
Medicinal Mushroom Extracts from Hericium coralloides and Trametes versicolor Exert Differential Immunomodulatory Effects on Immune Cells from Older Adults In Vitro
 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Population
2.2. Blood Processing and Peripheral Blood Mononuclear Cell Isolation
2.3. In Vitro Cell Culture of Peripheral Blood Mononuclear Cells
2.4. LEGENDplexTM Assay
2.5. Statistical Analysis
3. Results
3.1. Subject Characteristics
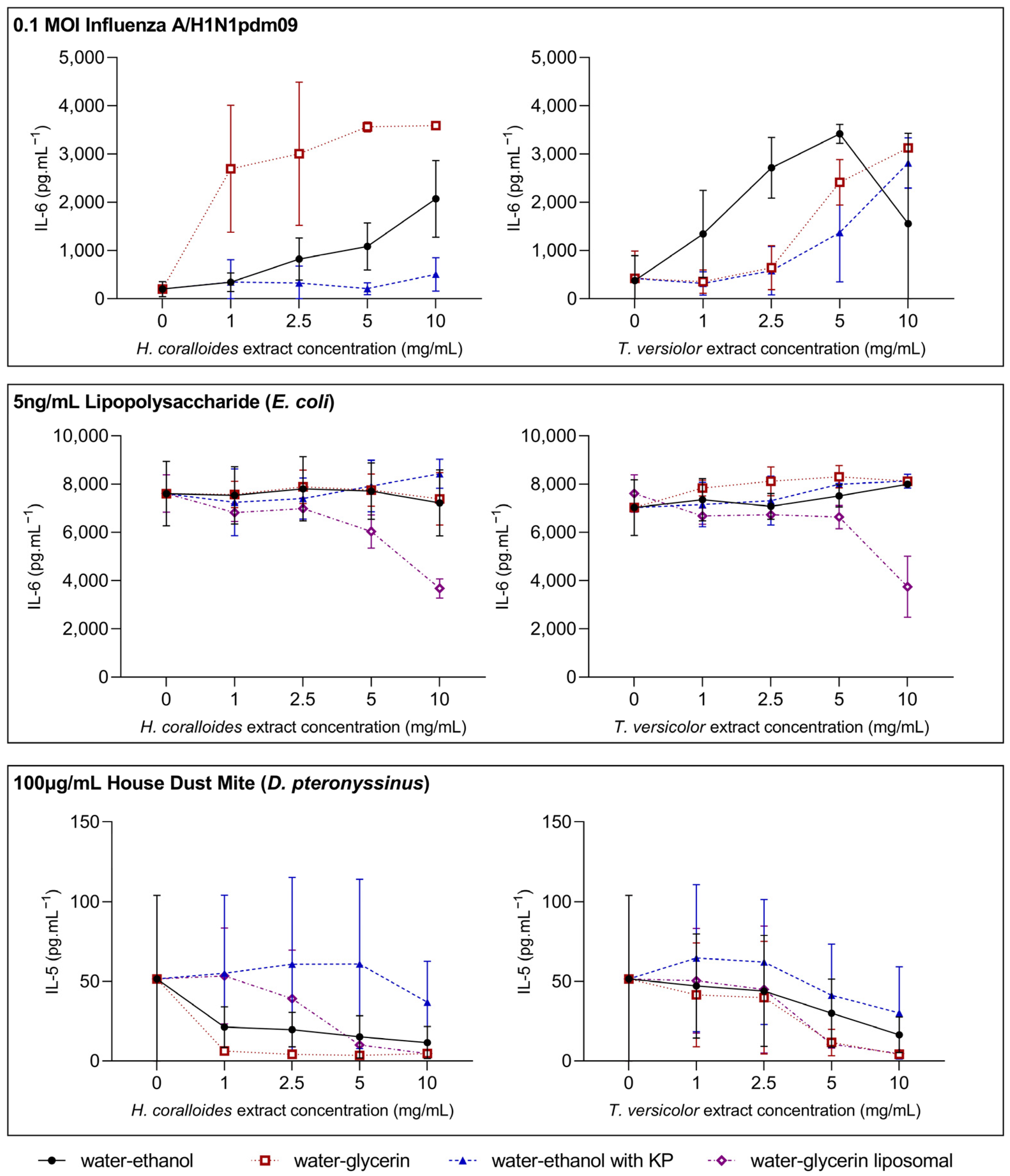
3.2. Dose–Response Relationships between Mushroom Extract Concentration and Inflammatory Mediator Release across Models of Stimulation
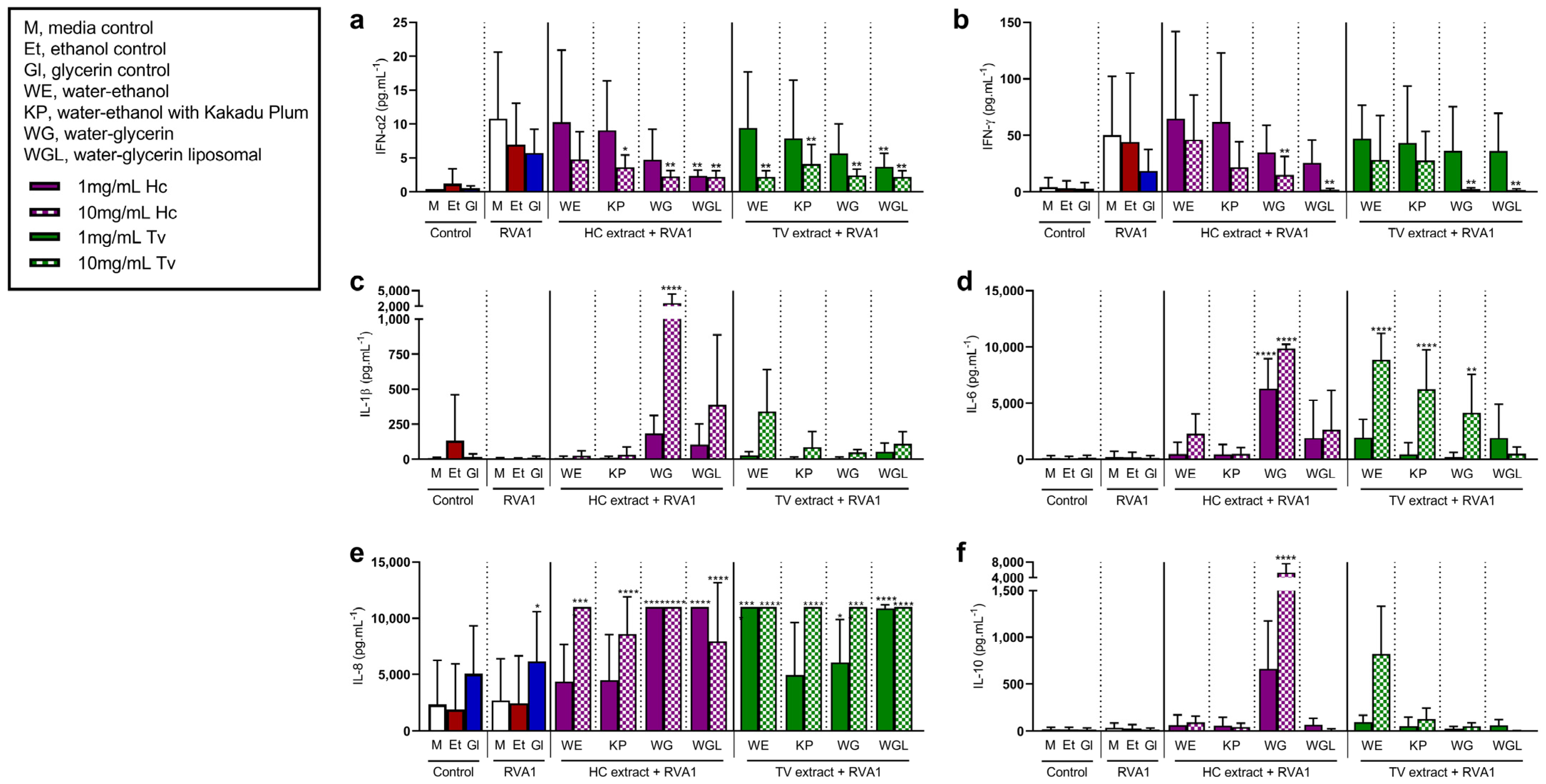
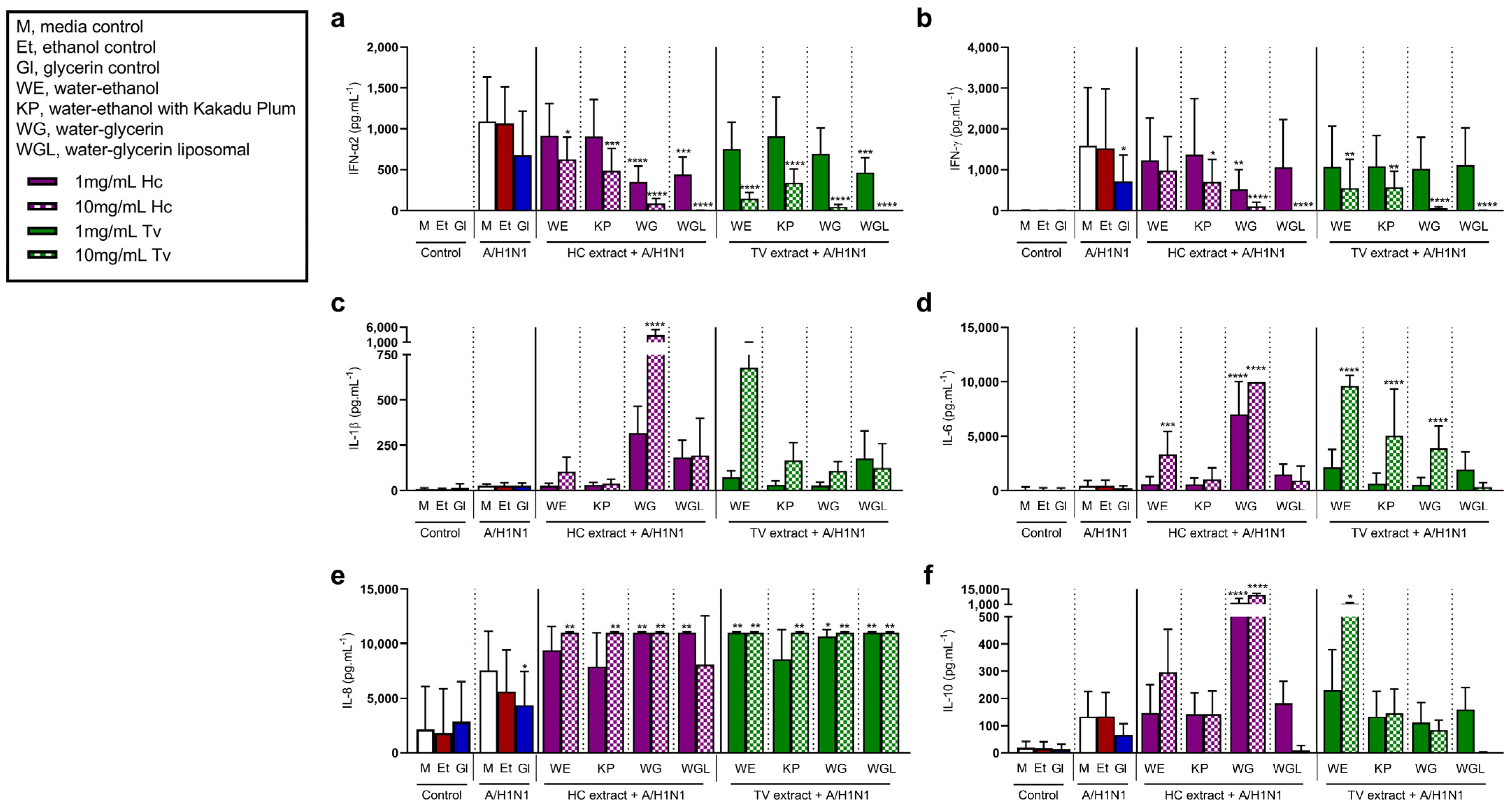
3.3. Mushroom Extracts Reduce Type I and II IFN, While Increasing Pro-Inflammatory Mediator Release to Rhinovirus and Influenza Stimulation
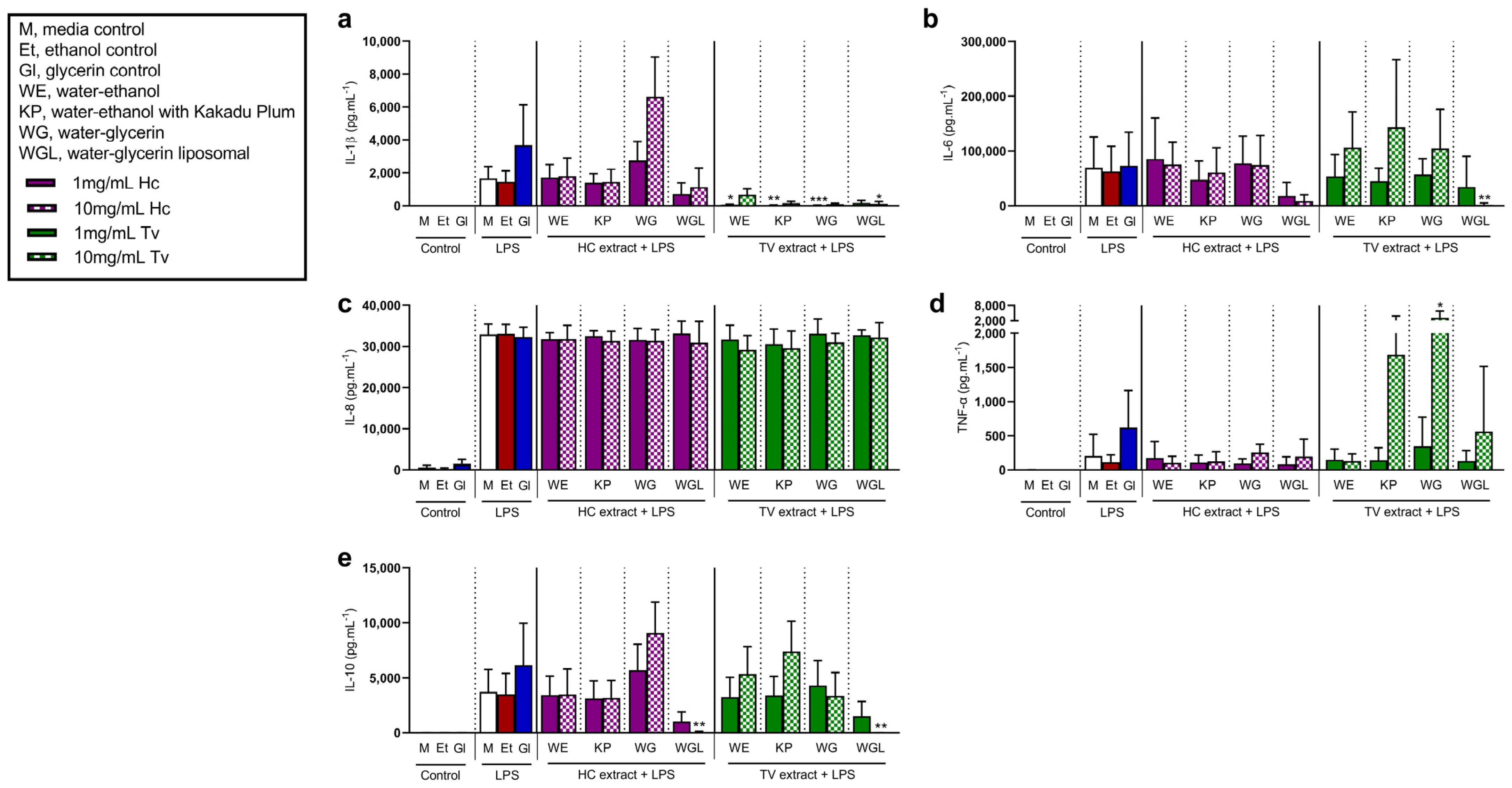
3.4. Interleukin-1β Is Decreased with Mushroom Extracts from Trametes versicolor in the Presence of Lipopolysaccharide
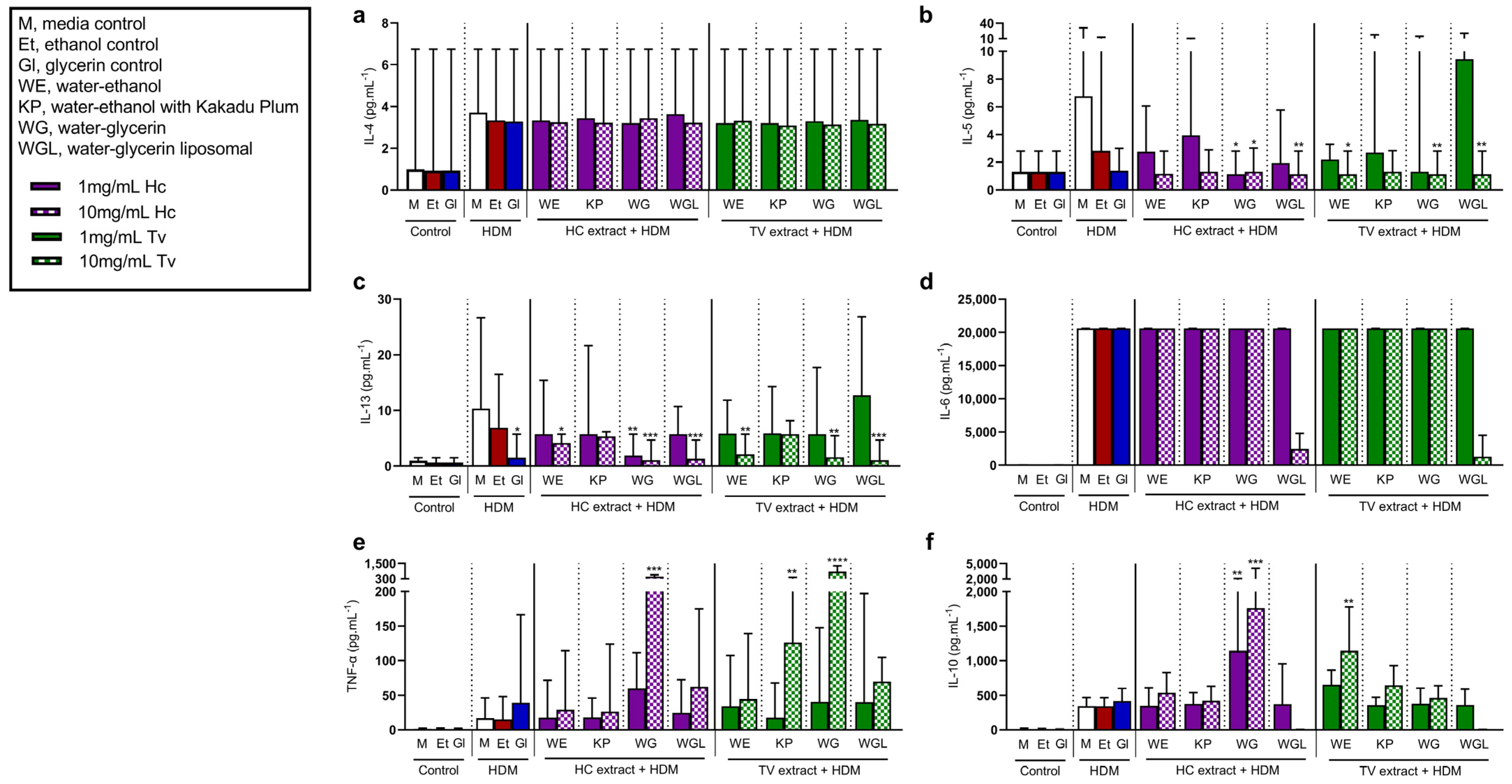
3.5. Type 2 Cytokines Induced by House Dust Mite Are Suppressed by Treatment with Mushroom Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soares, A.A.; De Sá-Nakanishi, A.B.; Bracht, A.; Da Costa, S.M.G.; Koehnlein, E.A.; De Souza, C.G.M.; Peralta, R.M. Hepatoprotective Effects of Mushrooms. Molecules 2013, 18, 7609–7630. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.K.; Dutta, S.D.; Ganguly, K.; Cho, S.-J.; Lim, K.-T. Mushroom-Derived Bioactive Molecules as Immunotherapeutic Agents: A Review. Molecules 2021, 26, 1359. [Google Scholar] [CrossRef] [PubMed]
