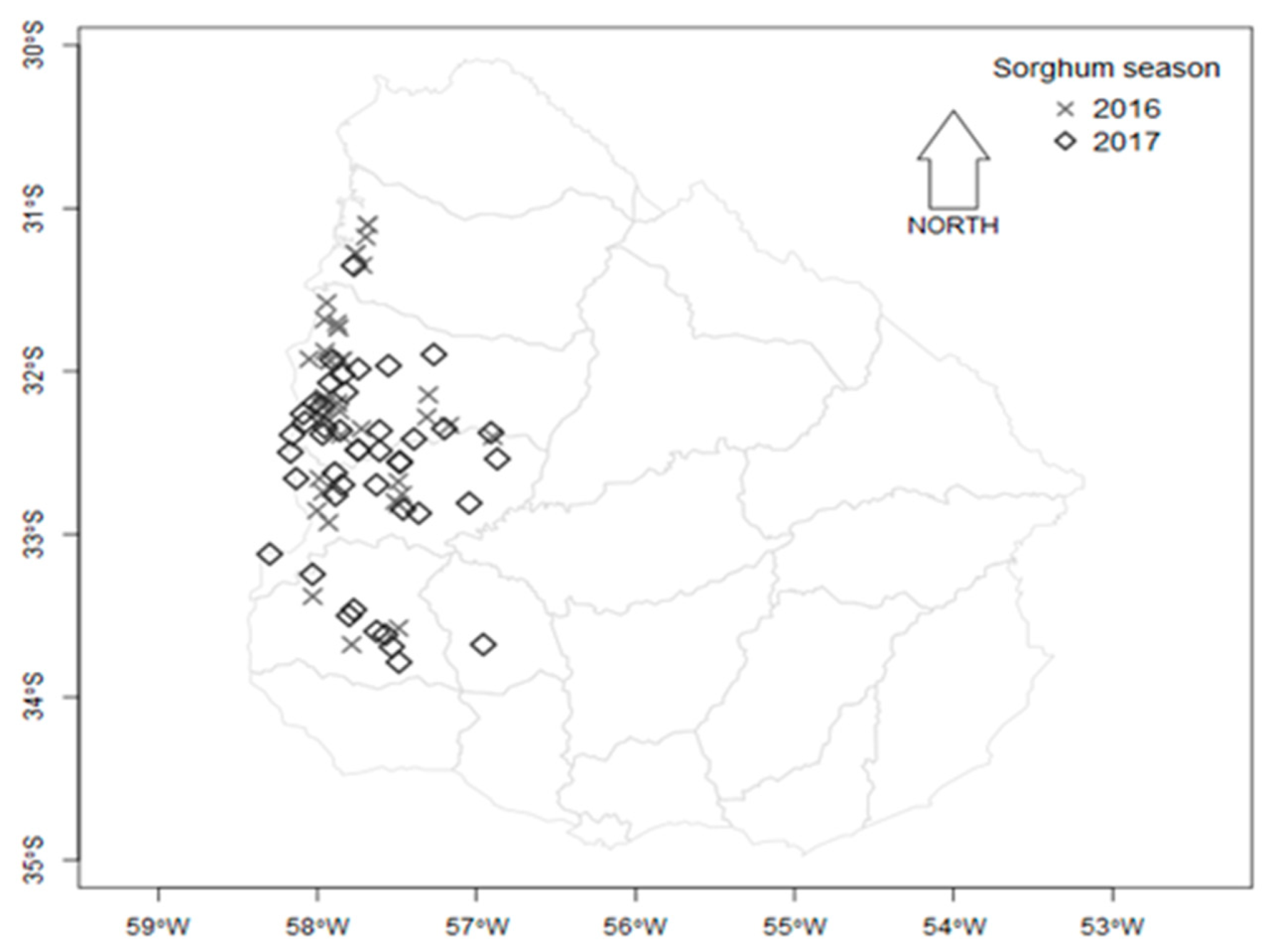
5.1. Sorghum Samples
5.2. Fusarium Isolation
5.3. Molecular Identification
5.4. Trichothecene Genotype Determination
5.5. Fusarium Mycotoxin Production
5.6. Determination of Mycotoxins in Sorghum
5.7. Fungicide Sensitivity Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. FAOSTAT Statistical Database. 2015. Available online: http://faostat3.fao.org/home/E (accessed on 6 May 2022).
