The Reduced Folate Carrier (RFC-1) 80A>G Polymorphism and Maternal Risk of Having a Child with Down Syndrome: A Meta-Analysis


Abstract
:1. Introduction
2. Experimental Section
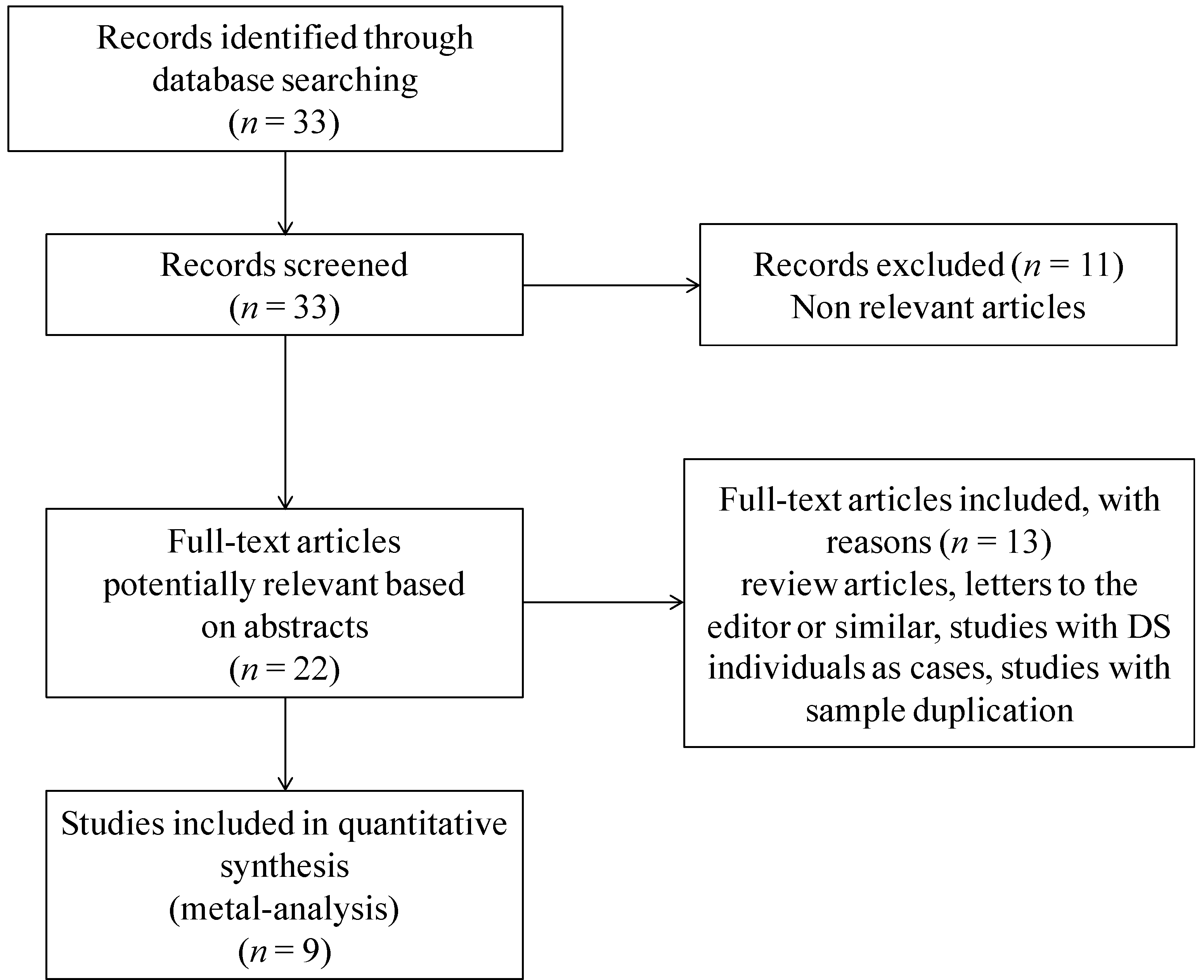
2.1. Selection of Manuscripts for Meta-Analysis

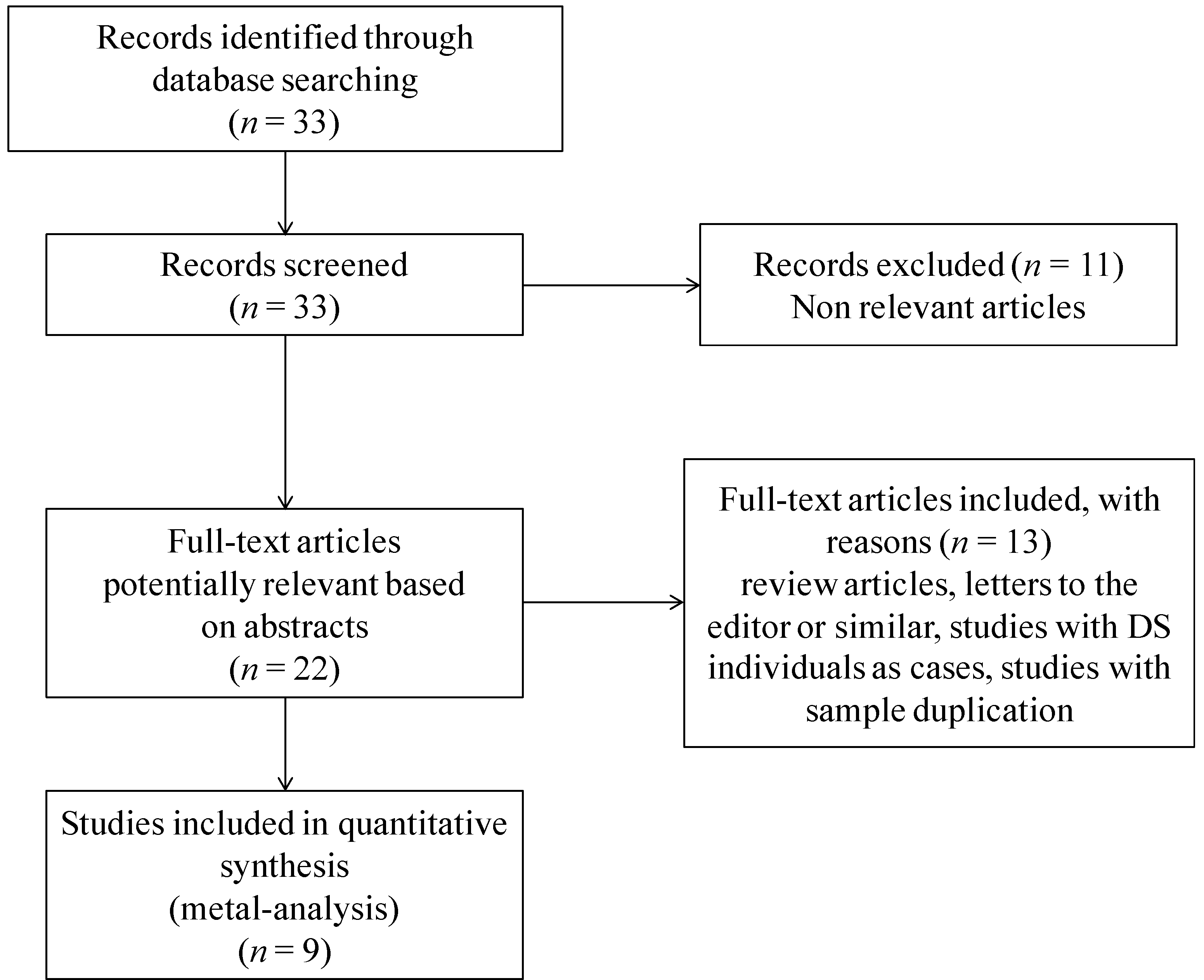{kind=link}
{kind=link}
| Author and year [Ref.] | Country | Ethnicity | MDS/MC | Age at delievry |
|---|---|---|---|---|
| Chango et al. 2005 [25] | France | white Caucasians | 119/94 a | MDS: 33.8 ± 6 years MC: 29.5 ± 6 years |
| Coppedè et al. 2006 [19] | Italy | white Caucasians | 69/93 | MDS and MC: both aged <35 years |
| Scala et al. 2006 [20] | Italy | white Caucasians | 94/263 | MDS: 32.4 ± 6.3 years MC: 30 ± 5.6 years |
| Fintelman-Rodrigues et al. 2009 [26] | Brazil | Brazilian (mixed) | 114/110 | MDS and MC: both aged <35 years |
| Brandalize et al. 2010 [24] | Brazil | Brazilian (whites) | 239/197 | MDS: 121 ≥ 35 years b MC: 29 ≥ 35 years |
| Liao et al. 2010 [23] | China | Asians | 60/68 | not available: article in Chinese |
| Neagos et al. 2010 [27] | Romania | white Caucasians | 26/46 | MDS and MC: range 20–42 years |
| Zampieri et al. 2012 [29] | Brazil | Brazilian (mixed) | 105/185 | MDS: 54 ≤ 35 years b MC: 173 ≤ 35 years |
| Wang et al. 2013 [21] | China | Asians | 104/184 | MDS and MC: both aged <35 years |
| Author and year [Ref.] | MDS Alleles | MC Alleles | MDS Genotype | MC Genotype | HW | p-Value |
|---|---|---|---|---|---|---|
| Chango et al. 2005 [25] | Allele A: 114 | Allele A: 84 | AA:24/AG:6/GG:2 | AA:16/AG:5/GG:2 | HWE yes | p = 0.24 |
| Allele G: 124 | Allele G: 104 | |||||
| Coppedè et al. 2006 [19] | Allele A: 55 | Allele A: 82 | AA:13/AG:29/GG:2 | AA:20/AG:42/GG:3 | HWE yes | p = 0.42 |
| Allele G: 83 | Allele G: 104 | |||||
| Scala et al. 2006 [20] | Allele A: 95 | Allele A: 317 | AA:27/AG:4/GG:2 | AA:102/AG:113/GG:4 | HWE yes | p = 0.09 |
