
Is There a FADS2-Modulated Link between Long-Chain Polyunsaturated Fatty Acids in Plasma Phospholipids and Polyphenol Intake in Adult Subjects Who Are Overweight?

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Clearance, Study Design and Intervention Treatments
2.2. Dietary Intake and Anthropometric Data Assessments
2.3. Blood Sample Collection and Processing
2.4. Plasma Phospholipid Fatty Acid Analysis
2.5. Genotyping
2.6. Statistical Analyses
3. Results
3.1. Clinical Characteristics and FADS2 Genotypes of the Study Subjects Who Are Overweight
3.2. Effects of the 4-Week Interventional Treatment with Varying Polyphenol Content on Changes in BMI, Total Body Weight and Levels of LC-PUFA in Plasma Phospholipids in the Subjects Who Are Overweight
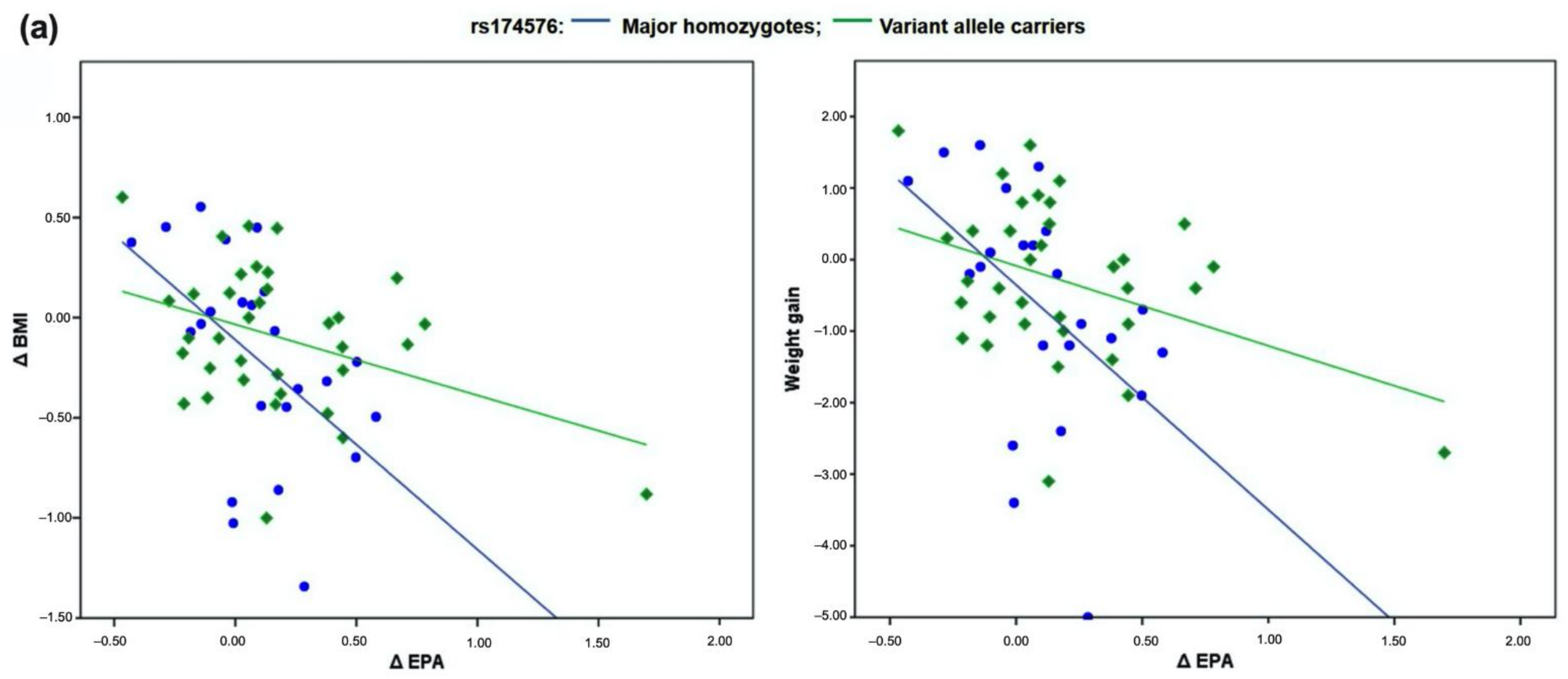
3.3. Relationship between Dietary Intake of Polyphenols from Aronia Juice, FADS2 Genotypes, Levels of LC-PUFA in Plasma Phospholipids and Obesity Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Total (n = 62) | AJ (n = 22) | MJ (n = 20) | PLB (n = 20) | |
|---|---|---|---|---|
| Age | 41.05 ± 0.85 | 42.73 ± 1.54 | 40.50 ± 1.64 | 39.75 ± 1.19 |
| Women, n(%) | 34 (54.8) | 13 (59.1) | 13 (65) | 8 (40) |
| FADS2 Distribution | ||||
| rs174593 T > C, n(%) c | 35 (56.5) | 11 (50) | 9 (45) | 15 (75) |
| rs174616 G > A, n(%) c | 48 (77.4) | 17 (77.3) | 15 (75) | 16 (80) |
| rs174576 C > A, n(%) c | 39 (62.9) | 13 (59.1) | 11 (55) | 15 (75) |
| Daily Dietary Intake | ||||
| TE, kcal | 1878.21 [1587.90, 2387.69] | 1849.68 [1654.49, 2436.93] | 1911.91 [1615.89, 2217.07] | 2165.51 [1456.10, 2748.90] |
| Carbohydrate, % TE | 44.13 ± 1.00 | 44.44 ± 1.26 | 42.51 ± 2.17 | 45.38 ± 1.82 |
| Protein, % TE | 16.05 [14.69, 17.49] | 16.04 [14.90, 17.58] | 16.53 [15.02, 17.54] | 15.83 [13.59, 17.40] |
| Fat, % TE | 39.67 ± 0.80 | 39.19 ± 1.01 | 40.69 ± 1.46 | 39.20 ± 1.74 |
| Omega-6, g | 18.05 [14.96, 24.16] | 17.24 [13.79, 24.85] | 19.84 [17.28, 23.16] | 16.62 [12.78, 26.65] |
| Omega-3, g | 1.12 [0.76, 1.59] | 0.96 [0.79, 1.41] | 1.16 [0.65, 1.77] | 1.16 [0.82, 1.63] |
| Omega-6/Omega-3 | 16.71 [12.71, 24.96] | 17.25 [12.92, 21.43] | 20.16 [12.56, 26.63] | 15.71 [12.31, 25.05] |
| Clinical Biomarkers | ||||
| Weight, kg | 92.25 [80.58, 105.65] | 89.40 [76.08, 99.78] | 89.90 [73.00, 107.53] | 93.45 [82.90, 109.85] |
| BMI, kg/m2 | 29.51 [26.45, 32.43] | 28.38 [26.44, 34.14] | 30.25 [26.33, 32.35] | 29.22 [27.11, 32.80] |
| FPG, mmol/L | 5.06 [4.73, 5.52] | 4.95 [4.66, 5.52] a | 4.95 [4.55, 5.18] a | 5.27 [4.94, 6.26] b |
| TAG, mmol/L | 1.24 [0.84, 1.70] | 1.16 [0.64, 1.91] | 1.16 [0.75, 1.35] | 1.44 [0.99, 1.73] |
| TC, mmol/L | 5.30 ± 0.13 | 5.43 ± 0.25 | 5.14 ± 0.21 | 5.32 ± 0.24 |
| HDL-c, mmol/L | 1.51 ± 0.05 | 1.56 ± 0.12 | 1.60 ± 0.07 | 1.36 ± 0.07 |
| LDL-c, mmol/L | 3.68 ± 0.13 | 3.74 ± 0.25 | 3.50 ± 0.20 | 3.79 ± 0.22 |
| Creatinine, μmol/L | 72.10 ± 1.62 | 72.96 ± 2.75 | 70.50 ± 3.36 | 72.77 ± 2.31 |
| Urea, mmol/L | 3.51 [2.94, 4.16] | 3.59 [3.11, 4.26] | 3.12 [2.75, 4.08] | 3.62 [2.66, 4.43] |
| ALT, U/L | 25.55 [20.40, 38.45] | 29.55 [21.70, 40.80] | 22.05 [17.60, 26.90] | 30.95 [21.28, 37.48] |
| AST, U/L | 20.90 [18.38, 25.10] | 24.45 [18.75, 28.60] | 19.90 [17.63, 23.88] | 20.50 [18.45, 25.05] |
| GGT, U/L | 16.85 [10.88, 24.68] | 20.80 [10.48, 34.65] | 14.00 [10.18, 22.33] | 18.25 [11.53, 22.85] |
| LDH, U/L | 149.70 [134.43, 169.78] | 160.20 [145.30, 179.90] | 146.80 [133.25, 168.83] | 142.70 [128.73, 156.58] |
References
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/en/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 16 November 2020).
