
Effect of Urban River Morphology on the Structure of Macroinvertebrate Communities in a Subtropical Urban River

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
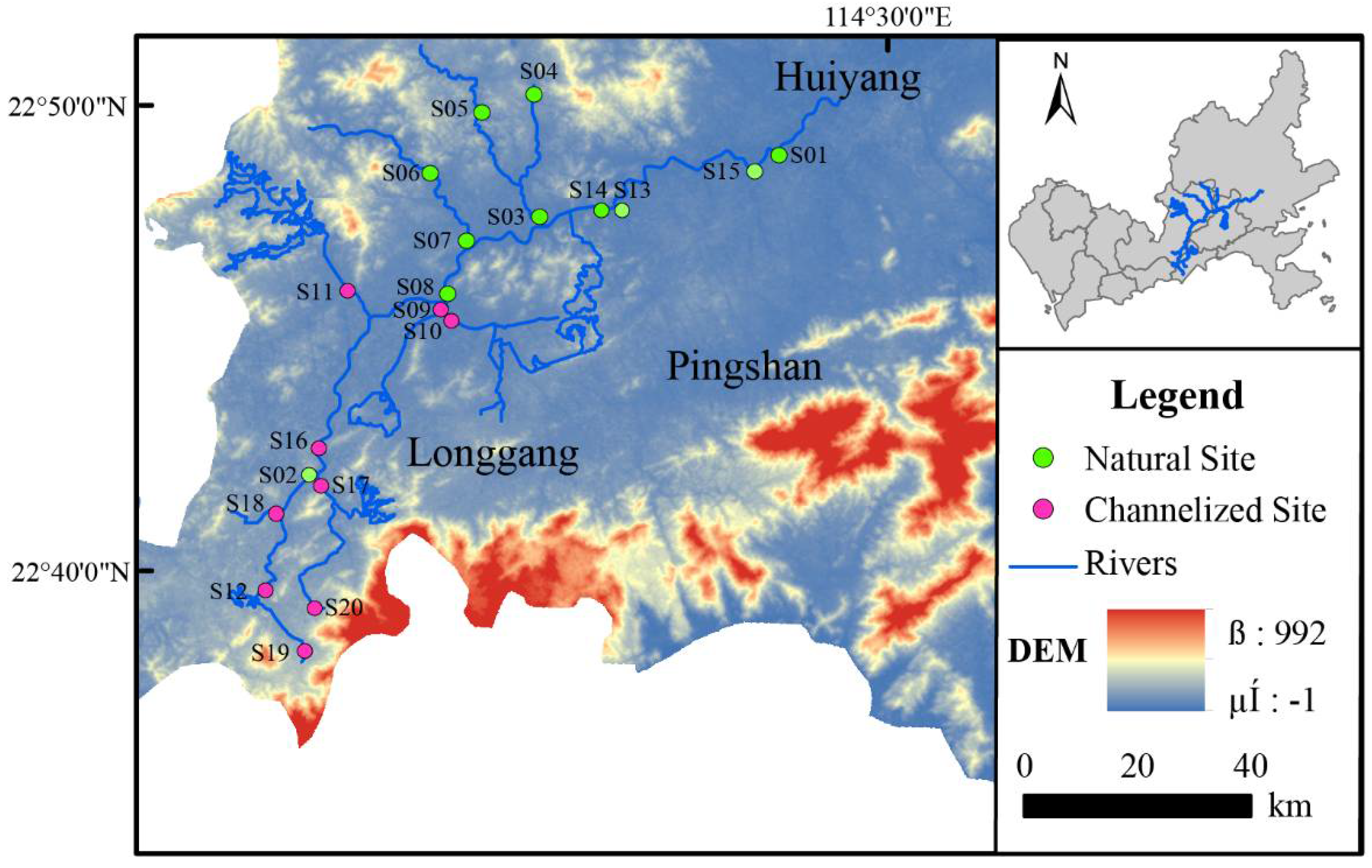
2.1. Study Area
2.2. Study Site and Sampling Design
2.3. Data Analysis
3. Results
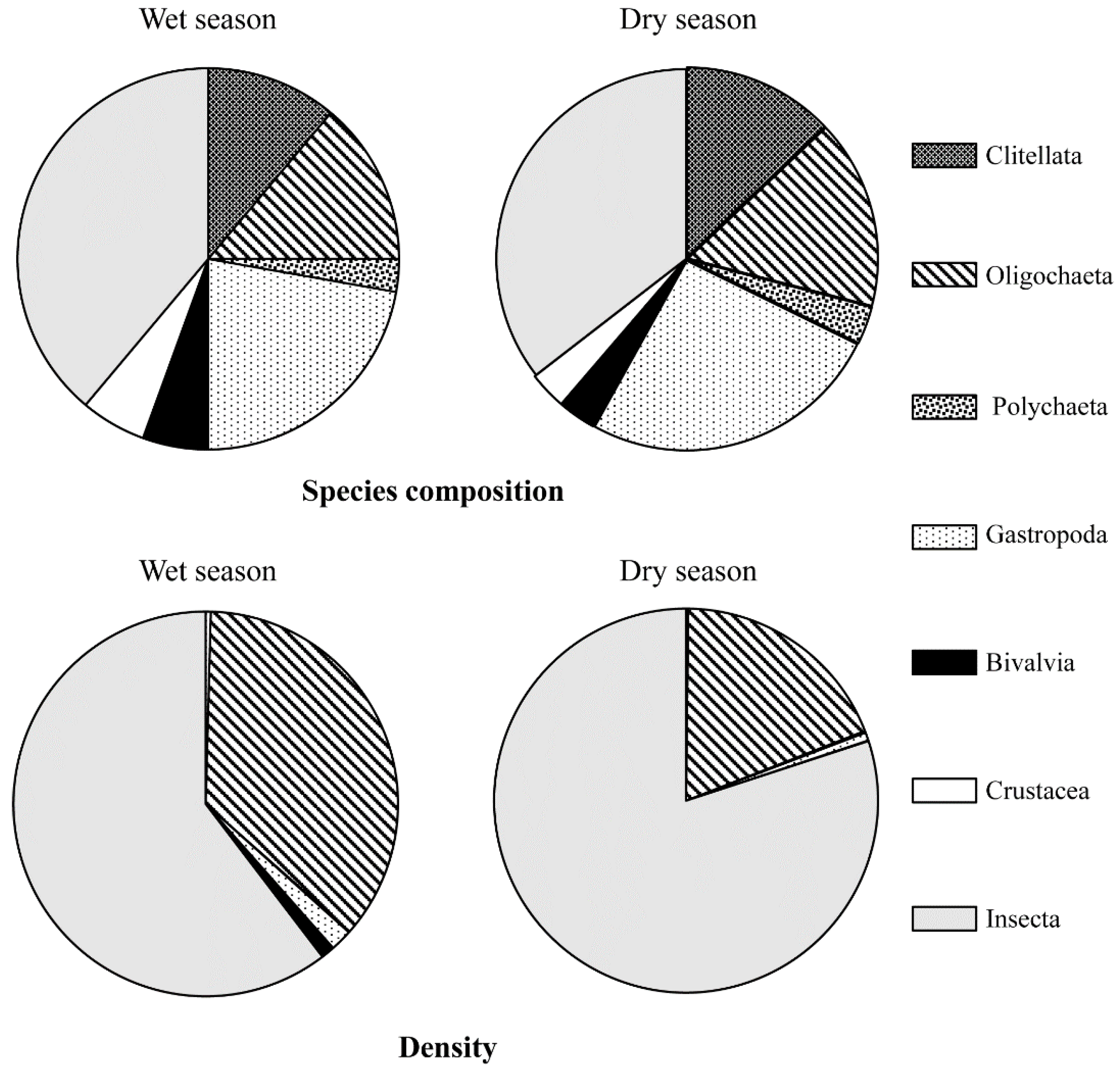
3.1. The Benthic Community in the Wet and Dry Season in the Longgang River
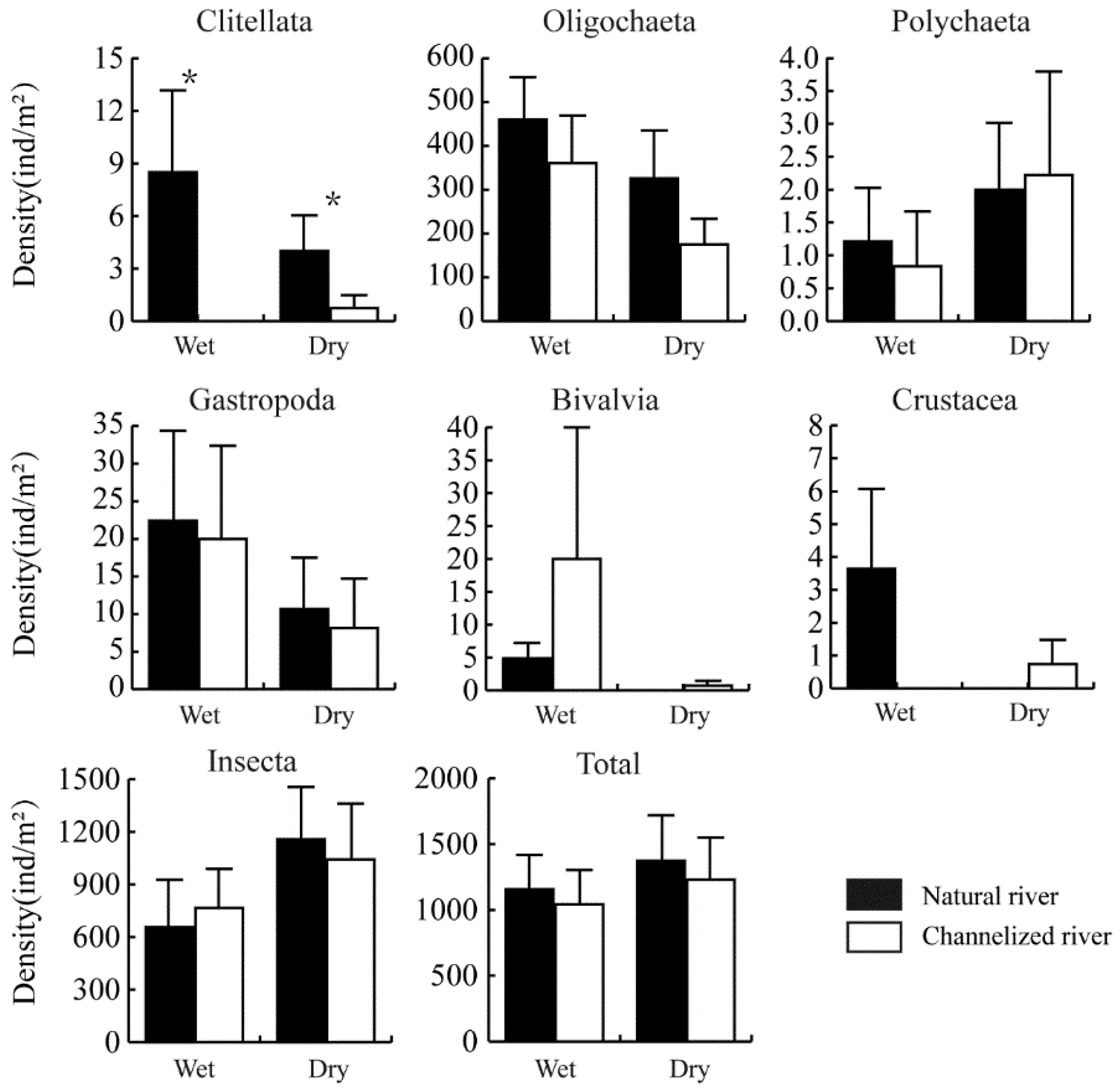
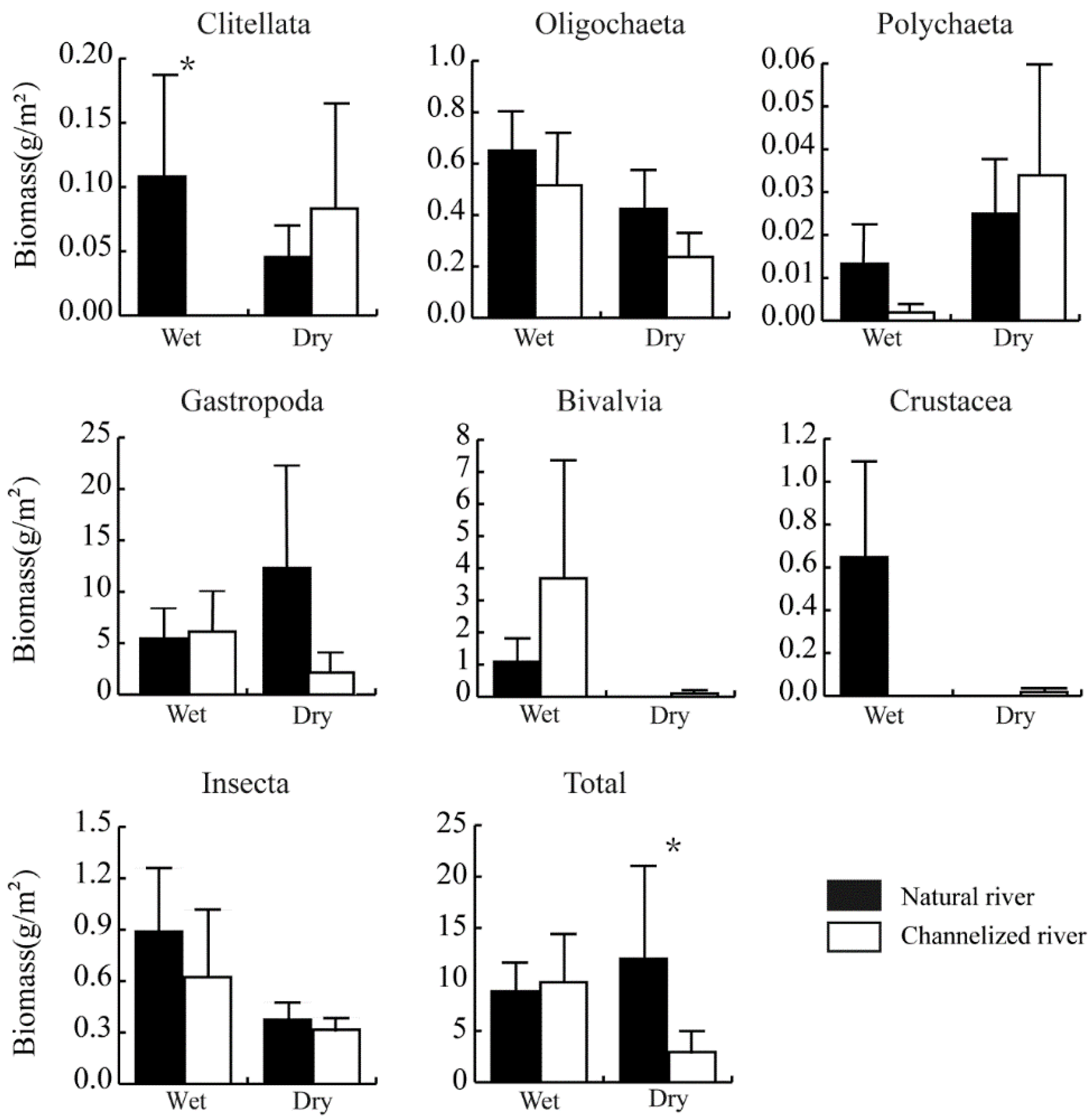
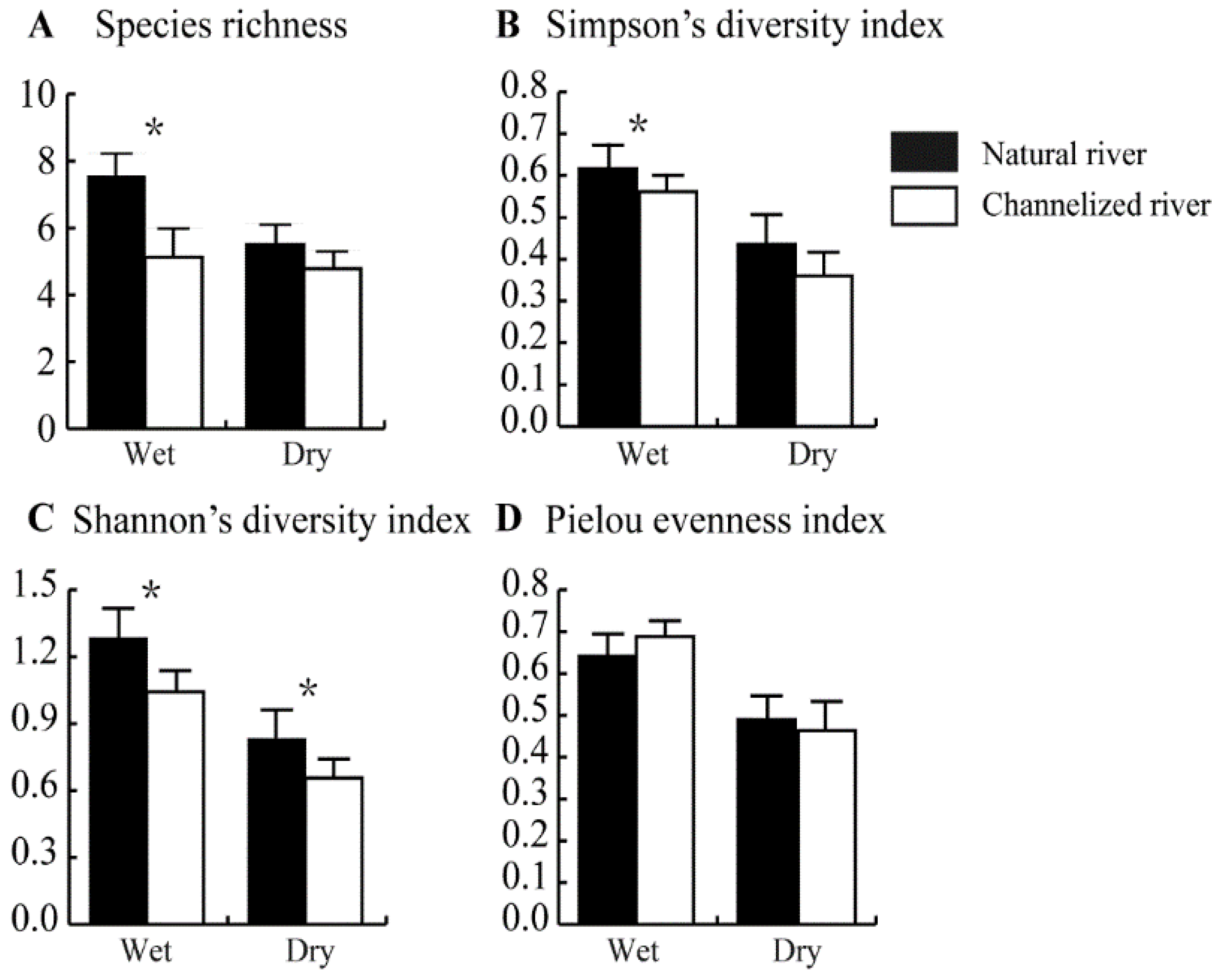
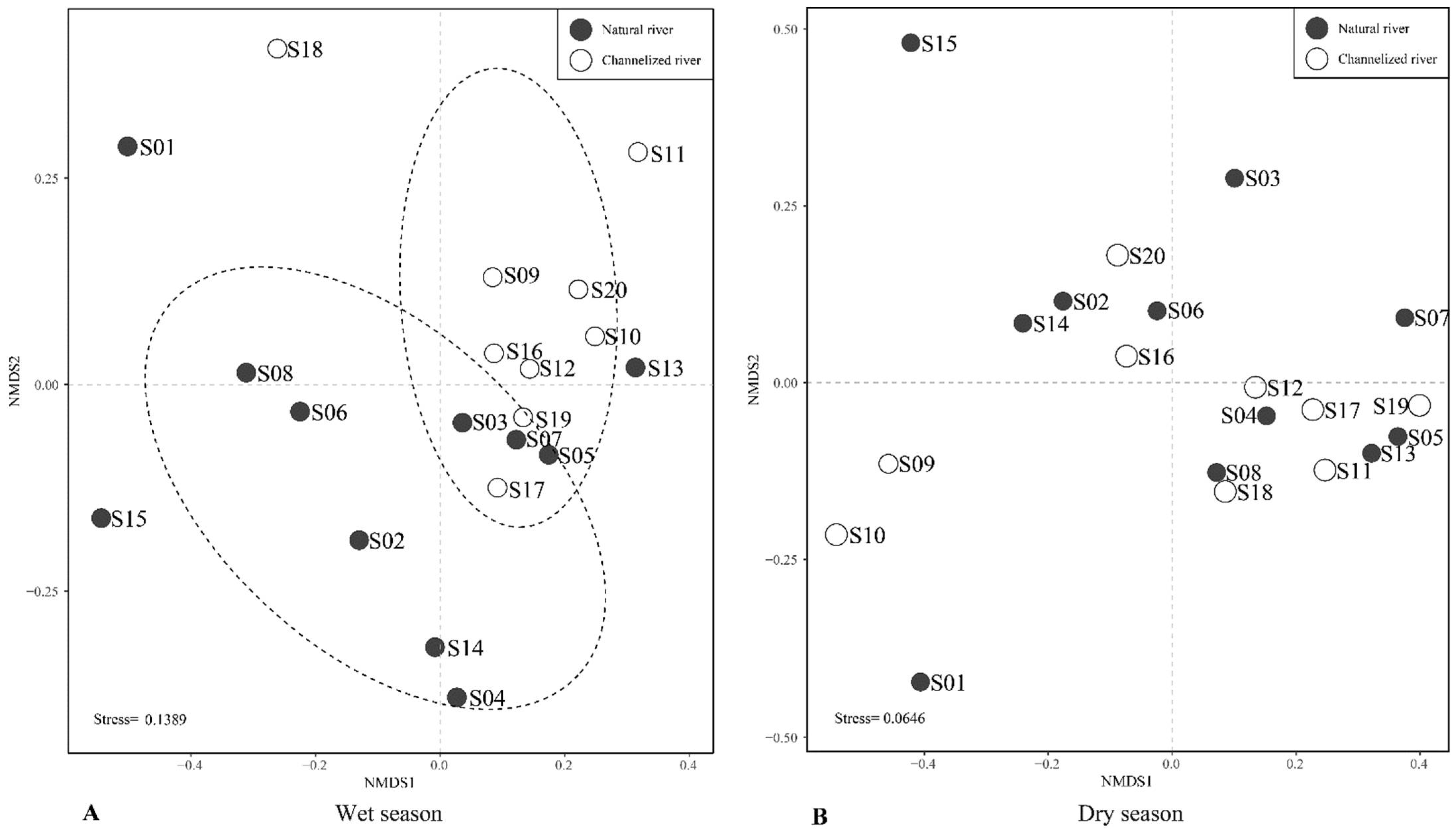
3.2. Differences in the Benthic Community between Natural and Channelized Rivers
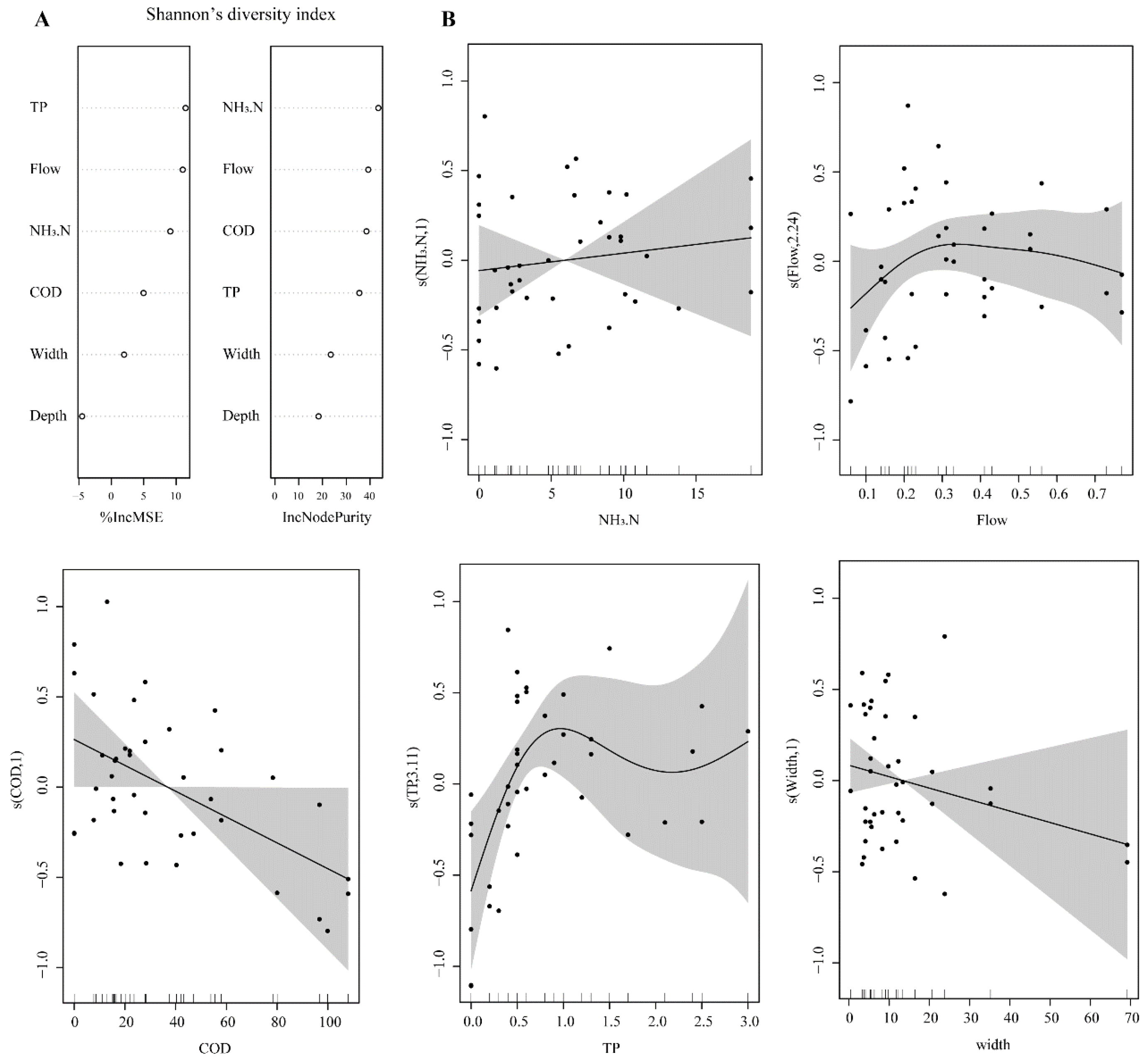
3.3. Correlations between the Diversity of Benthic Communities and Environmental Parameters
4. Discussion
4.1. The Benthic Community of the Longgang River, a Tropical Urban River
4.2. Effects of Urban River Morphology on the Benthic Community
4.3. Implications for Urban River Management
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wet Season | Dry Season | ||||
|---|---|---|---|---|---|
| Natural Area | Channelized Area | Natural Area | Channelized Area | ||
| Clitellata | Glossiphonia complanata | 0.61 ± 0.61 | 0.00 ± 0.00 | 0.61 ± 0.61 | 0.74 ± 0.74 |