| Allele G: 93 | Allele G: 209 | |||||
| Fintelman-Rodrigues et al. 2009 [26] | Allele A: 114 | Allele A: 113 | AA:25/AG:64/GG:25 | AA:29/AG:55/GG:2 | HWE yes | p = 0.99 |
| Allele G: 114 | Allele G: 107 | |||||
| Brandalize et al. 2010 [24] | Allele A: 247 | Allele A: 219 | AA:73/AG:101/GG:65 | AA:64/AG:91/GG:4 | HWE yes | p = 0.36 |
| Allele G: 231 | Allele G: 175 | |||||
| Liao et al. 2010 [23] | Allele A: 70 | Allele A: 64 | AA:24/AG:22/GG:14 | AA:12/AG:40/GG:16 | HWE yes | p = 0.14 |
| Allele G: 50 | Allele G: 72 | |||||
| Neagos et al. 2010 [27] | Allele A: 18 | Allele A: 40 | AA:1/AG:16/GG:9 | AA:5/AG:30/GG:11 | HWE no | p = 0.02 |
| Allele G: 34 | Allele G: 52 | |||||
| Zampieri et al. 2012 [29] | Allele A: 106 | Allele A: 194 | AA:29/AG:48/GG:28 | AA:53/AG:88/GG:44 | HWE yes | p = 0.53 |
| Allele G: 104 | Allele G: 176 | |||||
| Wang et al. 2013 [21] | Allele A: 135 | Allele A: 271 | AA:47/AG:41/GG:16 | AA:100/AG:71/GG:13 | HWE yes | p = 0.94 |
| Allele G: 73 | Allele G: 97 |
2.2. Statistical Analysis
3. Results
| Genetic Model | Studies | Fixed effects | p-value | Random effects | p-value | Q-statistics | p-value a | I2 |
|---|---|---|---|---|---|---|---|---|
| OR (CI95%) | OR (CI 95%) | |||||||
| G versus A | All | 1.14 | 0.032 | 1.14 | 0.121 | 12.56 | 0.128 | 2.0% |
| (1.01–1.30) | (0.97–1.33) | |||||||
| HWE | 1.14 | 0.047 | 1.12 | 0.187 | 12.12 | 0.097 | 2.0% | |
| (1.00–1.29) | (0.95–1.33) | |||||||
| HWE, no M b | 1.17 | 0.019 | 1.16 | 0.105 | 10.17 | 0.118 | 2.0% | |
| (1.02–1.34) | (0.97–1.39) | |||||||
| Dominant | All | 1.12 | 0.243 | 1.09 | 0.526 | 13.74 | 0.089 | 7.0% |
| (0.92–1.36) | (0.83–1.43) | |||||||
| HWE | 1.11 | 0.279 | 1.07 | 0.612 | 12.95 | 0.073 | 7.0% | |
| (0.91–1.35) | (0.81–1.42) | |||||||
| HWE, | 1.14 | 0.194 | 1.11 | 0.516 | 12.09 | 0.06 | 8.0% | |
