Abstract
Vitamin D (vitD) deficiency (25-hydroxy-vitamin D < 50 nmol/L) is common in pregnancy and associated with an increased risk of adverse pregnancy outcomes. High-dose vitD supplementation is suggested to improve pregnancy health, but there is limited knowledge about the effects on placental vitD transport and metabolism and the vitD status of newborns. Comparing the current standard maternal supplementation, 10 µg/day to a 90 µg vitD supplement, we investigated placental gene expression, maternal vitD transport and neonatal vitD status. Biological material was obtained from pregnant women randomized to 10 µg or 90 µg vitD supplements from week 11–16 onwards. Possible associations between maternal exposure, neonatal vitD status and placental expression of the vitD receptor (VDR), the transporters (Cubilin, CUBN and Megalin, LRP2) and the vitD-activating and -degrading enzymes (CYP24A1, CYP27B1) were investigated. Maternal vitD-binding protein (VDBP) was determined before and after supplementation. Overall, 51% of neonates in the 10 µg vitD group were vitD-deficient in contrast to 11% in the 90 µg group. High-dose vitD supplementation did not significantly affect VDBP or placental gene expression. However, the descriptive analyses indicate that maternal obesity may lead to the differential expression of CUBN, CYP24A1 and CYP27B1 and a changed VDBP response. High-dose vitD improves neonatal vitD status without affecting placental vitD regulation.
Keywords:
neonates; obesity; placenta; vitamin D; vitamin D-binding protein; vitamin D metabolism; pregnancy 1. Introduction
A sufficient vitamin D (vitD) supply during pregnancy is pivotal for both maternal, fetal and neonatal health. Over the years, observational studies have repeatedly linked maternal vitD deficiency to various adverse pregnancy outcomes like gestational diabetes, pre-eclampsia and fetal growth restriction [1,2,3,4,5]. In offspring, an insufficient vitD level in pregnancy is not only related to negative effects on bone development; numerous studies have also shown an increased risk of long-term health problems, such as diabetes, asthma, overweight and neurodevelopmental disorders [6,7,8,9].
The maternal blood stream is the sole source of vitD for the growing fetus. Maternal and placental factors are closely entangled and may affect fetal vitD supply. Within the placenta, active vitD, i.e., 1,25-dihydroxy-vitamin D (1,25(OH)2D), binds to the intracellular vitD receptor (VDR) [10], involved in the downstream regulation of numerous genes and physiological processes [11]. This presents a potential connection between maternal vitD status and placenta-associated pregnancy complications.
Placental uptake of vitD [12] is believed to mimic the active renal uptake of vitD [13,14], primarily facilitated by the megalin/cubilin complex, encoded by the LRP2 and CUBN genes. The pro-hormone 25-hydroxy-vitamin D (25(OH)D) is transported in the blood stream, mostly bound to vitD-binding protein (VDBP) or, to a lesser extent, albumin [15]. Once reaching the placenta, VDBP-bound 25(OH)D is internalized by receptor-mediated endocytosis in the decidual cells and the syncytiotrofoblasts [12,16]. To reach the fetus, 25(OH)D crosses the placenta [16] and is activated by transformation into 1,25(OH)2D in the fetal kidney and in the fetal part of the placenta. Likewise, 25(OH)D may accumulate in the placenta or metabolize into either the active metabolite 1,25(OH)2D or the inactive metabolite 24,25(OH)2D through the interaction with CYP27B1 or CYP24A1, respectively, as both enzymes are present in the placental tissue [17,18,19,20].
In most countries, including Denmark, USA and UK, health authorities recommend a 10 µg/day (400 IU/day) vitD supplement during pregnancy [21,22]. Despite this recommendation, vitD deficiency (25(OH)D < 50 nmol/L) continues to be a global problem with a prevalence ranging from 46 to 87% [23,24] in various pregnant populations. Moreover, current recommendations often fail to consider maternal factors directly interfering with the supply of vitD to the fetus. Notably, obesity, a common condition in pregnancy today [25,26], is known to increase maternal vitD deficiency [27,28]. This is believed to be due to the sequestering of vitamin D in fat tissue, due to the fat-soluble property of this vitamin, resulting in a decreased amount of vitamin D in the bloodstream [29]. In our previous study, we found that placental CYP24A1 gene expression increased with increasing maternal body weight [30]. Furthermore, it was previously reported that umbilical cord concentrations of 25(OH)D are affected by a high maternal body mass index (BMI) [31]. Though data are sparse, this underlines that a personalized approach may be needed to optimize nutrition in risk pregnancies such as those affected by maternal obesity [32]. The safety of markedly increasing the daily vitD supplementation up to 110 µg has been shown by others [33,34,35]. However, no previous supplementation study has focused on the impact of increased doses of maternal vitD supplementation on the placenta in a clinical setting and whether such placental factors could alter the amount of 25(OH)D reaching the fetus.
The aim of the present study was thus to investigate how increasing vitD supplementation to 90 µg/day during the last six months of pregnancy affects the umbilical cord blood concentration of 25(OH)D and placental vitD regulation, also considering if maternal pre-pregnancy BMI could be an independent factor affecting placental vitD metabolism.
2. Materials and Methods
2.1. Study Population
This study is a part of the randomized double-blinded trial GRAVITD registered at ClinicalTrial.gov on 17 February 2020 (NCT04291313). The study population and enrolment process are described in detail elsewhere [36]. In short, participants were invited during their first prenatal visit at the hospital (a free-of-charge nuchal translucency scan offered to all pregnant women in Denmark) in gestational week (GW) 11–16. Upon inclusion, participants were randomly allocated 1:1 to either 90 µg or 10 µg of vitD3 with even distribution over seasons. Exclusion criteria were age < 18 years, incapability to give written informed consent, disturbances in calcium metabolism, chronic kidney disease or a pre-existing high-dose (>10 µg/day) vitD supplementation initiated by a physician.
For all GRAVITD participants, serum samples were collected upon enrolment, i.e., GW 11–16. From approximately half of all participants, an additional sample was drawn in GW 24–38, at least three months apart from the first blood sample.
Of the 118 participants from whom placental samples were collected for this study (see Section 2.2), maternal blood samples from the first trimester (GW 11–16) were available from 113 pregnancies. For a smaller subgroup, blood samples from the third trimester (GW 29–38, n = 34) were available.