- Ślusarczyk, J.; Adamska, E.; Czerwik-Marcinkowska, J. Fungi and Algae as Sources of Medicinal and Other Biologically Active Compounds: A Review. Nutrients 2021, 13, 3178. [Google Scholar] [CrossRef] [PubMed]
- Motta, F.; Gershwin, M.E.; Selmi, C. Mushrooms and immunity. J. Autoimmun. 2021, 117, 102576. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Zhao, L.; Shui, X.; Wang, L.-a.; Wu, Y. Antioxidant and Anti-Aging Activities of Ethyl Acetate Extract of the Coral Tooth Mushroom, Hericium coralloides (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 561–570. [Google Scholar] [CrossRef]
- Tang, P.; Li, X.; Wang, D.; Feng, C.; Han, A. Effect of Hericium coralloides polysaccharide on the antioxidant and regulation of hepatic metabolism in rats. Edible Fungi China 2014, 33, 48–51. [Google Scholar]
- Tabibzadeh, F.; Alvandi, H.; Hatamian-Zarmi, A.; Kalitukha, L.; Aghajani, H.; Ebrahimi-Hosseinzadeh, B. Antioxidant activity and cytotoxicity of exopolysaccharide from mushroom Hericium coralloides in submerged fermentation. Biomass Convers. Biorefin. 2022, 1–11. [Google Scholar] [CrossRef]
- Zou, Y.-J.; Wang, H.-X.; Ng, T.-B.; Huang, C.-Y.; Zhang, J.-X. Purification and characterization of a novel laccase from the edible mushroom Hericium coralloides. J. Microbiol. 2012, 50, 72–78. [Google Scholar] [CrossRef]
- Benson, K.F.; Stamets, P.; Davis, R.; Nally, R.; Taylor, A.; Slater, S.; Jensen, G.S. The mycelium of the Trametes versicolor (Turkey tail) mushroom and its fermented substrate each show potent and complementary immune activating properties in vitro. BMC Complement. Altern. Med. 2019, 19, 342. [Google Scholar] [CrossRef]