| no M b | (0.93–1.40) | (0.82–1.50) | ||||||
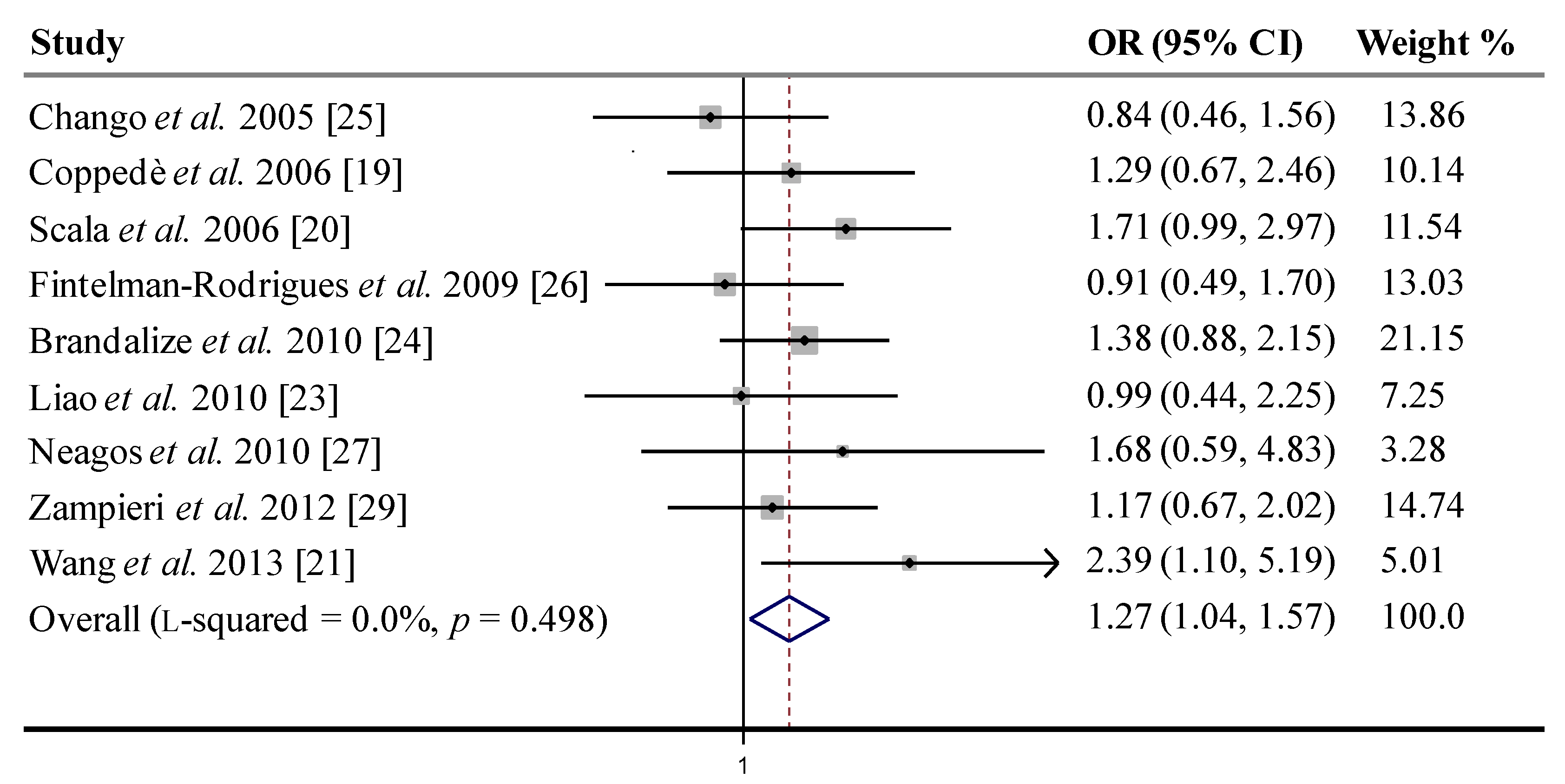
| Recessive | All | 1.27 | 0.020 | 1.27 | 0.020 | 7.37 | 0.5 | 0.0% |
| (1.04–1.57) | (1.04–1.57) | |||||||
| HWE | 1.26 | 0.030 | 1.26 | 0.031 | 7.09 | 0.42 | 0.0% | |
| (1.02–1.55) | (1.02–1.56) | |||||||
| HWE, | 1.33 | 0.012 | 1.33 | 0.012 | 5.21 | 0.517 | 0.0% | |
| no M b | (1.06–1.66) | (1.07–1.67) |
4. Discussion
Limitations
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Bailey, L.B.; Gregory, J.F., III. Folate metabolism and requirements. J. Nutr. 1999, 129, 779–782. [Google Scholar]
- Zhao, R.; Matherly, L.H.; Goldman, D.I. Membrane transporters and folate homeostasis: Intestinal absorption and transport into systemic compartments and tissues. Expert Rev. Mol. Med. 2009, 11, e4. [Google Scholar] [CrossRef]
- Fenech, M. The role of folic acid and vitamin B12 in genomic stability of human cells. Mutat. Res. 2001, 475, 57–67. [Google Scholar] [CrossRef]
- Stanger, O. Physiology of folic acid in health and disease. Curr. Drug Metab. 2002, 3, 211–223. [Google Scholar] [CrossRef]
- Antonarakis, S.E. Down syndrome. In Principle of Molecular Medicine; Jameson, J.L., Ed.; Humana Press Inc.: Totowa, NJ, USA, 1998; pp. 1069–1078. [Google Scholar]
- Lamb, N.E.; Freeman, S.B.; Savage-Austin, A.; Pettay, D.; Taft, L.; Hersey, J.; Gu, Y.; Shen, J.; Saker, D.; May, K.M.; et al. Susceptible chiasmate configurations of chromosome 21 predispose to non-disjunction in both maternal meiosis I and meiosis II. Nat. Genet. 1996, 14, 400–405. [Google Scholar] [CrossRef]
- Morris, J.K.; Mutton, D.E.; Alberman, E. Revised estimates of the maternal age specific live birth prevalence of Down’s syndrome. J. Med. Screen. 2002, 9, 2–6. [Google Scholar] [CrossRef]
