
Elevated Atmospheric CO2 Concentration Influences the Rooting Habits of Winter-Wheat (Triticum aestivum L.) Varieties


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Layout
2.2. Measurements
2.3. Analysis
3. Results
3.1. Effects of Water Shortage and Elevated Atmospheric CO2 Concentration on Yield Parameters
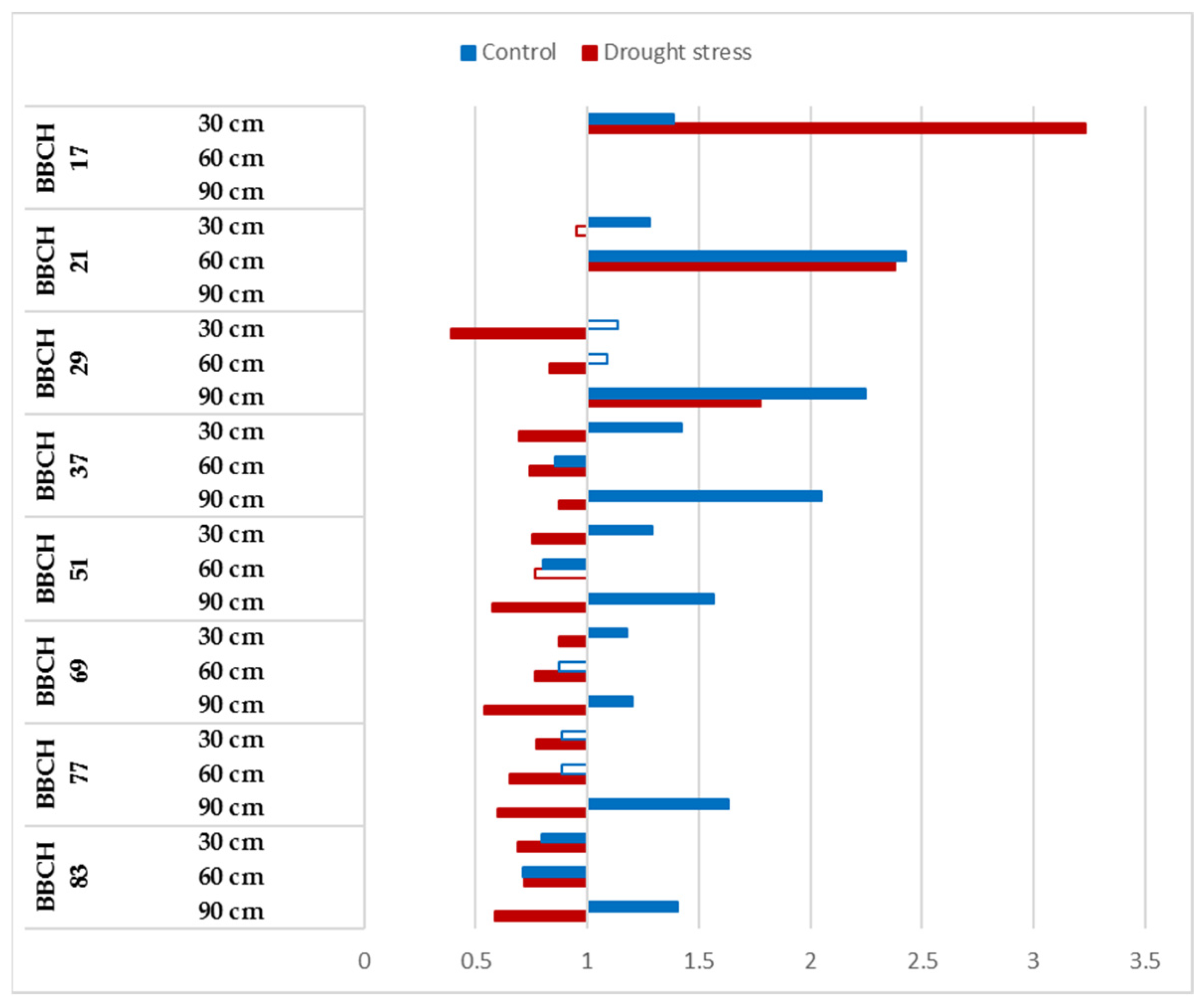
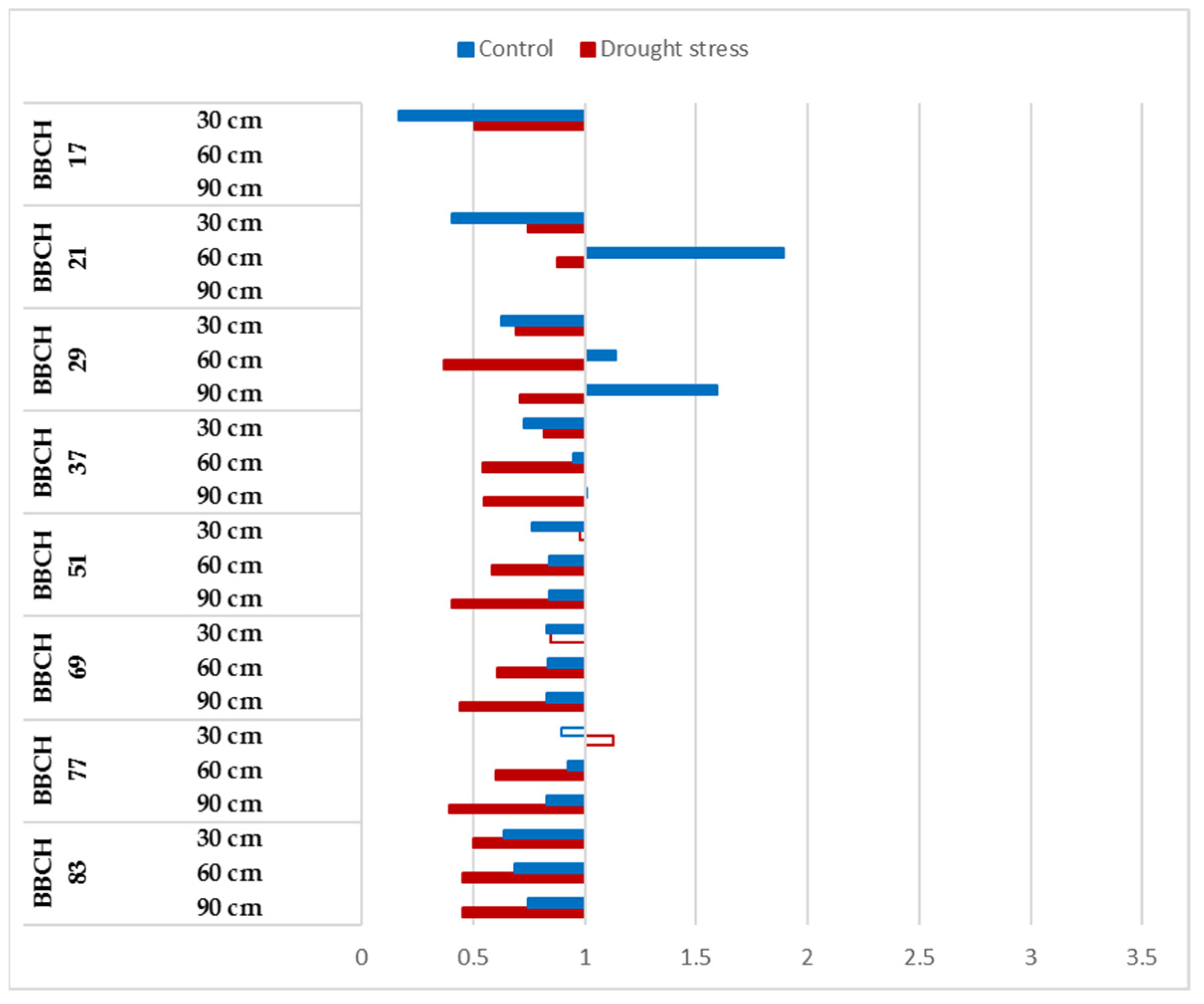
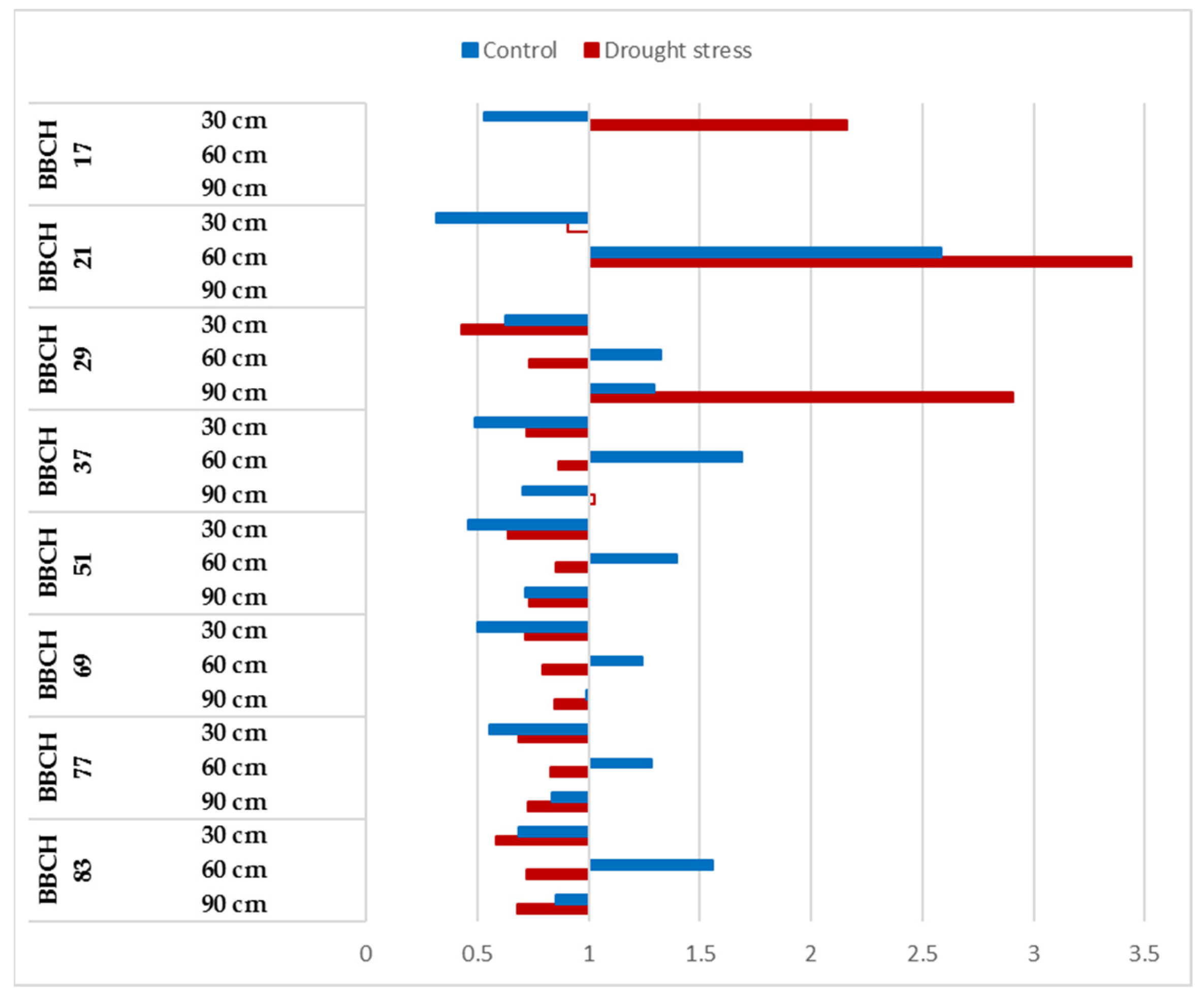
3.2. Dynamics of the Root Development of Winter-Wheat Varieties under Optimum Watering and Drought-Stressed Conditions Grown at Ambient and Elevated CO2 Concentrations
3.3. CO2 Reactions of Winter-Wheat Genotypes during Vegetation at Different Soil Layers under Well-Watered and Drought-Stressed Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change. Climate Change 2013: The Physical Science Basis–Working Group I Contribution to the IPCC Fifth Assessment Report; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Matiu, M.; Ankrest, D.P.; Menzel, A. Interactions between temperature and drought in global and regional crop yield variability during 1961–2014. PLoS ONE 2017, 12, e0178339. [Google Scholar] [CrossRef] [PubMed]
- Lobell, D.B.; Hammer, G.L.; McLean, G.; Messina, C.; Roberts, M.J.; Schlenker, W. The critical role of extreme heat for maize production in the United States. Nat. Clim. Chang. 2013, 3, 497–501. [Google Scholar] [CrossRef]
- Good, P.; Both, B.B.B.; Chadwick, R.; Hawkins, E.; Jonko, A.; Lowe, J.A. Large differences in regional precipitation change between a first and second 2 K of global warming. Nat. Commun. 2016, 7, 13667. [Google Scholar] [CrossRef] [PubMed]
- Leng, G.; Hall, J. Crop yield sensitivity of global major agricultural countries to droughts and the projected changes in the future. Sci. Total Environ. 2019, 654, 811–821. [Google Scholar] [CrossRef]
- Lesk, C.; Rowhani, P.; Ramankutty, N. Influence of extreme weather disasters on global crop production. Nature 2016, 529, 84–87. [Google Scholar] [CrossRef]
- Varga, B.; Vida, G.; Varga-László, E.; Hoffmann, B.; Veisz, O. Combined effects of drought stress and elevated atmospheric CO2 concentration on the yield parameters and water use properties of winter wheat (Triticum aestivum L.). J. Agron. Crop. Sci. 2017, 203, 192–205. [Google Scholar] [CrossRef]