- Yang, S.F.; Zhuang, T.F.; Si, Y.M.; Qi, K.Y.; Zhao, J. Coriolus versicolor mushroom polysaccharides exert immunoregulatory effects on mouse B cells via membrane Ig and TLR-4 to activate the MAPK and NF-κB signaling pathways. Mol. Immunol. 2015, 64, 144–151. [Google Scholar] [CrossRef]
- Krsmanović, N.; Rašeta, M.; Mišković, J.; Bekvalac, K.; Bogavac, M.; Karaman, M.; Isikhuemhen, O.S. Effects of UV Stress in Promoting Antioxidant Activities in Fungal Species Trametes versicolor (L.) Lloyd and Flammulina velutipes (Curtis) Singer. Antioxidants 2023, 12, 302. [Google Scholar] [CrossRef]
- Torkelson, C.J.; Sweet, E.; Martzen, M.R.; Sasagawa, M.; Wenner, C.A.; Gay, J.; Putiri, A.; Standish, L.J. Phase 1 Clinical Trial of Trametes versicolor in Women with Breast Cancer. ISRN Oncol. 2012, 2012, 251632. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.C. Aging. Proc. Am. Thorac. Soc. 2005, 2, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Pillai, P.S. Innate immunity to influenza virus infection. Nat. Rev. Immunol. 2014, 14, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Mitländer, H.; Vuorinen, T.; Finotto, S. Mechanism of Rhinovirus Immunity and Asthma. Front. Immunol. 2021, 12, 731846. [Google Scholar] [CrossRef]
- Park, B.S.; Lee, J.O. Recognition of lipopolysaccharide pattern by TLR4 complexes. Exp. Mol. Med. 2013, 45, e66. [Google Scholar] [CrossRef]