- Melamed-Bessudo, C.; Levy, A.A. Deficiency in DNA methylation increases meiotic crossover rates in euchromatic but not in heterochromatic regions in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, E981–E988. [Google Scholar] [CrossRef]
- Yelina, N.E.; Choi, K.; Chelysheva, L.; Macaulay, M.; de Snoo, B.; Wijnker, E.; Miller, N.; Drouaud, J.; Grelon, M.; Copenhaver, G.P.; et al. Epigenetic remodeling of meiotic crossover frequency in Arabidopsis thaliana DNA methyltransferase mutants. PLoS Genet. 2012, 8, e1002844. [Google Scholar] [CrossRef]
- Colomé-Tatché, M.; Cortijo, S.; Wardenaar, R.; Morgado, L.; Lahouze, B.; Sarazin, A.; Etcheverry, M.; Martin, A.; Feng, S.; Duvernois-Berthet, E.; et al. Features of the Arabidopsis recombination landscape resulting from the combined loss of sequence variation and DNA methylation. Proc. Natl. Acad. Sci. USA 2012, 109, 16240–16245. [Google Scholar] [CrossRef]
- James, S.J.; Pogribna, M.; Pogribny, I.P.; Melnyk, S.; Hine, R.J.; Gibson, J.B.; Yi, P.; Tafoya, D.L.; Swenson, D.H.; Wilson, V.L.; et al. Abnormal folate metabolism and mutation in the methylenetetrahydrofolate reductase gene may be maternal risk factors for Down syndrome. Am. J. Clin. Nutr. 1999, 70, 495–501. [Google Scholar]
- Wang, X.; Thomas, P.; Xue, J.; Fenech, M. Folate deficiency induces aneuploidy in human lymphocytes in vitro-evidence using cytokinesis-blocked cells and probes specific for chromosomes 17 and 21. Mutat. Res. 2004, 551, 167–180. [Google Scholar] [CrossRef]
- Beetstra, S.; Thomas, P.; Salisbury, C.; Turner, J.; Fenech, M. Folic acid deficiency increases chromosomal instability, chromosome 21 aneuploidy and sensitivity to radiation-induced micronuclei. Mutat. Res. 2005, 578, 317–326. [Google Scholar] [CrossRef]
- Coppedè, F. The complex relationship between folate/homocysteine metabolism and risk of Down syndrome. Mutat. Res. 2009, 682, 54–70. [Google Scholar] [CrossRef]
