Molecular Characterization and Functional Analysis of the Dipeptidyl Peptidase IV from Venom of the Ectoparasitoid Scleroderma guani


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
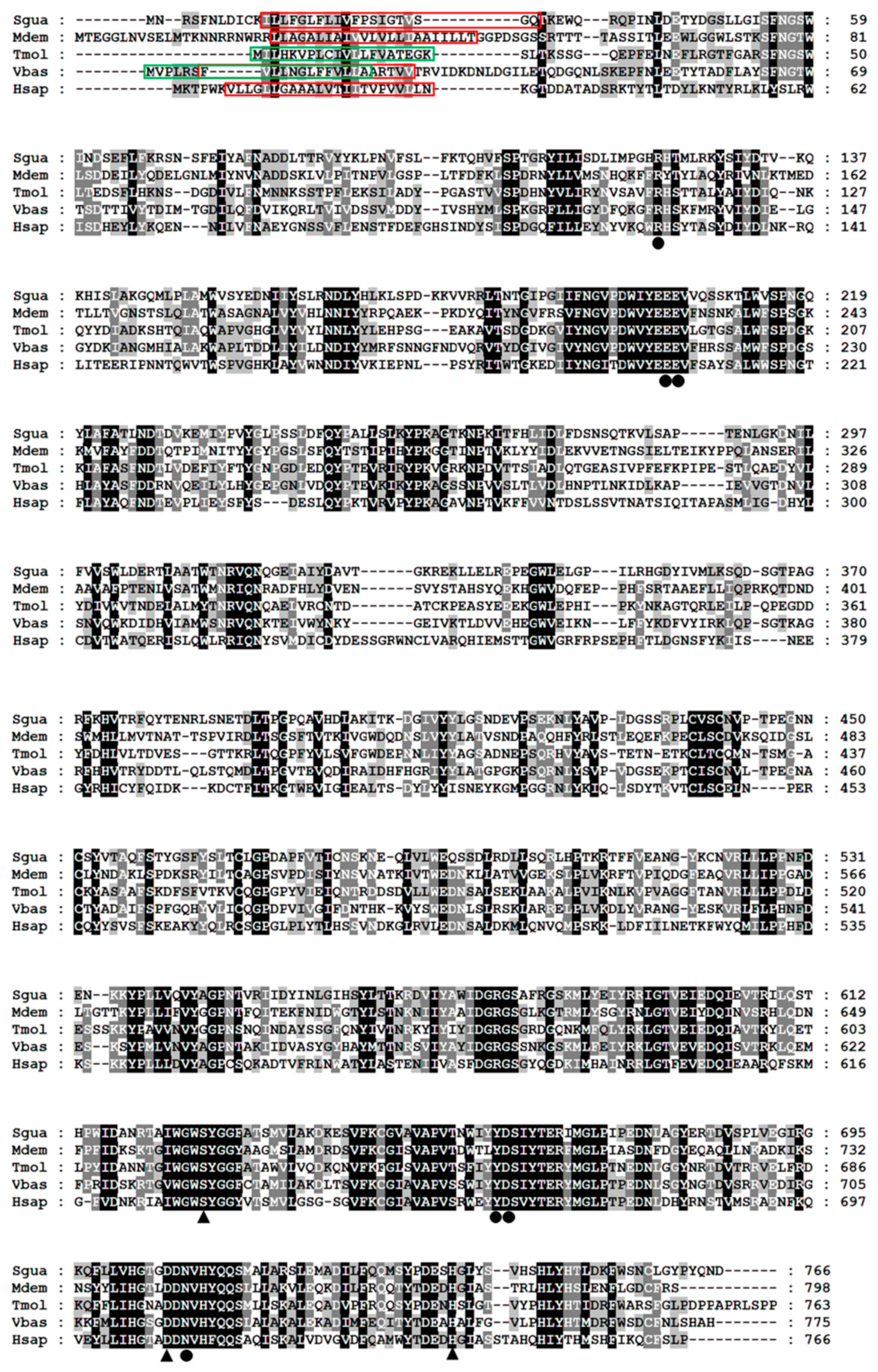
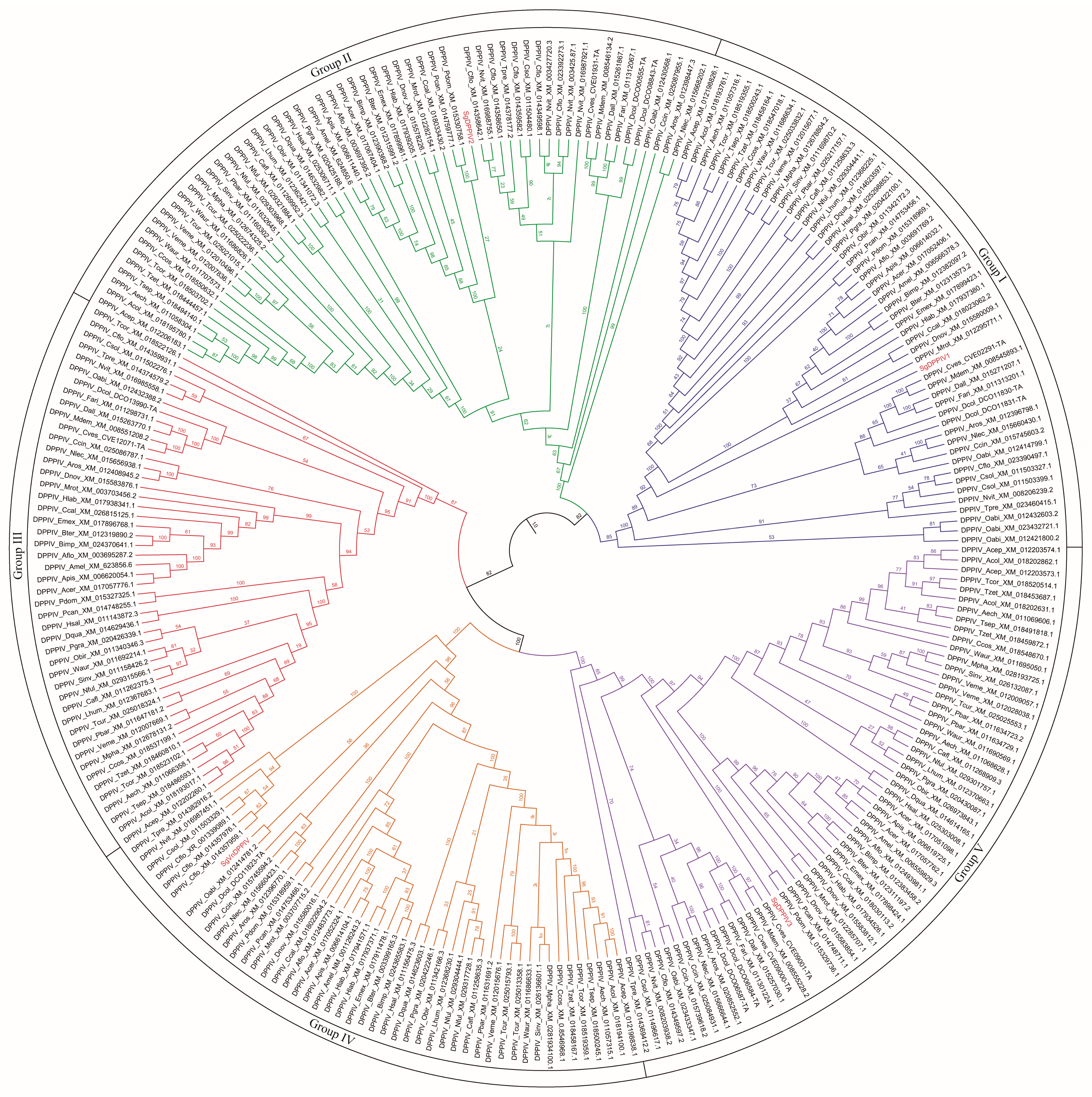
2.1. Molecular Characteristics of SgVnDPPIV
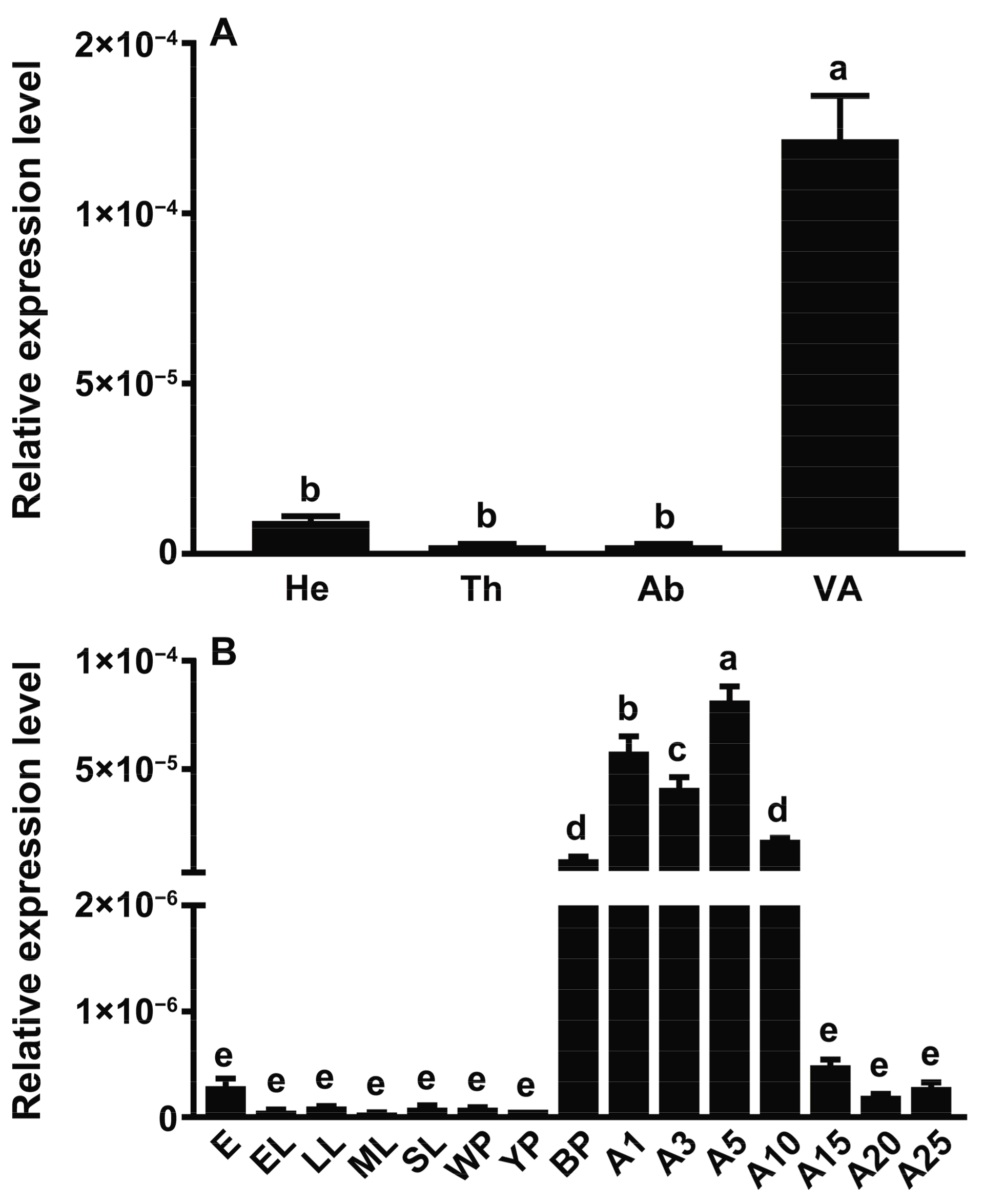
2.2. Gene Expression Pattern of SgVnDPPIV
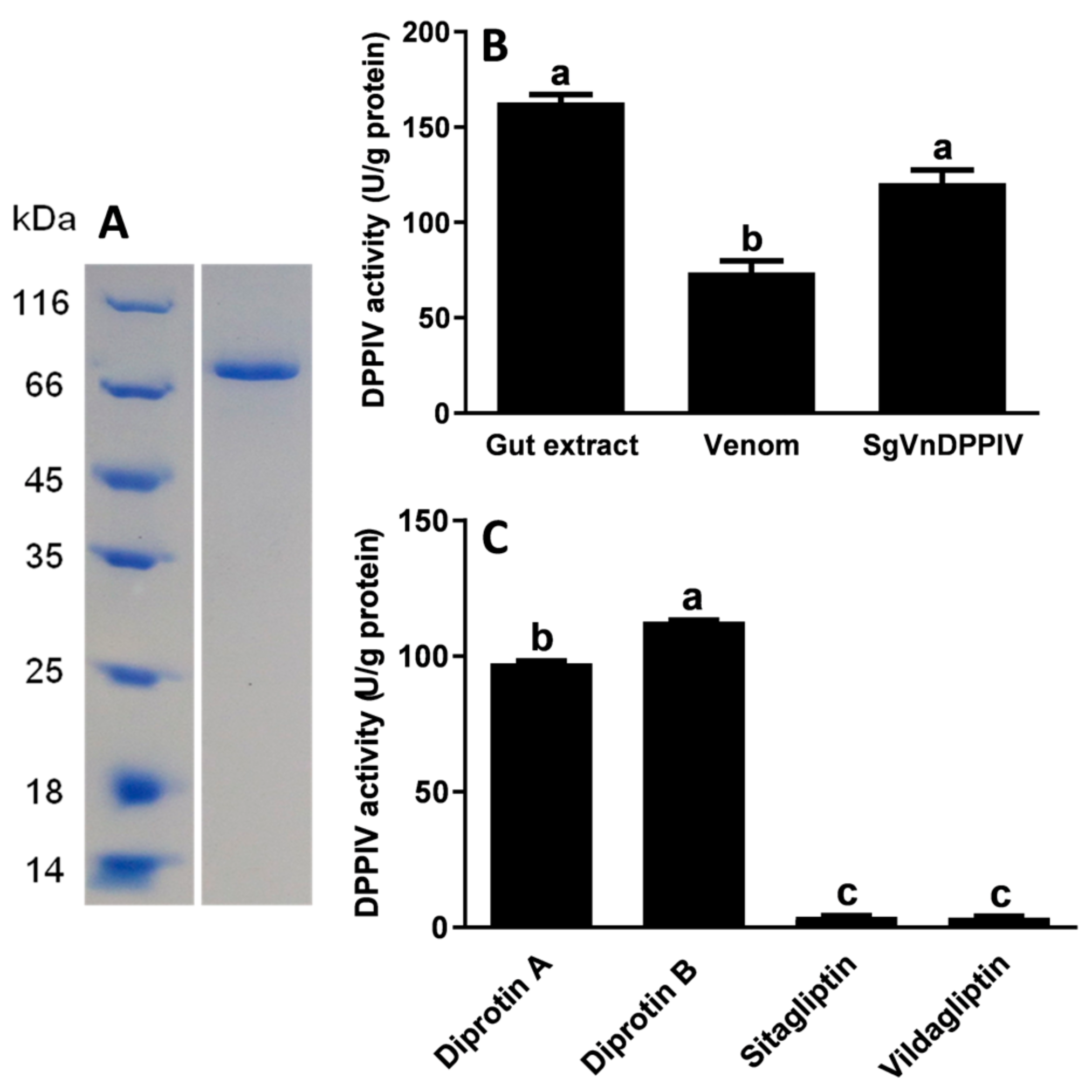
2.3. Enzymatic Activity of SgVnDPPIV
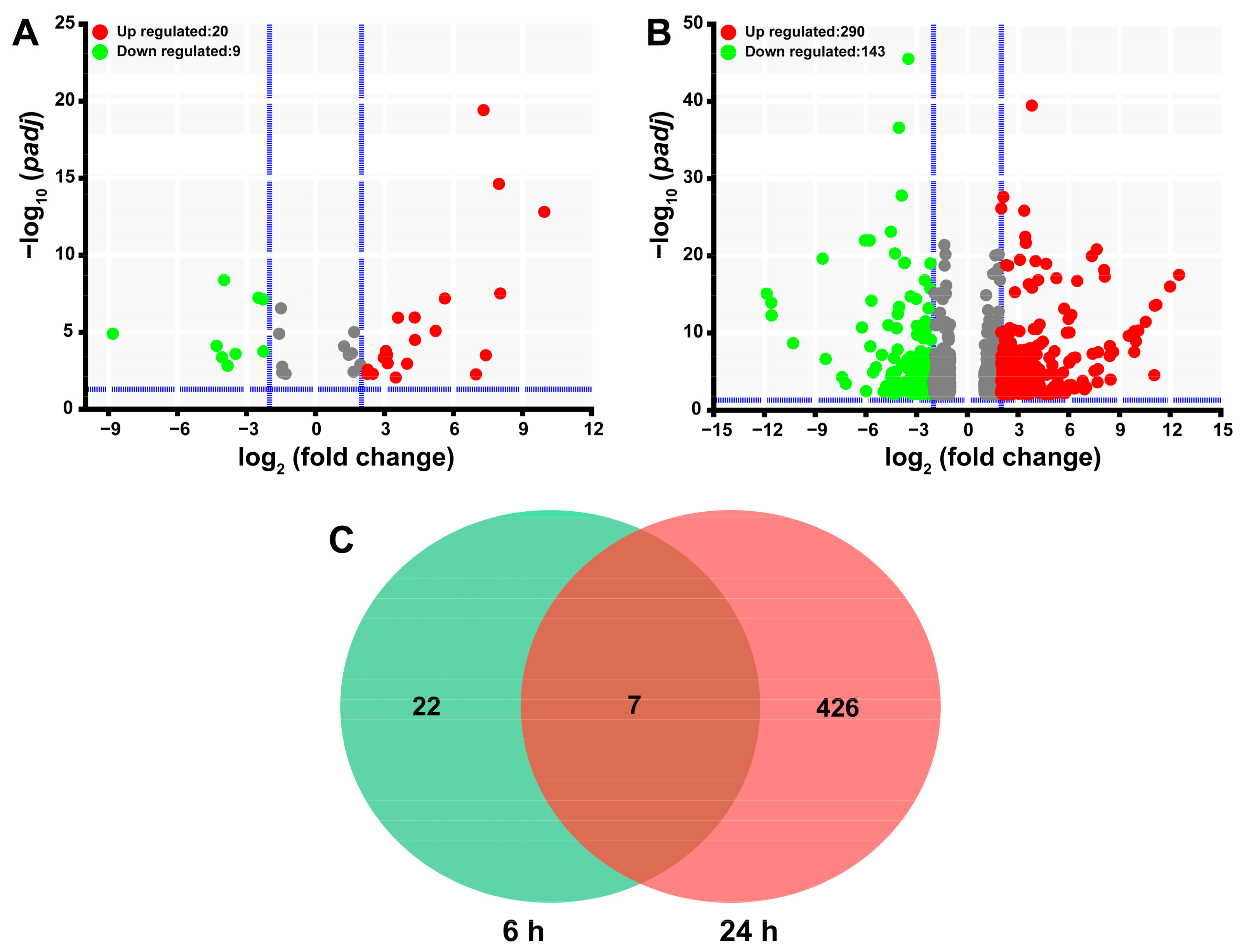
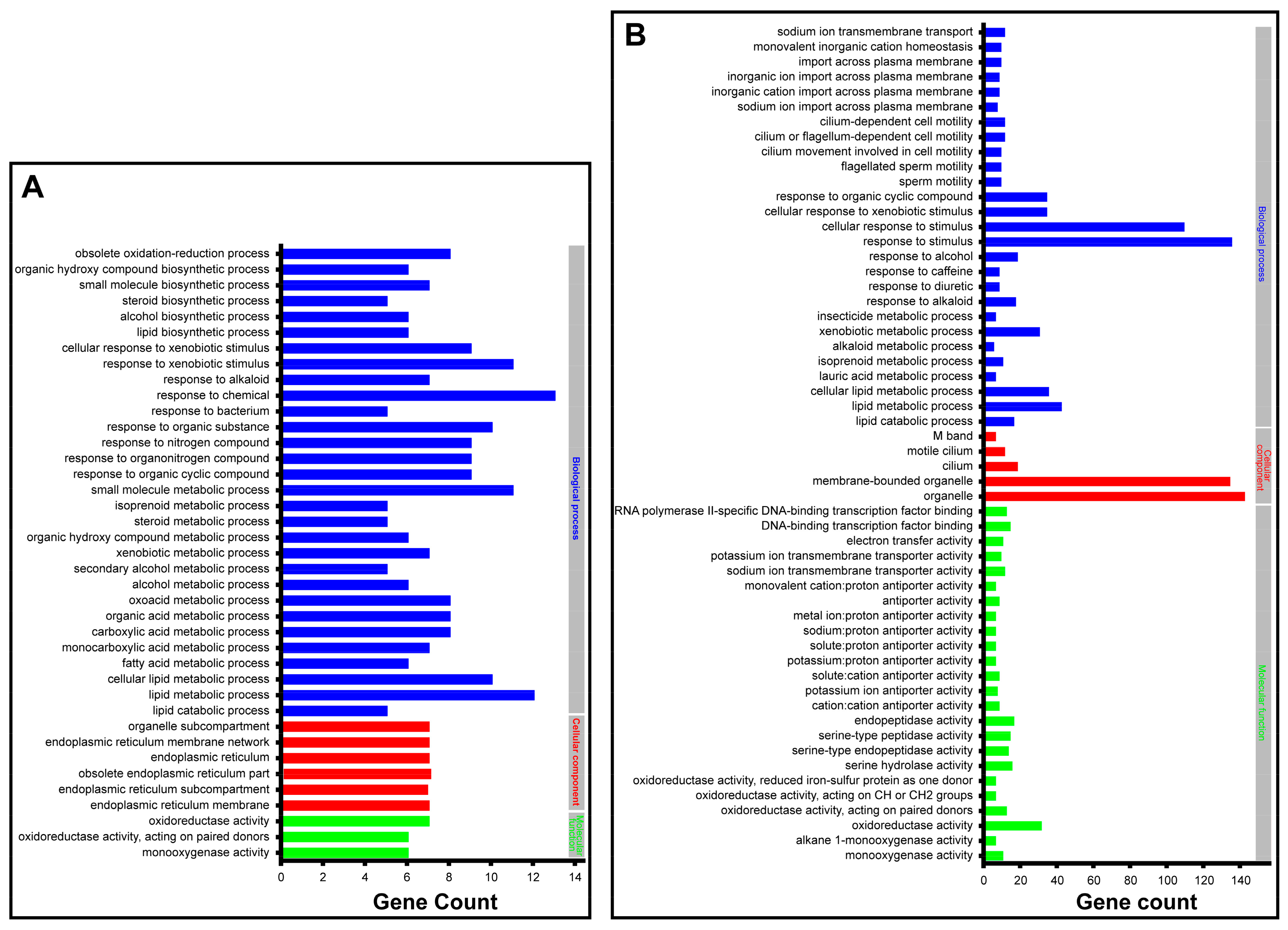
2.4. Functional Analysis of SgVnDPPIV
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insects
5.2. Gene Cloning and Sequence Analysis
5.3. Quantitative Real-Time PCR
5.4. Recombinant Production of Proteins
5.5. Enzyme Assay
5.6. Transcriptomic Sequencing and Analysis
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rawlings, N.D.; Waller, M.; Barrett, A.J.; Bateman, A. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2014, 42, D503–D509. [Google Scholar] [CrossRef] [PubMed]
- Borloo, M.; De Meester, I. Dipeptidyl peptidase IV: Development, design, synthesis and biological evaluation of inhibitors. Verh. K. Acad. Geneeskd. Belg. 1994, 56, 57–88. [Google Scholar] [PubMed]
- Tereshchenkova, V.F.; Goptar, I.A.; Kulemzina, I.A.; Zhuzhikov, D.P.; Serebryakova, M.V.; Belozersky, M.A.; Dunaevsky, Y.E.; Oppert, B.; Filippova, I.Y.; Elpidina, E.N. Dipeptidyl peptidase 4—An important digestive peptidase in Tenebrio molitor larvae. Insect Biochem. Mol. Biol. 2016, 76, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar] [CrossRef]