- Jacquet, A. Characterization of Innate Immune Responses to House Dust Mite Allergens: Pitfalls and Limitations. Front. Allergy 2021, 2, 662378. [Google Scholar] [CrossRef]
- Tolosa, J.M.; Parsons, K.S.; Hansbro, P.M.; Smith, R.; Wark, P.A. The placental protein syncytin-1 impairs antiviral responses and exaggerates inflammatory responses to influenza. PLoS ONE 2015, 10, e0118629. [Google Scholar] [CrossRef]
- Williams, E.J.; Guilleminault, L.; Berthon, B.S.; Eslick, S.; Wright, T.; Karihaloo, C.; Gately, M.; Baines, K.J.; Wood, L.G. Sulforaphane reduces pro-inflammatory response to palmitic acid in monocytes and adipose tissue macrophages. J. Nutr. Biochem. 2022, 104, 108978. [Google Scholar] [CrossRef]
- Eslick, S.; Williams, E.J.; Berthon, B.S.; Wright, T.; Karihaloo, C.; Gately, M.; Wood, L.G. Weight Loss and Short-Chain Fatty Acids Reduce Systemic Inflammation in Monocytes and Adipose Tissue Macrophages from Obese Subjects. Nutrients 2022, 14, 765. [Google Scholar] [CrossRef]
- Hosseini, B.; Berthon, B.S.; Starkey, M.R.; Collison, A.; McLoughlin, R.F.; Williams, E.J.; Nichol, K.; Wark, P.A.; Jensen, M.E.; Da Silva Sena, C.R.; et al. Children with Asthma Have Impaired Innate Immunity and Increased Numbers of Type 2 Innate Lymphoid Cells Compared with Healthy Controls. Front. Immunol. 2021, 12, 664668. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, B.; Berthon, B.S.; Jensen, M.E.; McLoughlin, R.F.; Wark, P.A.B.; Nichol, K.; Williams, E.J.; Baines, K.J.; Collison, A.; Starkey, M.R.; et al. The Effects of Increasing Fruit and Vegetable Intake in Children with Asthma on the Modulation of Innate Immune Responses. Nutrients 2022, 14, 3087. [Google Scholar] [CrossRef] [PubMed]