- Medica, I.; Maver, A.; Augusto, G.F.; Peterlin, B. Polymorphisms in genes involved in folate metabolism as maternal risk factors for Down syndrome—Meta-analysis. Cent. Eur. J. Med. 2009, 4, 395–408. [Google Scholar] [CrossRef]
- Amorim, M.R.; Lima, M.A. MTRR 66A>G polymorphism as maternal risk factor for Down Syndrome: A meta-analysis. Genet. Test. Mol. Biomarkers 2013, 17, 69–73. [Google Scholar] [CrossRef]
- Wu, X.; Wang, X.; Chan, Y.; Jia, S.; Luo, Y.; Tang, W. Folate metabolism gene polymorphisms MTHFR C677T and A1298C and risk for Down syndrome offspring: A meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 167, 154–159. [Google Scholar] [CrossRef]
- Chango, A.; Emery-Fillon, N.; de Courcy, G.P.; Lambert, D.; Pfister, M.; Rosenblatt, D.S.; Nicolas, J.P. A polymorphism (80G>A) in the reduced folate carrier gene and its associations with folate status and homocysteinemia. Mol. Genet. Metab. 2000, 70, 310–315. [Google Scholar] [CrossRef]
- Coppedè, F.; Marini, G.; Bargagna, S.; Stuppia, L.; Minichilli, F.; Fontana, I.; Colognato, R.; Astrea, G.; Palka, G.; Migliore, L. Folate gene polymorphisms and the risk of Down syndrome pregnancies in young Italian women. Am. J. Med. Genet. A 2006, 140, 1083–1091. [Google Scholar]
- Scala, I.; Granese, B.; Sellitto, M.; Salomè, S.; Sammartino, A.; Pepe, A.; Mastroiacovo, P.; Sebastio, G.; Andria, G. Analysis of seven maternal polymorphisms of genes involved in homocysteine/folate metabolism and risk of Down syndrome offspring. Genet. Med. 2006, 8, 409–416. [Google Scholar] [CrossRef]
- Wang, S.S.; Wang, C.; Qiao, F.Y.; Lv, J.J.; Feng, L. Polymorphisms in genes RFC-1/CBS as maternal risk factors for Down syndrome in China. Arch. Gynecol. Obstet. 2013, in press. [Google Scholar]