- Farkas, Z.; Varga-László, E.; Anda, A.; Veisz, O.; Varga, B. Effects of waterlogging, drought and their combination on yield and water-use efficiency of five Hungarian winter wheat varieties. Water 2020, 12, 1318. [Google Scholar] [CrossRef]
- Chun, J.A.; Wang, Q.; Timlin, D.; Fleischer, D.H.; Reddy, V.R. Effect of elevated carbon dioxide and water stress on gas exchange and water use efficiency in corn. Agric. For. Meteorol. 2011, 151, 378–384. [Google Scholar] [CrossRef]
- Meng, F.C.; Zhang, J.H.; Yao, F.M.; Hao, C. Interactive effects of elevated CO2 concentration and irrigation on photosynthetic parameters and yield of maize in Northeast China. PLoS ONE 2014, 5, e98318. [Google Scholar] [CrossRef]
- Wang, M.; Xie, B.; Fu, Y.; Dong, C.; Hui, L.; Guanghui, L.; Liu, H. Effects of different elevated CO2 concentrations on chlorophyll contents gas exchange, water use efficiency, and PSII activity on C3 and C4 cereal crops in a closed artificial ecosystem. Photosynth. Res. 2015, 126, 351–362. [Google Scholar] [CrossRef]
- Franzaring, J.; Holz, I.; Fangmeier, A. Responses of old and modern cereals to CO2-fertilisation. Crop Pasture Sci. 2013, 64, 943–956. [Google Scholar] [CrossRef]
- Rogers, E.D.; Benfrey, P.N. Regulation of plant root system architecture: Implication for crop advancement. Curr. Opin. Biotechnol. 2015, 32, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Ristova, D.; Busch, W. Natural variation of root traits: From development to nutrient uptake. Plant Physiol. 2014, 166, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Benlloch-Gonzalez, M.; Bochicchio, R.; Berger, J.; Bramley, H.; Palta, J.A. High temperature reduces the positive effects of elevated CO2 on wheat root system growth. Field Crop. Res. 2014, 165, 71–79. [Google Scholar] [CrossRef]
- Zuo, Q.; Jie, F.; Zhang, R.; Mend, L. A generalized function of wheat’s root length density distribution. Vadose Zone J. 2004, 3, 271–277. [Google Scholar] [CrossRef]
- Wasson, A.P.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Richards, R.A.; Watt, M. Soil coring at multiple field environments can directly quantify variation in deep root traits to select wheat genotypes for breeding. J. Exp. Bot. 2014, 65, 6231–6249. [Google Scholar] [CrossRef]