- Rawle, F.C.; Mitchell, E.B.; Platts-Mills, T.A. T cell responses to the major allergen from the house dust mite Dermatophagoides pteronyssinus, Antigen P1: Comparison of patients with asthma, atopic dermatitis, and perennial rhinitis. J. Immunol. 1984, 133, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.B.; Braciale, V.L.; Braciale, T.J. Influenza virus-specific CD4+ T helper type 2 T lymphocytes do not promote recovery from experimental virus infection. J. Exp. Med. 1994, 180, 1273–1282. [Google Scholar] [CrossRef] [PubMed]
- Moran, T.M.; Park, H.; Fernandez-Sesma, A.; Schulman, J.L. Th2 responses to inactivated influenza virus can Be converted to Th1 responses and facilitate recovery from heterosubtypic virus infection. J. Infect. Dis. 1999, 180, 579–585. [Google Scholar] [CrossRef]
- Kankkunen, P.; Teirilä, L.; Rintahaka, J.; Alenius, H.; Wolff, H.; Matikainen, S. (1,3)-beta-glucans activate both dectin-1 and NLRP3 inflammasome in human macrophages. J. Immunol. 2010, 184, 6335–6342. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, L.; Zhu, X.; Wang, Y.; Liu, W.; Gong, W. Polysaccharide Agaricus blazei Murill stimulates myeloid derived suppressor cell differentiation from M2 to M1 type, which mediates inhibition of tumour immune-evasion via the Toll-like receptor 2 pathway. Immunology 2015, 146, 379–391. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A.; Chávez-Valencia, V. High IL-6/IFN-γ ratio could be associated with severe disease in COVID-19 patients. J. Med. Virol. 2020, 92, 1789–1790. [Google Scholar] [CrossRef]
