

Eddy-Induced Chlorophyll Profile Characteristics and Underlying Dynamic Mechanisms in the South Pacific Ocean

Abstract
1. Introduction
2. Materials and Methods
2.1. Eddy Datasets
2.2. Multi-Satellite Merged Ocean Color Products
2.3. Argo and BGC-Argo Datasets
2.4. Data Processing
3. Results
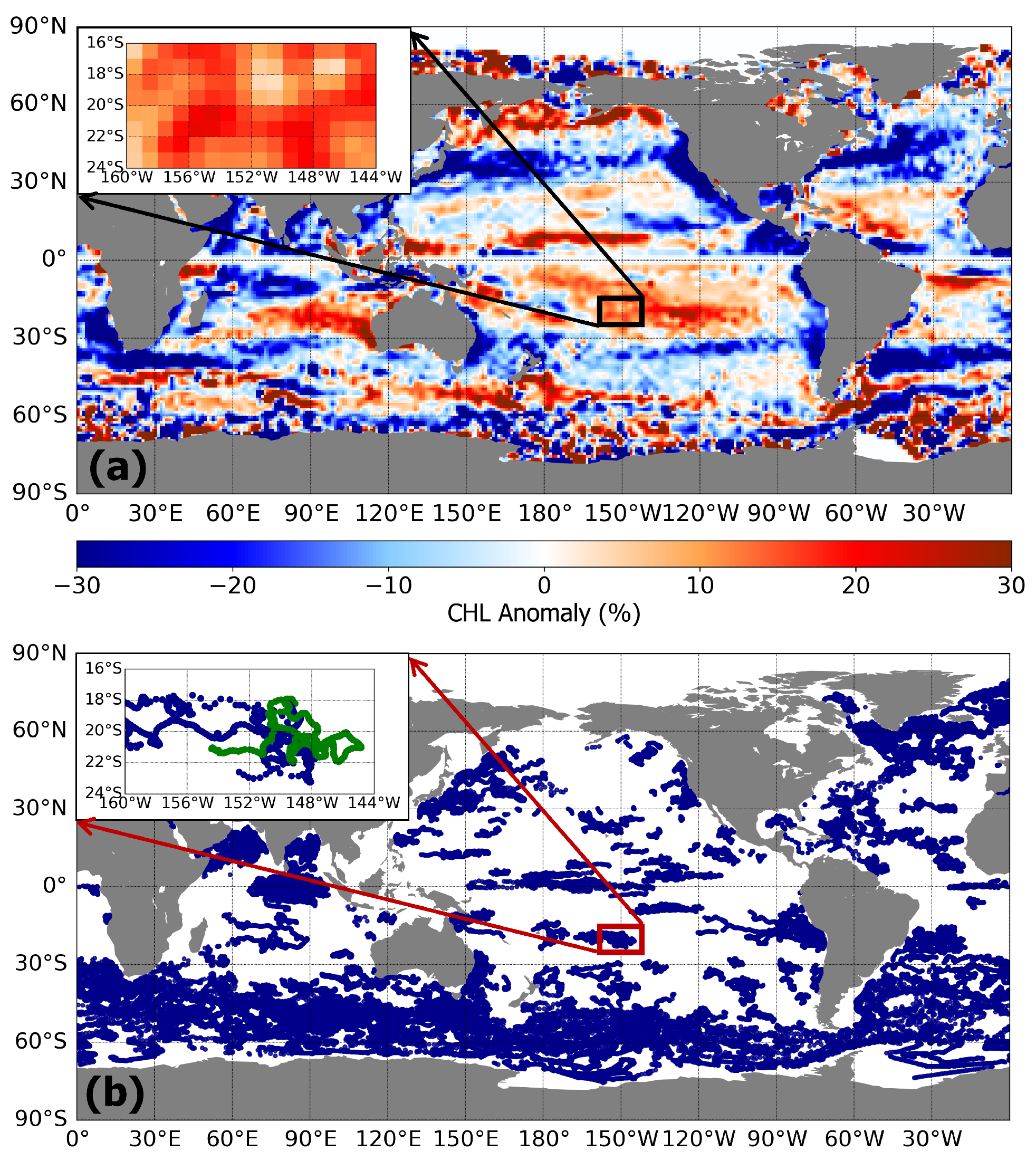
3.1. Chlorophyll Characteristics of Sea Surface
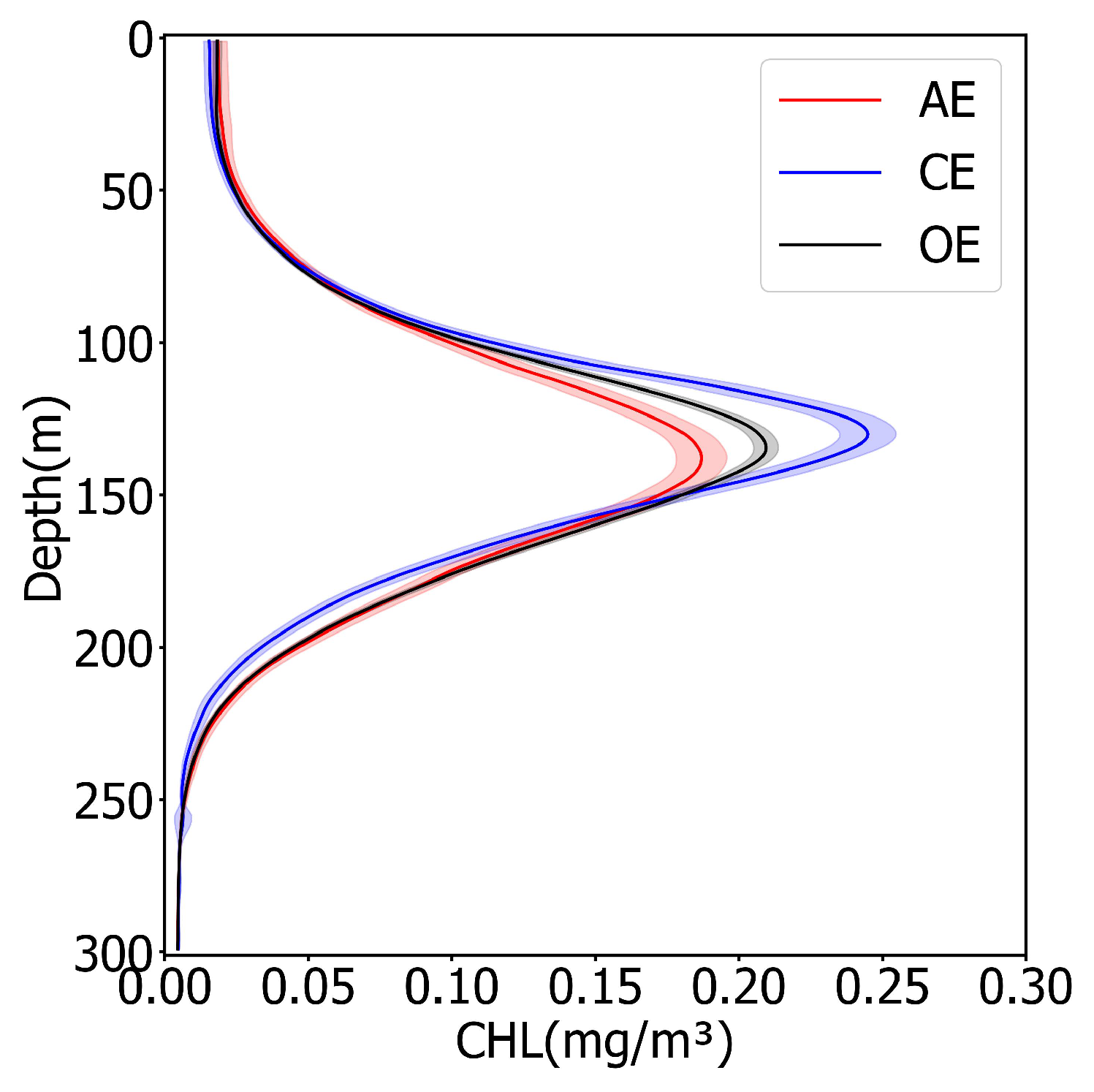
3.2. Subsurface Chlorophyll Structure in Eddies
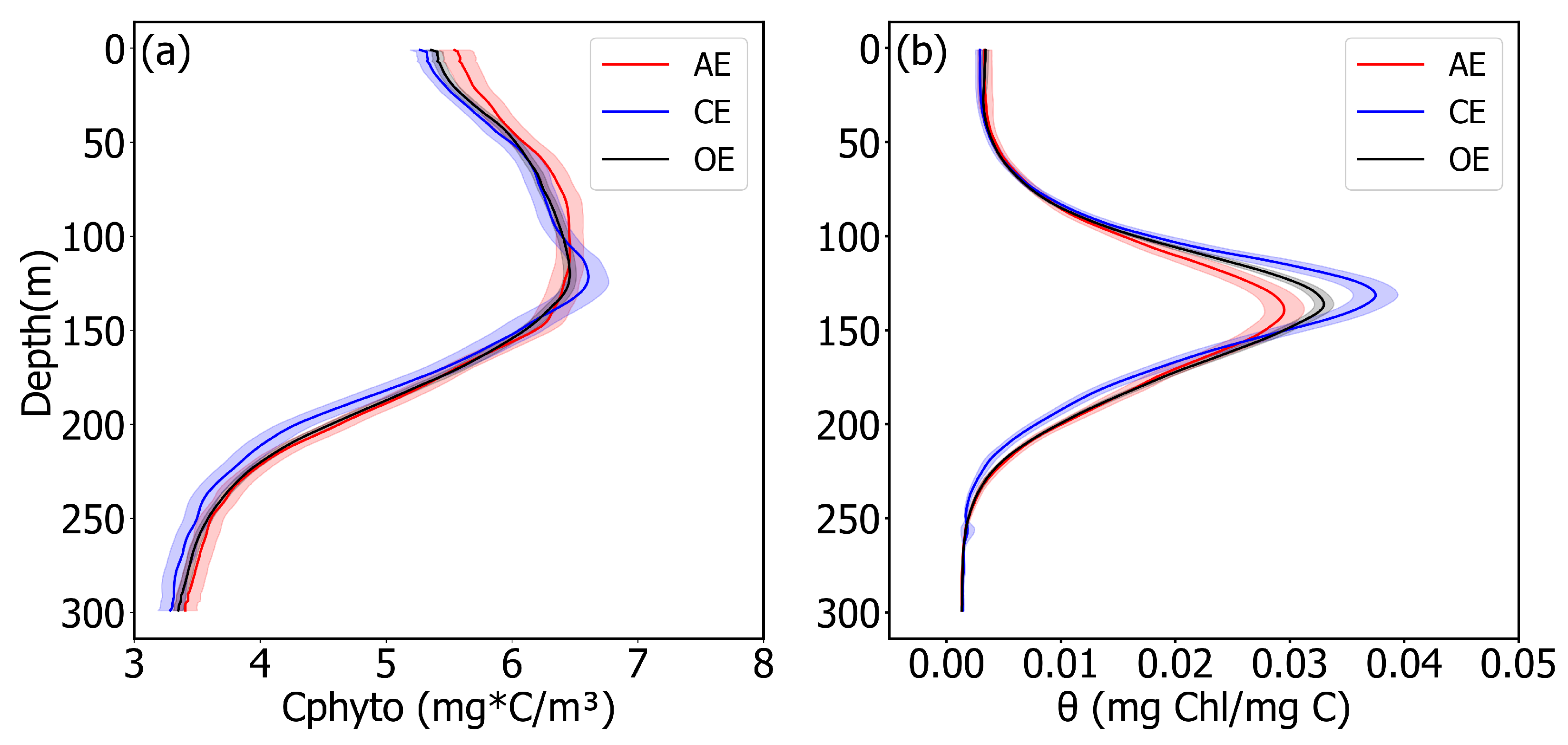
3.3. CPhyto and
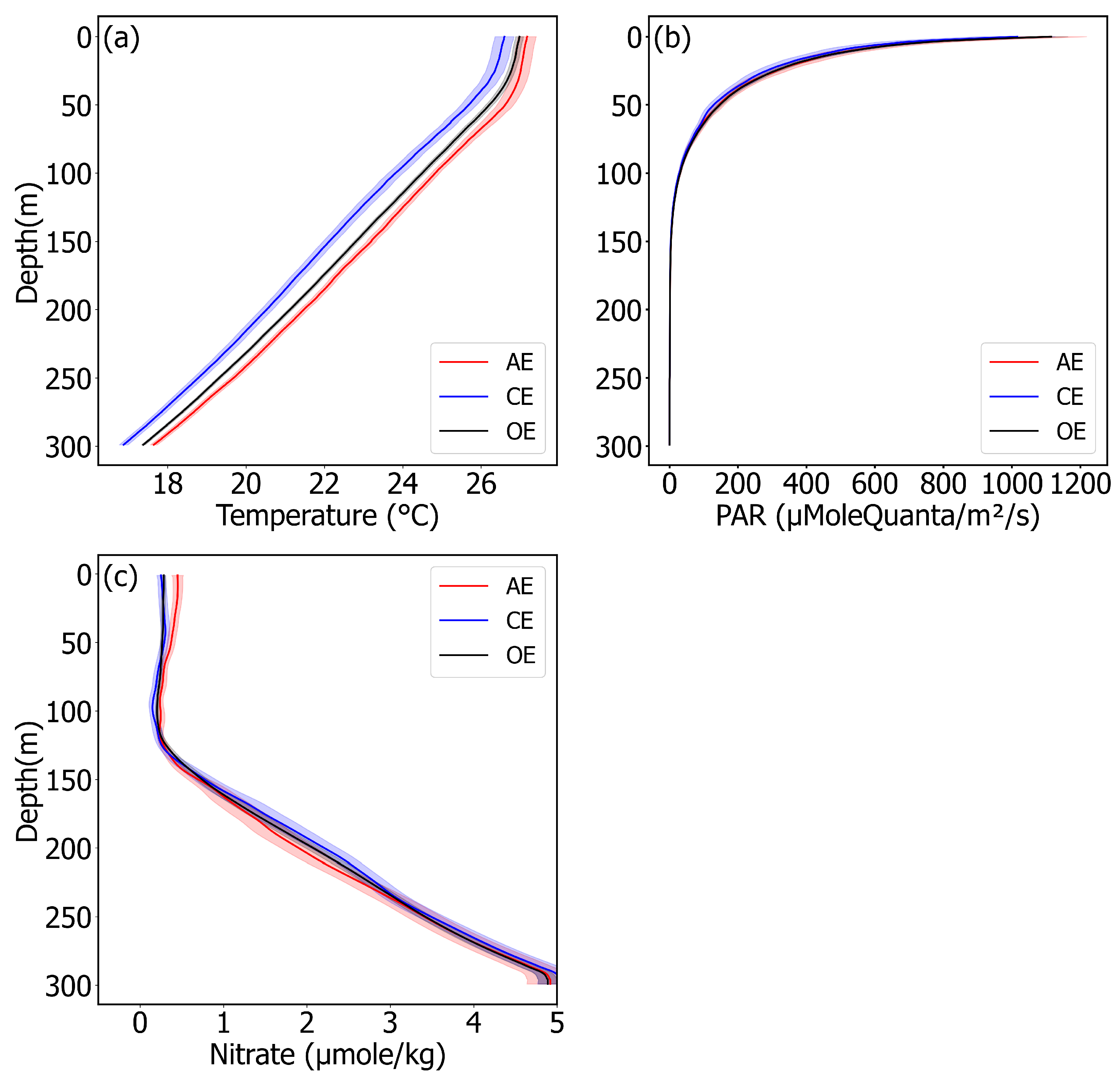
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef]
- Chassot, E.; Bonhommeau, S.; Dulvy, N.K.; Mélin, F.; Watson, R.; Gascuel, D.; Le Pape, O. Global marine primary production constrains fisheries catches. Ecol. Lett. 2010, 13, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Durant, J.M.; Stige, L.C.; Hessen, D.O.; Hjermann, D.Ø.; Zhu, L.; Llope, M.; Stenseth, N.C. Contrasting correlation patterns between environmental factors and chlorophyll levels in the global ocean. Glob. Biogeochem. Cycles 2015, 29, 2095–2107. [Google Scholar] [CrossRef]
- Zhao, D.; Gao, L.; Xu, Y. Quantification of the impact of environmental factors on chlorophyll in the open ocean. J. Oceanol. Limnol. 2021, 39, 447–457. [Google Scholar] [CrossRef]
- Halsey, K.H.; Jones, B.M. Phytoplankton strategies for photosynthetic energy allocation. Annu. Rev. Mar. Sci. 2015, 7, 265–297. [Google Scholar] [CrossRef] [PubMed]
- Behrenfeld, M.J.; O’Malley, R.T.; Boss, E.S.; Westberry, T.K.; Graff, J.R.; Halsey, K.H.; Milligan, A.J.; Siegel, D.A.; Brown, M.B. Revaluating ocean warming impacts on global phytoplankton. Nat. Clim. Chang. 2016, 6, 323–330. [Google Scholar] [CrossRef]