- Yang, Q.; Fu, B.; Luo, D.; Wang, H.B.; Cao, H.Y.; Chen, X.; Tian, L.; Yu, X.J. The multiple biological functions of dipeptidyl peptidase-4 in bone metabolism. Front. Endocrinol. 2022, 13, 856954. [Google Scholar] [CrossRef] [PubMed]
- Juillerat-Jeanneret, L. Dipeptidyl peptidase IV and its inhibitors: Therapeutics for type 2 diabetes and what else? J. Med. Chem. 2014, 57, 2197–2212. [Google Scholar] [CrossRef]
- Dávalos Terán, I.; Imai, K.; Lacroix, I.M.E.; Fogliano, V.; Udenigwe, C.C. Bioinformatics of edible yellow mealworm (Tenebrio molitor) proteome reveal the cuticular proteins as promising precursors of dipeptidyl peptidase-IV inhibitors. J. Food Biochem. 2020, 44, e13121. [Google Scholar] [CrossRef]
- Przezak, A.; Bielka, W.; Pawlik, A. Incretins in the therapy of diabetic kidney disease. Int. J. Mol. Sci. 2021, 22, 12312. [Google Scholar] [CrossRef]
- Martensen, I.; Koolman, J.; Mentlein, R. Proline-specific dipeptidyl peptidase from the blue blowfly Calliphora vicina hydrolyzes in vitro the ecdysiostatic peptide trypsin-modulating oostatic factor (Neb-TMOF). Arch. Insect Biochem. Physiol. 1998, 37, 146–157. [Google Scholar] [CrossRef]
- Nässel, D.R.; Mentlein, R.; Bollner, T.; Karlsson, A. Proline-specific dipeptidyl peptidase activity in the cockroach brain and intestine: Partial characterization, distribution, and inactivation of tachykinin-related peptides. J. Comp. Neurol. 2000, 418, 81–92. [Google Scholar] [CrossRef]
- Chihara, C.J.; Song, C.Y.; LaMonte, G.; Fetalvero, K.; Hinchman, K.; Phan, H.; Pineda, M.; Robinson, K.; Schneider, G.P. Identification and partial characterization of the enzyme of omega: One of five putative DPP IV genes in Drosophila melanogaster. J. Insect Sci. 2005, 5, 26. [Google Scholar] [PubMed]