- Bernardshaw, S.; Hetland, G.; Ellertsen, L.K.; Tryggestad, A.M.; Johnson, E. An extract of the medicinal mushroom Agaricus blazei Murill differentially stimulates production of pro-inflammatory cytokines in human monocytes and human vein endothelial cells in vitro. Inflammation 2005, 29, 147–153. [Google Scholar] [CrossRef]
- Sorimachi, K.; Akimoto, K.; Ikehara, Y.; Inafuku, K.; Okubo, A.; Yamazaki, S. Secretion of TNF-alpha, IL-8 and nitric oxide by macrophages activated with Agaricus blazei Murill fractions in vitro. Cell Struct. Funct. 2001, 26, 103–108. [Google Scholar] [CrossRef]
- Johnson, E.; Førland, D.T.; Saetre, L.; Bernardshaw, S.V.; Lyberg, T.; Hetland, G. Effect of an extract based on the medicinal mushroom Agaricus blazei murill on release of cytokines, chemokines and leukocyte growth factors in human blood ex vivo and in vivo. Scand. J. Immunol. 2009, 69, 242–250. [Google Scholar] [CrossRef]
- Diling, C.; Xin, Y.; Chaoqun, Z.; Jian, Y.; Xiaocui, T.; Jun, C.; Ou, S.; Yizhen, X. Extracts from Hericium erinaceus relieve inflammatory bowel disease by regulating immunity and gut microbiota. Oncotarget 2017, 8, 85838–85857. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.Y.; Kim, D.H.; Lee, E.K.; Chung, K.W.; Chung, S.; Lee, B.; Seo, A.Y.; Chung, J.H.; Jung, Y.S.; Im, E.; et al. Redefining Chronic Inflammation in Aging and Age-Related Diseases: Proposal of the Senoinflammation Concept. Aging Dis. 2019, 10, 367–382. [Google Scholar] [CrossRef]