- United States Department of Agriculture (USDA). World Agricultural Production. 2022. Available online: https://apps.fas.usda.gov/psdonline/circulars/production.pdf (accessed on 6 May 2022).
- Astoreca, A.; Emateguy, G.; Alconada, T. Fungal contamination and mycotoxins associated with sorghum crop: Its relevance today. Eur. J. Plant Pathol. 2019, 155, 381–392. [Google Scholar] [CrossRef]
- United States Department of Agriculture (USDA). Office of the Chief Economist. In Agricultural Projections to 2024; World Agricultural Outlook Board: Washington, DC, USA, 2015. [Google Scholar]
- DIEA. Anuario estadístico agropecuario, Ministerio de Ganadería Agricultura y Pesca. Montevideo. DIEA. 2013. Available online: https://descargas.mgap.gub.uy/DIEA/Documentos%20compartidos/Anuario2013/Diea_Anuario_2013.pdf (accessed on 10 August 2022).
- Mehtol, M. Maiz y sorgo: Situacion y perspectivas, 2021. In Anuario de OPYPA de 2021; Ministerio de Ganadería, Agricultura y Pesca: Montevideo, Uruguay, 2021; p. 193. [Google Scholar]
- Ackerman, A.; Wenndt, A.; Boyles, R. The sorghum grain mold disease complex: Pathogens, host responses, and the bioactive metabolites at play. Front. Plant Sci. 2021, 12, 660171. [Google Scholar] [CrossRef] [PubMed]
- Alves dos Reis, T.; Zorzete, P.; Rodrigues Pozzi, C.; Nascimiento da Silva, V.; Ortega, E.; Correa, B. Mycoflora and fumonisin contamination in Brazilian sorghum from sowing to harvest. J. Sci. Food Agric. 2010, 90, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Divakara, S.; Santosh, P.; Aiyaz, M.; Ramana, M.; Hariprasad, P.; Nayaka, S.; Niranjana, S. Molecular identification and characterization of Fusarium spp. associated with sorghum seeds. J. Sci. Food Agric. 2014, 94, 1132–1139. [Google Scholar] [CrossRef]
- Lahouar, A.; Crespo-Sempere, A.; Marín, S.; Saïd, S.; Sanchis, V. Toxigenic molds in Tunisian and Egyptian sorghum for human consumption. J. Stored Prod. Res. 2015, 63, 57–62. [Google Scholar] [CrossRef]
- Mohamed Nor, N.M.I.; Salleh, B.; Leslie, J.F. Fusarium Species from Sorghum in Thailand. Plant Pathol. J. 2019, 35, 301–312. [Google Scholar] [CrossRef]
- Pena, G.A.; Cavaglieri, L.R.; Chulze, S.N. Fusarium species and moniliformin occurrence in sorghum grains used as an ingredient for animal feed in Argentina. J. Sci. Food Agr. 2019, 99, 47–54. [Google Scholar] [CrossRef]
- Leslie, J.F.; Zeller, K.A.; Lamprecht, S.C.; Rheeder, J.P.; Marasas, W.F. Toxicity, pathogenicity, and genetic differentiation of five species of Fusarium from sorghum and millet. Phytopathology 2005, 95, 275–283. [Google Scholar] [CrossRef]
- Marasas, W.F.O.; Rheeder, J.P.; Lamprecht, S.C.; Zeller, K.A.; Leslie, J.F. Fusarium andiyazi sp. nov., a new species from sorghum. Mycologia 2001, 93, 1203–1210. [Google Scholar] [CrossRef]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Molto, J.C.; Manes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Garmendia, G.; Pattarino, L.; Negrin, C.; Martinez-Silveira, A.; Pereyra, S.; Ward, T.; Vero, S. Species composition, toxigenic potential and aggressiveness of Fusarium isolates causing Head blight of barley in Uruguay. Food Microbiol. 2018, 76, 426–433. [Google Scholar] [CrossRef]
- Pan, D.; Mionetto, A.; Calero, N.; Reynoso, M.M.; Torres, A.; Bettucci, L. Population genetic analysis and trichothecene profiling of Fusarium graminearum from wheat in Uruguay. Genet. Mol. Res. 2016, 15, 1–11. [Google Scholar] [CrossRef]