- Zhao, G.; Liu, J.; Ciu, J.; Wang, H.; Wen, G. Revealing the mechanism of the force dragging the soft bag in the dynamic process pf deep soil coring. Powder Technol. 2019, 344, 251–259. [Google Scholar] [CrossRef]
- Cai, G.; Vanderbought, J.; Klotzche, A.; van der Kruk, J.; Neumann, J.; Hermes, N.; Vereecken, H. Construction of Minirhizotron Facilities for Investigating Root Zone Processes. Vadose Zone J. 2016, 15, vzj2016.05.0043. [Google Scholar] [CrossRef]
- Weigand, M.; Kemna, A. Imaging and functional characterization of crop root systems using spectroscopic electrical impedance measurements. Plant Soil 2019, 435, 201–224. [Google Scholar] [CrossRef]
- Weigand, M.; Kemna, A. Multi-frequency electrical impedance tomography as a non-invasive tool to characterize and monitor the crop root system. Biogeosciences 2017, 14, 921–939. [Google Scholar] [CrossRef]
- Xu, Z.; Valdes, C.; Clarke, J. Existing and potential statistical and computational approaches for the analysis of 3D CT images of plant roots. Agronomy 2018, 8, 71. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Pound, M.P.; Bennett, M.J.; Wells, D.M. Uncovering the hidden half of the plants using new advances in root phenotyping. Curr. Opin. Biotechnol. 2019, 55, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Tracy, S.R.; Nagel, K.A.; Postman, J.A.; Fassbender, H.; Wasson, A.; Watt, M. Crop improvement from phenotyping roots: Highlights reveal expanding opportunities. Trends Plant Sci. 2020, 25, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Barnett, S.; Zhao, S.; Ballard, R.; Franco, C. Selection of microbes for control of Rhizoctonia root rot on wheat using a high throughput pathosystem. Biol. Control. 2017, 113, 45–57. [Google Scholar] [CrossRef]
- Jaradat, A.A. Wheat landraces: A mini-review. Emir. J. Food Agric. 2013, 25, 20–29. [Google Scholar] [CrossRef]
- Abido, W.E.A.; Zsombik, L. Effect of water stress on germination of some Hungarian wheat landraces varieties. Acta Ecol. Sin. 2018, 38, 422–428. [Google Scholar] [CrossRef]
- Nagy, É.; Lehoczki-Krsjak, S.; Lantos, C.; Pauk, J. Phenotyping for testing drought tolerance on wheat varieties of different origins. S. Afr. J. Bot. 2018, 116, 216–221. [Google Scholar] [CrossRef]
- Dowla, M.A.N.; Edwards, I.; O’Hara, G.; Islam, S.; Ma, W. Developing Wheat for Improved Yield and Adaptation Under a Changing Climate: Optimization of a Few Key Genes. Engineering 2018, 4, 514–522. [Google Scholar] [CrossRef]
