


The Spatio-Temporal Distribution of the Freshwater Bivalves Corbicula fluminea and Dreissena polymorpha in the Lower Sector of the Danube River and the Danube Delta

 ,
,  , and
, and
Abstract
1. Introduction
Worldwide Historical Records of Corbicula fluminea (Müller, 1774) and Dreissena polymorpha (Pallas, 1771)
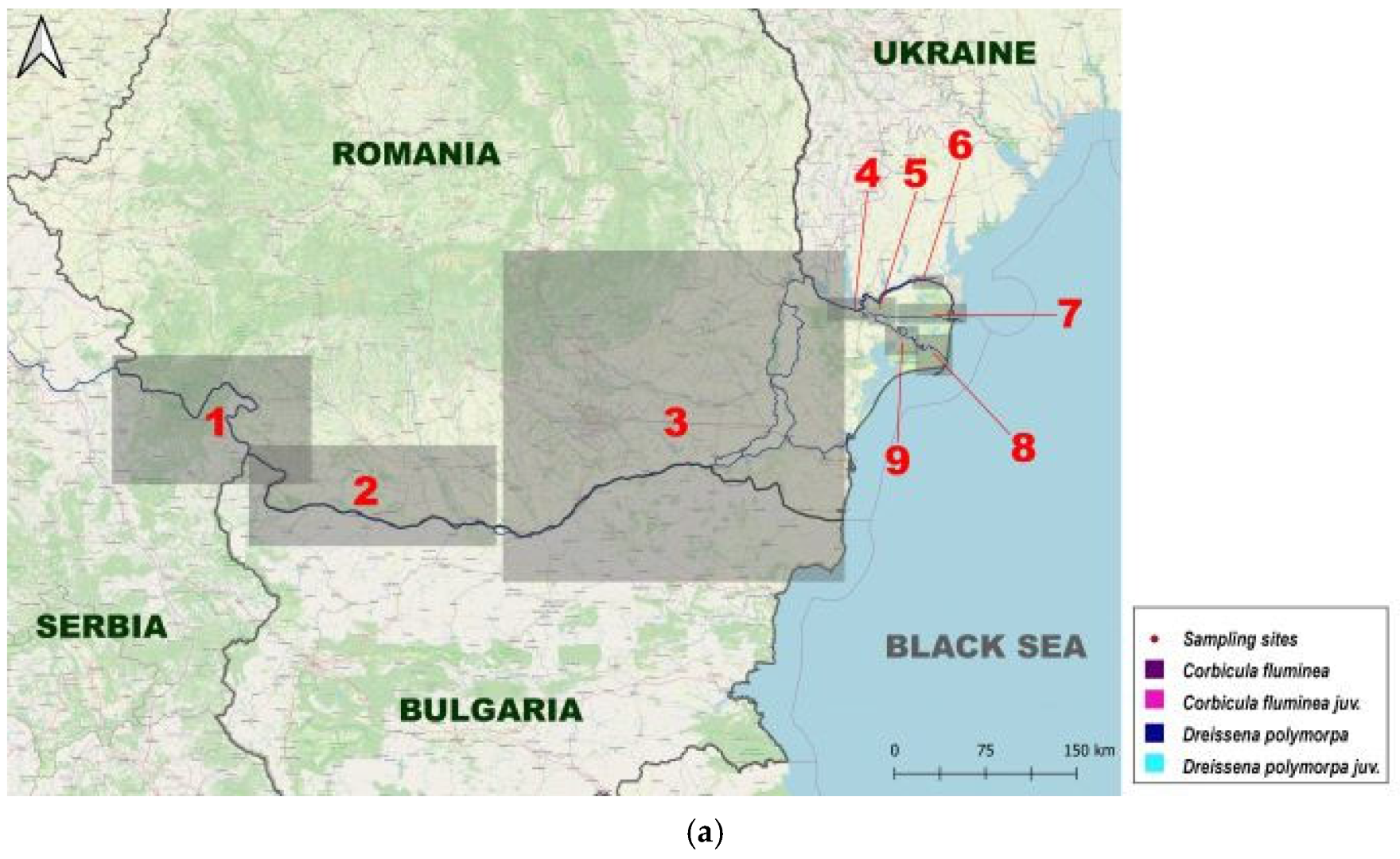
2. Materials and Methods
2.1. Sample Processing and Analysis
2.2. Statistical Analysis
2.3. Studied Species
2.3.1. Corbicula fluminea
2.3.2. Dreissena polymorpha
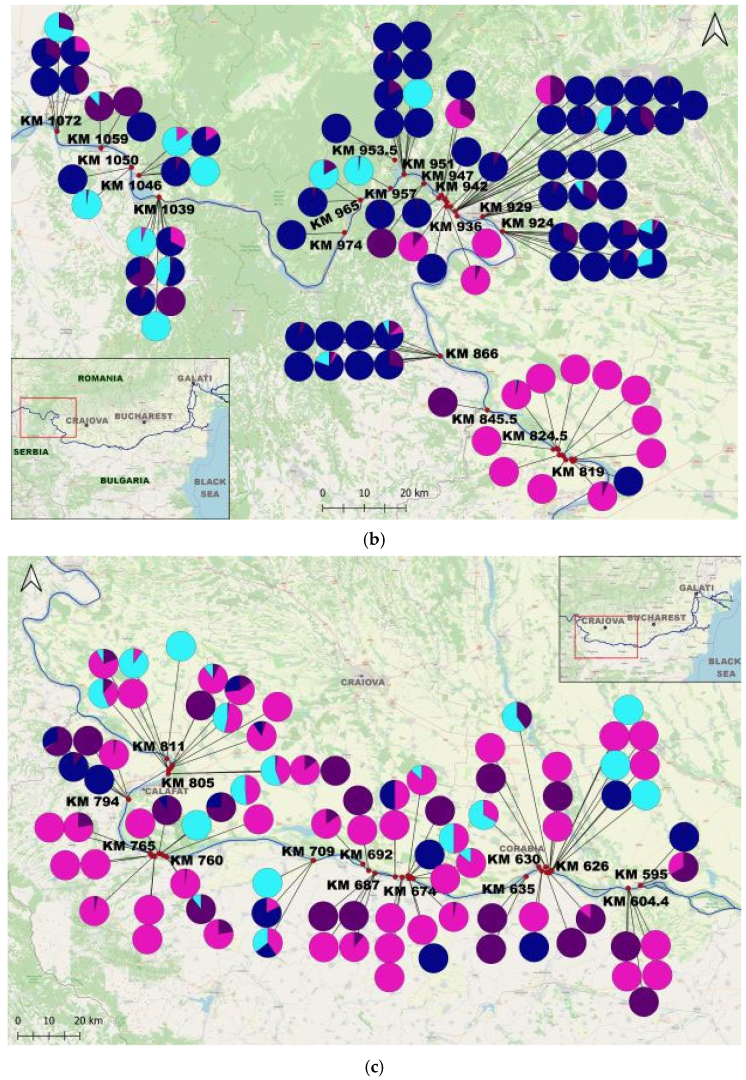
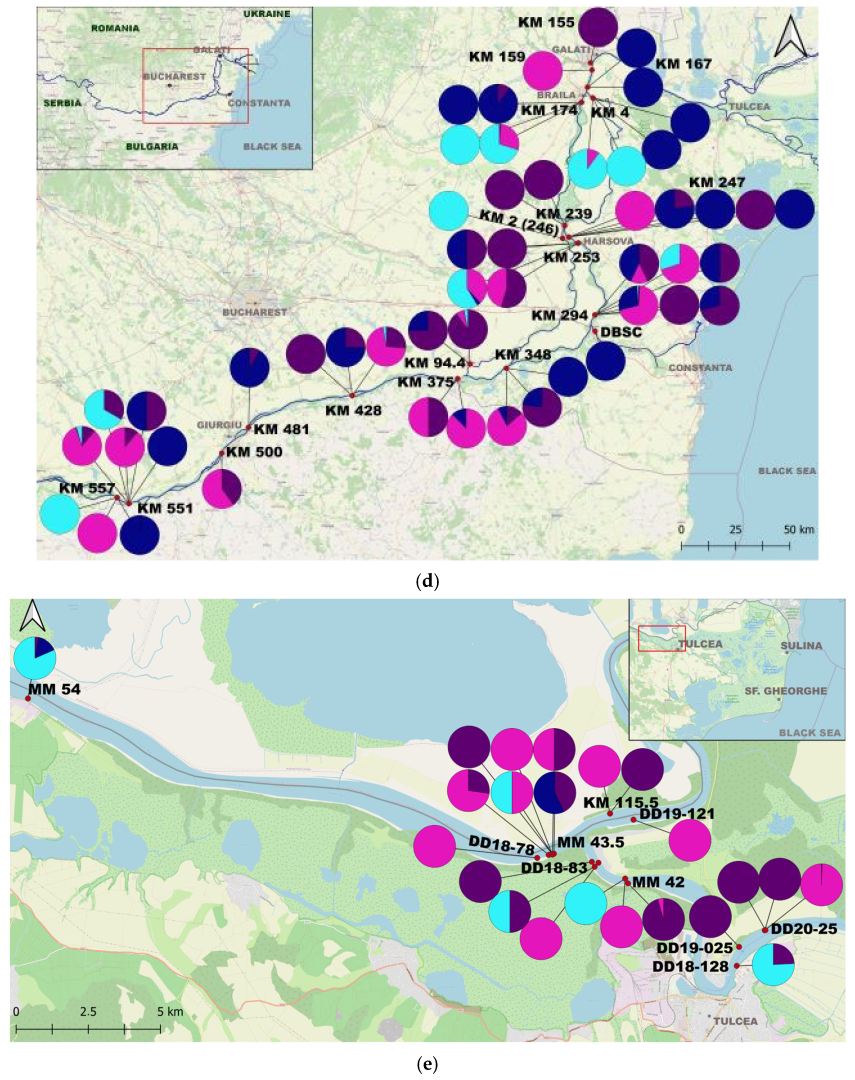
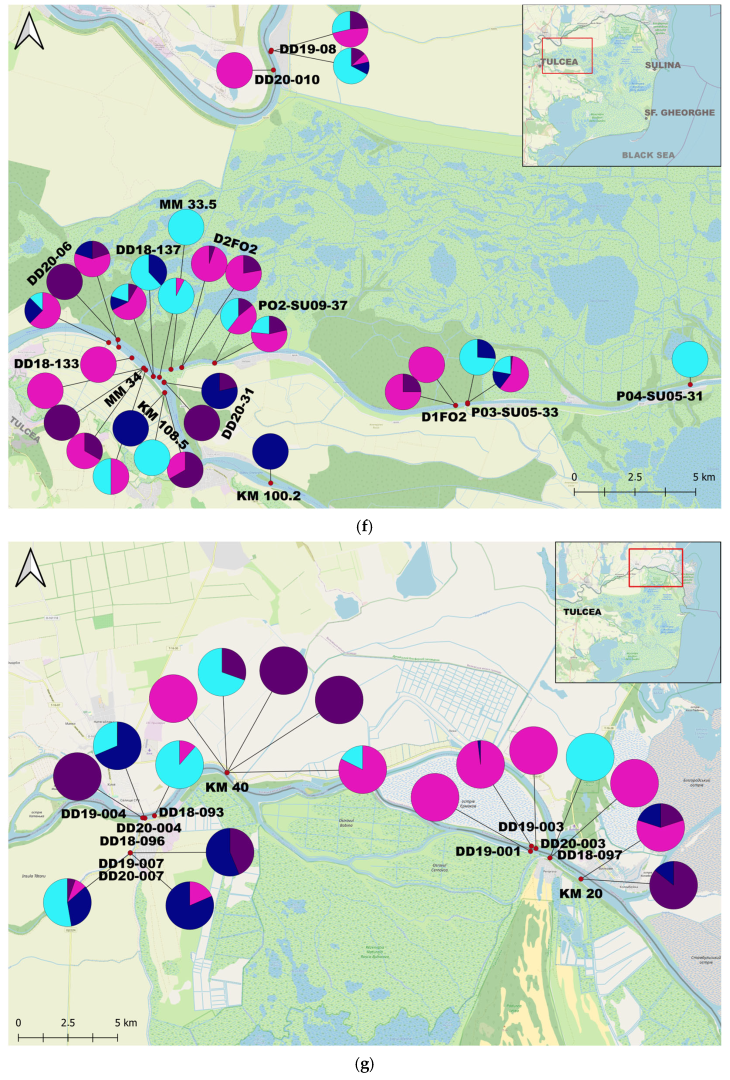
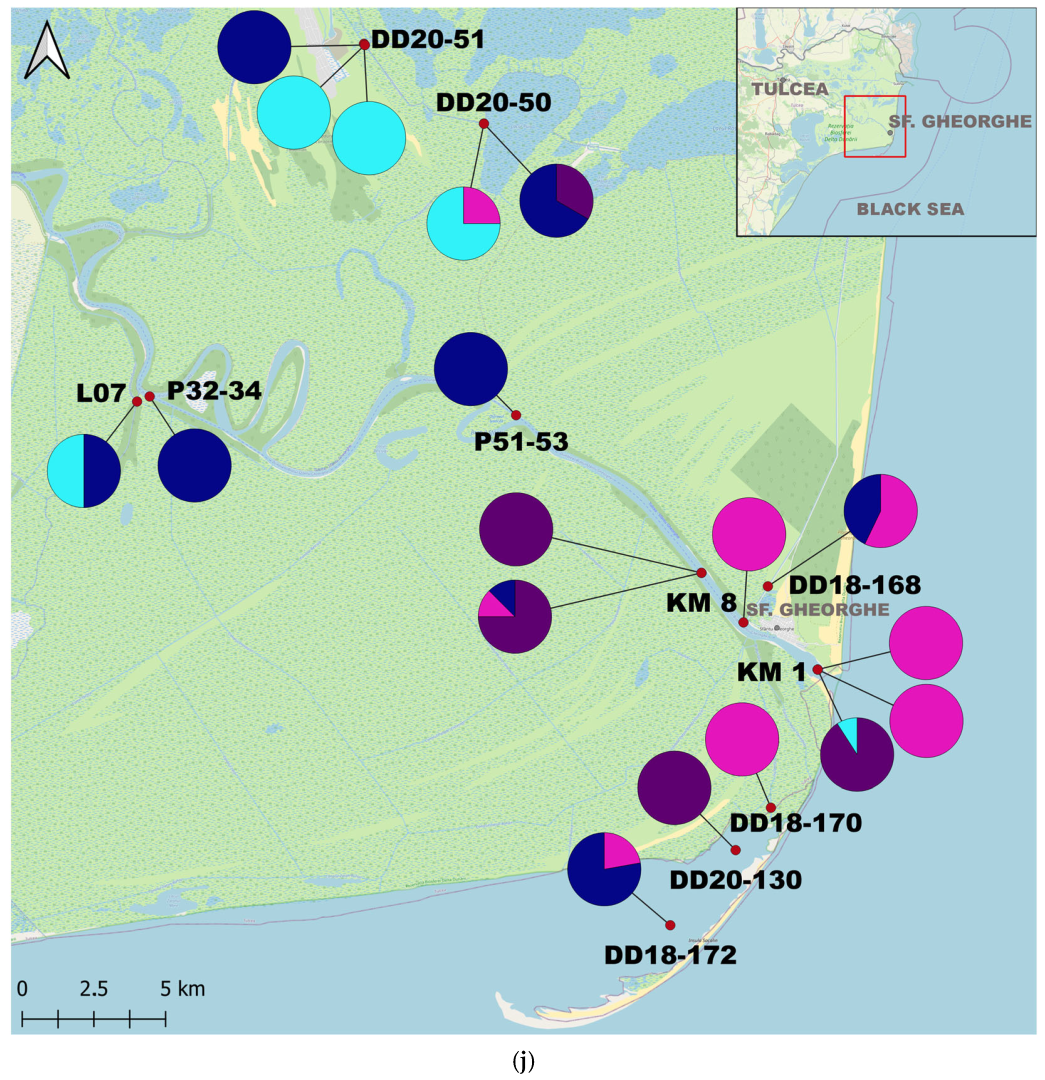
3. Results
4. Discussion
4.1. Historical and Modern Distribution Range Shifts
Ecological and Biological Prerequisites for Successful Invasion of D. polymorpha and C. fluminea and Implications of Their Invasiveness Potential
4.2. The Species Populations’ Dynamics and Trends
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lodge, D.M.; Stein, R.A.; Brown, K.M.; Covish, A.P.; Bronmark, C.; Garvey, J.E.; Klosiewki, S.P. Predicting impact of freshwater exotic species on native biodiversity: Challenges on spatial scaling. Aust. J. Ecol. 1998, 23, 53–67. [Google Scholar] [CrossRef]
- Ricciardi, A.; MacIsaac, H.J. Recent mass invasion of the North American Great Lakes by Ponto-Caspian species. Trends Ecol. Evol. 2000, 15, 62–65. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Ecological predictions and risk assessment for alien fishes in North America. Science 2002, 298, 1233–1236. [Google Scholar] [CrossRef]
- Strayer, D.L. Alien species in fresh waters: Ecological effects, interactions with other stressors, and prospects for the future. Freshw. Biol. 2010, 55 (Suppl. S1), 152–174. [Google Scholar] [CrossRef]
- Simberloff, D. How common are invasion-induced ecosystem impacts? Biol. Invasions 2011, 13, 1255–1268. [Google Scholar] [CrossRef]
- Sousa, R.; Morais, P.; Dias, E.; Antunes, C. Biological invasions and ecosystem functioning: Time to merge. Biol. Invasions 2011, 13, 1055–1058. [Google Scholar] [CrossRef]
- Vaughn, C.C.; Hakenkamp, C.C. The functional role of burrowing bivalves in freshwater ecosystems. Freshw. Biol. 2001, 46, 1431–1446. [Google Scholar] [CrossRef]
- Sousa, R.; Gutierrez, J.L.; Aldridge, D.C. Nonindigenous invasive bivalves as ecosystem engineers. Biol. Invasions 2009, 11, 2367–2385. [Google Scholar] [CrossRef]
- Araujo, R.; Moreno, D.; Ramos, M.A. The Asiatic clam Corbicula fluminea (Müller, 1774) (Bivalvia: Corbiculidae) in Europe. Am. Malacol. Bull. 1993, 10, 39–49. [Google Scholar]
- Zhadin, V. Mollusks of the Fresh and Brackish Waters of the USSR; Academy of Sciences of the USSR: Moscow, Russia; Leningrad, Russia, 1952; 376p. (In Russian) [Google Scholar]
- Morton, B. Corbicula in Asia—An Updated Synthesis. In American Malacological Bulletin, Special Edition; American Malacological Union: Buffalo, NY, USA, 1986; pp. 113–124. [Google Scholar]
- DAISIE (Delivering Alien Invasive Species Inventories to Europe). Handbook of Alien Species in Europe; Springer: Dordrecht, The Netherlands, 2009; pp. 269–374. [Google Scholar]
- Chen, H.; Zha, J.; Liang, X.; Bu, J.; Wang, M.; Wang, Z. Sequencing and de novo assembly of the Asian clam (Corbicula fluminea) transcriptome using the Illumina GAIIx method. PLoS ONE 2013, 8, e79516. [Google Scholar] [CrossRef] [PubMed]
- Chijimatsu, T.; Umeki, M.; Kataoka, Y.; Kobayashi, S.; Yamada, K.; Oda, H.; Mochizuki, S. Lipid components prepared from a freshwater Clam (Corbicula fluminea) extract ameliorate hypercholesterolaemia in rats fed highcholesterol diet. Food Chem. 2013, 136, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Liao, N.; Chen, S.; Ye, X.; Zhong, J.; Wu, N.; Dong, S.; Yang, B.; Liu, D. Antioxidant and anti-tumor activity of a polysaccharide from freshwater clam, Corbicula fluminea. Foods Funct. 2013, 4, 539–548. [Google Scholar] [CrossRef]
- Burch, J.W. Checklist of West American molluscs, family Corbiculidae. Conchol. Club South. Calif. Minutes 1944, 36, 1–18. [Google Scholar]
- Counts, C.L., III. Corbicula fluminea (Bivalvia: Sphaeriacea) in British Columbia. Nautika 1995, 95, 12–13. [Google Scholar]