2.2. Placental Collection
Placental samples were obtained within the period of February 2021 to March 2022. Only placentas from singleton pregnancies not diagnosed with pre-eclampsia, gestational diabetes or intrauterine growth restriction were used for the placental analysis. Placentas were placed at 5 °C shortly after delivery. To be eligible, tissue had to be sampled within 5 h after delivery, in order to ensure sufficient tissue quality without interfering with clinical work at the maternity ward, leaving a total of 118 placental samples (Supplementary Table S2) [37] for this study. In short, a medial villous sample from a randomly selected slice of the placenta was collected, rinsed in isotonic saline, placed in RNAlater and stored at −20 °C until analysis.
2.3. Umbilical Cord Blood
Umbilical cord blood was drawn in EDTA tubes by a midwife and placed at 5 °C shortly after delivery. Plasma samples were eligible for this study if prepared and snap-frozen within 18 hours by the research team. All samples were kept at −80 °C until analysis. Not distinguishing between healthy and diseased pregnancies, a total of 472 umbilical cord blood samples were available for the present study.
2.4. 25(OH)D Analysis
The concentration of 25(OH)D in maternal-serum samples and cord blood plasma samples were determined at the Department of Clinical Biochemistry at Aarhus University Hospital, Denmark, using high-performance liquid chromatography coupled with tandem mass spectrometry, which is considered the golden standard of 25(OH)D assays [38].
2.5. VDBP Analysis
For a subgroup of participants (n = 101), the VDBP content of maternal blood was determined using a commercially available enzyme-linked immunosorbent assay kit (Cloud-Clone Corp, Houston, TX, USA) according to the manufacturer’s protocol. Samples were diluted 1:200,000. To avoid an undue influence from differences related to gestational age [39], only dyads (early/late pregnancy) of maternal blood samples that could fulfil strict criteria regarding sampling time were used, i.e., to be eligible for VDBP measures, the first-trimester sample must have been collected at GW 12 + 0–13 + 4, and the third-trimester sample had to be obtained at GW 34 + 0–35 + 6.
2.6. RNA Extraction
Total RNA was purified from 30 mg of homogenized villous placental tissue using an RNeasy Mini Kit (cat. no. 74104, Qiagen, Germantown, MD, USA) according to the manufacturer’s protocol. RNA concentrations were quantified by an absorbance measurement using a NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA), and the quality of the extracted RNA was evaluated by agarose gel electrophoresis. cDNA was synthesized from 1000 ng of purified RNA using an ImProm™ Reverse Transcription System kit (Promega, Madison, WI, USA) with an oligoDt primer.
2.7. qPCR
The placental gene expression of vitD-related genes was assessed by qPCR specifically targeting genes encoding vitD transporters CUBN and LRP2, vitD-activating and -degrading enzymes CYP27B1 and CYP24A1 and the vitD receptor VDR.
For CUBN, LRP2 and VDR, gene expression analyses were performed using Fast start SYBR green (cat. no. 4913914001, Roche, Basel, Switzerland) and the primers shown in Table 1 on a Light-Cycler 480 II and standardized to the geomean of CycA and HPRT. For CYP24A1 and CYP27B1, gene expression analyses were performed using TaqMan Gene Expression (cat. no. 4369016, Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) and the probes shown in Table 1 on a QuantStudio 7 Flex (Applied biosystems) standardized to HPRT. Prior to statistical analysis, ΔCT (cycle threshold) values were converted into 2(−ΔCT) values.

Table 1.
Primers and probes used for qPCR.
Only samples fulfilling all the technical requirements for evaluation were included, leaving 107 data points for the evaluation of CUBN, 114 data points for LRP2 and 110 data points for VDR. In the CYP27B1 qPCR, the gene expression was below the detection limit in 46 samples. For these samples, the mean CT value of CYP27B1 was set to be 37 in the statistical analysis as this was the detection limit.
2.8. Statistics and Descriptive Analysis
All statistical analyses comparing the vitD dosing groups were performed as an intention to treat based on the randomization to 10 µg or 90 µg vitD supplementation. The dosing groups were compared using a Student’s t-test for the assessment of continuous variables when comparing two groups and a one-way ANOVA when comparing more than two groups. A chi-squared test was used for categorical variables. Continuous data were analyzed for a Gaussian distribution using QQ-plots, and a logarithmic transformation of the data was performed when appropriate.
In addition to the analysis based on the dosing groups, we investigated the possible associations with maternal 25(OH)D concentration using linear correlation analyses. Potential homoscedasticity was evaluated by the visual inspection of residual plots, and the normality of residuals was considered by the visual inspection of residual QQ plots.
We further described how women with pre-pregnancy overweight or obesity reacted to the vitD supplementation, using a spline model approach, stratifying according to the vitD dosing group. Spline models were constructed based on four not-prespecified knots in each model. The spline models were used for descriptive analysis only in order to visualize any potential association between concentration or expression and BMI in the two dosing groups. Spline models serve as a complement to categorical analyses and linear regression models and give a visual of the association within different intervals accommodating the complex nature of human biology. They are piecewise polynomials defined by the knots and smoothened to create a curve [40].
All analyses were performed using STATA (version 18, StataCorp, College Station, TX, USA) and Prism (version 10.2.2, GraphPad Software, Boston, MA, USA). A p-value < 0.05 was considered statistically significant.
3. Results
3.1. Placental Expression of vitD-Related Genes
Prior to comparing the placental gene expression of the two dosing groups, we investigated the pregnancy characteristics of each group, finding no differences in the distribution of offspring sex or overall maternal demographics, except for a slight difference in mean gestational age at delivery (10 µg: 40.2 weeks vs. 90 µg: 39.8 weeks, p = 0.031) (Supplementary Table S2).
Investigating the gene expression of the vitD-related genes CUBN, LRP2, CYP24A1, CYP27B1 and VDR, we found no statistically significant difference between the two vitD dosing groups (Figure 1a–e).
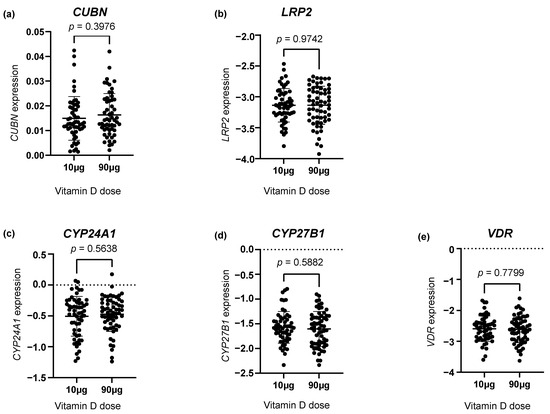
Figure 1.