- Blank, S.; Seismann, H.; Bockisch, B.; Braren, I.; Cifuentes, L.; McIntyre, M.; Rühl, D.; Ring, J.; Bredehorst, R.; Ollert, M.W.; et al. Identification, recombinant expression, and characterization of the 100 kDa high molecular weight Hymenoptera venom allergens Api m 5 and Ves v 3. J. Immunol. 2010, 184, 5403–5413. [Google Scholar] [PubMed]
- Jang, S.; Chung, T.Y.; Shin, J.; Lin, K.L.; Tzen, J.T.; Li, F.Y. Docking study of the precursor peptide of mastoparan onto its putative processing enzyme, dipeptidyl peptidase IV: A revisit to molecular ticketing. J. Comput. Aided Mol. Des. 2010, 24, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.S.Y.; Tu, W.C.; Jinn, T.R.; Peng, C.C.; Lin, L.J.; Tzen, J.T.C. Molecular cloning of the precursor polypeptide of mastoparan B and its putative processing enzyme, dipeptidyl peptidase IV, from the black-bellied hornet, Vespa basalis. Insect Mol. Biol. 2007, 16, 231–237. [Google Scholar] [CrossRef]
- Pretre, G.; Asturias, J.A.; Lizaso, M.T.; Tabar, A.I. Dipeptidyl peptidase IV of the Vespa velutina nigrithorax venom is recognized as a relevant allergen. Ann. Allergy Asthma Immunol. 2022, 129, 101–105. [Google Scholar]
- Schiener, M.; Hilger, C.; Eberlein, B.; Pascal, M.; Kuehn, A.; Revets, D.; Planchon, S.; Pietsch, G.; Serrano, P.; Moreno-Aguilar, C.; et al. The high molecular weight dipeptidyl peptidase IV Pol d 3 is a major allergen of Polistes dominula venom. Sci. Rep. 2018, 8, 1318. [Google Scholar]