- Hanna, G.D. Introduced mollusks of western North America. Calif. Acad. Sci. Occas. Pap. 1966, 48, 1–108. [Google Scholar]
- McMahon, R.F. The occurrence and spread of the introduced Asiatic freshwater clam, Corbicula fluminea (Müller) in North America: 1924–1982. Nautilus 1982, 96, 134–141. [Google Scholar]
- Foster, A.M.; Fuller, P.; Benson, A.; Constant, S.; Raikow, D.; Larson, J.; Fusaro, A. Corbicula fluminea; USGS Nonindigenous Aquatic Species Database: Gainesville, FL, USA, 2013. Available online: http://nas.er.usgs.gov/queries/FactSheet.aspx?speciesID=92 (accessed on 27 February 2023).
- CABI. Corbicula fluminea. In Invasive Species Compendium; CAB International: Wallingford, UK, 2013; Available online: www.cabi.org/isc (accessed on 4 January 2023).
- Bij de Vaate, A.; Greijdanus-Klaas, M. The Asiatic clam, Corbicula fluminea Muller, 1774 (Pelecypoda: Corbiculidae), a new immigrant in the Netherlands. Bull. Zool. Mus. 1990, 12, 173–178. [Google Scholar]
- Rey, P.; Ortlepp, J.; Küry, D. Wirbellose Neozoen im Hochrhein. Ausbreitung und ökologische Bedeutung. Schr. Umw. BUWAL Bern 2004, 380, 1–88. [Google Scholar]
- Schmidlin, S.; Bauer, B. Distribution and substrate preference of the invasive clam Corbicula fluminea in the river Rhine in the region of Basel (Switzerland, Germany, France). Aquat. Sci. 2007, 69, 153–161. [Google Scholar] [CrossRef]
- Schmidlin, S.; Schmera, D.; Ursenbacher, S.; Bauer, B. Separate introductions but lack of genetic variability in the invasive clam Corbicula spp. in Swiss lakes. Aquat. Invasions 2012, 7, 73–80. [Google Scholar] [CrossRef]
- Mouthon, J. Sur la presence en France et en Portugal de Corbicula (Bivalvi and Corbiculidae) onginaire d’Asia. Basterie 1981, 45, 109–116. [Google Scholar]
- Kinzelbach, R. Die Körbchenmuscheln Corbicula fluminalis, Corbicula fluminea und Corbicula fluviatilis in Europa (Bivalvia: Corbiculidae). Mainz. Nat. Arch. 1991, 29, 215–228. [Google Scholar]
- Grabow, K.; Martens, A. Vorkomen von Corbicula fluminea (O. F. Müller, 1774) und “C. fluminalis” (O. F. Müller, 1774) im östlichen Mittellandkanal (Bivalvia: Corbiculidae). Mitt. Der Dtsch. Malakozool. Ges. 1995, 56–57, 19–23. [Google Scholar]
- Tittizer, T.; Taxacher, M. Erstnachweis von Corbicula fluminea/fluminalis (Müller, 1774) (Corbiculidae, Mollusca) in der Donau. Lauterbornia 1997, 31, 103–107. [Google Scholar]
- Brauckmann, C.; Brauckmann, B.; Groning, E. Zur Ausbreitung der Korbchenmuschel Corbicula in Mitteleuropa. Jahresber. Nat. Ver. Wupp. 1999, 52, 221–228. [Google Scholar]
- Howlett, D.; Baker, R. Corbicula fluminea (Müller): New to UK. J. Conchol. 1999, 36, 83. [Google Scholar]
- Beran, L. First record of Corbicula fluminea (Mollusca: Bivalvia) in the Czech Republic. Acta Soc. Zool. Bohem. 2000, 64, 1–2. [Google Scholar]
- Beran, L. Spreading expansion of Corbicula fluminea (Mollusca: Bivalvia) in the Czech Republic. Heldia 2006, 65–66, 187–192. [Google Scholar]
- Vincent, T.; Brancotte, V. Répartition actuelle et modes de progression de Corbicula spp. en France. Bull. Soc. Zool. Fr. 2002, 127, 241–252. [Google Scholar]
- Willing, M.J. Sphaerium solidum and Corbicula fluminea: Two rare bivalve molluscs in the River Great Ouse system in Cambridgeshire. Nat. Cambs. 2007, 49, 39–49. [Google Scholar]
- Cianfanelli, S.; Lori, E.; Bodon, M. Non-indigenous freshwater mollusks and their distribution in Italy. In Biological Invaders in Inland Waters: Profiles, Distribution, and Threats. Invading Nature; Springer Series in Invasion, Ecology; Gherardi, F., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 103–121. [Google Scholar]
- Ayres, C. A new record of Asian clam Corbicula fluminea (Müller, 1774) in Galicia (Iberian Peninsula)—Ribeiras do Louro e Gándaras de Budiño wetland. Aquat. Invasions 2008, 3, 439–440. [Google Scholar] [CrossRef]
- Pérez-Quintero, J. Revision of the distribution of Corbicula fluminea (Müller, 1774) in the Iberian Peninsula. Aquat. Invasions 2008, 3, 355–358. [Google Scholar] [CrossRef]
- Elliott, P.; zu Ermgassen, P.S.E. The Asian clam (Corbicula fluminea) in the River Thames, London, England. Aquat. Invasions 2008, 3, 54–60. [Google Scholar] [CrossRef]
- Morais, P.; Teodósio, J.; Reis, J.; Chícharo, M.A.; Chícharo, L. The Asian clam Corbicula fluminea (Müller, 1774) in the Guadiana River Basin (southwestern Iberian Peninsula): Setting the record straight. Aquat. Invasions 2009, 4, 681–684. [Google Scholar] [CrossRef]