The placental gene expression of (a) CUBN, (b) LRP2, (c) CYP24A1, (d) CYP27B1 and (e) VDR in the two vitamin D dosing groups (10 µg vs. 90 µg). The error bars in the figure illustrate the mean expression ± SD. p-values were calculated using a Student’s t-test. A logarithmic transformation of LRP2, CYP24A1, CYP27B1 and VDR expression values was performed to achieve a Gaussian distribution of the data. Note that CYP27B1 expression was under the detection limit in 46 placental samples of which 28 were from the 90 µg vitamin D group and 18 from the 10 µg vitamin D group. CUBN, LRP2 and VDR expression was standardized to the geomean of CycA and HPRT, and CYP24A1 and CYP27B1 expression was standardized to the expression of HPRT.
3.1.1. Correlation to the Third-Trimester Maternal Serum 25(OH)D
In a subset of the participants, it was possible to analyze gene expression in relation to maternal 25(OH)D concentration in the third trimester (n = 34). A linear regression model revealed that maternal third-trimester 25(OH)D concentrations were positively correlated with the expression of LRP2 (p = 0.036), but not CUBN (p = 0.44), CYP24A1 (p = 0.42), CYP27B1 (p = 0.82) or VDR (p = 0.074) (Figure 2a–e).
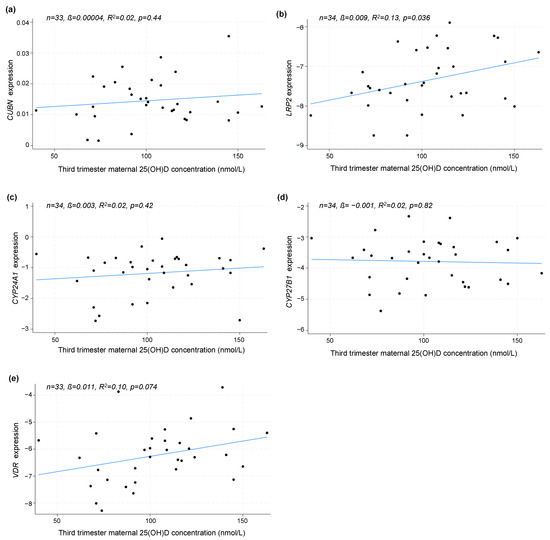
Figure 2.
The linear correlation analysis between the placental gene expression of (a) CUBN, (b) LRP2, (c) CYP24A1, (d) CYP27B1 and (e) VDR and maternal 25(OH)D concentration in the third trimester. The correlation is solely based on the measured 25(OH)D in the third trimester, not distinguishing between dosing groups. A logarithmic transformation of LRP2, CYP24A1, CYP27B1 and VDR expression values was performed to achieve a Gaussian distribution of the data. CUBN, LRP2 and VDR expression was standardized to the geomean of CycA and HPRT, and CYP24A1 and CYP27B1 expression was standardized to the expression of HPRT. 25(OH)D, 25-dihydroxy vitamin D.
3.1.2. The Effect of Pre-Pregnancy BMI
As depicted in Figure 3, no difference was found in the gene expression of any of the vitD-related genes among women with a pre-pregnancy BMI < 30 and >30 kg/m2. To further nuance the possible effect of BMI, we used a spline model to visualize any tendency toward an effect of pre-pregnancy BMI. This approach showed a tendency of an increased CUBN expression with increasing BMI in the 90 µg vitD group (Figure 4a). No differences with regard to LRP2 expression were observed (Figure 4b). For CYP24A1, the spline model approach indicated a higher CYP24A1 expression among overweight or obese women in the 90 µg vitD group with no apparent response to increasing BMI in the 10 µg vitD group (Figure 4c). With regard to CYP27B1, the 90 µg vitD group appeared to have a lower expression of CYP27B1 compared to the 10 µg vitD group in normal and overweight pregnancies, but at a BMI of 29 kg/m2, this correlation was reversed, and CYP27B1 expression was higher in the 90 µg vitD group (Figure 4d). For VDR, the spline model did not point toward an association between pre-pregnancy BMI and placental VDR expression (Figure 4e).
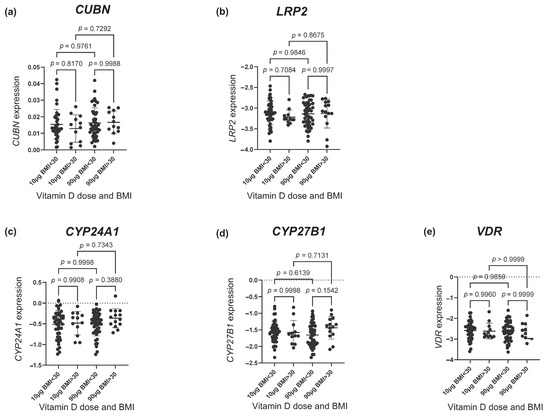
Figure 3.
The gene expression of (a) CUBN, (b) LRP2, (c) CYP24A1, (d) CYP27B1 and (e) VDR according to the vitamin D dosing group (10 µg vs. 90 µg) and BMI group (> and <30 kg/m2). The error bars in the figure illustrate the mean expression ± SD. p-values were calculated using a one-way ANOVA. A logarithmic transformation of LRP2, CYP24A1, CYP27B1 and VDR expression values was performed to achieve a Gaussian distribution of the data. Note that CYP27B1 expression was under the detection limit in 46 placental samples of which 28 were from the 90 µg vitamin D group and 18 from the 10 µg vitamin D group. CUBN, LRP2 and VDR expression was standardized to the geomean of CycA and HPRT, and CYP24A1 and CYP27B1 expression was standardized to the expression of HPRT. BMI, body mass index.