- Chelton, D.B.; Schlax, M.G.; Samelson, R.M. Global observations of nonlinear mesoscale eddies. Prog. Oceanogr. 2011, 91, 167–216. [Google Scholar] [CrossRef]
- He, Q.; Zhan, H.; Cai, S.; He, Y.; Huang, G.; Zhan, W. A new assessment of mesoscale eddies in the South China Sea: Surface features, three-dimensional structures, and thermohaline transports. J. Geophys. Res. Ocean. 2018, 123, 4906–4929. [Google Scholar] [CrossRef]
- Chen, G.; Han, G. Contrasting short-lived with long-lived mesoscale eddies in the global ocean. J. Geophys. Res. Ocean. 2019, 124, 3149–3167. [Google Scholar] [CrossRef]
- Chelton, D.B.; Gaube, P.; Schlax, M.G.; Early, J.J.; Samelson, R.M. The influence of nonlinear mesoscale eddies on near-surface oceanic chlorophyll. Science 2011, 334, 328–332. [Google Scholar] [CrossRef]
- Batten, S.D.; Crawford, W.R. The influence of coastal origin eddies on oceanic plankton distributions in the eastern Gulf of Alaska. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 991–1009. [Google Scholar] [CrossRef]
- Conway, T.M.; Palter, J.B.; de Souza, G.F. Gulf Stream rings as a source of iron to the North Atlantic subtropical gyre. Nat. Geosci. 2018, 11, 594–598. [Google Scholar] [CrossRef]
- Xu, G.; Dong, C.; Liu, Y.; Gaube, P.; Yang, J. Chlorophyll rings around ocean eddies in the North Pacific. Sci. Rep. 2019, 9, 2056. [Google Scholar] [CrossRef]
- Liu, Y.; Li, X.; Ren, Y. A deep learning model for oceanic mesoscale eddy detection based on multi-source remote sensing imagery. In Proceedings of the IGARSS 2020—2020 IEEE International Geoscience and Remote Sensing Symposium, Virtual, 26 September–2 October 2020; IEEE: Piscataway, NJ, USA, 2020; pp. 6762–6765. [Google Scholar]
- Xing, Q.; Yu, H.; Wang, H.; Ito, S.I.; Chai, F. Mesoscale eddies modulate the dynamics of human fishing activities in the global midlatitude ocean. Fish Fish. 2023, 24, 527–543. [Google Scholar] [CrossRef]