- Fabbri, R.; Landi, L. New records of exotic mussels, decapod, crustaceans and fishes from Emilia-Romagna and first record of Corbicula fluminea (O.F. Müller, 1774) in Italy. Quad. Studi E Not. Stor. Nat. Della Romagna 1999, 12, 9–20. [Google Scholar]
- Lois, S. New records of Corbicula fluminea (Müller, 1774) in Galicia (Northwest of the Iberian Peninsula): Mero, Sil and Deva rivers. Aquat. Invasions 2010, 5 (Suppl. S1), S17–S20. [Google Scholar] [CrossRef]
- Marescaux, J.; Pigneur, L.M.; Van Doninck, K. New records of Corbicula clams in French rivers. Aquat. Invasions 2010, 5 (Suppl. S1), S35–S39. [Google Scholar] [CrossRef]
- Lucy, F.E.; Karatayev, A.Y.; Burlakova, L.E. Predictions for the spread, population density, and impacts of Corbicula fluminea in Ireland. Aquat. Invasions 2012, 7, 465–474. [Google Scholar] [CrossRef]
- Kamburska, L.; Lauceri, R.; Beltrami, M.; Boggero, A.; Cardeccia, A.; Guarneri, I.; Manca, M.; Riccardi, N. Establishment of Corbicula fluminea (O.F. Müller, 1774) in Lake Maggiore: A spatial approach to trace the invasion dynamics. BioInvasions Rec. 2013, 2, 105–117. [Google Scholar] [CrossRef]
- Leuven, R.S.E.W.; van der Velde, G.; Baijens, I.; Snijders, J.; van der Zwart, C.; Lenders, H.H.R.; Bij De Vaate, A. The river Rhine: A global highway for dispersal of aquatic invasive species. Biol. Invasions 2009, 11, 1998–2008. [Google Scholar] [CrossRef]
- Tittizer, T.; Leuchs, H.; Banning, M. Das Makrozoobenthos der Donau im Abschnitt Kehlheim-Jochenstein (Donau- km 2414–2202). Limnol. Aktuell Stuttg. 1994, 2, 173–188. [Google Scholar]
- Alexandrov, B.; Boltachev, A.; Kharchenko, T.; Lyashenko, A.; Son, M.; Tsarenko, P.; Zhukinsky, V. Trends of aquatic alien species invasions in Ukraine. Aquat. Invasions 2007, 2, 215–242. [Google Scholar] [CrossRef]
- Son, M.O. Invasive mollusks (Mollusca, Bivalvia, Gastropoda) in the Danube Delta. Vestn. Zool. 2007, 41, 213–218. (In Russian) [Google Scholar]
- Paunović, M. Qualitative composition of the macroinvertebrate communities in the Serbian sector of the Sava River. Int. Assoc. Danub. Res. 2004, 35, 349–354. [Google Scholar]
- Paunović, M.; Csányi, B.; Knežević, S.; Simić, V.; Nenadić, D.; Jakovčev-Todorović, D.; Stojanović, B.; Cakić, P. Distribution of Asian clams Corbicula fluminea (Müller, 1774) and C. fluminalis (Müller, 1774) in Serbia. Aquat. Invasions 2007, 2, 99–106. [Google Scholar] [CrossRef]
- Munjiu, O.; Shubernetski, I. First record of Asian clam Corbicula fluminea (Müller, 1774) in the Republic of Moldova. Aquat. Invasions 2010, 5 (Suppl. S1), S67–S70. [Google Scholar] [CrossRef]
- Bódis, E.; Nosek, J.; Oertel, N.; Tóth, B.; Fehér, Z. A comparative study of two Corbicula morphs (Bivalvia, Corbiculidae) inhabiting River Danube. Int. Rev. Hydrobiol. 2011, 96, 257–273. [Google Scholar] [CrossRef]
- Bódis, E.; Sipkay, C.; Tóth, B.; Oertel, N.; Nosek, J.; Hornung, E. Spatial and temporal variation in biomass and size structure of Corbicula fluminea in Danube River catchment, Hungary. Biologia 2012, 67, 739–750. [Google Scholar] [CrossRef]
- Bij de Vaate, A.; Hulea, O. Range extension of the Asiatic clam Corbicula fluminea (Müller 1774) in the River Danube: First record from Romania. Lauterbornia 2000, 38, 23–26. [Google Scholar]
- Skolka, M.; Gomoiu, M.T. Alien invertebrate species in Romanian waters. Ovidius University Annals of Natural Sciences, Biology. Ecol. Ser. 2001, 5, 51–55. [Google Scholar]
- Popa, O.P. Studiul Unor Specii de Moluste Bivalve Strãine în Fauna României (Faunisticã, Biologie si Diversitate Geneticã a Populatiilor). Ph.D. Thesis, Universitatea din Bucuresti, Soala Doctoralã a Facultãtii de Biologie, Bucuresti, Romania, 2008. (In Romanian). [Google Scholar]
- Catianis, I.; Secrieru, D.; Pojar, I.; Grosu, D.; Scrieciu, A.; Pavel, A.B.; Vasiliu, D. Water Quality, Sediment Characteristics and Benthic Status of the Razim-Sinoie Lagoon System, Romania. Open Geosci. 2018, 10, 12–33. [Google Scholar] [CrossRef]
- Ferreira-Rodriguez, N.; Pavel, A.B.; Cogalniceanu, D. Integrating expert opinion and traditional ecological knowledge in invasive alien species management: Corbicula in Eastern Europe as a model. Biol. Invasions 2021, 23, 1087–1099. [Google Scholar] [CrossRef]