- Gulyás, G.; Bognár, Z.; Láng, L.; Bedő, Z. Distribution of dwarfing genes (Rht-B1b and Rht-D1b) in Martonvásár wheat breeding materials. Acta Agron. Hung. 2011, 59, 249–254. [Google Scholar] [CrossRef]
- Tischner, T.; Kőszegi, B.; Veisz, O. Climatic programmes used on Martonvásár phytotron most effectively in recent years. Acta Agron. Hung. 1997, 45, 85–104. [Google Scholar]
- Lancashire, P.D.; Bleiholder, H.; van den Boom, T.; Langlüdekke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Kimball, B.A.; Pinter, P.J.; Garcia, R.L.; LaMorte, R.L.; Wall, G.W.; Hunsaker, D.J. Productivity and water use of wheat under free-air CO2 enrichment. Glob. Chang. Biol. 1995, 1, 429–442. [Google Scholar] [CrossRef]
- Högy, P.; Wiese, H.; Koehler, P.; Schwadorf, K.; Breuer, J.; Franzaring, J. Effects of elevated CO2 on grain yield and quality of wheat: Results from a 3-year free-air CO2 enrichment experiment. Plant Biol. 2009, 11, 60–69. [Google Scholar] [CrossRef]
- Manderscheid, R.; Erbs, M.; Weigel, H.-J. Key physiological parameters related to differences in biomass production of maize and four sorghum cultivars under drought and free-air CO2 enrichment. Procedia Environ. Sci. 2015, 29, 89–90. [Google Scholar] [CrossRef][Green Version]
- Qiao, Y.; Miao, S.; Li, Q.; Jin, J.; Lou, X.; Tang, C. Elevated CO2 and temperature increase grain oil concentration but their impacts on grain yield differ between soybean and maize grown in a temperate region. Sci. Total Environ. 2019, 666, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, Y.; Yang, L. Response of rice yield traits to elevated atmospheric CO2 concentration and its interaction with cultivar, nitrogen application rate and temperature: A meta-analysis of 20 years FACE studies. Sci. Total Environ. 2020, 764, 142797. [Google Scholar] [CrossRef]
- Weigel, H.-J.; Manderscheid, R. Crop growth responses to free-air CO2 enrichment and nitrogen fertilization: Rotating barley, ryegrass, sugar beet and wheat. Eur. J. Agron. 2012, 43, 97–107. [Google Scholar] [CrossRef]
- Zheng, Y.; He, C.; Guo, L.; Hao, L.; Cheng, D.; Li, F.; Peng, Z.; Xu, M. Soil water status triggers CO2 fertilization effect on the growth of winter wheat (Triticum aestivum). Agric. For. Meteorol. 2020, 291, 108097. [Google Scholar] [CrossRef]