- Abdullah, A.; Peeters, A.; de Courten, M.; Stoelwinder, J. The magnitude of association between overweight and obesity and the risk of diabetes: A meta-analysis of prospective cohort studies. Diabetes Res. Clin. Pr. 2010, 89, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Fekete, K.; Györei, E.; Lohner, S.; Verduci, E.; Agostoni, C.; Decsi, T. Long-chain polyunsaturated fatty acid status in obesity: A systematic review and meta-analysis. Obes. Rev. 2015, 16, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Mayneris-Perxachs, J.; Guerendiain, M.; Castellote, A.I.; Estruch, R.; Covas, M.I.; Fitó, M.; Salas-Salvadó, J.; Martínez-González, M.A.; Aros, F.; Lamuela-Raventós, R.M.; et al. Plasma fatty acid composition, estimated desaturase activities, and their relation with the metabolic syndrome in a population at high risk of cardiovascular disease. Clin. Nutr. 2014, 33, 90–97. [Google Scholar] [CrossRef]
- Calder, P.C. Very long chain omega-3 (n − 3) fatty acids and human health. Eur. J. Lipid Sci. Technol. 2014, 116, 1280–1300. [Google Scholar] [CrossRef]
- Sonnweber, T.; Pizzini, A.; Nairz, M.; Weiss, G.; Tancevski, I. Arachidonic acid metabolites in cardiovascular and metabolic diseases. Int. J. Mol. Sci. 2018, 19, 3285. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; Reischl, E.; Tanjung, C.; Gonzalez-Casanova, I.; Ramakrishnan, U.; Meldrum, S.; Simmer, K.; Heinrich, J.; Demmelmair, H. FADS1 and FADS2 Polymorphisms Modulate Fatty Acid Metabolism and Dietary Impact on Health. Annu. Rev. Nutr. 2019, 39, 21–44. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, L.; Gohlke, H.; Müller, M.; Heid, I.M.; Palmer, L.J.; Kompauer, I.; Demmelmair, H.; Illig, T.; Koletzko, B.; Heinrich, J. Common genetic variants of the FADS1 FADS2 gene cluster and their reconstructed haplotypes are associated with the fatty acid composition in phospholipids. Hum. Mol. Genet. 2006, 15, 1745–1756. [Google Scholar] [CrossRef]
- Tanaka, T.; Shen, J.; Abecasis, G.R.; Kisialiou, A.; Ordovas, J.M.; Guralnik, J.M.; Singleton, A.; Bandinelli, S.; Cherubini, A.; Arnett, D.; et al. Genome-Wide Association Study of Plasma Polyunsaturated Fatty Acids in the InCHIANTI Study. PLoS Genet. 2009, 5, e1000338. [Google Scholar] [CrossRef]
- Sergeant, S.; Hugenschmidt, C.E.; Rudock, M.E.; Ziegler, J.T.; Ivester, P.; Ainsworth, H.C.; Vaidya, D.; Case, L.D.; Langefeld, C.D.; Freedman, B.I.; et al. Differences in arachidonic acid levels and fatty acid desaturase (FADS) gene variants in African Americans and European Americans with diabetes or the metabolic syndrome. Br. J. Nutr. 2012, 107, 547–555. [Google Scholar] [CrossRef]
- De la Garza Puentes, A.; Goyanes, R.M.; Tonato, A.M.C.; Torres-Espínola, F.J.; García, M.A.; de Almeida, L.; Aguirre, M.B.; Guerendiain, M.; Bargalló, A.I.C.; Moreno, M.S.; et al. Association of maternal weight with FADS and ELOVL genetic variants and fatty acid levels—The PREOBE follow-up. PLoS ONE 2017, 12, 1–16. [Google Scholar] [CrossRef]
- Khamlaoui, W.; Mehri, S.; Hammami, S.; Hammouda, S.; Chraeif, I.; Elosua, R.; Hammami, M. Association Between Genetic Variants in FADS1-FADS2 and ELOVL2 and Obesity, Lipid Traits, and Fatty Acids in Tunisian Population. Clin. Appl. Thromb. 2020, 26, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Krga, I.; Milenkovic, D.; Morand, C.; Monfoulet, L.-E. An update on the role of nutrigenomic modulations in mediating the cardiovascular protective effect of fruit polyphenols. Food Funct. 2016, 7, 3656–3676. [Google Scholar] [CrossRef] [PubMed]
- Krga, I.; Milenkovic, D. Anthocyanins: From Sources and Bioavailability to Cardiovascular-Health Benefits and Molecular Mechanisms of Action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef] [PubMed]
- Kardum, N.; Petrović-Oggiano, G.; Takic, M.; Glibetić, N.; Zec, M.; Debeljak-Martacic, J.; Konić-Ristić, A. Effects of glucomannan-enriched, aronia juice-based supplement on cellular antioxidant enzymes and membrane lipid status in subjects with abdominal obesity. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Kardum, N.; Takić, M.; Šavikin, K.; Zec, M.; Zdunić, G.; Spasić, S.; Konić-Ristić, A. Effects of polyphenol-rich chokeberry juice on cellular antioxidant enzymes and membrane lipid status in healthy women. J. Funct. Foods 2014, 9, 89–97. [Google Scholar] [CrossRef]
- Rienks, J.; Barbaresko, J.; Nöthlings, U. Association of polyphenol biomarkers with cardiovascular disease and mortality risk: A systematic review and Meta-Analysis of observational studies. Nutrients 2017, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Rienks, J.; Barbaresko, J.; Oluwagbemigun, K.; Schmid, M.; Nöthlings, U. Polyphenol exposure and risk of type 2 diabetes: Dose-response meta-analyses and systematic review of prospective cohort studies. Am. J. Clin. Nutr. 2018, 108, 49–61. [Google Scholar] [CrossRef]