- Umpiérrez-Failache, M.; Garmendia, G.; Pereyra, S.; Rodríguez-Haralambides, A.; Ward, T.J.; Vero, S. Regional differences in species composition and toxigenic potential among Fusarium head blight isolates from Uruguay indicate a risk of nivalenol contamination in new wheat production areas. Int. J. Food Microbiol. 2013, 166, 135–140. [Google Scholar] [CrossRef]
- del Palacio, A.; Mionetto, A.; Bettucci, L.; Pan, D. Evolution of fungal populations and mycotoxins in sorghum silage. Food Addit. Contam. A 2016, 33, 1864–1872. [Google Scholar] [CrossRef]
- Kelly, L.; Tan, Y.; Ryley, M.; Aitken, E. Fusarium species associated with stalk rot and head blight of grain sorghum in Queensland and New South Wales. Plant Pathol. 2017, 66, 1413–1423. [Google Scholar] [CrossRef]
- Sharma, R.; Thakur, R.P.; Senthilvel, S.; Nayak, S.; Reddy, S.V.; Rao, V.P.; Varshney, R.K. Identification and characterization of toxigenic Fusaria associated with sorghum grain mold complex in India. Mycopathologia 2011, 171, 223–230. [Google Scholar] [CrossRef][Green Version]
- Balota, M. Sorghum diseases, head mold. Virginia Polytechnic Institute and State University, Extension Program, Virginia State, Petersburg, VA, USA. 2012. [Google Scholar]
- Funnell-Harris, L.D.; Pedersen, J.F. Presence of Fusarium spp. in air and soil associated with sorghum fields. Plant Dis. 2011, 95, 648–656. [Google Scholar] [CrossRef]
- Leslie, J.F.; Marasas, W.F.O.; Diourté, M. Naturally occurring mycotoxins in sorghum and millet from Mali (abstract). In Proceedings of the INTSORMIL Principal Investigators Conference, Addis Ababa, Ethiopia, 18–20 November 2002. [Google Scholar]
- Petrovic, T.; Walsh, J.L.; Burgess, L.W.; Summerell, B.A. Fusarium species associated with stalk rot of grain sorghum in the northern grain belt of eastern Australia. Aust. Plant Pathol. 2009, 38, 373–379. [Google Scholar] [CrossRef]
- Pan, D.; Calero, N.; Mionetto, A.; Bettucci, L. Trichothecene genotypes of Fusarium graminearum from wheat in Uruguay. Int. J. Food Microbiol. 2013, 162, 120–123. [Google Scholar] [CrossRef]
- Del Ponte, E.M.; Moreira, G.M.; Ward, T.J.; O’Donnell, K.; Nicolli, C.P.; Machado, F.J.; Duffeck, M.R.; Alves, K.S.; Tessmann, D.J.; Waalwijk, C.; et al. Fusarium graminearum Species Complex: A Bibliographic Analysis and Web-Accessible Database for Global Mapping of Species and Trichothecene Toxin Chemotypes. Phytopathology 2022, 112, 741–751. [Google Scholar] [CrossRef] [PubMed]
- FDA. Mycotoxins. 2010. Available online: https://www.fda.gov/food/natural-toxins-food/mycotoxins (accessed on 4 May 2022).
- Chilaka, C.; De Boevre, M.; Atanda, O.; De Saeger, S. The status of Fusarium mycotoxins in Sub-Saharan Africa: A review of emerging trends and post-harvest mitigation strategies towards food control. Toxins 2017, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Hanvi, D.; Lawson-Evi, P.; De Boever, M.; Goto, C.E.; De Saeger, S.; Eklu-Gadegbeku, K. Natural occurrence of mycotoxins in maize and sorghum in Togo. Mycotoxin Res. 2019, 35, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Resolución S/N/001. 2001. Límites Máximos de DON en Alimentos Para Animales. 2001. Available online: https://www.gub.uy/ministerio-ganaderia-agricultura-pesca/institucional/normativa/resolucion-sn001-limites-maximos-don-alimentos-para-animales (accessed on 6 May 2022).