- Yu, H.N.; Yang, H.L.; Ma, D.Y.; Lv, Y.; Liu, T.G.; Zhang, K.Y.; Lai, R.; Liu, J.Z. Vespid chemotactic peptide precursor from the wasp, Vespa magnifica (Smith). Toxicon 2007, 50, 377–382. [Google Scholar] [CrossRef]
- Chen, W.H.; Yang, X.B.; Yang, X.L.; Zhai, L.; Lu, Z.K.; Liu, J.Z.; Yu, H.N. Antimicrobial peptides from the venoms of Vespa bicolor Fabricius. Peptides 2008, 29, 1887–1892. [Google Scholar] [CrossRef]
- Mrinalini; Werren, J.H. Parasitoid wasps and their venoms. In Evolution of Venomous Animals and Their Toxins; Starr, C., Ed.; Toxinology; Springer: Dordrecht, The Netherlands, 2017; pp. 1–26. [Google Scholar]
- Beckage, N.E.; Gelman, D.B. Wasp parasitoid disruption of host development: Implications for new biologically based strategies for insect control. Annu. Rev. Entomol. 2004, 49, 299–330. [Google Scholar]
- Moreau, S.J.M.; Asgari, S. Venom proteins from parasitoid wasps and their biological functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef]
- Gatti, J.L.; Belghazi, M.; Legeai, F.; Ravallec, M.; Frayssinet, M.; Robin, S.; Aboubakar-Souna, D.; Srinivasan, R.; Tamò, M.; Poirié, M.; et al. Proteo-trancriptomic analyses reveal a large expansion of metalloprotease-like proteins in atypical venom vesicles of the wasp Meteorus pulchricornis (Braconidae). Toxins 2021, 13, 502. [Google Scholar] [PubMed]
- Moreau, S.J.M.; Guillot, S. Advances and prospects on biosynthesis, structures and functions of venom proteins from parasitic wasps. Insect Biochem. Mol. Biol. 2005, 35, 1209–1223. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ye, X.H.; Dang, C.; Cao, Y.S.; Hong, R.; Sun, Y.H.; Xiao, S.; Mei, Y.; Xu, L.; Fang, Q.; et al. Genome of the pincer wasp Gonatopus flavifemur reveals unique venom evolution and a dual adaptation to parasitism and predation. BMC Biol. 2021, 19, 145. [Google Scholar] [CrossRef] [PubMed]