- Biselli, J.M.; Goloni-Bertollo, E.M.; Zampieri, B.L.; Haddad, R.; Eberlin, M.N.; Pavarino-Bertelli, E.C. Genetic polymorphisms involved in folate metabolism and elevated plasma concentrations of homocysteine: Maternal risk factors for Down syndrome in Brazil. Genet. Mol. Res. 2008, 7, 33–42. [Google Scholar] [CrossRef]
- Liao, Y.P.; Bao, M.S.; Liu, C.Q.; Liu, H.; Zhang, D. Folate gene polymorphism and the risk of Down syndrome pregnancies in young Chinese women. Yi Chuan 2010, 32, 461–466. [Google Scholar] [CrossRef]
- Brandalize, A.P.; Bandinelli, E.; Dos Santos, P.A.; Schüler-Faccini, L. Maternal gene polymorphisms involved in folate metabolism as risk factors for Down syndrome offspring in Southern Brazil. Dis. Markers 2010, 29, 95–101. [Google Scholar] [CrossRef]
- Chango, A.; Fillon-Emery, N.; Mircher, C.; Bléhaut, H.; Lambert, D.; Herbeth, B.; James, S.J.; Réthoré, M.O.; Nicolas, J.P. No association between common polymorphisms in genes of folate and homocysteine metabolism and the risk of Down’s syndrome among French mothers. Br. J. Nutr. 2005, 94, 166–169. [Google Scholar] [CrossRef]
- Fintelman-Rodrigues, N.; Corrêa, J.C.; Santos, J.M.; Pimentel, M.M.; Santos-Rebouças, C.B. Investigation of CBS, MTR, RFC-1 and TC polymorphisms as maternal risk factors for Down syndrome. Dis. Markers 2009, 26, 155–161. [Google Scholar] [CrossRef]
- Neagos, D.; Cretu, R.; Tutulan-Cunita, A.; Stoian, V.; Bohiltea, L.C. RFC-1 gene polymorphism and the risk of Down syndrome in romanian population. Maedica (Buchar) 2010, 5, 280–285. [Google Scholar]
- Stanisławska-Sachadyn, A.; Mitchell, L.E.; Woodside, J.V.; Buckley, P.T.; Kealey, C.; Young, I.S.; Scott, J.M.; Murray, L.; Boreham, C.A.; McNulty, H.; et al. The reduced folate carrier (SLC19A1) c.80G>A polymorphism is associated with red cell folate concentrations among women. Ann. Hum. Genet. 2009, 73, 484–491. [Google Scholar] [CrossRef]