- Zhang, X.; Shi, Z.; Jiang, D.; Högy, P.; Fangmeier, A. Independent and combined effects of elevated CO2 and post-anthesis heat stress on protein quantity and quality in spring wheat grains. Food Chem. 2019, 277, 524–530. [Google Scholar] [CrossRef]
- Sinha, P.G.; Saradhi, P.P.; Upverty, D.C.; Bhatnagar, A.K. Effect of elevated CO2 concentration on photosynthesis and flowering in three wheat species belonging to different ploidies. Agric Ecosyst. Environ. 2011, 142, 432–436. [Google Scholar] [CrossRef]
- Manderscheid, R.; Dier, M.; Erbst, M.; Sickora, J.; Weigel, H.-J. Nitrogen supply–A determinant in water use efficiency of winter wheat grown under free-air CO2 enrichment. Agric Water Manag. 2018, 210, 70–77. [Google Scholar] [CrossRef]
- Severini, A.D.; Wasson, A.P.; Evans, J.R.; Richards, R.A.; Watt, M. Root phenotypes at maturity in diverse wheat and triticale genotypes grown in three field experiments: Relationships to shoot selection, biomass, grain yield, flowering time, and environment. Field Crop. Res. 2020, 255, 107870. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Condon, A.G.; Macdonald, B.; Rebetzke, G.J.; Awasi, M.-A.; Borgognone, M.G.; Peake, A.; Pinare-Chavez, F.J.; Hungt, A.; Jackway, P.; et al. Genotypic variation for lodging tolerance in spring wheat: Wider and deeper root plates, a feature of low lodging, high yielding germplasm. Field Crop Res. 2020, 258, 1078942. [Google Scholar] [CrossRef]
- Hansen, E.M.O.; Hauggaard-Nielsen, H.; Launay, M.; Rose, P.; Mikkelsen, T.N. The impact of ozone exposure, temperature and CO2 on the growth and yield of three spring wheat varieties. Environ. Exp. Bot. 2019, 168, 103868. [Google Scholar] [CrossRef]
- Erice, G.; Sanz-Sáez, Á.; González-Torralba, J.; Méndez-Espinoza, A.M.; Urretavizcaya, I.; Nieto, M.T.; Serret, M.D.; Araus, J.L.; Irigoyen, J.J.; Aranjuelo, I. Impact of elevated CO2 and drought on yiled and quality traits of a historical (Blanqueta) and a modern (Sula), durum wheat. J. Cereal. Sci. 2019, 87, 194–201. [Google Scholar] [CrossRef]
- Asseng, S.; Ritchie, J.T.; Smucker, A.J.M.; Robertson, M.J. Root growth and water uptake during water deficit and recovering in wheat. Plant Soil 1998, 201, 265–273. [Google Scholar] [CrossRef]