- Dawson, H.R.; Strutton, P.G.; Gaube, P. The unusual surface chlorophyll signatures of Southern Ocean eddies. J. Geophys. Res. Ocean. 2018, 123, 6053–6069. [Google Scholar] [CrossRef]
- McGillicuddy, D.J., Jr. Mechanisms of physical-biological-biogeochemical interaction at the oceanic mesoscale. Annu. Rev. Mar. Sci. 2016, 8, 125–159. [Google Scholar] [CrossRef]
- Gaube, P.; Chelton, D.B.; Strutton, P.G.; Behrenfeld, M.J. Satellite observations of chlorophyll, phytoplankton biomass, and Ekman pumping in nonlinear mesoscale eddies. J. Geophys. Res. Ocean. 2013, 118, 6349–6370. [Google Scholar] [CrossRef]
- Dufois, F.; Hardman-Mountford, N.J.; Greenwood, J.; Richardson, A.J.; Feng, M.; Matear, R.J. Anticyclonic eddies are more productive than cyclonic eddies in subtropical gyres because of winter mixing. Sci. Adv. 2016, 2, e1600282. [Google Scholar] [CrossRef]
- Arteaga, L.; Pahlow, M.; Oschlies, A. Modeled Chl: C ratio and derived estimates of phytoplankton carbon biomass and its contribution to total particulate organic carbon in the global surface ocean. Glob. Biogeochem. Cycles 2016, 30, 1791–1810. [Google Scholar] [CrossRef]
- He, Q.; Zhan, H.; Cai, S.; Zhan, W. Eddy-induced near-surface chlorophyll anomalies in the subtropical gyres: Biomass or physiology? Geophys. Res. Lett. 2021, 48, e2020GL091975. [Google Scholar] [CrossRef]
- Gordon, H.R.; McCluney, W. Estimation of the depth of sunlight penetration in the sea for remote sensing. Appl. Opt. 1975, 14, 413–416. [Google Scholar] [CrossRef]
- Cornec, M.; Claustre, H.; Mignot, A.; Guidi, L.; Lacour, L.; Poteau, A.; d’Ortenzio, F.; Gentili, B.; Schmechtig, C. Deep chlorophyll maxima in the global ocean: Occurrences, drivers and characteristics. Glob. Biogeochem. Cycles 2021, 35, e2020GB006759. [Google Scholar] [CrossRef] [PubMed]