- Zampieri, B.L.; Biselli, J.M.; Goloni-Bertollo, E.M.; Vannucchi, H.; Carvalho, V.M.; Cordeiro, J.A.; Pavarino, E.C. Maternal risk for Down syndrome is modulated by genes involved in folate metabolism. Dis. Markers 2012, 32, 73–81. [Google Scholar] [CrossRef]
- Lubec, G.; Bajo, M.; Cheon, M.S.; Bajova, H.; Matherly, L.H. Increased expression of human reduced folate carrier in fetal Down syndrome brain. J. Neural Transm. Suppl. 2003, 67, 95–103. [Google Scholar] [CrossRef]
- Höger, J.; Patterson, D.; Höger, H.; Shim, K.S.; Bubna-Littitz, H.; Lubec, G. Mice transgenic for reduced folate carrier: An animal model of Down syndrome? Amino Acids 2009, 36, 349–357. [Google Scholar]
- Locke, A.E.; Dooley, K.J.; Tinker, S.W.; Cheong, S.Y.; Feingold, E.; Allen, E.G.; Freeman, S.B.; Torfs, C.P.; Cua, C.L.; Epstein, M.P.; et al. Variation in folate pathway genes contributes to risk of congenital heart defects among individuals with Down syndrome. Genet. Epidemiol. 2010, 34, 613–623. [Google Scholar] [CrossRef]
- Xavier, A.C.; Ge, Y.; Taub, J. Unique clinical and biological features of leukemia in Down syndrome children. Expert Rev. Hematol. 2010, 3, 175–186. [Google Scholar] [CrossRef]
- Biselli, J.M.; Brumati, D.; Frigeri, V.F.; Zampieri, B.L.; Goloni-Bertollo, E.M.; Pavarino-Bertelli, E.C. A80G polymorphism of reduced folate carrier 1 (RFC1) and C776G polymorphism of transcobalamin 2 (TC2) genes in Down’s syndrome etiology. Sao Paulo Med. J. 2008, 126, 329–332. [Google Scholar] [CrossRef]
- Mantel, N.; Haenszel, W. Statistical aspect of the analysis of data from retrospective studies of disease. J. Natl. Cancer Inst. 1959, 22, 719–748. [Google Scholar]
- DerSimonian, R.; Laird, N. Meta-analysis in clinical trials. Control. Clin. Trials 1986, 7, 177–188. [Google Scholar] [CrossRef]
- Higgins, J.P.; Thompson, S.J. Quantifying heterogeneity in a meta-analysis. Stat. Med. 2002, 21, 1539–1558. [Google Scholar] [CrossRef]