- Dorémus, T.; Urbach, S.; Jouan, V.; Cousserans, F.; Ravallec, M.; Demettre, E.; Wajnberg, E.; Poulain, J.; Azéma-Dossat, C.; Darboux, I.; et al. Venom gland extract is not required for successful parasitism in the polydnavirus-associated endoparasitoid Hyposoter didymator (Hym. Ichneumonidae) despite the presence of numerous novel and conserved venom proteins. Insect Biochem. Mol. Biol. 2013, 43, 292–307. [Google Scholar] [PubMed]
- Liu, N.Y.; Xu, Z.W.; Yan, W.; Ren, X.M.; Zhang, Z.Q.; Zhu, J.Y. Venomics reveals novel ion transport peptide-likes (ITPLs) from the parasitoid wasp Tetrastichus brontispae. Toxicon 2018, 141, 88–93. [Google Scholar] [CrossRef]
- Chen, J.; Cheng, H.Z. Advances in applied research on Scleroderma spp. Chin. J. Biol. Control 2000, 16, 166–170. [Google Scholar]
- Li, X.R.; Lu, D.G.; Liu, X.X.; Zhang, Q.W.; Zhou, X.G. Ultrastructural characterization of olfactory sensilla and immunolocalization of odorant binding and chemosensory proteins from an ectoparasitoid Scleroderma guani (Hymenoptera: Bethylidae). Int. J. Biol. Sci. 2011, 7, 848–868. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Y. Deciphering the main venom components of the ectoparasitic ant-like bethylid wasp, Scleroderma guani. Toxicon 2016, 113, 32–40. [Google Scholar]
- Martinson, E.O.; Mrinalini, W.J.H.; Kelkar, Y.D.; Chang, C.H.; Werren, J.H. The evolution of venom by co-option of single-copy genes. Curr. Biol. 2017, 27, 2007–2013.e8. [Google Scholar]