- Letelier, R.M.; Karl, D.M.; Abbott, M.R.; Flament, P.; Freilich, M.; Lukas, R.; Strub, T. Role of late winter mesoscale events in the biogeochemical variability of the upper water column of the North Pacific Subtropical Gyre. J. Geophys. Res. Ocean. 2000, 105, 28723–28739. [Google Scholar] [CrossRef]
- Cullen, J.J. Subsurface chlorophyll maximum layers: Enduring enigma or mystery solved? Annu. Rev. Mar. Sci. 2015, 7, 207–239. [Google Scholar] [CrossRef] [PubMed]
- Cornec, M.; Laxenaire, R.; Speich, S.; Claustre, H. Impact of mesoscale eddies on deep chlorophyll maxima. Geophys. Res. Lett. 2021, 48, e2021GL093470. [Google Scholar] [CrossRef] [PubMed]
- Schlax, M.G.; Chelton, D.B. The “Growing Method” of Eddy Identification and Tracking in Two and Three Dimensions; College of Earth, Ocean and Atmospheric Sciences, Oregon State University: Corvallis, OR, USA, 2016; Volume 8, p. 23. [Google Scholar]
- Peng, L.; Chen, G.; Guan, L.; Tian, F. Contrasting westward and eastward propagating mesoscale eddies in the global ocean. IEEE Trans. Geosci. Remote Sens. 2021, 60, 4504710. [Google Scholar] [CrossRef]
- Wong, A.P.; Wijffels, S.E.; Riser, S.C.; Pouliquen, S.; Hosoda, S.; Roemmich, D.; Gilson, J.; Johnson, G.C.; Martini, K.; Murphy, D.J.; et al. Argo data 1999–2019: Two million temperature-salinity profiles and subsurface velocity observations from a global array of profiling floats. Front. Mar. Sci. 2020, 7, 700. [Google Scholar] [CrossRef]
- Johnson, K.; Claustre, H. The scientific rationale, design and Implementation Plan for a Biogeochemical-Argo float array. Biogeochem.-Argo Plan. Group 2016, 58, 46601. [Google Scholar]