- Del Bo, C.; Bernardi, S.; Marino, M.; Porrini, M.; Tucci, M.; Guglielmetti, S.; Cherubini, A.; Carrieri, B.; Kirkup, B.; Kroon, P.; et al. Systematic Review on Polyphenol Intake and Health Outcomes: Is there Sufficient Evidence to Define a Health-Promoting Polyphenol-Rich Dietary Pattern? Nutrients 2019, 11, 1355. [Google Scholar] [CrossRef]
- Istas, G.; Wood, E.; Le Sayec, M.; Rawlings, C.; Yoon, J.; Dandavate, V.; Cera, D.; Rampelli, S.; Costabile, A.; Fromentin, E.; et al. Effects of aronia berry (poly)phenols on vascular function and gut microbiota: A double-blind randomized controlled trial in adult men. Am. J. Clin. Nutr. 2019, 110, 316–329. [Google Scholar] [CrossRef]
- Krga, I.; Vidovic, N.; Milenkovic, D.; Konic-Ristic, A.; Stojanovic, F.; Morand, C.; Glibetic, M. Effects of anthocyanins and their gut metabolites on adenosine diphosphate-induced platelet activation and their aggregation with monocytes and neutrophils. Arch. Biochem. Biophys. 2018, 645, 34–41. [Google Scholar] [CrossRef]
- Krga, I.; Tamaian, R.; Mercier, S.; Boby, C.; Monfoulet, L.-E.; Glibetic, M.; Morand, C.; Milenkovic, D. Anthocyanins and their gut metabolites attenuate monocyte adhesion and transendothelial migration through nutrigenomic mechanisms regulating endothelial cell permeability. Free Radic. Biol. Med. 2018, 124, 364–379. [Google Scholar] [CrossRef] [PubMed]
- García-Conesa, M.T.; Chambers, K.; Combet, E.; Pinto, P.; Garcia-Aloy, M.; Andrés-Lacueva, C.; de Pascual-Teresa, S.; Mena, P.; Ristic, A.K.; Hollands, W.J.; et al. Meta-analysis of the effects of foods and derived products containing ellagitannins and anthocyanins on cardiometabolic biomarkers: Analysis of factors influencing variability of the individual responses. Int. J. Mol. Sci. 2018, 19, 694. [Google Scholar] [CrossRef] [PubMed]
- Ruskovska, T.; Massaro, M.; Carluccio, M.A.; Arola-Arnal, A.; Muguerza, B.; Berghe, W.V.; Declerck, K.; Bravo, F.I.; Calabriso, N.; Combet, E.; et al. Systematic bioinformatic analysis of nutrigenomic data of flavanols in cell models of cardiometabolic disease. Food Funct. 2020, 11, 5040–5064. [Google Scholar] [CrossRef] [PubMed]
- Barth, S.W.; Koch, T.C.L.; Watzl, B.; Dietrich, H.; Will, F.; Bub, A. Moderate effects of apple juice consumption on obesity-related markers in obese men: Impact of diet–gene interaction on body fat content. Eur. J. Nutr. 2012, 51, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, D.; Morand, C.; Cassidy, A.; Konic-Ristic, A.; Tomás-Barberán, F.; Ordovas, J.M.; Kroon, P.; de Caterina, R.; Rodriguez-Mateos, A. Interindividual variability in biomarkers of cardiometabolic health after consumption of major plant-food bioactive compounds and the determinants involved. Adv. Nutr. 2017, 8, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, F.; Conti, C.; Dogliotti, E.; Terranegra, A.; Salvi, E.; Braga, D.; Ricca, F.; Lupoli, S.; Mingione, A.; Pivari, F.; et al. Interaction between polyphenols intake and PON1 gene variants on markers of cardiovascular disease: A nutrigenetic observational study. J. Transl. Med. 2016, 14, 186. [Google Scholar] [CrossRef] [PubMed]
- Zec, M.M.; Stojković, L.; Zeković, M.; Pokimica, B.; Zivkovic, M.; Stankovic, A.; Glibetic, M. FADS2 polymorphisms are associated with plasma arachidonic acid and estimated desaturase-5 activity in a cross-sectional study. Nutr. Res. 2020, 83, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Pokimica, B.; García-Conesa, M.-T.; Zec, M.; Debeljak-Martačić, J.; Ranković, S.; Vidović, N.; Petrović-Oggiano, G.; Konić-Ristić, A.; Glibetić, M. Chokeberry Juice Containing Polyphenols Does Not Affect Cholesterol or Blood Pressure but Modifies the Composition of Plasma Phospholipids Fatty Acids in Individuals at Cardiovascular Risk. Nutrients 2019, 11, 850. [Google Scholar] [CrossRef]
- Kardum, N.; Ristic, A.K.; Zec, M.; Kojadinovic, M.; Petrovic-Oggiano, G.; Zekovic, M.; Kroon, P.A.; Glibetić, M. Design, formulation and sensory evaluation of a polyphenol-rich food placebo: An example of aronia juice for food intervention studies. Int. J. Food Sci. Nutr. 2017, 68, 742–749. [Google Scholar] [CrossRef]
- Zec, M.; Martačić, J.D.; Ranković, S.; Pokimica, B.; Tomić, M.; Ignjatović, D.; Glibetić, M.; Popović, T. Effects of Aronia melanocarpa juice on plasma and liver phospholipid fatty acid composition in Wistar rats. Acta Vet. 2017, 67, 107–120. [Google Scholar] [CrossRef][Green Version]