- Chung, K.W.; Lee, E.K.; Kim, D.H.; An, H.J.; Kim, N.D.; Im, D.S.; Lee, J.; Yu, B.P.; Chung, H.Y. Age-related sensitivity to endotoxin-induced liver inflammation: Implication of inflammasome/IL-1β for steatohepatitis. Aging Cell 2015, 14, 524–533. [Google Scholar] [CrossRef]
- Therkelsen, S.P.; Hetland, G.; Lyberg, T.; Lygren, I.; Johnson, E. Cytokine Levels After Consumption of a Medicinal Agaricus blazei Murill-Based Mushroom Extract, AndoSan(™), in Patients with Crohn’s Disease and Ulcerative Colitis in a Randomized Single-Blinded Placebo-Controlled Study. Scand. J. Immunol. 2016, 84, 323–331. [Google Scholar] [CrossRef]
- Mahmood, F.; Hetland, G.; Nentwich, I.; Mirlashari, M.R.; Ghiasvand, R.; Nissen-Meyer, L.S.H. Agaricus blazei-Based Mushroom Extract Supplementation to Birch Allergic Blood Donors: A Randomized Clinical Trial. Nutrients 2019, 11, 2339. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.P.; Kim, Y.M.; Choi, H.I.; Choi, S.J.; Park, H.T.; Lee, W.H.; Gho, Y.S.; Jee, Y.K.; Jeon, S.G.; Kim, Y.K. An important role of tumor necrosis factor receptor-2 on natural killer T cells on the development of dsRNA-enhanced Th2 cell response to inhaled allergens. Allergy 2014, 69, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.P.; Lee, S.M.; Choi, H.I.; Kim, M.H.; Jeon, S.G.; Jang, M.H.; Jee, Y.K.; Yang, S.; Cho, Y.J.; Kim, Y.K. House Dust Mite-Derived Chitin Enhances Th2 Cell Response to Inhaled Allergens, Mainly via a TNF-α-Dependent Pathway. Allergy Asthma Immunol. Res. 2016, 8, 362–374. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | 15 |