- Bittig, H.C.; Maurer, T.L.; Plant, J.N.; Schmechtig, C.; Wong, A.P.; Claustre, H.; Trull, T.W.; Udaya Bhaskar, T.; Boss, E.; Dall’Olmo, G.; et al. A BGC-Argo guide: Planning, deployment, data handling and usage. Front. Mar. Sci. 2019, 6, 502. [Google Scholar] [CrossRef]
- Sukigara, C.; Inoue, R.; Sato, K.; Mino, Y.; Nagai, T.; Fassbender, A.J.; Takeshita, Y.; Bishop, S.; Oka, E. Observing intermittent biological productivity and vertical carbon transports during the spring transition with BGC Argo floats in the western North Pacific. Biogeosci. Discuss. 2022, 2022, 1–52. [Google Scholar]
- Xing, X.; Claustre, H.; Uitz, J.; Mignot, A.; Poteau, A.; Wang, H. Seasonal variations of bio-optical properties and their interrelationships observed by B io-A rgo floats in the subpolar N orth A tlantic. J. Geophys. Res. Ocean. 2014, 119, 7372–7388. [Google Scholar] [CrossRef]
- Chai, F.; Johnson, K.S.; Claustre, H.; Xing, X.; Wang, Y.; Boss, E.; Riser, S.; Fennel, K.; Schofield, O.; Sutton, A. Monitoring ocean biogeochemistry with autonomous platforms. Nat. Rev. Earth Environ. 2020, 1, 315–326. [Google Scholar] [CrossRef]
- de Boyer Montégut, C.; Madec, G.; Fischer, A.S.; Lazar, A.; Iudicone, D. Mixed layer depth over the global ocean: An examination of profile data and a profile-based climatology. J. Geophys. Res. Ocean. 2004, 109. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, J.; Chen, G. Euphotic Zone Depth Anomaly in Global Mesoscale Eddies by Multi-Mission Fusion Data. Remote Sens. 2023, 15, 1062. [Google Scholar] [CrossRef]
- Oubelkheir, K. Biogeochemical Characterization of Various Oceanic Provinces through Optical Indicators over Various Space and Time Scales. Ph.D. Thesis, Université de la Méditerraneé/CNRS, Marseille, France, 2001; p. 216. [Google Scholar]