- Gurinović, M.; Milešević, J.; Kadvan, A.; Nikolić, M.; Zeković, M.; Djekić-Ivanković, M.; Dupouy, E.; Finglas, P.; Glibetić, M. Development, features and application of DIET ASSESS & PLAN (DAP) software in supporting public health nutrition research in Central Eastern European Countries (CEEC). Food Chem. 2018, 238, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Gurinović, M.; Milešević, J.; Kadvan, A.; Djekić-Ivanković, M.; Debeljak-Martačić, J.; Takić, M.; Nikolić, M.; Ranković, S.; Finglas, P.; Glibetić, M. Establishment and advances in the online Serbian food and recipe data base harmonized with EuroFIR™ standards. Food Chem. 2016, 193, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Kunkel, L.M.; Smith, K.D.; Boyer, S.H.; Borgaonkar, D.S.; Wachtel, S.S.; Miller, O.J.; Breg, W.R.; Jones, H.W.; Rary, J.M. Analysis of human Y chromosome specific reiterated DNA in chromosome variants. Proc. Natl. Acad. Sci. USA 1977, 74, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.J.; Jackson, K.G.; Dadd, T.; Mayes, A.E.; Brown, A.L.; Lovegrove, J.A.; Minihane, A.M. The impact of the catechol-O-methyltransferase genotype on vascular function and blood pressure after acute green tea ingestion. Mol. Nutr. Food Res. 2012, 56, 966–975. [Google Scholar] [CrossRef]
- Gonzalez-Soto, M.; Mutch, D.M. Diet Regulation of Long-Chain PUFA Synthesis: Role of Macronutrients, Micronutrients, and Polyphenols on Δ-5/Δ-6 Desaturases and Elongases 2/5. Adv. Nutr. 2020. online ahead of print. [Google Scholar] [CrossRef]
- Martin, K.R. Polyphenols as dietary supplements: A double-edged sword. Nutr. Diet. Suppl. 2009, 2, 1–12. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Tjønneland, A.; Kyrø, C.; Fagherazzi, G.; Boutron-Ruault, M.-C.; et al. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef]
- Shrime, M.G.; Bauer, S.R.; McDonald, A.C.; Chowdhury, N.H.; Coltart, C.E.M.; Ding, E.L. Flavonoid-Rich Cocoa Consumption Affects Multiple Cardiovascular Risk Factors in a Meta-Analysis of Short-Term Studies. J. Nutr. 2011, 141, 1982–1988. [Google Scholar] [CrossRef]
- Rodriguez-Mateos, A.; Rendeiro, C.; Bergillos-meca, T.; Tabatabaee, S.; George, T.W.; Heiss, C.; Spencer, J.P.E. Intake and time dependence of blueberry flavonoid—Induced improvements in vascular function: A randomized, controlled, double-blind, crossover intervention study with mechanistic insights into biological activity. Am. J. Clin. Nutr. 2013, 98, 1179–1191. [Google Scholar] [CrossRef]
- Rodriguez-Mateos, A.; Feliciano, R.P.; Boeres, A.; Weber, T.; dos Santos, C.N.; Ventura, M.R.; Heiss, C. Cranberry (poly)phenol metabolites correlate with improvements in vascular function: A double-blind, randomized, controlled, dose-response, crossover study. Mol. Nutr. Food Res. 2016, 60, 2130–2140. [Google Scholar] [CrossRef] [PubMed]
- Bertoia, M.L.; Rimm, E.B.; Mukamal, K.J.; Hu, F.B.; Willett, W.C.; Cassidy, A. Dietary flavonoid intake and weight maintenance: Three prospective cohorts of 124 086 US men and women followed for up to 24 years. BMJ 2016, 352. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.A.E.; Arts, I.C.W.; Ambergen, T.; Brants, H.A.M.; Dagnelie, P.C.; Goldbohm, R.A.; van den Brandt, P.A.; Weijenberg, M.P. Higher dietary flavone, flavonol, and catechin intakes are associated with less of an increase in BMI over time in women: A longitudinal analysis from the Netherlands Cohort Study. Am. J. Clin. Nutr. 2008, 88, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Tabrizi, R.; Tamtaji, O.R.; Lankarani, K.B.; Akbari, M.; Dadgostar, E.; Dabbaghmanesh, M.H.; Kolahdooz, F.; Shamshirian, A.; Momen-Heravi, M.; Asemi, Z. The effects of resveratrol intake on weight loss: A systematic review and meta-analysis of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 375–390. [Google Scholar] [CrossRef]
- Mousavi, S.M.; Milajerdi, A.; Sheikhi, A.; Kord-Varkaneh, H.; Feinle-Bisset, C.; Larijani, B.; Esmaillzadeh, A. Resveratrol supplementation significantly influences obesity measures: A systematic review and dose–response meta-analysis of randomized controlled trials. Obes. Rev. 2019, 20, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef]
- Mohammadpour, S.; Amini, M.R.; Shahinfar, H.; Tijani, A.J.; Shahavandi, M.; Ghorbaninejad, P.; Djafarian, K.; Shab-Bidar, S. Effects of glucomannan supplementation on weight loss in overweight and obese adults: A systematic review and meta-analysis of randomized controlled trials. Obes. Med. 2020, 19, 100276. [Google Scholar] [CrossRef]
- Ojwang, A.A.; Kruger, H.S.; Zec, M.; Ricci, C.; Pieters, M.; Kruger, I.M.; Wentzel-Viljoen, E.; Smuts, C.M. Plasma phospholipid fatty acid patterns are associated with adiposity and the metabolic syndrome in black South Africans: A cross-sectional study. Cardiovasc. J. Afr. 2019, 30, 228–238. [Google Scholar] [CrossRef]
- Loo, B.-M.; Erlund, I.; Koli, R.; Puukka, P.; Hellström, J.; Wähälä, K.; Mattila, P.; Jula, A. Consumption of chokeberry (Aronia mitschurinii) products modestly lowered blood pressure and reduced low-grade inflammation in patients with mildly elevated blood pressure. Nutr. Res. 2016, 36, 1222–1230. [Google Scholar] [CrossRef]
- Kim, B.; Park, Y.; Wegner, C.J.; Bolling, B.W.; Lee, J. Polyphenol-rich black chokeberry (Aronia melanocarpa) extract regulates the expression of genes critical for intestinal cholesterol flux in Caco-2 cells. J. Nutr. Biochem. 2013, 24, 1564–1570. [Google Scholar] [CrossRef]