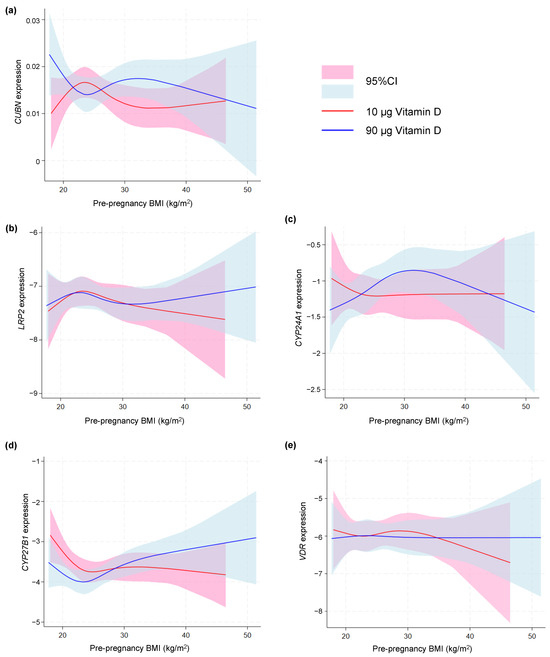
Figure 4.
Spline models depicting the gene expression of (a) CUBN, (b) LRP2, (c) CYP24A1, (d) CYP27B1 and (e) VDR correlated to maternal pre-pregnancy BMI in the two vitamin D dosing groups (10 µg vs. 90 µg). A logarithmic transformation of LRP2, CYP24A1, CYP27B1 and VDR expression values was performed to achieve a Gaussian distribution of the data. Note that CYP27B1 expression was under the detection limit in 46 placental samples of which 28 were from the 90 µg vitamin D group and 18 from the 10 µg vitamin D group. CUBN, LRP2 and VDR expression was standardized to the geomean of CycA and HPRT, and CYP24A1 and CYP27B1 expression was standardized to the expression of HPRT. BMI, body mass index; 95% CI, 95% confidence interval.
3.1.3. Impact of Early vitD Status
As the randomization was conducted at the end of the first trimester, weeks after the placentation, it is relevant to test if the maternal 25(OH)D concentration at study entry had any association to the placental gene expression at delivery. Using a linear correlation model, we found no association between first-trimester 25(OH)D concentration and the gene expression of CUBN (p = 0.68), LRP2 (p = 0.30), CYP24A1 (p = 0.39) or CYP27B1 (p = 0.87). However, for the gene expression of VDR, a statistically significant positive correlation (p = 0.02) was found (Figure 5).
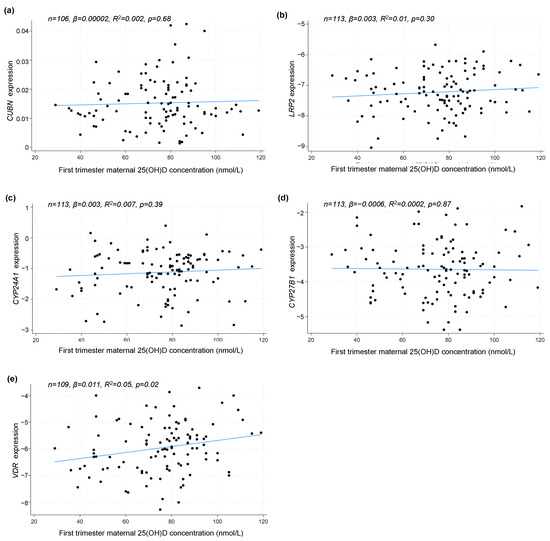
Figure 5.
Linear correlation analyses between placental (a) CUBN, (b) LRP2, (c) CYP24A1, (d) CYP27B1 and (e) VDR expression and maternal 25(OH)D concentration in the first trimester. A logarithmic transformation of LRP2, CYP24A1, CYP27B1 and VDR expression values was performed to achieve a Gaussian distribution of the data. CUBN, LRP2 and VDR expression was standardized to the geomean of CycA and HPRT, and CYP24A1 and CYP27B1 expression was standardized to the expression of HPRT. 25(OH)D, 25-hydroxy vitamin D.
3.1.4. Delivery Mode and Offspring Sex
As we have previously found that fetal sex is reflected in the placental transcriptome [41], we tested if the vitD-associated genes were differently expressed in placentas from pregnancies with a male offspring compared to their female counterparts, finding no statistically significant impact of offspring sex for any of the analyzed genes (CUBN (p = 0.39), LRP2 (p = 0.85), CYP24A1 (p = 0.92), CYP27B1 (p = 0.93) or VDR (p = 0.13)). We also tested if mode of delivery (vaginal vs. sectio) altered the gene expression but found no association for any of the analyzed genes (CUBN (p = 0.81), LRP2 (p = 0.35), CYP24A1 (p = 0.45), CYP27B1 (p = 0.75) or VDR (p = 0.21)).
3.2. Maternal VDBP during Pregnancy
Overall, offspring sex and maternal demographic data were similar in the two dosing groups among the subset of participants (n = 101) in whom VDBP was measured in maternal serum, albeit the 90 µg vitD group did include more women of non-Scandinavian origin, although the vast majority of women were Scandinavian in both groups (Supplementary Table S3). VDBP results were available from 99 participants.
As VDBP was determined in GW 12 + 0–13 + 4 and GW 34 + 0–35 + 6, respectively, we initially correlated the absolute values to maternal 25(OH)D concentration at these time points, finding no linear correlation between maternal 25(OH)D and VDBP in neither the first (p = 0.14) nor the third trimester (p = 0.64) (Figure 6). Moreover, the VDBP concentration in neither the first nor third trimester was associated with offspring sex (p = 0.65 and p = 0.72, respectively).
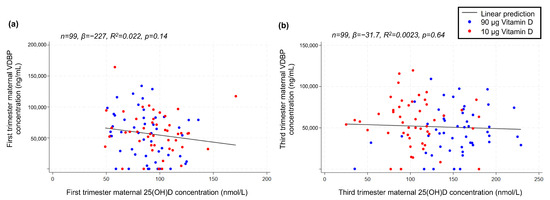
Figure 6.
Linear correlations between (a) maternal-serum VDBP and 25(OH)D concentration in the first trimester and (b) maternal-serum VDBP and 25(OH)D concentration in the third trimester. VDBP, vitamin D-binding protein; 25(OH)D, 25-hydroxy vitamin D.
The mean VDBP concentration measured in the third trimester was lower in the 90 µg group, i.e., 46,496 ng/mL, 95% CI [38,716, 54,275], compared to 56,385 ng/mL, 95% CI [48,230, 64,541] in the 10 µg group; however, this difference was not statistically significant (p = 0.08).