|---|---|
| Age, years (range) | 68.7 ± 9.5 |
| Weight, kg | 82.2 ± 16.6 |
| BMI 1, kg/m2 | 28.87 ± 5.05 |
| Male, n (%) | 6 (40) |
| Female, n (%) | 9 (60) |
| Ex-smokers, n (%) | 7 (47) |
| Smoking history, pack years | 13.6 ± 7.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, L.M.; Berthon, B.S.; Stoodley, I.L.; Williams, E.J.; Wood, L.G. Medicinal Mushroom Extracts from Hericium coralloides and Trametes versicolor Exert Differential Immunomodulatory Effects on Immune Cells from Older Adults In Vitro. Nutrients 2023, 15, 2227. https://doi.org/10.3390/nu15092227
Williams LM, Berthon BS, Stoodley IL, Williams EJ, Wood LG. Medicinal Mushroom Extracts from Hericium coralloides and Trametes versicolor Exert Differential Immunomodulatory Effects on Immune Cells from Older Adults In Vitro. Nutrients. 2023; 15(9):2227. https://doi.org/10.3390/nu15092227
Chicago/Turabian StyleWilliams, Lily M., Bronwyn S. Berthon, Isobel L. Stoodley, Evan J. Williams, and Lisa G. Wood. 2023. "Medicinal Mushroom Extracts from Hericium coralloides and Trametes versicolor Exert Differential Immunomodulatory Effects on Immune Cells from Older Adults In Vitro" Nutrients 15, no. 9: 2227. https://doi.org/10.3390/nu15092227
APA StyleWilliams, L. M., Berthon, B. S., Stoodley, I. L., Williams, E. J., & Wood, L. G. (2023). Medicinal Mushroom Extracts from Hericium coralloides and Trametes versicolor Exert Differential Immunomodulatory Effects on Immune Cells from Older Adults In Vitro. Nutrients, 15(9), 2227. https://doi.org/10.3390/nu15092227