- Behrenfeld, M.J.; Boss, E.; Siegel, D.A.; Shea, D.M. Carbon-based ocean productivity and phytoplankton physiology from space. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Stramski, D.; Reynolds, R.A.; Babin, M.; Kaczmarek, S.; Lewis, M.R.; Röttgers, R.; Sciandra, A.; Stramska, M.; Twardowski, M.; Franz, B.; et al. Relationships between the surface concentration of particulate organic carbon and optical properties in the eastern South Pacific and eastern Atlantic Oceans. Biogeosciences 2008, 5, 171–201. [Google Scholar] [CrossRef]
- Xing, X.; Wells, M.L.; Chen, S.; Lin, S.; Chai, F. Enhanced winter carbon export observed by BGC-Argo in the Northwest Pacific Ocean. Geophys. Res. Lett. 2020, 47, e2020GL089847. [Google Scholar] [CrossRef]
- Kerimoglu, O.; Hofmeister, R.; Maerz, J.; Riethmüller, R.; Wirtz, K.W. The acclimative biogeochemical model of the southern North Sea. Biogeosciences 2017, 14, 4499–4531. [Google Scholar] [CrossRef]
- Martin, A.; Pondaven, P. On estimates for the vertical nitrate flux due to eddy pumping. J. Geophys. Res. Ocean. 2003, 108. [Google Scholar] [CrossRef]
- Gregg, W.W.; Casey, N.W.; McClain, C.R. Recent trends in global ocean chlorophyll. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Poppeschi, C.; Charria, G.; Daniel, A.; Verney, R.; Rimmelin-Maury, P.; Retho, M.; Goberville, E.; Grossteffan, E.; Plus, M. Interannual variability of the initiation of the phytoplankton growing period in two French coastal ecosystems. Biogeosciences Discuss. 2022, 2022, 5667–5687. [Google Scholar] [CrossRef]
- Wu, M.L.; Wang, Y.S.; Wang, Y.T.; Yin, J.P.; Dong, J.D.; Jiang, Z.Y.; Sun, F.L. Scenarios of nutrient alterations and responses of phytoplankton in a changing Daya Bay, South China Sea. J. Mar. Syst. 2017, 165, 1–12. [Google Scholar] [CrossRef]