- Golikov, A.; Starobogatov, Y. Zoogeographical characteristics of the gastropod mollusks of Black and Azov Seas. In Biological Studies of the Black Sea and Its Fishery Resources; Nauka Publ.: Moscow, Russia, 1968; pp. 70–83. (In Russian) [Google Scholar]
- Skarlato, O.; Starobogatov, Y. Class of Bivalve Mollusks—Bivalvia. In The Key for the Fauna of the Black and Azov Seas; Mordukhay-Boltovskoy, F., Ed.; Naukova Dumka Publ.: Sydney, Australia, 1972; pp. 178–249. (In Russian) [Google Scholar]
- Valkanov, A.; Petrova, V.; Roshdestvenski, A.; Marinov, T.; Naidenow, W. Black Sea lakes. In The Black Sea; Valkanov, A., Marinov, H., Danov, H., Vladev, P., Eds.; G. Bakalov Publ.: Varna, Bulgaria, 1978; pp. 262–283. (In Bulgarian) [Google Scholar]
- Marinov, T. The Zoobenthos from the Bulgarian Sector of the Black Sea; Publishing House of the Bulgarian Academy of Sciences: Sofia, Bulgaria, 1990; 195p. (In Bulgarian) [Google Scholar]
- Kerney, M.P.; Morton, B.S. The distribution of Dreissena polymorpha in Britain. J. Conchol. 1970, 27, 97–100. [Google Scholar]
- Olenin, S.; Orlova, M.; Minchin, D. Dreissena polymorpha (Pallas, 1771). In Case Histories on Introduced Species: Their General Biology, Distribution, Range Expansion and Impact; Gollasch, S., Minchin, D., Rosenthal, H., Voigt, M., Eds.; Logos-Verlag: Berlin, Germany, 1999; pp. 37–42. [Google Scholar]
- Aldridge, D.C.; Elliot, P.; Moggridge, G.D. The recent and rapid spread of the zebra mussel (Dreissena polymorpha) in Great Britain. Biol. Conserv. 2004, 119, 253–261. [Google Scholar] [CrossRef]
- Kobelt, W.; Haas, F. Iconographie der Land Süsswasser-Mollusken mit vorzüglicher Berücksichtigung der europäischen noch nicht abgebildeten Arten von E. A. Rossmässler. Neue Folge. Siebzehnter Band Wiesb. 1911, 1–60, 451–480. [Google Scholar]
- Wohlberedt, O. Zur Molluskenfauna von Bulgarien. Abh. Ber. Nat. Ges. Görlitz 1911, 27, 167–234. [Google Scholar]
- Hesse, P. Zur Kenntnis der Molluskenfauna von Ostrumelien. III. Nachr. Dtsch. Malakozool. Ges. 1914, 46, 49–58. [Google Scholar]
- Drensky, P. Synopsis and distribution of freshwater Mollusca in Bulgaria. Annuaire de l’Universite de Sofia. Fac. Phys. Math. 1947, 43, 33–54. (In Bulgarian) [Google Scholar]
- Petrbok, J. The freshwater molluscs of the lakes of Varna and of Gebedže. Arbeiten aus der Biologischen Meeresstation in Varna. Bulgarien 1947, 13, 71–75. (In Bulgarian) [Google Scholar]
- Valkanov, A. Katalog unserer Schwarzmeerfauna. Arbeiten aus der Biologischen Meeresstation in Varna. Bulgarien 1957, 19, 1–62. (In Bulgarian) [Google Scholar]
- Russev, B.; Yaneva, I.; Detcheva, R.; Karapetkova, M. Zusammensetzung der Hydrofauna. In Limnologie der Bulgarischen Donauzuflüsse; Russev, B., Ed.; Knizhen Tigar Publ.: Sofia, Bulgaria, 1994; pp. 130–174. (In Bulgarian) [Google Scholar]
- Angelov, A. Mollusca: Gastropoda et Bivalvia aquae dulcis. In Catalogus Faunae Bulgaricae, 4; Pensoft: Sofia, Bulgaria, 2000; 57p. [Google Scholar]
- Shopov, V. Distribution of Upper Quaternary Mollusca Communities in the Outer Zone of the South Bulgarian Black Sea Shelf; Geologia Balcanica, BAS Publ.: Sofia, Bulgaria, 1979; Volume 9, pp. 51–66. [Google Scholar]
- Shopov, V. Biostratigraphy of the Upper Quaternary Sediments from the South-West Parts of the Black Sea; Geologica Balcanica, BAS Publ.: Sofia, Bulgaria, 1984; Volume 14, pp. 17–38. (In Russian) [Google Scholar]
- Hrischev, H.; Shopov, V. Marine Pleistocen in the Bay of Burgas and the Problem of Relations between Uzunlar and Karangat Beds; Geologica Balcanica, BAS Publ.: Sofia, Bulgaria, 1979; Volume 9, pp. 69–84. (In Russian) [Google Scholar]
- Liutzkanov, D. Morphology, Taxonomy and Ecology of Fossil and Recent Dreissena polymorpha Pall. in Bulgaria. Ph.D. Thesis, Institute of Zoology, Bulgarian Academy of Sciences, Sofia, Bulgaria, 1981; 253p. (In Bulgarian). [Google Scholar]
- Hubenov, Z. Dreissena (Bivalvia: Dreissenidae)—Systematics, autochthonous and anthropogenic areas. Acta Zool. Bulg. 2005, 57, 259–268. [Google Scholar]
- Kerney, M.P. Atlas of the Land and Freshwater Molluscs of Britain and Ireland; Harley Books: Colchester, UK, 1999. [Google Scholar]