- Ninomiya, T.; Nagata, M.; Hata, J.; Hirakawa, Y.; Ozawa, M.; Yoshida, D.; Ohara, T.; Kishimoto, H.; Mukai, N.; Fukuhara, M.; et al. Association between ratio of serum eicosapentaenoic acid to arachidonic acid and risk of cardiovascular disease: The Hisayama Study. Atherosclerosis 2013, 231, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Kawano, T.; Ukawa, Y.; Sagesaka, Y.M.; Fukuhara, I. Green tea beverages enriched with catechins with a galloyl moiety reduce body fat in moderately obese adults: A randomized double-blind placebo-controlled trial. Food Funct. 2016, 7, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Moustaid-Moussa, N.; Chen, L.; Mo, H.; Shastri, A.; Su, R.; Bapat, P.; Kwun, I.; Shen, C.-L. Novel insights of dietary polyphenols and obesity. J. Nutr. Biochem. 2014, 25, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Galleano, M.; Calabro, V.; Prince, P.D.; Litterio, M.C.; Piotrkowski, B.; Vazquez-Prieto, M.A.; Miatello, R.M.; Oteiza, P.I.; Fraga, C.G. Flavonoids and metabolic syndrome. Ann. N. Y. Acad. Sci. 2012, 1259, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Chimhashu, T.; Malan, L.; Baumgartner, J.; van Jaarsveld, P.J.; Galetti, V.; Moretti, D.; Smuts, C.M.; Zimmermann, M.B. Sensitivity of fatty acid desaturation and elongation to plasma zinc concentration: A randomised controlled trial in Beninese children. Br. J. Nutr. 2018, 119, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Knez, M.; Pantovic, A.; Zekovic, M.; Pavlovic, Z.; Glibetic, M.; Zec, M. Is there a link between zinc intake and status with plasma fatty acid profile and desaturase activities in dyslipidemic subjects? Nutrients 2020, 12, 93. [Google Scholar] [CrossRef]
- Calder, P.C. Eicosapentaenoic and docosahexaenoic acid derived specialised pro-resolving mediators: Concentrations in humans and the effects of age, sex, disease and increased omega-3 fatty acid intake. Biochimie 2020, 178, 105–123. [Google Scholar] [CrossRef]
- Takahashi, M.; Ando, J.; Shimada, K.; Nishizaki, Y.; Tani, S.; Ogawa, T.; Yamamoto, M.; Nagao, K.; Hirayama, A.; Yoshimura, M.; et al. The ratio of serum n-3 to n-6 polyunsaturated fatty acids is associated with diabetes mellitus in patients with prior myocardial infarction: A multicenter cross-sectional study. BMC Cardiovasc. Disord. 2017, 17, 41. [Google Scholar] [CrossRef]
- Tutino, V.; de Nunzio, V.; Caruso, M.G.; Bonfiglio, C.; Franco, I.; Mirizzi, A.; de Leonardis, G.; Cozzolongo, R.; Giannuzzi, V.; Giannelli, G.; et al. Aerobic physical activity and a low glycemic diet reduce the AA/EPA ratio in red blood cell membranes of patients with NAFLD. Nutrients 2018, 10, 1299. [Google Scholar] [CrossRef]
- Chilton, F.; Dutta, R.; Reynolds, L.; Sergeant, S.; Mathias, R.; Seeds, M. Precision Nutrition and Omega-3 Polyunsaturated Fatty Acids: A Case for Personalized Supplementation Approaches for the Prevention and Management of Human Diseases. Nutrients 2017, 9, 1165. [Google Scholar] [CrossRef]



| rs174593 | rs174616 | rs174576 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ΔR2 | B ± S.E. | p | ΔR2 | B ± S.E. | p | ΔR2 | B ± S.E. | p | ||
| Δ Linoleic acid, C18:3n − 6 | ||||||||||
| M1 | TS | 0.001 | −0.15 ± 0.57 | 0.789 | 0.067 | −1.05 ± 0.53 | 0.051 | 0.029 | −0.72 ± 0.56 | 0.204 |
| M2 | 0.042 | −0.11 ± 0.58 | 0.851 | 0.034 | −0.98 ± 0.53 | 0.070 | 0.032 | −0.57 ± 0.57 | 0.320 | |
| M1 | AJ | 0.250 | −2.85 ± 1.11 | 0.018 | 0.341 | −2.47 ± 0.77 | 0.004 | 0.239 | −2.46 ± 0.98 | 0.021 |
| M2 | 0.181 | −2.12 ± 1.06 | 0.062 | 0.175 | −2.16 ± 0.77 | 0.012 | 0.197 | −1.98 ± 0.97 | 0.056 | |
| M1 | MJ | 0.058 | −0.86 ± 0.86 | 0.337 | 0.131 | −1.33 ± 0.86 | 0.140 | 0.345 | −2.12 ± 0.73 | 0.010 |
| M2 | 0.236 | −1.16 ± 0.83 | 0.183 | 0.193 | −1.33 ± 0.81 | 0.123 | 0.074 | −1.81 ± 0.78 | 0.036 | |
| M1 | PLB | 0.149 | 1.51 ± 0.93 | 0.126 | 0.022 | 0.58 ± 1.00 | 0.569 | 0.092 | 1.19 ± 0.96 | 0.237 |
| M2 | 0.110 | 1.88 ± 1.01 | 0.085 | 0.079 | 0.85 ± 1.13 | 0.465 | 0.096 | 1.51 ± 1.06 | 0.178 | |
| Δ Arachidonic acid, C20:4n − 6 | ||||||||||
| M1 | TS | 0.038 | 0.62 ± 0.43 | 0.149 | 0.014 | −0.37 ± 0.42 | 0.382 | 0.000 | −0.07 ± 0.43 | 0.875 |
| M2 | 0.012 | 0.68 ± 0.44 | 0.130 | 0.005 | −0.34 ± 0.43 | 0.422 | 0.006 | −0.02 ± 0.45 | 0.972 | |
| M1 | AJ MJ | 0.198 | 1.10 ± 0.50 | 0.038 | 0.003 | −0.10 ± 0.41 | 0.812 | 0.078 | 0.61 ± 0.47 | 0.208 |
| M2 | 0.015 | 1.14 ± 0.54 | 0.049 | 0.017 | −0.11 ± 0.48 | 0.818 | 0.033 | 0.73 ± 0.53 | 0.183 | |
| M1 | MJ | 0.000 | 0.06 ± 0.78 | 0.943 | 0.184 | −1.38 ± 0.73 | 0.076 | 0.005 | −0.23 ± 0.79 | 0.773 |
| M2 | 0.014 | −0.04 ± 0.86 | 0.967 | 0.016 | −1.39 ± 0.77 | 0.094 | 0.012 | −0.18 ± 0.89 | 0.844 | |
| M1 | PLB | 0.000 | 0.02 ± 0.98 | 0.987 | 0.021 | −0.54 ± 0.97 | 0.582 | 0.117 | −1.29 ± 0.92 | 0.179 |
| M2 | 0.151 | −0.15 ± 1.04 | 0.886 | 0.172 | −0.87 ± 1.04 | 0.417 | 0.201 | −1.69 ± 0.94 | 0.096 | |