| Helobdella stagnalis | 1.21 ± 0.81 | 0.00 ± 0.00 | 1.21 ± 1.21 | 0.00 ± 0.00 | |
| Glossiphonia lata | 1.82 ± 0.94 | 0.00 ± 0.00 | 0.61 ± 0.61 | 0.00 ± 0.00 | |
| Erpobdella octoculata | 4.85 ± 4.23 | 0.00 ± 0.00 | 1.21 ± 0.81 | 0.00 ± 0.00 | |
| Oligochaeta | Rhyacodrilus sinicus | 6.06 ± 3.86 | 2.22 ± 2.22 | 3.03 ± 2.08 | 0.00 ± 0.00 |
| Limnodrilus hoffmeisteri | 387.88 ± 91.38 | 237.78 ± 84.96 | 258.79 ± 83.89 | 154.07 ± 50.97 | |
| Branchiura sowerbyi | 60.00 ± 29.23 | 80.74 ± 38.27 | 41.82 ± 20.60 | 20.74 ± 9.46 | |
| Limnodrilus grandisetosus | 1.21 ± 1.21 | 2.96 ± 2.96 | 1.21 ± 1.21 | 0.00 ± 0.00 | |
| Teneridrilus mastix | 4.85 ± 3.72 | 1.48 ± 1.48 | 1.21 ± 1.21 | 0.74 ± 0.74 | |
| Polychaeta | Nephtys oligobranchia | 1.21 ± 0.81 | 0.74 ± 0.74 | 1.82 ± 0.94 | 2.22 ± 1.57 |
| Gastropoda | Semisulcospira libertina | 1.82 ± 1.82 | 0.74 ± 0.74 | 1.21 ± 1.21 | 4.44 ± 4.44 |
| Pomacea canaliculata | 0.61 ± 0.61 | 0.00 ± 0.00 | 1.22 ± 0.61 | 0.00 ± 0.00 | |
| Parafossarulus eximius | 3.64 ± 3.64 | 8.89 ± 8.89 | 0.00 ± 0.00 | 0.74 ± 0.74 | |
| Bellamya purificata | 0.61 ± 0.61 | 2.22 ± 2.22 | 1.21 ± 0.81 | 0.00 ± 0.00 | |
| Melanoides tuberculata | 13.94 ± 8.74 | 3.70 ± 3.70 | 6.06 ± 5.43 | 1.48 ± 1.48 | |