- Araujo, R.; Alvarez, R.M. El mejillo’n cebra en el Ebro: Un grave caso de riesgo ambiental en Arago’n. Nat. Aragonesa 2001, 8, 39–46. [Google Scholar]
- Minchin, D.; Lucy, F.; Sullivan, M. Zebra mussel: Impacts and spread. In Invasive Aquatic Species of Europe: Distribution, Impact and Management; Leppäkoski, E., Gollasch, S., Olenin, S., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 135–146. [Google Scholar]
- Pollux, B.; Minchin, D.; van der Velde, G.; van Alen, T.; Moon-van der Staay, S.Y.; Hackstein, J. Zebra mussels (Dreissena polymorpha) in Ireland, AFLP-fingerprinting and boat traffic both indicate an origin from Britain. Freshw. Biol. 2003, 48, 1127–1139. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Muncaster, B.W.; Mackie, G.L. Ecological and genetic studies on Dreissena polymorpha (Pallas): A new mollusc in the Great Lakes. Can. J. Fish. Aquat. Sci. 1989, 46, 1491–1587. [Google Scholar] [CrossRef]
- Nalepa, T.F.; Schloesser, D.W. (Eds.) Zebra Mussels Biology, Impacts, and Control, 2nd ed.; Lewis Publishers, CRC Press: Boca Raton, FL, USA, 2013; 815p, ISBN 9780429110863. [Google Scholar]
- MacIsaac, H.J. Potential abiotic and biotic impacts of zebra mussels on the inland waters of North America. Am. Zool. 1996, 36, 287–299. [Google Scholar] [CrossRef]
- SR EN ISO 10870; Water Quality—Guidelines for the Selection of Sampling Methods and DEVICES for Benthic Macroinvertebrates in Fresh Waters. ISS: Belgrade, Serbia, 2012.
- SR EN ISO 16150; Water Quality—Guidance on Pro-Rata Multi-Habitat Sampling of Benthic Macro-Invertebrates from Wade Able Rivers. ISS: Belgrade, Serbia, 2012.
- SR EN ISO 5661–1; Water Quality. Part 1, General Guidelines for Establishing Sampling Programs and Techniques. ISS: Belgrade, Serbia, 2008.
- Godeanu, S. Continental waters, overview. In The Diversity of the Living World. Illustrated Identification Manual of the Flora and Fauna of Romania; Godeanu, S.P., Ed.; Bucura Mond Publishing: Bucharest, Romania, 2002; pp. 1–24. ISBN 973-98248-5-4. [Google Scholar]
- Wentworth, C.K. A scale of grade and class terms for clastic sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Verdonschot, P.F.M. Beken in Beeld; IBN: Wageningen, The Netherlands; RIZA: Leystad, The Netherlands; Ministerie van Verkeer Waterstaat: Den Haag, The Netherlands, 1999; 25p. (In Dutch)
- Black, C.A. Methods of Soil Analysis, Part I; American Society of Agronomy: Madison, WI, USA, 1965; 1572p. [Google Scholar]
- Gaudette, H.; Flight, W.; Toner, L.; Folger, D. An inexpensive titration method for the determination of organic carbon in recent sediments. J. Sediment. Petrol. 1974, 44, 249–253. [Google Scholar]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft: New York, NY, USA, 2020; Available online: https://www.xlstat.com (accessed on 1 February 2023).
- Yang, Y.; Liu, Z.; Chen, F.; Wu, S.; Zhang, L.; Kang, M.; Li, J. Assessment of trace element contamination in sediment cores from the Pearl River and estuary, South China: Geochemical and multivariate analysis approaches. Environ. Monit. Assess. 2014, 186, 8089–8107. [Google Scholar] [CrossRef]
- McMahon, R. Ecology of an invasive pest bivalve, Corbicula. In The Mollusca; Russell-Hunter, W., Ed.; Academic Press: New York, NY, USA, 1983; pp. 505–561. [Google Scholar]
- Linstow, O.V. Beitrag zur Geschichte und Verbreitung von Corbicula fluminalis. Arch. Für Molluskenkd. 1922, 54, 113–144. [Google Scholar]
- Ellis, A.E. British Freshwater Bivalva Mollusca. In Synopses of the British Fauna (New Series) Vol. 11, Linnean Society of London; Academic Press: London, UK, 1978; 109p. [Google Scholar]
- Britton, J.C.; Morton, B. Corbicula in North América: The evidence reviewed and evaluated. In Proceedings of the First International Corbicula Symposium 1977, Fort Worth, TX, USA, 13–15 October 1977; Britton, J.C., Ed.; Texas Christian University: Fort Worth, TX, USA, 1979; pp. 250–287. [Google Scholar]
- Woodward, S.L.; Quinn, J.A. Encyclopedia of Invasive Species; ABC-CLIO, LLC.: Goleta, CA, USA, 2011; 764p. [Google Scholar]
- Birnbaum, C. NOBANIS—Invasive Alien Species Fact Sheet—Dreissena polymorpha—From: Online Database of the European Network on Invasive Alien Species—NOBANIS. 2011. Available online: www.nobanis.org (accessed on 1 January 2023).