- He, Q.; Zhan, H.; Shuai, Y.; Cai, S.; Li, Q.P.; Huang, G.; Li, J. Phytoplankton bloom triggered by an anticyclonic eddy: The combined effect of eddy-E kman pumping and winter mixing. J. Geophys. Res. Ocean. 2017, 122, 4886–4901. [Google Scholar] [CrossRef]
- Wang, T.; Ning, J.; Xu, Q. The Enhancement of Upper Ocean Nutrients Concentration in the Peripheries of Two Anti-Cyclonic Eddies. In Proceedings of the IGARSS 2018—2018 IEEE International Geoscience and Remote Sensing Symposium, Valencia, Spain, 22–27 July 2018; IEEE: Piscataway, NJ, USA, 2018; pp. 7624–7627. [Google Scholar]
- Gaube, P.; McGillicuddy, D.J., Jr.; Moulin, A.J. Mesoscale eddies modulate mixed layer depth globally. Geophys. Res. Lett. 2019, 46, 1505–1512. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Boss, E.S. Resurrecting the ecological underpinnings of ocean plankton blooms. Annu. Rev. Mar. Sci. 2014, 6, 167–194. [Google Scholar] [CrossRef] [PubMed]
- Geider, R.J. Light and temperature dependence of the carbon to chlorophyll a ratio in microalgae and cyanobacteria: Implications for physiology and growth of phytoplankton. New Phytol. 1987, 106, 1–34. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Halsey, K.H.; Milligan, A.J. Evolved physiological responses of phytoplankton to their integrated growth environment. Philos. Trans. R. Soc. Biol. Sci. 2008, 363, 2687–2703. [Google Scholar] [CrossRef] [PubMed]
- d’Ortenzio, F.; Taillandier, V.; Claustre, H.; Coppola, L.; Conan, P.; Dumas, F.; Durrieu du Madron, X.; Fourrier, M.; Gogou, A.; Karageorgis, A.; et al. BGC-Argo floats observe nitrate injection and spring phytoplankton increase in the surface layer of Levantine Sea (Eastern Mediterranean). Geophys. Res. Lett. 2021, 48, e2020GL091649. [Google Scholar] [CrossRef]