When calculating the absolute and relative VDBP change (first vs. third trimester), we found no statistically significant difference between the two dosing groups (p = 0.41 and p = 0.94, respectively). However, when we investigated the possible effect of pre-pregnancy BMI using spline models for the absolute (Figure 7a) and relative VDBP change (Figure 7b), we observed that the relative change in VDBP appeared to be affected by increasing BMI in the 90 µg group, whereas BMI did not elicit a marked VDBP change in the 10 µg dosing group.
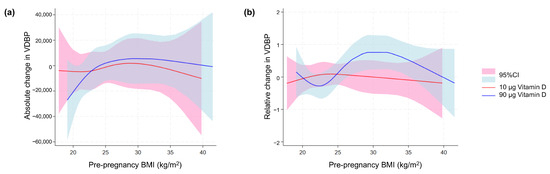
Figure 7.
VDBP in maternal blood in relation to maternal body weight. Spline models of the association between maternal pre-pregnancy BMI and (a) the absolute change and (b) relative change in maternal-serum VDBP concentration from the first to third trimester in relation to and stratified for the vitamin D dosing group (10 µg vs. 90 µg). BMI, body mass index; VDBP, vitamin D-binding protein; 95% CI, 95% confidence interval.
3.3. Neonatal vitD Status
As the fetal vitD supply is based on the transplacental vitD transfer, we finally determined the vitD status in the umbilical cord blood from 472 newborns. The offspring sex and maternal demographics of this subset of the GRAVITD cohort, which did not distinguish between healthy and complicated pregnancies, did not find any significant differences in the demographic and clinical factors between the two dosing groups (Supplementary Table S4).
It was evident that newborns from the 90 µg dosing group had a markedly higher mean 25(OH)D concentration compared to the 10 µg group, i.e., 83.5 nmol/L, 95% CI [80.0, 87.1] vs. 50.9 nmol/L, 95% CI [48.4, 53.4], (p < 0.0001).
Correspondingly, the prevalence of vitD deficiency was markedly reduced among newborns from the high-dose vitD group, i.e., 11 % (n = 26) in contrast to a prevalence of 51% (n = 124) in the 10 µg group. A total of 6% (n = 14) of newborns in the 10 µg vitD group had severe vitD deficiency <25 nmol/L, while this was only the case for 3 % (n = 7) of newborns in the 90 µg group.
Looking at the offspring from women with a pre-pregnancy BMI > 30 kg/m2 specifically, we found an increased frequency of vitD deficiency in the offspring from these pregnancies in both dosing groups, i.e., 17% in the 90 µg group and 61% in the 10 µg group.
4. Discussion
The findings from this study support that increased maternal vitD supplementation significantly reduces vitD deficiency among newborns and has a beneficial effect on neonatal vitD status, without signs of a reduced placental vitD uptake or an increased placental vitD degradation. The effects of high-dosage vitD supplementation on placental function have not previously been scrutinized. In this study, we analyzed a large number of placentas from pregnant women supplemented with either 10 µg or 90 µg of vitD3 aiming at reducing this knowledge gap.
Although our analysis based on an intention to treat found no significant differences between the dosing groups, there were indications that high maternal 25(OH)D concentrations may lead to an increased transplacental transport capacity of vitD. Though limited by the fact that serum collected later in the pregnancy, e.g., in the third trimester was only available from around one third of the participants, we thus found that an increased maternal third-trimester 25(OH)D concentration was associated with increased placental LRP2 expression. This is supported by a previous small-scale study by Park et al. who further reported a positive correlation with placental CUBN [19]. Previous studies are, however, not conclusive on this point as others did not find any correlation between placental CUBN and LRP2 expression and maternal 25(OH)D in the third trimester [12]. In the present study, we investigated the effect of two markedly different vitD supplementation doses on placental gene expression which was not conducted in the previous studies [12,19]. Our findings thus resemble the results one would see if clinical recommendations were altered. However, there are some limitations such as the absence of pill counts, and the set-up did not make it possible to register individual variations in sun exposure. The RCT set-up considers seasonal variation during randomization, but it is possible that the increased vitD uptake would be more profound if data from the winter season were analyzed separately. Also, our analysis might reflect that it was carried out in a cohort with a limited frequency of vitD deficiency, especially very limited numbers of severe vitD deficiency, just as complicated pregnancies were not included in the placental study.
As previous research has shown that 25(OH)D bound to VDBP rather than free 25(OH)D is preferentially taken up by the placenta through the megalin/cubilin complex [12,19,42], it could be hypothesized that alterations in VDBP concentration may also influence placental vitD uptake. However, we did not observe a significant increase in VDBP associated with a difference in vitD supplementation. We cannot exclude that the sample size in which VDBP was tested and the fact that we are employing an intention-to-treat analysis may affect the results. On the other hand, a natural rise in VDBP of 40–50% in pregnancy has been described with a peak in the early third trimester [15,39], indicating that in the pregnant state, vitD metabolism is already maximized to adequately supply the fetus.
Interestingly, the spline model did hint toward an effect on the change in VDBP over time related to pre-pregnancy BMI. This could indicate that increasing the vitD supplement of women with high BMI to 90µg triggers an increase in VDBP that allows them to make more use of the vitD given. Similarly, our spline model also indicated an increase in CUBN in this group of women. On the other hand, the 10 µg of vitD supplement did not appear to induce a change in VDBP or CUBN, even though these women might need more vitD.
In a previous study, we found a positive association between the gene expression of CYP24A1 in term placentas and pre-pregnancy BMI [30]. Though not statistically significant in this study, our spline model did indicate an increased expression of CYP24A1 in obese and overweight pregnancies. This could support a higher vitD need in obese pregnancies, as the frequency of vitD-deficient newborns was higher in these pregnancies. However, further studies are warranted to examine the maternal, placental and fetal responses to high-dose vitD supplementation in pregnancy, further comparing obese, overweight and lean women to determine if the advice on vitD supplementation should consider body composition.
With regard to our study population as a whole, this study shows that the currently recommended vitD supplementation of 10 µg/day does not prevent vitD deficiency in as many as 51% of all newborns, whereas increasing the supplementation to 90 µg results in vitD sufficiency in the majority of newborns. Moreover, the prevalence of vitD deficiency might even be underestimated in the present study, as 25(OH)D concentrations determined in plasma, as was the case with the umbilical cord samples, tend to be higher than those measured in serum [43].