- Uddin, S.; Löw, M.; Parvin, S.; Fitzgerald, G.; Bahrami, H.; Tausz-Posch, S.; Armstrong, R.; O’Leary, G.; Tausz, M. Water use and growth responses of dryland wheat grown under elevated [CO2] are associated with root length in deeper, but not upper soil layer. Field Crops Res. 2018, 224, 170–181. [Google Scholar] [CrossRef]
- Mitchell, J.H.; Chapman, S.C.; Rebetzke, G.J.; Bonnett, D.G.; Fukai, S. Evaluation of a reduced-tilling (tin) gene in wheat lines grown across different production environments. Crop Pasture Sci 2012, 63, 128–141. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Christopher, J.; Devoil, P.; Hammer, G.L. The role of root architectural traits in adaptation of wheat to water-limited environments. Funct. Plant Biol. 2006, 33, 823–837. [Google Scholar] [CrossRef]
- Siddique, K.H.M.; Belford, R.K.; Tennant, D. Root: Shoot ratios of old and modern, tall and semi-dwarf wheat in a Mediterranean environment. Plant Soil 1990, 121, 89–98. [Google Scholar] [CrossRef]
- Miralles, D.J.; Slafer, G.A.; Lynch, V. Rooting patterns in near-isogenic lines of spring wheat for dwarfism. Plant Soil 1997, 197, 79–86. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BBCH Codes | Explanation |
|---|---|
| BBCH 17 | Leaf development: 7 or more leaves unfolded |
| BBCH 29 | End of tillering: the maximum number of tillers detectable |
| BBCH 37 | Flag leaf just visible, still rolled |
| BBCH 51 | Beginning of heading: the tip of inflorescence emerged from the sheath, the first spikelet just visible |
| BBCH 69 | End of flowering: all spikelets have completed flowering but some dehydrated anthers may remain |
| BBCH 77 | Late milk stage |
| BBCH 83 | Ripening, early dough |
| Factors | Variables | |||||
|---|---|---|---|---|---|---|
| Genotypes | CO2 Levels | Watering | BM (g) | GY (g) | TKW (g) | HI (%) |
| Mv Pálma | NC | C | 67.4 | 26.06 | 36.87 | 38.57 |
| NC | D | 62.0 | 23.37 | 41.57 | 37.54 | |
| EC | C | 73.6 | 30.47 | 35.63 | 41.95 | |
| EC | D | 65.3 | 25.65 | 40.53 | 39.62 | |
| Mv Karéj | NC | C | 73.8 | 27.47 | 43.00 | 37.18 |
| NC | D | 66.4 | 24.45 | 42.60 | 37.05 | |
| EC | C | 75.6 | 28.44 | 41.47 | 37.90 | |
| EC | D | 67.7 | 25.83 | 40.52 | 38.15 | |
| Mv Karizma | NC | C | 77.4 | 32.12 | 41.77 | 41.76 |
| NC | D | 70.6 | 28.96 | 37.87 | 41.17 | |