| Bellamya aeruginosa | 0.61 ± 0.61 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Gyraulus convexiusculus | 0.61 ± 0.61 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Radix swinhoei | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.74 ± 0.74 | |
| Tarebia granifera | 0.61 ± 0.61 | 2.22 ± 1.57 | 0.00 ± 0.00 | 0.74 ± 0.74 | |
| Bivalvia | Corbicula fluminea | 2.42 ± 1.86 | 8.15 ± 8.15 | 0.00 ± 0.00 | 0.74 ± 0.74 |
| Limnoperna fortunei | 2.42 ± 1.86 | 9.63 ± 9.63 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Crustacea | Exopalaemon modestus | 3.03 ± 2.08 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.74 ± 0.74 |
| Palaemonetes sinensis | 0.61 ± 0.61 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Insecta | Orthocladius rivulorum | 33.33 ± 14.13 | 0.00 ± 0.00 | 16.36 ± 11.06 | 0.00 ± 0.00 |
| Propsilocerus akamusi | 41.82 ± 21.98 | 45.93 ± 32.35 | 12.12 ± 12.12 | 0.00 ± 0.00 | |
| Procladius choreus | 22.42 ± 17.76 | 8.89 ± 8.89 | 0.00 ± 0.00 | 6.67 ± 6.67 | |
| Orthetrum sp. | 1.21 ± 1.21 | 0.00 ± 0.00 | 0.61 ± 0.61 | 0.00 ± 0.00 | |
| Pericoma sp. | 0.61 ± 0.61 | 0.00 ± 0.00 | 0.61 ± 0.61 | 0.74 ± 0.74 | |
| Labrogomphus sp. | 2.42 ± 1.86 | 0.74 ± 0.74 | 0.61 ± 0.61 | 2.96 ± 1.61 | |
| Microchironomus tener | 3.64 ± 3.64 | 24.44 ± 24.44 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Dicrotendipes tritomus | 8.49 ± 7.84 | 80.00 ± 63.75 | 70.91 ± 70.91 | 38.52 ± 27.37 | |