- Sousa, R.; Antunes, C.; Guilhermino, L. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: An overview. Ann. Limnol. Int. J. Limnol. 2008, 44, 85–94. [Google Scholar] [CrossRef]
- Zaiko, A.; Daunys, D.; Olenin, S. Habitat engineering by the invasive zebra mussel Dreissena polymorpha (Pallas) in a boreal coastal lagoon: Impact on biodiversity. Helgol. Mar Res. 2009, 63, 85–94. [Google Scholar] [CrossRef]
- Bij de Vaate, A. Colonization of the German part of the River Rhine by the Asiatic clam, Corbicula fluminea Muller, 1774 (Pelecypoda, Corbiculidae). Bull. Zool. Mus. 1991, 13, 13–16. [Google Scholar]
- Csányi, B. Spreading invaders along the Danubian highway: First record of Corbicula fluminea (O.F. Müller 1774) and C. fluminalis (O.F. Müller 1774) in Hungary (Mollusca, Bivalvia). Folia Hist. Nat. Musei Matra. 1999, 23, 343–345. [Google Scholar]
- Fischer, W.; Schultz, P. Erstnachweis Corbicula cf. Fluminea (O.F. Müller 1774) (Mollusca: Bivalvia: Corbiculidae) aus Österreich, sowie ein Nachweis von lebenden Microcolpia daudebartii acicularis (Ferussac 1821) (Mollusca: Gastropoda: Melanopsidae) aus Bad Deutsch-Altenburg (NÖ, Österreich). Club Conchyl. Inf. 1999, 31, 23–26. [Google Scholar]
- Hubenov, Z. Corbiculidae—A new family for the Bulgarian recent malacofauna (Mollusca, Bivalvia). Acta Zool. Bulg. 2001, 53, 61–66. [Google Scholar]
- Vrabec, V.; Čejka, T.; Šporka, F.; Hamerlík, L.; Král, D. First record of Corbicula fluminea (Mollusca, Bivalvia) from Slovakia with a note about its dispersion in Central Europe. Biologia 2003, 58, 942–952. [Google Scholar]
- Lyashenko, A.V.; Sinitzina, O.O.; Voloshkevich, E.V. Exotic benthic invertebrates in the water bodies of the lower reaches of the Danube. Hydrobiol. Zhurnal 2005, 41, 46–56. (In Russian) [Google Scholar]
- Popa, O.P.; Murariu, D. Freshwater bivalve molluscs invasive in Romania. In Biological Invasions: Towards a Synthesis; Pyšek, P., Pergl, J., Eds.; Institute of Ecology of the TU: Berlin, Germany, 2009; pp. 123–133. [Google Scholar]
- Lyashenko, A.V.; Makovskii, V.V. Molluscs of genus Corbicula in the Ukrainian section of the Danube. Hydrobiol. Zhurnal 2011, 47, 43–52. (In Russian) [Google Scholar]
- Sweeney, P. First record of Asian clam Corbicula fluminea (Müller, 1774) in Ireland. Ir. Nat. J. 2009, 30, 147–148. [Google Scholar]
- Caffrey, J.; Evers, S.; Millane, M.; Moran, H. Current status of Ireland’s newest invasive species—The Asian clam Corbicula fluminea (Müller, 1774). Aquat. Invasions 2011, 6, 291–299. [Google Scholar] [CrossRef]
- Hubenov, Z.; Trichkova, T.; Kenderov, L.; Kozuharov, D. Distribution of Corbicula fluminea (Mollusca: Corbiculidae) over an Eleven-Year Period of its Invasion in Bulgaria. Acta Zool. Bulg. 2013, 65, 315–326. [Google Scholar]
- Strayer, D.L.; Smith, L.C. Distribution of the zebra mussel (Dreissena polymorpha) in estuaries and brackish waters. In Zebra Mussels: Biology, Impacts, and Control; Lewis Publishers: Boca Raton, FL, USA, 1993; pp. 715–726. [Google Scholar]
- Ludyanskiy, M.L.; McDonald, D.; MacNeill, D. Impact of the zebra mussel, a bivalve invader. BioScience 1993, 43, 533–544. [Google Scholar] [CrossRef]
- Ram, J.L.; Karim, A.S.; Banno, F.; Kashian, D.R. Invading the invaders: Reproductive and other mechanisms mediating the displacement of zebra mussels by quagga mussels. Int. J. Invertebr. Reprod. Dev. 2011, 56, 21–32. [Google Scholar] [CrossRef]
- Carlton, J.T. Dispersal Mechanisms of the Zebra Mussel (Dreissena polymorpha). In Zebra Mussels: Biology, Impacts, and Control; Lewis Publishers: Boca Raton, FL, USA, 1993; pp. 677–704. [Google Scholar]
- Johnson, L.E.; Ricciardi, A.; Carlton, J.T. Overland dispersal of aquatic invasive species: A risk assessment of transient recreational boating. Ecol. Appl. 2001, 11, 1789–1799. [Google Scholar] [CrossRef]
- Karatayev, A.Y.; Burlakova, L.E.; Mastitsky, S.E.; Padilla, D.K.; Mills, E.L. Contrasting rates of spread of two congeners, Dreissena polymorpha and Dreissena rostriformis bugensis, at different spatial scales. J. Shellfish. Res. 2011, 30, 923–931. [Google Scholar] [CrossRef]
- Kelly, N.E.; Wantola, K.; Weisz, E.; Yan, N.D. Recreational boats as a vector of secondary spread for aquatic invasive species and native crustacean zooplankton. Biol. Invasions 2012, 15, 509–519. [Google Scholar] [CrossRef]
- McMahon, R.F. Evolutionary and physiological adaptations of aquatic invasive animals: R selection versus resistance. Can. J. Fish. Aquat. Sci. 2002, 59, 1235–1244. [Google Scholar] [CrossRef]
- Sousa, R.; Rufino, M.; Gaspar, M.; Antunes, C.; Guilhermino, L. Abiotic impacts on spatial and temporal distribution of Corbicula fluminea (Müller, 1774) in the River Minho Estuary, Portugal. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 98–110. [Google Scholar] [CrossRef]
- van de Velde, S.; Jorissen, E.L.; Neubauer, T.A.; Radan, S.; Pavel, A.B.; Stoica, M.; Van Baak, C.G.C.; Gandara, A.M.; Popa, L.; de Stigter, H.; et al. A conservation palaeobiological approach to assess faunal response of threatened biota under natural and anthropogenic environmental change. Biogeosciences 2019, 16, 2423–2442. [Google Scholar] [CrossRef]
- Gogaladze, A.; Raes, N.; Biesmeijer, J.C.; Ionescu, C.; Pavel, A.B.; Son, M.O.; Gozak, N.; Anistratenko, V.V.; Wesselingh, F.P. Social network analysis and the implications for Pontocaspian biodiversity conservation in Romania and Ukraine: A comparative study. PLoS ONE 2020, 15, e0221833. [Google Scholar] [CrossRef]
- Gogaladze, A.; Son, M.O.; Lattuada, M.; Anistratenko, V.V.; Syomin, V.L.; Pavel, A.B.; Popa, O.P.; Popa, L.O.; ter Poorten, J.J.; Biesmeijer, J.C.; et al. Decline of unique Pontocaspian biodiversity in the Black Sea Basin: A review. Ecol. Evol. 2021, 11, 12923–12947. [Google Scholar] [CrossRef]
- Ingram, W.M. Asiatic clams as potential pests in California water supplies. J. Am. Water Work. Assoc. 1959, 51, 363–370. [Google Scholar] [CrossRef]
- Isom, B.G.; Bowman, C.F.; Johnson, J.T.; Rodgers, E.B. Controlling Corbicula (Asiatic clam) in complex power plant and industrial water systems. In American Malacological Bulletin, Special Edition; American Malacological Union: Buffalo, NY, USA, 1986; Volume 2, pp. 95–98. [Google Scholar]
- Prokopovich, N.P.; Hebert, D.J. Sedimentation in the Delta-Mendota Canal. J. Am. Water Work. Assoc. 1965, 57, 375–382. [Google Scholar] [CrossRef]