| Δ Adrenic acid, C22:4n − 6 | ||||||||||
| M1 | TS | 0.070 | 0.09 ± 0.04 | 0.047 | 0.015 | 0.04 ± 0.04 | 0.360 | 0.126 | 0.12 ± 0.04 | 0.007 |
| M2 | 0.040 | 0.08 ± 0.04 | 0.063 | 0.045 | 0.03 ± 0.04 | 0.452 | 0.024 | 0.11 ± 0.04 | 0.016 | |
| M1 | AJ | 0.007 | −0.03 ± 0.07 | 0.708 | 0.064 | 0.06 ± 0.05 | 0.255 | 0.037 | 0.05 ± 0.06 | 0.393 |
| M2 | 0.303 | −0.04 ± 0.06 | 0.540 | 0.237 | 0.02 ± 0.05 | 0.706 | 0.259 | 0.01 ± 0.06 | 0.881 | |
| M1 | MJ | 0.077 | 0.13 ± 0.11 | 0.264 | 0.025 | −0.08 ± 0.12 | 0.533 | 0.199 | 0.21 ± 0.11 | 0.064 |
| M2 | 0.120 | 0.13 ± 0.11 | 0.263 | 0.121 | −0.08 ± 0.12 | 0.519 | 0.055 | 0.18 ± 0.12 | 0.135 | |
| M1 | PLB | 0.203 | 0.11 ± 0.06 | 0.070 | 0.127 | 0.09 ± 0.06 | 0.161 | 0.117 | 0.08 ± 0.06 | 0.179 |
| M2 | 0.101 | 0.08 ± 0.06 | 0.199 | 0.118 | 0.05 ± 0.06 | 0.427 | 0.125 | 0.05 ± 0.06 | 0.444 | |
| ΔEicosapentaenoic acid, C20:5 n − 3 | ||||||||||
| M1 | TS | 0.008 | −0.05 ± 0.07 | 0.516 | 0.003 | 0.03 ± 0.07 | 0.672 | 0.001 | 0.02 ± 0.07 | 0.784 |
| M2 | 0.020 | −0.05 ± 0.08 | 0.514 | 0.018 | 0.03 ± 0.07 | 0.734 | 0.018 | 0.01 ± 0.08 | 0.917 | |
| M1 | AJ | 0.000 | −0.01 ± 0.19 | 0.968 | 0.100 | 0.20 ± 0.13 | 0.152 | 0.047 | 0.16 ± 0.16 | 0.331 |
| M2 | 0.125 | −0.05 ± 0.20 | 0.813 | 0.062 | 0.14 ± 0.15 | 0.370 | 0.086 | 0.09 ± 0.18 | 0.641 | |
| M1 | MJ | 0.079 | −0.14 ± 0.12 | 0.259 | 0.234 | −0.26 ± 0.12 | 0.042 | 0.021 | −0.07 ± 0.13 | 0.570 |
| M2 | 0.102 | −0.15 ± 0.13 | 0.253 | 0.103 | −0.26 ± 0.12 | 0.041 | 0.140 | −0.14 ± 0.13 | 0.323 | |
| M1 | PLB | 0.016 | 0.04 ± 0.09 | 0.627 | 0.028 | 0.06 ± 0.09 | 0.523 | 0.001 | 0.01 ± 0.09 | 0.903 |
| M2 | 0.078 | 0.01 ± 0.10 | 0.954 | 0.068 | 0.02 ± 0.10 | 0.866 | 0.101 | −0.03 ± 0.10 | 0.736 | |
| Δ Clupanodonic acid, C22:5n − 3 | ||||||||||
| M1 | TS | 0.046 | 0.13 ± 0.08 | 0.110 | 0.007 | 0.05 ± 0.08 | 0.541 | 0.024 | 0.09 ± 0.08 | 0.254 |
| M2 | 0.018 | 0.12 ± 0.08 | 0.153 | 0.027 | 0.05 ± 0.08 | 0.540 | 0.029 | 0.10 ± 0.08 | 0.236 | |
| M1 | AJ | 0.014 | 0.03 ± 0.05 | 0.596 | 0.030 | −0.03 ± 0.04 | 0.438 | 0.003 | 0.01 ± 0.05 | 0.797 |
| M2 | 0.025 | 0.02 ± 0.06 | 0.726 | 0.082 | −0.06 ± 0.04 | 0.220 | 0.029 | 0.00 ± 0.05 | 0.953 | |
| M1 | MJ | 0.045 | 0.21 ± 0.24 | 0.396 | 0.003 | 0.05 ± 0.25 | 0.840 | 0.011 | 0.11 ± 0.24 | 0.673 |
| M2 | 0.021 | 0.18 ± 0.26 | 0.507 | 0.036 | 0.05 ± 0.26 | 0.852 | 0.050 | 0.17 ± 0.27 | 0.546 | |
| M1 | PLB | 0.196 | 0.16 ± 0.08 | 0.075 | 0.254 | 0.18 ± 0.08 | 0.039 | 0.228 | 0.17 ± 0.08 | 0.053 |
| M2 | 0.042 | 0.13 ± 0.09 | 0.174 | 0.030 | 0.16 ± 0.09 | 0.106 | 0.035 | 0.15 ± 0.09 | 0.133 | |
| ΔDocosahexaenoic acid, C22:6n − 3 | ||||||||||
| M1 | TS | 0.004 | 0.10 ± 0.21 | 0.630 | 0.007 | −0.12 ± 0.20 | 0.548 | 0.014 | 0.18 ± 0.21 | 0.388 |
| M2 | 0.052 | 0.09 ± 0.21 | 0.676 | 0.058 | −0.16 ± 0.20 | 0.439 | 0.046 | 0.12 ± 0.21 | 0.579 | |
| M1 | AJ | 0.103 | 0.77 ± 0.51 | 0.146 | 0.010 | 0.18 ± 0.40 | 0.658 | 0.121 | 0.74 ± 0.44 | 0.113 |
| M2 | 0.020 | 0.68 ± 0.55 | 0.233 | 0.038 | 0.02 ± 0.46 | 0.968 | 0.011 | 0.66 ± 0.51 | 0.205 | |
| M1 | MJ | 0.001 | −0.05 ± 0.39 | 0.908 | 0.187 | −0.69 ± 0.36 | 0.073 | 0.003 | 0.08 ± 0.39 | 0.832 |
| M2 | 0.203 | 0.01 ± 0.38 | 0.977 | 0.208 | −0.70 ± 0.33 | 0.054 | 0.210 | −0.16 ± 0.39 | 0.688 | |
| M1 | PLB | 0.011 | −0.10 ± 0.24 | 0.685 | 0.001 | 0.03 ± 0.24 | 0.897 | 0.008 | −0.08 ± 0.24 | 0.735 |
| M2 | 0.009 | −0.06 ± 0.28 | 0.822 | 0.024 | 0.10 ± 0.29 | 0.733 | 0.010 | −0.04 ± 0.28 | 0.879 | |
| Δ Destaurase-5 predicted activity, AA/DGLA | ||||||||||
| M1 | TS | 0.004 | −0.15 ± 0.32 | 0.646 | 0.034 | −0.43 ± 0.31 | 0.167 | 0.077 | −0.66 ± 0.31 | 0.037 |
| M2 | 0.050 | −0.12 ± 0.32 | 0.718 | 0.044 | −0.38 ± 0.31 | 0.219 | 0.031 | −0.58 ± 0.31 | 0.073 | |
| M1 | AJ | 0.097 | −0.78 ± 0.53 | 0.159 | 0.001 | −0.06 ± 0.42 | 0.880 | 0.135 | −0.81 ± 0.46 | 0.092 |
| M2 | 0.070 | −0.62 ± 0.56 | 0.284 | 0.124 | 0.25 ± 0.46 | 0.585 | 0.042 | −0.62 ± 0.51 | 0.241 | |
| M1 | MJ | 0.002 | −0.09 ± 0.49 | 0.851 | 0.420 | −1.30 ± 0.38 | 0.004 | 0.050 | −0.44 ± 0.48 | 0.372 |
| M2 | 0.036 | −0.19 ± 0.53 | 0.729 | 0.033 | −1.31 ± 0.40 | 0.005 | 0.021 | −0.43 ± 0.54 | 0.441 | |
| M1 | PLB | 0.002 | −0.15 ± 0.76 | 0.850 | 0.023 | −0.45 ± 0.75 | 0.562 | 0.134 | −1.08 ± 0.71 | 0.148 |
| M2 | 0.153 | −0.24 ± 0.81 | 0.771 | 0.165 | −0.63 ± 0.81 | 0.450 | 0.189 | −1.33 ± 0.73 | 0.091 | |
| Δ Destaurase-6 predicted activity, DGLA/LA | ||||||||||
| M1 | TS | 0.041 | 0.01 ± 0.01 | 0.130 | 0.075 | 0.02 ± 0.01 | 0.039 | 0.139 | 0.02 ± 0.01 | 0.004 |
| M2 | 0.061 | 0.01 ± 0.01 | 0.128 | 0.051 | 0.02 ± 0.01 | 0.054 | 0.035 | 0.02 ± 0.01 | 0.010 | |
| M1 | AJ | 0.232 | 0.04 ± 0.02 | 0.023 | 0.095 | 0.02 ± 0.01 | 0.163 | 0.332 | 0.04 ± 0.01 | 0.005 |
| M2 | 0.118 | 0.03 ± 0.02 | 0.052 | 0.116 | 0.01 ± 0.01 | 0.551 | 0.054 | 0.03 ± 0.01 | 0.029 | |
| M1 | MJ | 0.018 | 0.01 ± 0.02 | 0.600 | 0.157 | 0.03 ± 0.02 | 0.103 | 0.103 | 0.02 ± 0.02 | 0.194 |