| Baetis sp. | 7.27 ± 4.88 | 0.74 ± 0.74 | 0.61 ± 0.61 | 1.48 ± 0.98 | |
| Copera sp. | 7.88 ± 4.48 | 0.74 ± 0.74 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Chaoborus sp. | 13.33 ± 13.33 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Chironomus plumosus | 465.45 ± 280.53 | 506.67 ± 173.40 | 917.58 ± 277.09 | 991.11 ± 319.84 | |
| Tanypus chinensis | 3.03 ± 2.08 | 13.33 ± 13.33 | 14.55 ± 14.55 | 0.00 ± 0.00 | |
| Glyptotendipes barbipes | 45.45 ± 24.60 | 0.00 ± 0.00 | 17.58 ± 14.05 | 0.00 ± 0.00 | |
| Total | 1156.98 ± 262.04 | 1042.96 ± 260.71 | 1372.73 ± 345.53 | 1229.63 ± 321.10 | |
References
- Chen, M.; Zhang, H.; Liu, W.; Zhang, W. The global pattern of urbanization and economic growth: Evidence from the last three decades. PLoS ONE 2014, 9, e103799. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, Q.; Liu, X.; Zhuang, Y.; Hong, S. Global urbanization research from 1991 to 2009: A systematic research review. Landsc. Urban Plan. 2012, 104, 299–309. [Google Scholar] [CrossRef]
- Zhang, X.Q. The trends, promises and challenges of urbanisation in the world. Habitat. Int. 2016, 54, 241–252. [Google Scholar] [CrossRef]