- Rosa, I.C.; Pereira, J.L.; Gomes, J.; Saraiva, P.M.; Gonçalves, F.; Costa, R. The Asian clam Corbicula fluminea in the European freshwater-dependent industry: A latent threat or a friendly enemy? Ecol. Econ. 2011, 70, 1805–1813. [Google Scholar] [CrossRef]
- Clarke, K.B. The infestation of waterworks by Dreissena polymorpha, a freshwater mussel. J. Inst. Water Eng. 1952, 6, 370–379. [Google Scholar]
- Mackie, G.L. Biology of the exotic zebra mussel, Dreissena polymorpha, in relation to native bivalves and its potential impact in Lake St. Clair. Hydrobiologia 1991, 219, 251–268. [Google Scholar] [CrossRef]
- MacIsaac, H.J.; Lonnee, C.J.; Leach, J.H. Suppression of microzooplankton by zebra mussels: Importance of mussel size. Freshw. Biol. 1995, 34, 379–387. [Google Scholar] [CrossRef]
- Ricciardi, A.; Whoriskey, F.G.; Rasmussen, J.B. Impact of the Dreissena invasion on native unionid bivalves in the upper St. Lawrence River. Can. J. Fish. Aquat. Sci. 1996, 53, 1434–1444. [Google Scholar] [CrossRef]
- Schloesser, D.W.; Nalepa, T.F.; Mackie, G.L. Zebra mussel infestation of unionid bivalves (Unionidae) in North America. Am. Zool. 1996, 36, 300–310. [Google Scholar] [CrossRef]
- Nalepa, T.F.; Hartson, D.J.; Gostenik, G.W.; Fanslow, D.L.; Lang, G.A. Changes in the freshwater mussel community of Lake St. Clair: From Unionidae to Dreissena polymorpha in eight years. J. Great Lakes Res. 1996, 22, 354–369. [Google Scholar] [CrossRef]
- Nalepa, T.F.; Hartson, D.J.; Fanslow, D.L.; Lang, G.A. Recent population changes in freshwater mussels (Bivalvia: Unionidae) and zebra mussels (Dreissena polymorpha) in Lake St. Clair, USA. Am. Malacol. Bull. 2001, 16, 141–145. [Google Scholar]
- Karatayev, A.Y.; Burlakova, L.E.; Padilla, D.K. The effects of Dreissena polymorpha (Pallas) invasion on aquatic communities in eastern Europe. J. Shellfish Res. 1997, 16, 187–203. [Google Scholar]
- Caraco, N.F.; Cole, J.J.; Raymond, P.A.; Strayer, D.L.; Pace, M.L.; Findlay, S.E.G.; Fischer, D.T. Zebra mussel invasion in a large, turbid river: Phytoplankton response to increased grazing. Ecology 1997, 78, 588–602. [Google Scholar] [CrossRef]
- Bastviken, D.T.E.; Caraco, N.F.; Cole, J.J. Experimental measurements of zebra mussel (Dreissena polymorpha) impact on phytoplankton community composition. Freshw. Biol. 1998, 39, 375–386. [Google Scholar] [CrossRef]
- Pace, M.L.; Findlay, S.E.G.; Fischer, D. Effects of an invasive bivalve on the zooplankton community of the Hudson River. Freshw. Biol. 1998, 39, 103–116. [Google Scholar] [CrossRef]
- Burlakova, L.E.; Karatayev, A.Y.; Padilla, D.K. The impact of Dreissena polymorpha (Pallas) invasion on unionid bivalves. Int. Rev. Hydrobiol. 2000, 85, 529–541. [Google Scholar] [CrossRef]
- Erben, R.; Lajtner, J.; Lucic, A.; Maguire, I.; Klobucar, G.I.V. Attachment of the zebra mussel on the artificial substrates in the reservoir Dubrava (River Drava, Croatia). Int. Assoc. Danub. Res. 2000, 33, 225–231. [Google Scholar]
- Strayer, D.L.; Hattala, K.; Kahnle, A. Effects of an invasive bivalve (Dreissena polymorpha) on fish populations in the Hudson River estuary. Can. J. Fish. Aquat. Sci. 2004, 61, 924–941. [Google Scholar] [CrossRef]
- Maguire, M.; Grey, J. Determination of zooplankton dietary shift following a zebra mussel invasion, as indicated by stable isotope analysis. Freshw. Biol. 2006, 51, 1310–1319. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Nalepa, T.F.; Fahnenstiel, G.L. Dreissena polymorpha in the Saginaw Bay, Lake Huron Ecosystem: Overview and perspective. J. Great Lakes Res. 1995, 21, 411–416. [Google Scholar] [CrossRef]
- Olenin, S. Comparative study of the south-eastern Baltic coastal zone and the Curonian Lagoon bottom communities. In Proceedings of the 13th Baltic Marine Biologists Symposium; Andrushaitis, A., Ed.; Institute of Aquatic Ecology, University of Latvia: Riga, Latvia, 1997; pp. 151–159. [Google Scholar]
- Stewart, T.W.; Miner, J.G.; Lowe, R.L. Quantifying mechanisms for zebra mussel eVects on benthic macroinvertebrates: Organic matter production and shell-generated habitat. J. N. Am. Benthol. Soc. 1998, 17, 81–94. [Google Scholar] [CrossRef]
- Karatayev, A.Y.; Burlakova, L.E.; Padilla, D.K. Impacts of zebra mussels on aquatic communities and their role as ecosystem engineers. In Invasive Aquatic Species of Europe: Distribution, Impacts and Management; Leppakoski, E., Gollasch, S., Olenin, S., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 433–446. [Google Scholar]
- Vanderploeg, H.A.; Nalepa, T.F.; Jude, D.J.; Mills, E.L.; Holeck, K.T.; Liebig, J.R.; Grigorovich, I.A.; Ojaveer, H. Dispersal and emerging ecological impacts of Ponto-Caspian species in the Laurentian Great Lakes. Can. J. Fish. Aquat Sci. 2002, 59, 1209–1228. [Google Scholar]
- Bially, A.; MacIsaac, H.J. Fouling mussels (Dreissena spp.) colonize soft sediments in Lake Erie and facilitate benthic invertebrates. Freshw. Biol. 2000, 43, 85–97. [Google Scholar] [CrossRef]
- Beekey, M.A.; McCabe, D.J.; Marsden, J.E. Zebra mussel colonisation of soft sediments facilitates invertebrate communities. Freshw. Biol. 2004, 49, 535–545. [Google Scholar] [CrossRef]
- Hecky, R.E.; Smith, R.E.H.; Barton, D.R.; Guilford, S.J.; Taylor, W.D.; Charlton, M.N.; Howell, T. The nearshore phosphorus shunt: A consequence of ecosystem engineering by dreissenids in the Laurentian Great Lakes. Can. J. Fish. Aquat Sci. 2004, 61, 1285–1293. [Google Scholar] [CrossRef]
- Karatayev, A.Y.; Lyakhnovich, V.P.; Afanasiev, S.A.; Burlakova, L.E.; Zakutsky, V.P.; Lyakhov, S.M.; Miroshnichenko, M.P.; Moroz, T.G.; Nekrasova, M.Y.; Skalskaya, I.A.; et al. The place of species in ecosystem. In Freshwater Zebra Mussel Dreissena polymorpha (Pall.) (Bivalvia, Dreissenidae); Starobogatov, J.I., Ed.; Systematics, Ecology, Practical Meaning; Nauka Press: Moscow, Russia, 1994; pp. 206–221. (In Russian) [Google Scholar]
- Botts, P.; Silver, B.; Patterson, A.; Schloesser, D.W. Zebra mussel effects on benthic invertebrates: Physical or biotic? J. N. Am. Benthol. Soc. 1996, 15, 179–184. [Google Scholar] [CrossRef]
- Gutierrez, J.L.; Jones, C.G.; Strayer, D.L.; Iribame, O.O. Mollusks as ecosystem engineers: The role of shell production in aquatic habitats. Oikos 2003, 101, 79–90. [Google Scholar] [CrossRef]
- Beekey, M.A.; McCabe, D.J.; Marsden, J.E. Zebra mussels affect benthic predator foraging success and habitat choice on soft sediments. Oecologia 2004, 141, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Karatayev, A.Y.; Howells, R.G.; Burlakova, L.E.; Sewell, B.D. History of spread and current distribution of Corbicula fluminea (Müller) in Texas. J. Shellfish. Res. 2005, 24, 553–559. [Google Scholar]