| EC | C | 79.4 | 33.70 | 41.46 | 42.71 | |
| EC | D | 75.9 | 32.20 | 35.11 | 42.64 | |
| HSD5% values | Watering | 1.817 | 1.223 | n.s. | n.s. | |
| CO2 | n.s. | 1.079 | 6.42 | 7.69 | ||
| Genotype | 1.198 | 0.998 | 6.59 | 8.08 | ||
| 400 ppm | 750 ppm | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C30 | D30 | C60 | D60 | C90 | D90 | C30 | D30 | C60 | D60 | C90 | D90 | |
| BBCH 17 | 362a4 | 160b5 | n.r. | n.r. | n.r. | n.r. | 503a4 | 518a3 | n.r. | n.r. | n.r. | n.r. |
| BBCH 21 | 884a3 | 897a4 | 262a4 | 276a4 | n.r. | n.r. | 1133a3 | 851b2 | 634a4 | 656a2 | n.r. | n.r. |
| BBCH 29 | 1215b2 | 1516a1 | 858b3 | 1225a23 | 457a4 | 666a3 | 1380a2 | 596b3 | 933a23 | 1021a1 | 1029b3 | 1180a4 |
| BBCH 37 | 1249b2 | 1532a1 | 1385a1 | 1452a1 | 951b3 | 2228a2 | 1778a1 | 1063b1 | 1185a1 | 1079a1 | 1947a1 | 1952a1 |
| BBCH 51 | 1276b1 | 1452a1 | 1537a1 | 1492a1 | 1194b2 | 3190a1 | 1649a1 | 1097b1 | 1231a1 | 1146a1 | 1871a1,2 | 1843a1,2 |
| BBCH 69 | 1253a2 | 1242a2,3 | 1234a2 | 1337a1 | 1540b1 | 2877a1 | 1477a2 | 1087b1 | 1079a1,2 | 1027a1 | 1852a1,2 | 1548b3 |
| BBCH 77 | 1506a1 | 1137b3 | 927a3 | 1034a3 | 1008b2,3 | 2730a1 | 1334a2 | 876b2 | 821a3 | 674b2 | 1646a2 | 1637a2,3 |
| BBCH 83 | 1293a2 | 1439a1,2 | 1209b2 | 1387a1,2 | 1340b1,2 | 3200a1 | 1026a3 | 991a1,2 | 862a3 | 995a1 | 1887a1,2 | 1882a1,2 |
| 400 ppm | 750 ppm | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C30 | D30 | C60 | D60 | C90 | D90 | C30 | D30 | C60 | D60 | C90 | D90 | |
| BBCH 17 | 357a4 | 412a4 | n.r. | n.r. | n.r. | n.r. | 60b6 | 208a4 | n.r. | 0 | 0 | 0 |
| BBCH 21 | 1450a3 | 1425a2 | 288a3 | 237a3 | n.r. | n.r. | 588b5 | 1060a3 | 544a3 | 208b4 | 0 | 0 |
| BBCH 29 | 1982a1,2 | 1818a1 | 982b2 | 1280a1,2 | 470a3 | 785a3 | 1240a4 | 1257a2,3 | 1121a2 | 472b3 | 750a3 | 559b3 |
| BBCH 37 | 2111a1 | 1890a1 | 1476a1 | 1375a1 | 2326b2 | 2832a2 | 1539a1 | 1550a1 | 1398a1 | 746b1,2 | 2341a1,2 | 1562b1,2 |
| BBCH 51 | 2167a1 | 1582b2 | 1659a1 | 1474a1 | 3205b1 | 3804a1 | 1657a1 | 1548a1 | 1396a1 | 862b1 | 2703a1 | 1551b1,2 |
| BBCH 69 | 1870a2 | 1575a2 | 1484a1 | 1334a1,2 | 2867b1 | 3747a1 | 1551a1,2 | 1338b1 | 1242a1,2 | 809b1,2 | 2387a1,2 | 1664b1 |
| BBCH 77 | 1578a3 | 1153b3 | 1122a2 | 1122a2 | 2454b2 | 3480a1 | 1411a2,3 | 1298a2 | 1041a2 | 673b2 | 2038a2 | 1361b2 |