- Zhang, Z.; Peng, J.; Xu, Z.; Wang, X.; Meersmans, J. Ecosystem services supply and demand response to urbanization: A case study of the Pearl River Delta, China. Ecosyst. Serv. 2021, 49, 101274. [Google Scholar] [CrossRef]
- Booth, D.B.; Roy, A.H.; Smith, B.; Capps, K.A. Global perspectives on the urban stream syndrome. Freshw. Sci. 2016, 35, 412–420. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, W.; Wang, L.; Ding, J.; Li, C.; Wang, B. Consideration of river governance based on the concept of urban spatial resilience. IOP Conf. Ser. Earth Environ. Sci. 2022, 983, 12087. [Google Scholar] [CrossRef]
- Fan, X.; Cui, B.; Zhao, H.; Zhang, Z. The Changes of Wetland Network Pattern Associated with Water Quality in the Pearl River Delta, China. CLEAN–Soil Air Water 2012, 40, 1064–1075. [Google Scholar] [CrossRef]
- Douglas, I.; Kobold, M.; Lawson, N.; Pasche, E.; White, I. Characterisation of Urban Streams and Urban Flooding; CRC Press: Boca Raton, FL, USA, 2007; pp. 41–70. ISBN 0429224346. [Google Scholar]
- Luo, Z.; Shao, Q.; Zuo, Q.; Cui, Y. Impact of land use and urbanization on river water quality and ecology in a dam dominated basin. J. Hydrol. 2020, 584, 124655. [Google Scholar] [CrossRef]
- Qin, H.; Su, Q.; Khu, S.; Tang, N. Water quality changes during rapid urbanization in the Shenzhen River Catchment: An integrated view of socio-economic and infrastructure development. Sustainability 2014, 6, 7433–7451. [Google Scholar] [CrossRef]