- Arostegui, M.C.; Gaube, P.; Woodworth-Jefcoats, P.A.; Kobayashi, D.R.; Braun, C.D. Anticyclonic eddies aggregate pelagic predators in a subtropical gyre. Nature 2022, 609, 535–540. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AE | CE | OE | |
|---|---|---|---|
| MLD (m) | 51 ± 0.73 | 40 ± 0.65 | 44 ± 0.59 |
| ZEU (m) | 126.4 ± 0.27 | 125.1 ± 0.25 | 126.6 ± 0.36 |
| SCMD (m) | 139 ± 0.76 | 131 ± 0.67 | 135 ± 0.49 |
| Depth | Chl′ | CPhyto′ | θ′ | Temp′ | PAR′ | Nitrate′ |
|---|---|---|---|---|---|---|
| 0-MLD | 21% ± 2.9% | 5.1% ± 1.1% | 15.6% ± 1.8% | 2.2% ± 0.2% | −6.3% ± 0.1% | 65% ± 19.1% |
| 50–110 m | −7.9% ± 0.5% | 1.7% ± 0.4% | −9.2% ± 1.2% | 3.9% ± 0.3% | 6.1% ± 0.7% | 35.8% ± 16.5% |
| 110–150 m | −25% ± 1.3% | −1.2% ± 0.3% | −23.6% ± 2.5% | 4.5% ± 0.1% | 9.9% ± 3.2% | −10% ± 3.6% |
| 150–300 m | 12.6% ± 1.7% | 4.1% ± 0.9% | 4.5% ± 0.2% | 5.8% ± 1.5% | −5.4% ± 0.3% | |
| SCM | −27.7% ± 3.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, M.; Yang, J.; Chen, G. Eddy-Induced Chlorophyll Profile Characteristics and Underlying Dynamic Mechanisms in the South Pacific Ocean. Remote Sens. 2024, 16, 2628. https://doi.org/10.3390/rs16142628
Hou M, Yang J, Chen G. Eddy-Induced Chlorophyll Profile Characteristics and Underlying Dynamic Mechanisms in the South Pacific Ocean. Remote Sensing. 2024; 16(14):2628. https://doi.org/10.3390/rs16142628
Chicago/Turabian StyleHou, Meng, Jie Yang, and Ge Chen. 2024. "Eddy-Induced Chlorophyll Profile Characteristics and Underlying Dynamic Mechanisms in the South Pacific Ocean" Remote Sensing 16, no. 14: 2628. https://doi.org/10.3390/rs16142628
APA StyleHou, M., Yang, J., & Chen, G. (2024). Eddy-Induced Chlorophyll Profile Characteristics and Underlying Dynamic Mechanisms in the South Pacific Ocean. Remote Sensing, 16(14), 2628. https://doi.org/10.3390/rs16142628