From a health perspective, our findings are of high clinical relevance as a sufficient fetal and neonatal vitD status is important for bone health [44,45]. Furthermore, intrauterine vitD exposure may potentially reduce later-life disease risk by strengthening the immune system [46], thereby potentially reducing the risk of immune-system-related conditions like type 1 diabetes, asthma and allergy [6,9,47]. VitD insufficiency during pregnancy and the first vulnerable years of life also markedly increases the risk of common and potentially fatal infections such as respiratory syncytial virus (RSV) [48]. A previous study found a six-fold increase in RSV frequency in children who were vitD-deficient at birth [49], and a reduction in RSV susceptibility would reduce hospitalization rates and mortality in young children.
Placental VDR expression per se has previously been linked to fetal growth and development [12]. Though we found no direct association between supplementation or actual maternal vitD status in late pregnancy and VDR, our previous next-generation RNA sequencing study on vitD supplementation and the placental transcriptome found that alterations in placental function occur with the high-dose regime tested here [41]. Such changes, e.g., those linked to neurodevelopment, are likely to further benefit the growing fetus, in addition to the direct increase in fetal vitD supply. Notably, we found that VDR expression appeared unaffected by high-dose supplementation through the last six months of pregnancy, albeit significantly associated with first-trimester vitD measurements, highlighting the importance of also paying specific attention to pre-pregnancy vitD status for optimal placental function.
As VDR activation depends on the presence of the active vitD metabolite 1,25(OH)2D, we further investigated the effects of vitD supplementation on the gene encoding the vitD-activating enzyme CYP27B1. Notably, we found that the placental gene expression of CYP27B1 was below the detection limit in 39% of the samples. This likely relates to the low content of the enzyme in villus tissue compared to other parts of the placenta [42]. We did not see significant effects of either dosing group or maternal vitD concentration. However, we did observe indications of differences in CYP27B1 expression in relation to pre-pregnancy BMI. This could further suggest a complex vitD metabolism and uptake in women with obesity and overweight that may be associated with the numerous clinical problems seen in such pregnancies; however, more research is needed before such conclusions can be made for certain.
5. Conclusions
A high dose of vitD supplementation during pregnancy significantly reduces vitD deficiency among newborns and thus improves the vitD status without remarkably reducing the placental vitD uptake or affecting placental vitD metabolism. There is, however, a need for further studies on vitD metabolism and maternal–fetal vitD transfer in women with overweight or obesity to elucidate if maternal body weight affects the nutritional needs in pregnancy.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/nu16132145/s1, Table S1: Clinical characteristics of pregnancies for the 118 placentas studied, Table S2: Demographic and clinical data for the 118 women with a placenta eligible for gene analysis, Table S3: Demographic data for the 101 participants with available first- and third-trimester samples for VDBP analysis, Table S4: Demographic data for the 472 participants with cord blood available for 25(OH)D.
Author Contributions
Conceptualization, A.L.V., A.L. and P.B.; methodology, A.L.V., M.K.A., A.L. and P.B.; validation, A.L.V., M.K.A., H.H.A. and K.A.B.; formal analysis, A.L.V., M.K.A., H.H.A. and K.A.B.; investigation, A.L.V., M.K.A., H.H.A. and K.A.B.; resources, A.L.V., M.K.A., A.L. and P.B.; data curation, A.L.V., M.K.A., H.H.A. and K.A.B.; writing—original draft preparation, A.L.V.; writing—review and editing, A.L.V., M.K.A., A.L. and P.B.; visualization, A.L.V. and H.H.A.; supervision, A.L. and P.B.; project administration, A.L.V. and M.K.A.; funding acquisition, A.L.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Aarhus University; Aase and Ejner Danielsens Foundation; Axel Muusfeldt Foundation; the Augustinus Foundation; the Danish Medical Association Research Foundation; the Toyota Foundation; Director Emil C. Hertz and Wife Inger Hertz’ Foundation; Dagmar Marshall Foundation and the Frimodt-Heineke Foundation. All vitamin and placebo tablets used in the trial were donated by Orkla Care, Denmark.
Institutional Review Board Statement
This study was conducted in accordance with the Declaration of Helsinki and approved by the scientific ethical committee of the Central Denmark Region on 9 September 2019 (1–10-72–54-19).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study after receiving both oral and written information about the study.
Data Availability Statement
The full dataset is not publicly available as we do not have consent from the participants to publish the full dataset. However, a de-identified dataset will be available from the corresponding author upon reasonable request.
Acknowledgments
A great thank you to the midwifes and other personnel at the delivery ward at Randers Regional Hospital for helping with the placental sample collection. Substantial gratitude to the participating women who donated blood samples and placental tissue for the study. And lastly, a thank you to lab technician Henriette Gram Johanson for the preparation and handling of samples and assistance in the analysis.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript or in the decision to publish the results.