- Almeida, C.A.; Quintar, S.; González, P.; Mallea, M.A. Influence of urbanization and tourist activities on the water quality of the Potrero de los Funes River (San Luis–Argentina). Environ. Monit. Assess. 2007, 133, 459–465. [Google Scholar] [CrossRef]
- Luo, K.; Hu, X.; He, Q.; Wu, Z.; Cheng, H.; Hu, Z.; Mazumder, A. Impacts of rapid urbanization on the water quality and macroinvertebrate communities of streams: A case study in Liangjiang New Area, China. Sci. Total Environ. 2018, 621, 1601–1614. [Google Scholar] [CrossRef] [PubMed]
- Mcdonald, R.I.; Weber, K.; Padowski, J.; Flörke, M.; Schneider, C.; Green, P.A.; Gleeson, T.; Eckman, S.; Lehner, B.; Balk, D.; et al. Water on an urban planet: Urbanization and the reach of urban water infrastructure. Glob. Environ. Chang. 2014, 27, 96–105. [Google Scholar] [CrossRef]
- Roy, A.H.; Rosemond, A.D.; Paul, M.J.; Leigh, D.S.; Wallace, J.B. Stream macroinvertebrate response to catchment urbanisation (Georgia, U.S.A.). Freshw. Biol. 2003, 48, 329–346. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Cheng, L.; Cai, Y.; Yin, H.; Gao, J.; Gao, Y. Macroinvertebrate assemblages in streams and rivers of a highly developed region (Lake Taihu Basin, China). Aquat. Biol. 2014, 23, 15–28. [Google Scholar] [CrossRef]