- Ogawa, Y.; Mamura, Y.; Murayama, N.; Yanoshita, R. Characterization and cDNA cloning of dipeptidyl peptidase IV from the venom of Gloydius blomhoffi brevicaudus. Comp. Biochem. Physiol. 2006, 145, 35–42. [Google Scholar] [CrossRef]
- Qian, C.; Fang, Q.; Wang, L.; Ye, G.Y. Molecular cloning and functional studies of two kazal-type serine protease inhibitors specifically expressed by Nasonia vitripennis venom apparatus. Toxins 2015, 7, 2888–2905. [Google Scholar] [CrossRef] [PubMed]
- Misumi, Y.; Hayashi, Y.; Arakawa, F.; Ikehara, Y. Molecular cloning and sequence analysis of human dipeptidyl peptidase IV, a serine proteinase on the cell surface. Biochim. Biophys. Acta 1992, 1131, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Meadows, S.A.; Edosada, C.Y.; Mayeda, M.; Tran, T.; Quan, C.; Raab, H.; Wiesmann, C.; Wolf, B.B. Ala657 and conserved active site residues promote fibroblast activation protein endopeptidase activity via distinct mechanisms of transition state stabilization. Biochemistry 2007, 46, 4598–4605. [Google Scholar] [CrossRef]
- Nabeno, M.; Akahoshi, F.; Kishida, H.; Miyaguchi, I.; Tanaka, Y.; Ishii, S.; Kadowaki, T. A comparative study of the binding modes of recently launched dipeptidyl peptidase IV inhibitors in the active site. Biochem. Biophys. Res. Commun. 2013, 434, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential properties of venom peptides and proteins in solitary vs. social hunting wasps. Toxins 2016, 8, 32. [Google Scholar] [CrossRef]