| M2 | 0.030 | 0.01 ± 0.02 | 0.525 | 0.020 | 0.03 ± 0.02 | 0.123 | 0.010 | 0.02 ± 0.02 | 0.243 | |
| M1 | PLB | 0.002 | 0.00 ± 0.02 | 0.882 | 0.018 | 0.01 ± 0.02 | 0.604 | 0.067 | 0.02 ± 0.01 | 0.316 |
| M2 | 0.071 | 0.00 ± 0.02 | 0.878 | 0.075 | 0.01 ± 0.02 | 0.577 | 0.081 | 0.02 ± 0.02 | 0.298 | |
| Δ(Arachidonic acid/Eicosapentaenoic acid) | ||||||||||
| M1 | TS | 0.003 | 2.54 ± 6.19 | 0.684 | 0.003 | 2.38 ± 5.98 | 0.692 | 0.000 | 0.54 ± 6.18 | 0.931 |
| M2 | 0.046 | 4.47 ± 6.28 | 0.480 | 0.042 | 3.14 ± 5.99 | 0.602 | 0.042 | 1.93 ± 6.29 | 0.760 | |
| M1 | AJ | 0.015 | −6.89 ± 12.33 | 0.582 | 0.026 | −6.69 ± 9.09 | 0.470 | 0.043 | −10.12 ± 10.71 | 0.356 |
| M2 | 0.017 | −4.68 ± 13.47 | 0.732 | 0.014 | −5.53 ± 10.60 | 0.608 | 0.012 | −9.14 ± 12.19 | 0.463 | |
| M1 | MJ | 0.000 | −0.09 ± 13.91 | 0.995 | 0.060 | 14.12 ± 13.94 | 0.326 | 0.000 | 0.19 ± 14.11 | 0.989 |
| M2 | 0.144 | 2.71 ± 14.19 | 0.851 | 0.146 | 14.58 ± 13.70 | 0.305 | 0.148 | 4.76 ± 14.71 | 0.751 | |
| M1 | PLB | 0.045 | 7.06 ± 8.41 | 0.414 | 0.000 | 0.72 ± 8.60 | 0.934 | 0.027 | 5.44 ± 8.49 | 0.531 |
| M2 | 0.055 | 10.20 ± 9.45 | 0.300 | 0.026 | 3.17 ± 10.02 | 0.756 | 0.047 | 8.47 ± 9.63 | 0.395 | |
| Δ (Arachidonic acid/Docosahexaenoic acid) | ||||||||||
| M1 | TS | 0.001 | −0.09 ± 0.35 | 0.789 | 0.001 | 0.09 ± 0.34 | 0.792 | 0.015 | −0.31 ± 0.34 | 0.369 |
| M2 | 0.047 | −0.06 ± 0.35 | 0.872 | 0.049 | 0.14 ± 0.34 | 0.669 | 0.039 | −0.21 ± 0.35 | 0.551 | |
| M1 | AJ | 0.107 | −1.32 ± 0.85 | 0.138 | 0.008 | −0.27 ± 0.67 | 0.692 | 0.151 | −1.38 ± 0.73 | 0.073 |
| M2 | 0.017 | −1.16 ± 0.93 | 0.230 | 0.041 | −0.05 ± 0.77 | 0.954 | 0.018 | −1.34 ± 0.83 | 0.124 | |
| M1 | MJ | 0.001 | −0.08 ± 0.59 | 0.898 | 0.028 | 0.41 ± 0.60 | 0.508 | 0.000 | 0.05 ± 0.60 | 0.940 |
| M2 | 0.141 | −0.21 ± 0.60 | 0.729 | 0.135 | 0.41 ± 0.60 | 0.503 | 0.155 | 0.36 ± 0.62 | 0.569 | |
| M1 | PLB | 0.024 | 0.20 ± 0.32 | 0.550 | 0.004 | −0.08 ± 0.33 | 0.819 | 0.017 | −0.17 ± 0.33 | 0.616 |
| M2 | 0.174 | 0.11 ± 0.34 | 0.746 | 0.216 | −0.23 ± 0.34 | 0.508 | 0.226 | −0.31 ± 0.33 | 0.362 | |
| Δ BMI | Δ Body Weight | ||||||
|---|---|---|---|---|---|---|---|
| ΔR2 | B ± S.E. | p | ΔR2 | B ± S.E. | p | ||
| Δ Linoleic acid, C18:2n − 6 | |||||||
| M1 | TS | 0.001 | 0.01 ± 0.02 | 0.825 | 0.002 | 0.02 ± 0.07 | 0.729 |
| M2 | 0.040 | 0.00 ± 0.02 | 0.929 | 0.045 | 0.00 ± 0.07 | 0.985 | |
| M1 | AJ | 0.000 | 0.00 ± 0.04 | 0.998 | 0.002 | 0.02 ± 0.12 | 0.863 |
| M2 | 0.229 | −0.04 ± 0.04 | 0.291 | 0.232 | −0.12 ± 0.13 | 0.341 | |
| M1 | MJ | 0.000 | 0.00 ± 0.05 | 0.937 | 0.000 | 0.00 ± 0.15 | 0.994 |
| M2 | 0.150 | −0.02 ± 0.06 | 0.674 | 0.150 | −0.09 ± 0.16 | 0.587 | |
| M1 | PLB | 0.007 | 0.01 ± 0.04 | 0.755 | 0.007 | 0.04 ± 0.11 | 0.754 |
| M2 | 0.027 | 0.01 ± 0.04 | 0.888 | 0.026 | 0.02 ± 0.12 | 0.884 | |
| Δ Arachidonic acid, C20:4n − 6 | |||||||
| M1 | TS | 0.014 | −0.02 ± 0.03 | 0.387 | 0.007 | −0.05 ± 0.09 | 0.542 |
| M2 | 0.045 | −0.03 ± 0.03 | 0.323 | 0.050 | −0.07 ± 0.09 | 0.459 | |
| M1 | AJ | 0.042 | −0.07 ± 0.08 | 0.358 | 0.036 | −0.23 ± 0.26 | 0.397 |
| M2 | 0.177 | −0.07 ± 0.08 | 0.340 | 0.190 | −0.22 ± 0.25 | 0.390 | |
| M1 | MJ | 0.106 | −0.08 ± 0.06 | 0.187 | 0.091 | −0.20 ± 0.16 | 0.222 |
| M2 | 0.142 | −0.08 ± 0.05 | 0.174 | 0.138 | −0.21 ± 0.16 | 0.204 | |
| M1 | PLB | 0.002 | 0.01 ± 0.04 | 0.859 | 0.006 | 0.03 ± 0.11 | 0.771 |
| M2 | 0.046 | 0.02 ± 0.04 | 0.652 | 0.049 | 0.08 ± 0.13 | 0.574 | |
| Adrenic acid, C22:4n − 6 | |||||||
| M1 | TS | 0.008 | −0.18 ± 0.27 | 0.521 | 0.005 | −0.46 ± 0.85 | 0.597 |
| M2 | 0.035 | −0.10 ± 0.28 | 0.735 | 0.042 | −0.18 ± 0.87 | 0.834 | |
| M1 | AJ | 0.004 | 0.19 ± 0.65 | 0.775 | 0.003 | 0.52 ± 2.17 | 0.812 |
| M2 | 0.242 | 0.91 ± 0.71 | 0.218 | 0.245 | 2.73 ± 2.37 | 0.263 | |
| M1 | MJ | 0.129 | −0.56 ± 0.36 | 0.143 | 0.121 | −1.56 ± 1.05 | 0.156 |
| M2 | 0.070 | −0.41 ± 0.39 | 0.321 | 0.067 | −1.14 ± 1.15 | 0.340 | |
| M1 | PLB | 0.048 | 0.51 ± 0.58 | 0.397 | 0.044 | 1.46 ± 1.76 | 0.421 |
| M2 | 0.038 | 0.60 ± 0.69 | 0.398 | 0.034 | 1.70 ± 2.09 | 0.431 | |
| ΔEicosapentaenoic acid, C20:5n − 3 | |||||||
| M1 | TS | 0.147 | −0.47 ± 0.15 | 0.003 | 0.142 | −1.45 ± 0.48 | 0.004 |
| M2 | 0.023 | −0.45 ± 0.16 | 0.006 | 0.028 | −1.36 ± 0.49 | 0.007 | |
| M1 | AJ | 0.122 | −0.36 ± 0.22 | 0.112 | 0.103 | −1.11 ± 0.73 | 0.145 |
| M2 | 0.116 | −0.27 ± 0.23 | 0.252 | 0.137 | −0.81 ± 0.76 | 0.303 | |
| M1 | MJ | 0.264 | −0.73 ± 0.30 | 0.029 | 0.280 | −2.17 ± 0.87 | 0.024 |
| M2 | 0.051 | −0.63 ± 0.33 | 0.078 | 0.043 | −1.89 ± 0.95 | 0.066 | |
| M1 | PLB | 0.067 | −0.42 ± 0.41 | 0.316 | 0.069 | −1.30 ± 1.23 | 0.307 |
| M2 | 0.028 | −0.43 ± 0.45 | 0.358 | 0.029 | −1.34 ± 1.37 | 0.344 | |
| Δ Clupanodonic acid, C22:5n − 3 | |||||||
| M1 | TS | 0.004 | −0.08 ± 0.15 | 0.625 | 0.003 | −0.20 ± 0.48 | 0.682 |
| M2 | 0.043 | −0.10 ± 0.16 | 0.544 | 0.049 | −0.27 ± 0.49 | 0.583 | |
| M1 | AJ | 0.001 | −0.09 ± 0.82 | 0.912 | 0.000 | −0.24 ± 2.74 | 0.933 |