| BBCH 83 | 2173a1 | 1933a1 | 1623a1 | 1466a1 | 3232b1 | 3802a1 | 1382a3 | 969a3 | 1117a2 | 662b2 | 2404a1,2 | 1716b1 |
| 400 ppm | 750 ppm | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C30 | D30 | C60 | D60 | C90 | D90 | C30 | D30 | C60 | D60 | C90 | D90 | |
| BBCH 17 | 375a4 | 156b5 | n.r. | n.r. | n.r. | n.r. | 200a4 | 338b4 | n.r. | n.r. | n.r. | n.r. |
| BBCH 21 | 2013a3 | 1057b4 | 114b4 | 153a4 | n.r. | n.r. | 641a3 | 956b3 | 294b5 | 525a4 | n.r. | n.r. |
| BBCH 29 | 2729a1,2 | 1711b1,2 | 561b3 | 1843a3 | 528a4 | 195b4 | 1170a2 | 1067a2,3 | 742b4 | 1355a3 | 684b3 | 568a3 |
| BBCH 37 | 2820a1 | 1829b1,2 | 810b2 | 2356a1,2 | 2847a2 | 3251a3 | 1381a1 | 1322a1 | 1366b2,3 | 2041a1 | 1997b2 | 3345a1 |
| BBCH 51 | 2931a1 | 1853b1 | 1142b1 | 2560a1 | 3469b1 | 4411a1 | 1338a1,2 | 1181a1,2 | 1592b2 | 2188a1 | 2483b2 | 3242a1 |
| BBCH 69 | 2470a2 | 1609b2 | 1156b1 | 2086a2,3 | 2433b3 | 3744a2 | 1244a1,2 | 1155a1,2 | 1432b2,3 | 1660a2 | 2407b2 | 3178a1 |
| BBCH 77 | 2128a3 | 1476b2,3 | 991b1,2 | 2012a2,3 | 2629b2,3 | 3481a3 | 1180a1,2 | 1013a2 | 1271b3 | 1665a2 | 2196b2 | 2535a2 |
| BBCH 83 | 1963a3 | 1766b1,2 | 1182b1 | 2474a1 | 3365b1 | 4468a1 | 1342a1,2 | 1031b2 | 1845a1 | 1792a2 | 2864a1 | 3050a1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varga, B.; Farkas, Z.; Varga-László, E.; Vida, G.; Veisz, O. Elevated Atmospheric CO2 Concentration Influences the Rooting Habits of Winter-Wheat (Triticum aestivum L.) Varieties. Sustainability 2022, 14, 3304. https://doi.org/10.3390/su14063304
Varga B, Farkas Z, Varga-László E, Vida G, Veisz O. Elevated Atmospheric CO2 Concentration Influences the Rooting Habits of Winter-Wheat (Triticum aestivum L.) Varieties. Sustainability. 2022; 14(6):3304. https://doi.org/10.3390/su14063304
Chicago/Turabian StyleVarga, Balázs, Zsuzsanna Farkas, Emese Varga-László, Gyula Vida, and Ottó Veisz. 2022. "Elevated Atmospheric CO2 Concentration Influences the Rooting Habits of Winter-Wheat (Triticum aestivum L.) Varieties" Sustainability 14, no. 6: 3304. https://doi.org/10.3390/su14063304
APA StyleVarga, B., Farkas, Z., Varga-László, E., Vida, G., & Veisz, O. (2022). Elevated Atmospheric CO2 Concentration Influences the Rooting Habits of Winter-Wheat (Triticum aestivum L.) Varieties. Sustainability, 14(6), 3304. https://doi.org/10.3390/su14063304