- Wertheim, B. Adaptations and counter-adaptations in Drosophila host-parasitoid interactions: Advances in the molecular mechanisms. Curr. Opin. Insect Sci. 2022, 51, 100896. [Google Scholar] [CrossRef]
- Yoon, K.A.; Kim, K.; Kim, W.J.; Bang, W.Y.; Ahn, N.H.; Bae, C.H.; Yeo, J.H.; Lee, S.H. Characterization of venom components and their phylogenetic properties in some aculeate bumblebees and wasps. Toxins 2020, 12, 47. [Google Scholar] [CrossRef]
- Becchimanzi, A.; Avolio, M.; Bostan, H.; Colantuono, C.; Cozzolino, F.; Mancini, D.; Chiusano, M.L.; Pucci, P.; Caccia, S.; Pennacchio, F. Venomics of the ectoparasitoid wasp Bracon nigricans. BMC Genom. 2020, 21, 34. [Google Scholar] [CrossRef]
- Dos Santos-Pinto, J.R.A.; Perez-Riverol, A.; Lasa, A.M.; Palma, M.S. Diversity of peptidic and proteinaceous toxins from social Hymenoptera venoms. Toxicon 2018, 148, 172–196. [Google Scholar] [CrossRef]
- Martinson, E.O.; Siebert, A.L.; He, M.; Kelkar, Y.D.; Doucette, L.A.; Werren, J.H. Evaluating the evolution and function of the dynamic venom Y protein in ectoparasitoid wasps. Insect Mol. Biol. 2019, 28, 499–508. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Yang, P.; Zhang, Z.; Wu, G.X.; Yang, B. Transcriptomic immune response of Tenebrio molitor pupae to parasitization by Scleroderma guani. PLoS ONE 2013, 8, e54411. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xiong, A.S.; Yao, Q.H.; Peng, R.H.; Duan, H.; Li, X.; Fan, H.Q.; Cheng, Z.M.; Li, Y. PCR-based accurate synthesis of long DNA sequences. Nat. Protoc. 2006, 1, 791–797. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Song, Q.S.; Stanley, D. Genomic and transcriptomic analysis of Tenebrio molitor: Preface. Arch. Insect Biochem. Physiol. 2022, 111, e21969. [Google Scholar] [CrossRef]