- Pacioglu, O.; Duţu, L.; Duţu, F.; Pavel, A.B. Habitat preferences and trophic interactions of the benthic invertebrate communities inhabiting depositional and erosional banks of a meander from Danube Delta (Romania). Glob. Ecol. Conserv. 2022, 38, e02213. [Google Scholar] [CrossRef]
- Pacioglu, O.; Duţu, F.; Pavel, A.B.; Duţu, L.T. The influence of hydrology and sediment grain-size on the spatial distribution of macroinvertebrate communities in two submerged dunes from the Danube Delta (Romania). Limnetica 2022, 41, 85–100. [Google Scholar] [CrossRef]
- Burlakova, L.E.; Karatayev, A.Z.; Karatayev, A.V. Invasive mussels induce com-munity changes by increasing habitat complexity. Hydrobiologia 2012, 685, 121–134. [Google Scholar] [CrossRef]
- Dieterich, A.; Mörtl, M.; Eckmann, R. The effects of zebra mussels (Dreissena polymorpha) on the foraging success of Eurasian perch (Perca fluviatilis) and ruffe (Gymnocephalus cernuus). Int. Rev. Hydrobiol. 2004, 89, 229–237. [Google Scholar] [CrossRef]
- Mayer, C.M.; Rudstam, L.G.; Mills, E.L.; Cardiff, S.G.; Bloom, C.A. Zebra mussels (Dreissena polymorpha), habitat alteration, and yellow perch (Perca flavescens) foraging: System-wide effects and behavioural mechanisms. Can. J. Fish. Aquat. Sci. 2001, 58, 2459–2467. [Google Scholar] [CrossRef]
- Ricciardi, A.; Whoriskey, F.G.; Rasmussen, J.B. The role of the zebra mussel (Dreissena polymorpha) in structuring macroinvertebrate communities on hard substrata. Can. J. Fish. Aquat. Sci. 1997, 54, 2596–2608. [Google Scholar] [CrossRef]
- Nosek, J.; Oertel, N.; Bódis, E.; Tóth, B. A bentikus szervesanyag és a makroger-inctelen társulások tér- és id”obeli változása a Duna Kismaros (1688 fkm) és Göd (1668 fkm) közötti szakaszán (Spatial and temporal changes of benthic organicmatter and macroinvertebrate communities in the Kismaros (1688 rkm)-Göd (1668 rkm) section of the River Danube). Acta Biol. Debrecina Suppl. Oecol. Hung. 2009, 20, 165–179. [Google Scholar]
- Pavel, A.B.; Menabit, S.; Pop, I.C.; Stanescu, I.; Lupascu, N. The spatio-temporal distribution of the Ponto-Caspian polychaete in the Lower Sector of the Danube River and in Danube Delta. Glob. Ecol. Conserv. 2021, 28, e01623. [Google Scholar] [CrossRef]
- Aldridge, D.C.; Müller, S.J. The Asiatic clam, Corbicula fluminea, in Britain: Current status and potential impacts. J. Conchol. 2001, 37, 177–183. [Google Scholar]
- Beran, L. Současný stav invaze a neobvyklá lokalita korbikuly asijské [The current state of the invasion and the unusual habitat of the Asian clam]. Živa 2013, 1, 25. (In Czech) [Google Scholar]
- Sebestye´n, O. Colonization of two new fauna elements of Pontus-origin (Dreissena polymorpha Pall. and Corophium curvispinum G.O. Sars forma devium Wundsch) in Lake Balaton. Verh. Int. Ver. Theor. Angew. Limnol. 1938, 8, 169–181. [Google Scholar]
- Balogh, C.; Musko, I.B.; G-To´th, L.; Nagy, L. Quantitative trends of zebra mussels in Lake Balaton (Hungary) in 2003–2005 at different water levels. Hydrobiologia 2008, 613, 57–69. [Google Scholar] [CrossRef]
- Orlova, M.I.; Panov, V.E. Establishment of the zebra mussel, Dreissena polymorpha (Pallas), in the Neva Estuary (Gulf of Finland, Baltic Sea): Distribution, population structure and possible impact on local unionid bivalves. Hydrobiologia 2004, 514, 207–217. [Google Scholar] [CrossRef]
- Sporka, F.; Nagy, S. The macrozoobenthos of parapotamon-type side arms of the Danube River in Slovakia and its response to flowing conditions. Biol. Bratisl. 1998, 53/5, 633–643. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
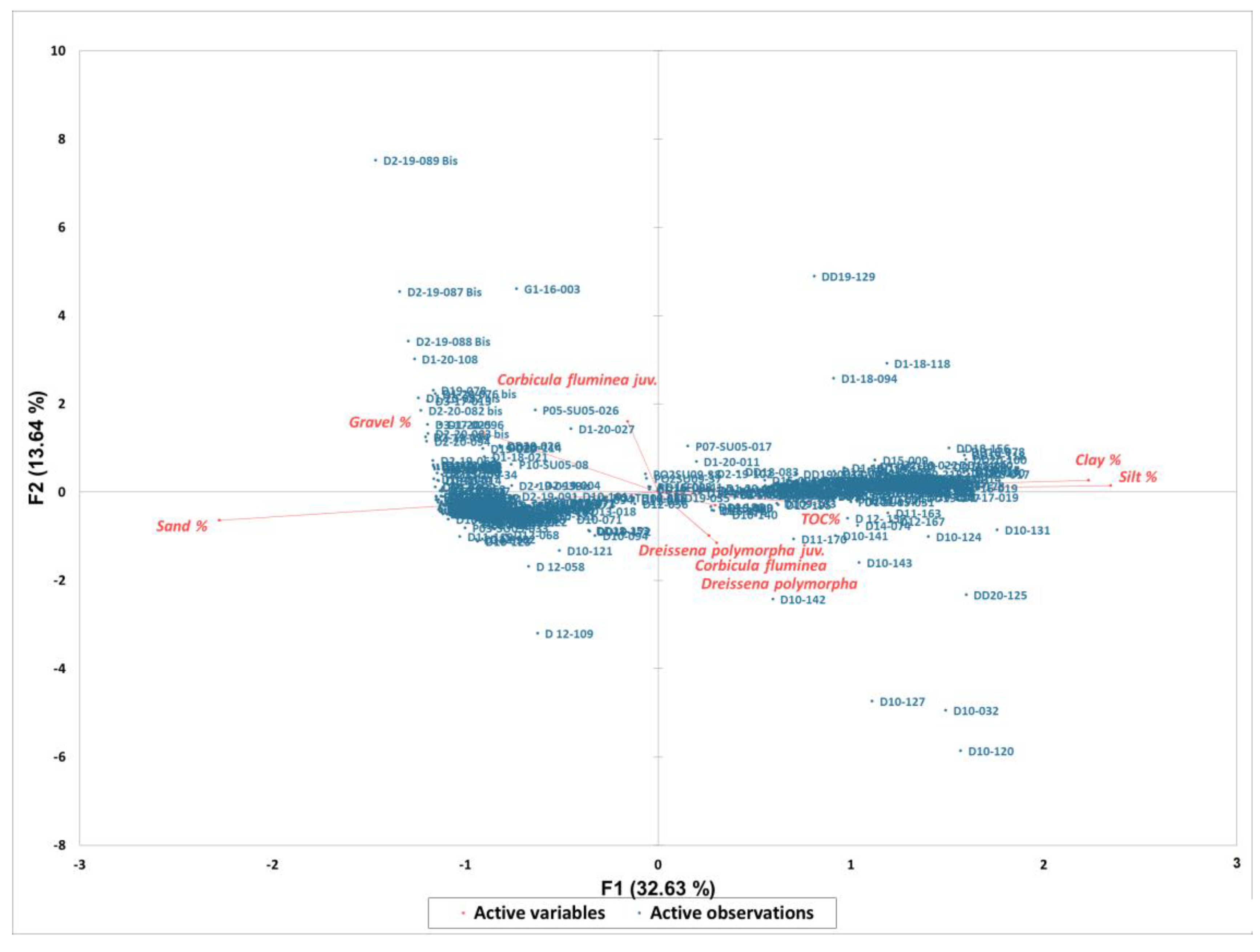
| Correlation Matrix Pearson (n)): | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Variables | Corbicula fluminea | Corbicula fluminea juv. | Dreissena polymorpha | Dreissena polymorpha juv. | Gravel % | Sand % | Silt % | Clay % | TOC% |
| Corbicula fluminea | 1 | −0.009 | 0.182 | 0.066 | −0.028 | −0.061 | 0.074 | 0.038 | 0.017 |
| Corbicula fluminea juv. | −0.009 | 1 | −0.042 | −0.027 | 0.198 | −0.035 | −0.031 | −0.018 | −0.078 |
| Dreissena polymorpha | 0.182 | −0.042 | 1 | −0.009 | −0.067 | −0.065 | 0.086 | 0.059 | −0.005 |
| Dreissena polymorpha juv. | 0.066 | −0.027 | −0.009 | 1 | −0.032 | −0.075 | 0.081 | 0.068 | 0.013 |
| Gravel % | −0.028 | 0.198 | −0.067 | −0.032 | 1 | 0.041 | −0.345 | −0.301 | −0.119 |
| Sand % | −0.061 | −0.035 | −0.065 | −0.075 | 0.041 | 1 | −0.934 | −0.859 | −0.234 |
| Silt % | 0.074 | −0.031 | 0.086 | 0.081 | −0.345 | −0.934 | 1 | 0.804 | 0.272 |
| Clay % | 0.038 | −0.018 | 0.059 | 0.068 | −0.301 | −0.859 | 0.804 | 1 | 0.181 |
| TOC% | 0.017 | −0.078 | −0.005 | 0.013 | −0.119 | −0.234 | 0.272 | 0.181 | 1 |
| F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | |
|---|---|---|---|---|---|---|---|---|
| Eigenvalue | 2.937 | 1.228 | 1.150 | 1.002 | 0.908 | 0.795 | 0.779 | 0.201 |
| Variability (%) | 32.628 | 13.640 | 12.780 | 11.138 | 10.085 | 8.835 | 8.659 | 2.235 |
| Cumulative % | 32.628 | 46.268 | 59.048 | 70.186 | 80.271 | 89.106 | 97.765 | 100.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavel, A.B.; Gheablau, C.; Kreuter, S.; Catianis, I.; Scrieciu, A.; Enache, A. The Spatio-Temporal Distribution of the Freshwater Bivalves Corbicula fluminea and Dreissena polymorpha in the Lower Sector of the Danube River and the Danube Delta. Sustainability 2023, 15, 8526. https://doi.org/10.3390/su15118526
Pavel AB, Gheablau C, Kreuter S, Catianis I, Scrieciu A, Enache A. The Spatio-Temporal Distribution of the Freshwater Bivalves Corbicula fluminea and Dreissena polymorpha in the Lower Sector of the Danube River and the Danube Delta. Sustainability. 2023; 15(11):8526. https://doi.org/10.3390/su15118526
Chicago/Turabian StylePavel, Ana Bianca, Catalin Gheablau, Sylvain Kreuter, Irina Catianis, Albert Scrieciu, and Alin Enache. 2023. "The Spatio-Temporal Distribution of the Freshwater Bivalves Corbicula fluminea and Dreissena polymorpha in the Lower Sector of the Danube River and the Danube Delta" Sustainability 15, no. 11: 8526. https://doi.org/10.3390/su15118526
APA StylePavel, A. B., Gheablau, C., Kreuter, S., Catianis, I., Scrieciu, A., & Enache, A. (2023). The Spatio-Temporal Distribution of the Freshwater Bivalves Corbicula fluminea and Dreissena polymorpha in the Lower Sector of the Danube River and the Danube Delta. Sustainability, 15(11), 8526. https://doi.org/10.3390/su15118526