References
- Raia-Barjat, T.; Sarkis, C.; Rancon, F.; Thibaudin, L.; Gris, J.C.; Alfaidy, N.; Chauleur, C. Vitamin D deficiency during late pregnancy mediates placenta-associated complications. Sci. Rep. 2021, 11, 20708. [Google Scholar] [CrossRef]
- O’Callaghan, K.M.; Kiely, M. Systematic Review of Vitamin D and Hypertensive Disorders of Pregnancy. Nutrients 2018, 10, 294. [Google Scholar] [CrossRef]
- Wei, S.-Q.; Qi, H.-P.; Luo, Z.-C.; Fraser, W.D. Maternal vitamin D status and adverse pregnancy outcomes: A systematic review and meta-analysis. J. Matern.-Fetal Neonatal Med. 2013, 26, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Ness, R.B.; Markovic, N.; Roberts, J.M. The risk of preeclampsia rises with increasing prepregnancy body mass index. Ann. Epidemiol. 2005, 15, 475–482. [Google Scholar] [CrossRef]
- Kiely, M.E.; Zhang, J.Y.; Kinsella, M.; Khashan, A.S.; Kenny, L.C. Vitamin D status is associated with uteroplacental dysfunction indicated by pre-eclampsia and small-for-gestational-age birth in a large prospective pregnancy cohort in Ireland with low vitamin D status. Am. J. Clin. Nutr. 2016, 104, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Ponsonby, A.L.; Lucas, R.M.; Lewis, S.; Halliday, J. Vitamin D status during pregnancy and aspects of offspring health. Nutrients 2010, 2, 389–407. [Google Scholar] [CrossRef]
- Wagner, C.L.; Hollis, B.W.; Kotsa, K.; Fakhoury, H.; Karras, S.N. Vitamin D administration during pregnancy as prevention for pregnancy, neonatal and postnatal complications. Rev. Endocr. Metab. Disord. 2017, 18, 307–322. [Google Scholar] [CrossRef]
- Ideraabdullah, F.Y.; Belenchia, A.M.; Rosenfeld, C.S.; Kullman, S.W.; Knuth, M.; Mahapatra, D.; Bereman, M.; Levin, E.D.; Peterson, C.A. Maternal vitamin D deficiency and developmental origins of health and disease (DOHaD). J. Endocrinol. 2019, 241, R65–R80. [Google Scholar] [CrossRef] [PubMed]
- Thorsteinsdottir, F.; Cardoso, I.; Keller, A.; Stougaard, M.; Frederiksen, P.; Cohen, A.S.; Maslova, E.; Jacobsen, R.; Backer, V.; Heitmann, B.L. Neonatal Vitamin D Status and Risk of Asthma in Childhood: Results from the D-Tect Study. Nutrients 2020, 12, 842. [Google Scholar] [CrossRef]
- Bikle, D. Nonclassic actions of vitamin D. J. Clin. Endocrinol. Metab. 2009, 94, 26–34. [Google Scholar] [CrossRef]
- Norman, A.W. From vitamin D to hormone D: Fundamentals of the vitamin D endocrine system essential for good health. Am. J. Clin. Nutr. 2008, 88, 491s–499s. [Google Scholar] [CrossRef] [PubMed]
- Ashley, B.; Simner, C.; Manousopoulou, A.; Jenkinson, C.; Hey, F.; Frost, J.M.; Rezwan, F.I.; White, C.H.; Lofthouse, E.M.; Hyde, E.; et al. Placental uptake and metabolism of 25(OH)vitamin D determine its activity within the fetoplacental unit. Elife 2022, 11, e71094. [Google Scholar] [CrossRef] [PubMed]
- Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [CrossRef] [PubMed]
- Akour, A.A.; Gerk, P.; Kennedy, M.J. Megalin expression in human term and preterm placental villous tissues: Effect of gestational age and sample processing and storage time. J. Pharmacol. Toxicol. Methods 2015, 71, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Fernando, M.; Ellery, S.J.; Marquina, C.; Lim, S.; Naderpoor, N.; Mousa, A. Vitamin D-Binding Protein in Pregnancy and Reproductive Health. Nutrients 2020, 12, 1489. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.Q.; Hewison, M. Vitamin D, the placenta and pregnancy. Arch. Biochem. Biophys. 2012, 523, 37–47. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, K.O.; Li, S.; Cao, C.; Kent, T.; Young, B.V.; Queenan, R.A.; Pressman, E.K.; Cooper, E.M. Placental CYP27B1 and CYP24A1 expression in human placental tissue and their association with maternal and neonatal calcitropic hormones. J. Clin. Endocrinol. Metab. 2014, 99, 1348–1356. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ma, R.; Gu, Y.; Zhao, S.; Sun, J.; Groome, L.J.; Wang, Y. Expressions of vitamin D metabolic components VDBP, CYP2R1, CYP27B1, CYP24A1, and VDR in placentas from normal and preeclamptic pregnancies. Am. J. Physiol. 2012, 303, E928–E935. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Wood, M.R.; Malysheva, O.V.; Jones, S.; Mehta, S.; Brannon, P.M.; Caudill, M.A. Placental vitamin D metabolism and its associations with circulating vitamin D metabolites in pregnant women. Am. J. Clin. Nutr. 2017, 106, 1439–1448. [Google Scholar] [CrossRef]
- Schulz, E.V.; Cruze, L.; Wei, W.; Gehris, J.; Wagner, C.L. Maternal vitamin D sufficiency and reduced placental gene expression in angiogenic biomarkers related to comorbidities of pregnancy. J. Steroid Biochem. Mol. Biol. 2017, 173, 273–279. [Google Scholar] [CrossRef]
- Mousa, A.; Abell, S.; Scragg, R.; de Courten, B. Vitamin D in Reproductive Health and Pregnancy. Semin. Reprod. Med. 2016, 34, e1–e13. [Google Scholar] [CrossRef] [PubMed]
- Authority, T.D.H. Anbefalinger om Tilskud Med D-Vitamin. Available online: https://sst.dk/da/viden/forebyggelse/ernaering/anbefalinger-om-d-vitamin (accessed on 14 June 2023).