- Purcell, A.H.; Bressler, D.W.; Paul, M.J.; Barbour, M.T.; Rankin, E.T.; Carter, J.L.; Resh, V.H. Assessment tools for urban catchments: Developing biological indicators based on benthic macroinvertebrates 1. JAWRA J. Am. Water Resour. Assoc. 2009, 45, 306–319. [Google Scholar] [CrossRef]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P. The urban stream syndrome: Current knowledge and the search for a cure. J. N. Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Ramírez, A.; Engman, A.; Rosas, K.G.; Perez-Reyes, O.; Martinó-Cardona, D.M. Urban impacts on tropical island streams: Some key aspects influencing ecosystem response. Urban Ecosyst. 2012, 15, 315–325. [Google Scholar] [CrossRef]
- Anderson, M.G.; Steinberg, M.; Tontar, C. A New Urban Revival: Floodplain Acquisition in Lawrence, Massachusetts. In Times Are Changing—Flood Mitigation Technology: Proceedings of the 22nd Annual Conference, 18–22 May 1998; Natural Hazards Research and Applications Information Center, University of Colorado: Boulder, CO, USA, 1998; p. 201. [Google Scholar]
- Nakano, D.; Nagayama, S.; Kawaguchi, Y.; Nakamura, F. River restoration for macroinvertebrate communities in lowland rivers: Insights from restorations of the Shibetsu River, north Japan. Landsc. Ecol. Eng. 2008, 4, 63–68. [Google Scholar] [CrossRef]