- Eleftheriou, E.; Aury, J.M.; Vacherie, B.; Istace, B.; Orcid, C.B.; Noel, B.; Moret, Y. Chromosome-scale assembly of the yellow mealworm genome. Open Res. Eur. 2022, 1, 94. [Google Scholar] [CrossRef]
- Kaur, S.; Stinson, S.A.; DiCenzo, G.C. Whole genome assemblies of Zophobas morio and Tenebrio molitor. G3 Genes 2023, jkad079. [Google Scholar] [CrossRef]
- Li, W.Z.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Zhang, C.X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Yang, C.; Wang, Y.; Wang, J.; Zhu, J. Molecular Characterization and Functional Analysis of the Dipeptidyl Peptidase IV from Venom of the Ectoparasitoid Scleroderma guani. Toxins 2023, 15, 311. https://doi.org/10.3390/toxins15050311
Wu C, Yang C, Wang Y, Wang J, Zhu J. Molecular Characterization and Functional Analysis of the Dipeptidyl Peptidase IV from Venom of the Ectoparasitoid Scleroderma guani. Toxins. 2023; 15(5):311. https://doi.org/10.3390/toxins15050311
Chicago/Turabian StyleWu, Chaoyan, Cheng Yang, Yuqin Wang, Jun Wang, and Jiaying Zhu. 2023. "Molecular Characterization and Functional Analysis of the Dipeptidyl Peptidase IV from Venom of the Ectoparasitoid Scleroderma guani" Toxins 15, no. 5: 311. https://doi.org/10.3390/toxins15050311
APA StyleWu, C., Yang, C., Wang, Y., Wang, J., & Zhu, J. (2023). Molecular Characterization and Functional Analysis of the Dipeptidyl Peptidase IV from Venom of the Ectoparasitoid Scleroderma guani. Toxins, 15(5), 311. https://doi.org/10.3390/toxins15050311