- Huedo-Medina, T.B.; Sanchez-Meca, J.; Martin-Martinez, F.; Botella, J. Assessing heterogeneity in meta-analysis: Q statistics or I2 index? Psychol. Methods 2006, 11, 193–202. [Google Scholar] [CrossRef]
- Egger, M.; Smith, G.D. Metaanalysis. Potentials and promise. BMJ 1997, 315, 1371–1374. [Google Scholar] [CrossRef]
- DerSimonian, R.; Kacker, R. Random-effects model for meta-analysis of clinical trials: An update. Contemp. Clin. Trials 2007, 28, 105–114. [Google Scholar] [CrossRef]
- Hollis, N.D.; Allen, E.G.; Oliver, T.R.; Tinker, S.W.; Druschel, C.; Hobbs, C.A.; O’Leary, L.A.; Romitti, P.A.; Royle, M.H.; Torfs, C.P.; et al. Preconception folic acid supplementation and risk for chromosome 21 nondisjunction: A report from the National Down Syndrome Project. Am. J. Med. Genet. A 2013, 161, 438–444. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Coppedè, F.; Lorenzoni, V.; Migliore, L. The Reduced Folate Carrier (RFC-1) 80A>G Polymorphism and Maternal Risk of Having a Child with Down Syndrome: A Meta-Analysis. Nutrients 2013, 5, 2551-2563. https://doi.org/10.3390/nu5072551
Coppedè F, Lorenzoni V, Migliore L. The Reduced Folate Carrier (RFC-1) 80A>G Polymorphism and Maternal Risk of Having a Child with Down Syndrome: A Meta-Analysis. Nutrients. 2013; 5(7):2551-2563. https://doi.org/10.3390/nu5072551
Chicago/Turabian StyleCoppedè, Fabio, Valentina Lorenzoni, and Lucia Migliore. 2013. "The Reduced Folate Carrier (RFC-1) 80A>G Polymorphism and Maternal Risk of Having a Child with Down Syndrome: A Meta-Analysis" Nutrients 5, no. 7: 2551-2563. https://doi.org/10.3390/nu5072551
APA StyleCoppedè, F., Lorenzoni, V., & Migliore, L. (2013). The Reduced Folate Carrier (RFC-1) 80A>G Polymorphism and Maternal Risk of Having a Child with Down Syndrome: A Meta-Analysis. Nutrients, 5(7), 2551-2563. https://doi.org/10.3390/nu5072551