| M2 | 0.180 | 0.19 ± 0.80 | 0.813 | 0.195 | 0.73 ± 2.64 | 0.785 | |
| M1 | MJ | 0.032 | −0.13 ± 0.18 | 0.477 | 0.028 | −0.36 ± 0.53 | 0.506 |
| M2 | 0.147 | −0.15 ± 0.18 | 0.421 | 0.142 | −0.43 ± 0.54 | 0.433 | |
| M1 | PLB | 0.011 | 0.16 ± 0.40 | 0.686 | 0.007 | 0.40 ± 1.21 | 0.748 |
| M2 | 0.031 | 0.17 ± 0.45 | 0.715 | 0.030 | 0.38 ± 1.36 | 0.785 | |
| ΔDocosahexaenoic acid, C22:6n − 3 | |||||||
| M1 | TS | 0.008 | −0.04 ± 0.06 | 0.512 | 0.004 | −0.09 ± 0.18 | 0.624 |
| M2 | 0.035 | −0.02 ± 0.06 | 0.749 | 0.043 | −0.02 ± 0.18 | 0.901 | |
| M1 | AJ | 0.010 | 0.04 ± 0.08 | 0.664 | 0.013 | 0.14 ± 0.27 | 0.609 |
| M2 | 0.206 | 0.07 ± 0.08 | 0.365 | 0.226 | 0.27 ± 0.26 | 0.306 | |
| M1 | MJ | 0.133 | −0.17 ± 0.11 | 0.137 | 0.117 | −0.46 ± 0.32 | 0.165 |
| M2 | 0.064 | −0.13 ± 0.13 | 0.328 | 0.060 | −0.32 ± 0.37 | 0.397 | |
| M1 | PLB | 0.025 | −0.09 ± 0.15 | 0.541 | 0.034 | −0.33 ± 0.45 | 0.476 |
| M2 | 0.034 | −0.10 ± 0.16 | 0.549 | 0.033 | −0.34 ± 0.48 | 0.489 | |
| Δ Desaturase-5 predicted activity, AA/DGLA | |||||||
| M1 | TS | 0.003 | 0.02 ± 0.04 | 0.677 | 0.003 | 0.05 ± 0.12 | 0.697 |
| M2 | 0.038 | 0.00 ± 0.04 | 0.934 | 0.044 | 0.00 ± 0.12 | 0.974 | |
| M1 | AJ | 0.007 | −0.03 ± 0.08 | 0.704 | 0.007 | −0.10 ± 0.26 | 0.705 |
| M2 | 0.228 | −0.09 ± 0.08 | 0.261 | 0.245 | −0.30 ± 0.25 | 0.246 | |
| M1 | MJ | 0.134 | 0.14 ± 0.09 | 0.135 | 0.142 | 0.41 ± 0.25 | 0.123 |
| M2 | 0.142 | 0.14 ± 0.09 | 0.125 | 0.129 | 0.41 ± 0.25 | 0.123 | |
| M1 | PLB | 0.003 | −0.01 ± 0.05 | 0.844 | 0.004 | −0.04 ± 0.15 | 0.813 |
| M2 | 0.030 | 0.00 ± 0.06 | 0.945 | 0.027 | 0.00 ± 0.17 | 0.981 | |
| Δ Desaturase-6 predicted activity, DGLA/LA | |||||||
| M1 | TS | 0.012 | −1.15 ± 1.40 | 0.418 | 0.008 | −2.87 ± 4.40 | 0.516 |
| M2 | 0.032 | −0.66 ± 1.45 | 0.649 | 0.040 | −1.19 ± 4.53 | 0.794 | |
| M1 | AJ | 0.000 | 0.06 ± 2.44 | 0.981 | 0.000 | −0.05 ± 8.12 | 0.995 |
| M2 | 0.223 | 2.58 ± 2.53 | 0.321 | 0.237 | 8.51 ± 8.34 | 0.321 | |
| M1 | MJ | 0.224 | −4.99 ± 2.32 | 0.047 | 0.216 | −14.19 ± 6.76 | 0.052 |
| M2 | 0.134 | −4.98 ± 2.27 | 0.046 | 0.122 | −13.99 ± 6.71 | 0.056 | |
| M1 | PLB | 0.027 | 1.59 ± 2.49 | 0.532 | 0.041 | 6.02 ± 7.48 | 0.434 |
| M2 | 0.020 | 1.22 ± 2.75 | 0.664 | 0.016 | 5.00 ± 8.26 | 0.556 | |
| Δ(Arachidonic acid/ Eicosapentaenoic acid) | |||||||
| M1 | TS | 0.042 | 0.00 ± 0.00 | 0.127 | 0.038 | 0.01 ± 0.01 | 0.146 |
| M2 | 0.035 | 0.00 ± 0.00 | 0.154 | 0.042 | 0.01 ± 0.01 | 0.172 | |
| M1 | AJ | 0.030 | 0.00 ± 0.00 | 0.441 | 0.026 | 0.01 ± 0.01 | 0.470 |
| M2 | 0.161 | 0.00 ± 0.00 | 0.588 | 0.176 | 0.01 ± 0.01 | 0.638 | |
| M1 | MJ | 0.057 | 0.00 ± 0.00 | 0.339 | 0.056 | 0.01 ± 0.01 | 0.346 |
| M2 | 0.095 | 0.00 ± 0.00 | 0.641 | 0.091 | 0.01 ± 0.01 | 0.621 | |
| M1 | PLB | 0.061 | 0.00 ± 0.00 | 0.339 | 0.056 | 0.01 ± 0.01 | 0.361 |
| M2 | 0.030 | 0.00 ± 0.00 | 0.374 | 0.030 | 0.01 ± 0.01 | 0.395 | |
| Δ (Arachidonic acid/ Docosahexaenoic acid) | |||||||
| M1 | TS | 0.007 | −0.02 ± 0.04 | 0.537 | 0.007 | −0.07 ± 0.11 | 0.525 |
| M2 | 0.051 | −0.04 ± 0.04 | 0.334 | 0.058 | −0.11 ± 0.11 | 0.313 | |
| M1 | AJ | 0.037 | −0.04 ± 0.05 | 0.390 | 0.041 | −0.15 ± 0.16 | 0.365 |
| M2 | 0.220 | −0.06 ± 0.05 | 0.183 | 0.241 | −0.22 ± 0.15 | 0.152 | |
| M1 | MJ | 0.000 | −0.01 ± 0.08 | 0.937 | 0.001 | −0.03 ± 0.22 | 0.912 |
| M2 | 0.154 | −0.04 ± 0.08 | 0.614 | 0.151 | −0.14 ± 0.24 | 0.568 | |
| M1 | PLB | 0.013 | 0.05 ± 0.11 | 0.667 | 0.025 | 0.21 ± 0.34 | 0.547 |
| M2 | 0.063 | 0.10 ± 0.13 | 0.450 | 0.071 | 0.37 ± 0.39 | 0.353 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zec, M.M.; Krga, I.; Stojković, L.; Živković, M.; Pokimica, B.; Stanković, A.; Glibetic, M. Is There a FADS2-Modulated Link between Long-Chain Polyunsaturated Fatty Acids in Plasma Phospholipids and Polyphenol Intake in Adult Subjects Who Are Overweight? Nutrients 2021, 13, 296. https://doi.org/10.3390/nu13020296
Zec MM, Krga I, Stojković L, Živković M, Pokimica B, Stanković A, Glibetic M. Is There a FADS2-Modulated Link between Long-Chain Polyunsaturated Fatty Acids in Plasma Phospholipids and Polyphenol Intake in Adult Subjects Who Are Overweight? Nutrients. 2021; 13(2):296. https://doi.org/10.3390/nu13020296
Chicago/Turabian StyleZec, Manja M., Irena Krga, Ljiljana Stojković, Maja Živković, Biljana Pokimica, Aleksandra Stanković, and Maria Glibetic. 2021. "Is There a FADS2-Modulated Link between Long-Chain Polyunsaturated Fatty Acids in Plasma Phospholipids and Polyphenol Intake in Adult Subjects Who Are Overweight?" Nutrients 13, no. 2: 296. https://doi.org/10.3390/nu13020296
APA StyleZec, M. M., Krga, I., Stojković, L., Živković, M., Pokimica, B., Stanković, A., & Glibetic, M. (2021). Is There a FADS2-Modulated Link between Long-Chain Polyunsaturated Fatty Acids in Plasma Phospholipids and Polyphenol Intake in Adult Subjects Who Are Overweight? Nutrients, 13(2), 296. https://doi.org/10.3390/nu13020296