- Gallo, S.; McDermid, J.M.; Al-Nimr, R.I.; Hakeem, R.; Moreschi, J.M.; Pari-Keener, M.; Stahnke, B.; Papoutsakis, C.; Handu, D.; Cheng, F.W. Vitamin D Supplementation during Pregnancy: An Evidence Analysis Center Systematic Review and Meta-Analysis. J. Acad. Nutr. Diet. 2020, 120, 898–924.e4. [Google Scholar] [CrossRef] [PubMed]
- Saraf, R.; Morton, S.M.; Camargo, C.A., Jr.; Grant, C.C. Global summary of maternal and newborn vitamin D status—A systematic review. Matern. Child Nutr. 2016, 12, 647–668. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.M. Collateral Damage: Maternal Obesity During Pregnancy Continues to Rise. Obstet. Gynecol. Surv. 2020, 75, 39–49. [Google Scholar] [CrossRef]
- Devlieger, R.; Benhalima, K.; Damm, P.; Van Assche, A.; Mathieu, C.; Mahmood, T.; Dunne, F.; Bogaerts, A. Maternal obesity in Europe: Where do we stand and how to move forward?: A scientific paper commissioned by the European Board and College of Obstetrics and Gynaecology (EBCOG). Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 201, 203–208. [Google Scholar] [CrossRef]
- Pereira-Santos, M. Obesity and vitamin D deficiency: A systematic review and meta-analysis. Obes. Rev. 2015, 16, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, L.M.; Catov, J.M.; Roberts, J.M.; Simhan, H.N. Prepregnancy obesity predicts poor vitamin D status in mothers and their neonates. J. Nutr. 2007, 137, 2437–2442. [Google Scholar] [CrossRef]
- Wortsman, J.; Matsuoka, L.Y.; Chen, T.C.; Lu, Z.; Holick, M.F. Decreased bioavailability of vitamin D in obesity. Am. J. Clin. Nutr. 2000, 72, 690–693. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, A.L.; Justesen, S.; Volqvartz, T.; Aagaard, S.K.; Andreasen, M.F.; Lesnikova, I.; Uldbjerg, N.; Larsen, A.; Bor, P. Vitamin D insufficiency among Danish pregnant women-Prevalence and association with adverse obstetric outcomes and placental vitamin D metabolism. Acta Obstet. Gynecol. Scand. 2021, 100, 480–488. [Google Scholar] [CrossRef]
- Josefson, J.L.; Feinglass, J.; Rademaker, A.W.; Metzger, B.E.; Zeiss, D.M.; Price, H.E.; Langman, C.B. Maternal obesity and vitamin D sufficiency are associated with cord blood vitamin D insufficiency. J. Clin. Endocrinol. Metab. 2013, 98, 114–119. [Google Scholar] [CrossRef]
- Paredes, C.; Hsu, R.C.; Tong, A.; Johnson, J.R. Obesity and Pregnancy. Neoreviews 2021, 22, e78–e87. [Google Scholar] [CrossRef] [PubMed]
- Hollis, B.W.; Johnson, D.; Hulsey, T.C.; Ebeling, M.; Wagner, C.L. Vitamin D supplementation during pregnancy: Double-blind, randomized clinical trial of safety and effectiveness. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2011, 26, 2341–2357. [Google Scholar] [CrossRef]
- Roth, D.E.; Morris, S.K.; Zlotkin, S.; Gernand, A.D.; Ahmed, T.; Shanta, S.S.; Papp, E.; Korsiak, J.; Shi, J.; Islam, M.M.; et al. Vitamin D Supplementation in Pregnancy and Lactation and Infant Growth. N. Engl. J. Med. 2018, 379, 535–546. [Google Scholar] [CrossRef]
- Wagner, C.L.; McNeil, R.; Hamilton, S.A.; Winkler, J.; Rodriguez Cook, C.; Warner, G.; Bivens, B.; Davis, D.J.; Smith, P.G.; Murphy, M.; et al. A randomized trial of vitamin D supplementation in 2 community health center networks in South Carolina. Am. J. Obstet. Gynecol. 2013, 208, 137.e1–137.e13. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, A.L.; Christensen, M.; Andreasen, M.F.; Larsen, A.; Bor, P. Vitamin D in pregnancy (GRAVITD)—A randomised controlled trial identifying associations and mechanisms linking maternal Vitamin D deficiency to placental dysfunction and adverse pregnancy outcomes—Study protocol. BMC Pregnancy Childbirth 2023, 23, 177. [Google Scholar] [CrossRef]
- Nelson, M.; Burton, G.J. A technical note to improve the reporting of studies of the human placenta. Placenta 2010, 32, 195–196. [Google Scholar] [CrossRef] [PubMed]
- Garg, U. 25-Hydroxyvitamin D Testing: Immunoassays Versus Tandem Mass Spectrometry. Clin. Lab. Med. 2018, 38, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Lucey, A.J.; Horgan, R.; Kenny, L.C.; Kiely, M. Impact of pregnancy on vitamin D status: A longitudinal study. Br. J. Nutr. 2014, 112, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Steenland, K.; Deddens, J.A. A practical guide to dose-response analyses and risk assessment in occupational epidemiology. Epidemiology 2004, 15, 63–70. [Google Scholar] [CrossRef]
- Vestergaard, A.L.; Andersen, M.K.; Olesen, R.V.; Bor, P.; Larsen, A. High-Dose Vitamin D Supplementation Significantly Affects the Placental Transcriptome. Nutrients 2023, 15, 5032. [Google Scholar] [CrossRef]
- Tamblyn, J.A.; Susarla, R.; Jenkinson, C.; Jeffery, L.E.; Ohizua, O.; Chun, R.F.; Chan, S.Y.; Kilby, M.D.; Hewison, M. Dysregulation of maternal and placental vitamin D metabolism in preeclampsia. Placenta 2017, 50, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Harvey, S.M.; Murphy, V.E.; Gibson, P.G.; Clarke, M.; Jensen, M.E. The Impact of Sample Type on Vitamin D Quantification and Clinical Classification during Pregnancy. Nutrients 2020, 12, 3872. [Google Scholar] [CrossRef] [PubMed]
- Javaid, M.K.; Crozier, S.R.; Harvey, N.C.; Gale, C.R.; Dennison, E.M.; Boucher, B.J.; Arden, N.K.; Godfrey, K.M.; Cooper, C. Maternal vitamin D status during pregnancy and childhood bone mass at age 9 years: A longitudinal study. Lancet 2006, 367, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Mahon, P.; Harvey, N.; Crozier, S.; Inskip, H.; Robinson, S.; Arden, N.; Swaminathan, R.; Cooper, C.; Godfrey, K. Low maternal vitamin D status and fetal bone development: Cohort study. J. Bone Miner. Res. 2010, 25, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Hornsby, E.; Pfeffer, P.E.; Laranjo, N.; Cruikshank, W.; Tuzova, M.; Litonjua, A.A.; Weiss, S.T.; Carey, V.J.; O’Connor, G.; Hawrylowicz, C. Vitamin D supplementation during pregnancy: Effect on the neonatal immune system in a randomized controlled trial. J. Allergy Clin. Immunol. 2018, 141, 269–278.e1. [Google Scholar] [CrossRef]
- Pacheco-González, R.M.; García-Marcos, L.; Morales, E. Prenatal vitamin D status and respiratory and allergic outcomes in childhood: A meta-analysis of observational studies. Pediatr. Allergy Immunol. 2018, 29, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Loddo, F.; Nauleau, S.; Lapalus, D.; Tardieu, S.; Bernard, O.; Boubred, F. Association of Maternal Gestational Vitamin D Supplementation with Respiratory Health of Young Children. Nutrients 2023, 15, 2380. [Google Scholar] [CrossRef]
- Belderbos, M.E.; Houben, M.L.; Wilbrink, B.; Lentjes, E.; Bloemen, E.M.; Kimpen, J.L.; Rovers, M.; Bont, L. Cord blood vitamin D deficiency is associated with respiratory syncytial virus bronchiolitis. Pediatrics 2011, 127, e1513–e1520. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).