- Aoyama, K.; Ishikuro, E.; Nishiwaki, M.; Ichinoe, M. Zearalenone contamination and the causative fungi in sorghum. Shokuhin Eiseigaku Zasshi 2009, 50, 47–51. [Google Scholar] [CrossRef]
- Ayalew, A.; Fehrmann, H.; Lepschy, J.; Beck, R.; Abate, D. Natural occurrence of mycotoxins in staple cereals from Ethiopia. Mycopathologia 2006, 162, 57–63. [Google Scholar] [CrossRef]
- Chala, A.; Taye, W.; Ayalew, A.; Krska, R.; Sulyok, M.; Logrieco, A. Multi mycotoxin analysis of sorghum (Sorghum bicolor L. Moench) and finger millet (Eleusine coracana L. Garten) from Ethiopia. Food Control 2014, 45, 29–35. [Google Scholar] [CrossRef]
- Cendoya, M.; Nichea, J.; Monge, M.; Zachett, V.; Stella, M.; Ramirez, M. Effect of fungicides commonly used for Fusarium head blight management on growth and fumonisin production by Fusarium proliferatum. Rev. Argent. Microbiol. 2021, 53, 64–74. [Google Scholar] [CrossRef]
- Marín, P.; De Ory, A.; Cruz, A.; Magan, N.; González-Jaén, M.T. Potential effects of environmental conditions on the efficiency of the antifungal tebuconazole controlling Fusarium verticillioides and Fusarium proliferatum growth rate and fumonisin biosynthesis. Int. J. Food Microbiol. 2013, 165, 251–258. [Google Scholar] [CrossRef]
- Xu, S.; Wang, J.; Wang, H.; Bao, Y.; Li, Y.; Govindaraju, M.; Yao, W.; Chen, B.; Zhang, M. Molecular characterization of carbendazim resistance of Fusarium species complex that causes sugarcane pokkah boeng disease. BMC Genomics 2019, 20, 115. [Google Scholar] [CrossRef]
- Liu, S.; Fu, L.; Chen, J.; Chen, J.; Wang, S.; Liu, J.; Jiang, J.; Che, Z.; Tian, Y.; Chen, G. Baseline sensitivity and control efficacy of epoxiconazole against Fusarium graminearum in Henan Province, China. Eur J Plant Pathol. 2020, 157, 825–833. [Google Scholar] [CrossRef]
- Ivić, D.; Sever, Z.; Kumanovska, B. In vitro sensitivity of Fusarium graminearum, F. avenaceum and F. verticillioides to carbendazim, tebuconazole, flutriafol, metconazole and prochloraz. Pestic i Phytomed (Belgrade) 2011, 26, 35–42. [Google Scholar] [CrossRef]
- Machado, F.J.; Santana, F.M.; Lau, D.; Del Ponte, E.M. Quantitative Review of the effects of triazole and benzimidazole fungicides on Fusarium head blight and wheat yield in Brazil. Plant Dis. 2017, 101, 1633–1641. [Google Scholar] [CrossRef]
- Spolti, P.; Del Ponte, E.M.; Dong, Y.; Cummings, J.A.; Bergstrom, G.C. Triazole sensitivity in a contemporary population of Fusarium graminearum from New York wheat and competitiveness of a tebuconazole-resistant isolate. Plant Dis. 2014, 98, 607–613. [Google Scholar] [CrossRef]
- Yin, Y.; Liu, X.; Li, B.; Ma, Z. Characterization of sterol demethylation inhibitor-resistant isolates of Fusarium asiaticum and F. graminearum collected from wheat in China. Phytopathology 2009, 99, 487–497. [Google Scholar] [CrossRef]
- Anderson, N.; Freije, A.; Bergstrom, G.; Bradley, C.; Cowger, C.; Faske, T.; Hollier, C.; Kleczewski, N.; Padgett0, G.; Paul, P.; et al. Sensitivity of Fusarium graminearum to Metconazole and Tebuconazole Fungicides Before and After Widespread Use in Wheat in the United States. Plant Health Prog. 2020, 21, 85–90. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Professional: Ames, IA, USA, 2006. [Google Scholar]
- Garmendia, G.; Umpierrez-Failache, M.; Ward, T.; Vero, S. Development of a PCR-RFLP method based on the transcription elongation factor 1-α gene to differentiate Fusarium graminearum from other species within the Fusarium graminearum species complex. Food Microbiol. 2018, 70, 28–32. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Molecular Biology and Evolution 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Ward, T.J.; Bielawski, J.P.; Kistler, H.C.; Sullivan, E.; O’Donnell, K. Ancestral polymorphism and adaptive evolution in the trichothecene mycotoxin gene cluster of phytopathogenic Fusarium. Proc. Natl. Acad. Sci. USA 2002, 99, 9278–9283. [Google Scholar] [CrossRef]
- Chandler, E.; Simpson, D.; Thomsett, M.; Nicholson, P. Development of PCR Assays to Tri7 and Tril3 Trichothecene Biosynthetic GeneRevista Argentina de Microbiologías, and Characterization of Chemotypes of Fusarium. Physiol. Mol. Plant Pathol. 2003, 62, 355–367. [Google Scholar] [CrossRef]
- Reynoso, M.M.; Ramirez, M.L.; Torres, A.M.; Chulze, S.N. Trichothecene genotypes and chemotypes in Fusarium graminearum strains isolated from wheat in Argentina. Int J Food Microbiol. 2011, 145, 444–448. [Google Scholar] [CrossRef] [PubMed]
- AOAC. International Official Methods of Analysis of AOAC International, 20th ed.; AOAC: Gaithersburg, MD, USA, 2016. [Google Scholar]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef]
- Tateishi, H.; Miyake, T.; Mori, M.; Kimura, R.; Sakuma, Y.; Saishoji, T. Sensitivity of Japanese Fusarium graminearum species complex isolates to metconazole. J. Pestic. Sci. 2010, 35, 419–430. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis (2nd Ed). J. Inst. Actuar. 1952, 78, 388–390. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Mean (µg/kg) ± SD | Positive Samples (%) | Contamination in Positive Samples (µg/kg) | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | |||||
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | |||
| DON | 1315 ± 1597 | 565 ± 567 | 88 | 93 | 1507 ± 1623 | 59–6071 | 619 ± 564 | 59–2133 |
| NIV | nd * | 4 ± 19 | 0 | 5 | -- | -- | 79 ± 36 | 50–120 |
| ZEN | 1453 ± 2194 | 752 ± 1297 | 97 | 90 | 1567 ± 2239 | 30–9280 | 841 ± 1344 | 30–6560 |
| FB1 | 67 ± 244 | 58 ± 148 | 16 | 23 | 411 ± 493 | 80–1693 | 255 ± 217 | 80–753 |
| FB2 | 5 ± 36 | 13 ± 98 | 2 | 2 | -- | 80–270 | -- | 80–742 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corallo, A.B.; del Palacio, A.; Oliver, M.; Tiscornia, S.; Simoens, M.; Cea, J.; de Aurrecoechea, I.; Martínez, I.; Sanchez, A.; Stewart, S.; et al. Fusarium Species and Mycotoxins Associated with Sorghum Grains in Uruguay. Toxins 2023, 15, 484. https://doi.org/10.3390/toxins15080484
Corallo AB, del Palacio A, Oliver M, Tiscornia S, Simoens M, Cea J, de Aurrecoechea I, Martínez I, Sanchez A, Stewart S, et al. Fusarium Species and Mycotoxins Associated with Sorghum Grains in Uruguay. Toxins. 2023; 15(8):484. https://doi.org/10.3390/toxins15080484
Chicago/Turabian StyleCorallo, Ana Belén, Agustina del Palacio, María Oliver, Susana Tiscornia, Macarena Simoens, Jaqueline Cea, Inés de Aurrecoechea, Inés Martínez, Alicia Sanchez, Silvina Stewart, and et al. 2023. "Fusarium Species and Mycotoxins Associated with Sorghum Grains in Uruguay" Toxins 15, no. 8: 484. https://doi.org/10.3390/toxins15080484
APA StyleCorallo, A. B., del Palacio, A., Oliver, M., Tiscornia, S., Simoens, M., Cea, J., de Aurrecoechea, I., Martínez, I., Sanchez, A., Stewart, S., & Pan, D. (2023). Fusarium Species and Mycotoxins Associated with Sorghum Grains in Uruguay. Toxins, 15(8), 484. https://doi.org/10.3390/toxins15080484