- Garcia, X.; Schnauder, I.; Pusch, M.T. Complex hydromorphology of meanders can support benthic invertebrate diversity in rivers. Hydrobiologia 2012, 685, 49–68. [Google Scholar] [CrossRef]
- Zika, U.; Peter, A. The introduction of woody debris into a channelized stream: Effect on trout populations and habitat. River Res. Appl. 2002, 18, 355–366. [Google Scholar] [CrossRef]
- Wang, S.; Wang, T.; Lin, H.; Stewart, S.D.; Cheng, G.; Li, W.; Yang, F.; Huang, W.; Chen, Z.; Xie, S. Impacts of environmental factors on the food web structure, energy flows, and system attributes along a subtropical urban river in southern China. Sci. Total Environ. 2021, 794, 148673. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Wang, H.; Xia, B.; Jiang, C.; Wu, W.; Li, Z.; Li, S.; Su, H.; Bai, Z.; Xu, S. Pollution Characterization and Comprehensive Water Quality Assessment of Rain-source River: A Case Study of the Longgang River in Shenzhen. Huan Jing Ke Xue = Huanjing Kexue 2022, 43, 782–794. [Google Scholar]
- Yang, Y.; Hu, M.; Lu, W.; Xue, L.; Lin, X.; Liu, E. Occurrence, distribution, and risk assessment of PPCPs in water and sediments of Longgang River in Shenzhen City, south China. Desalin. Water Treat. 2020, 189, 196–206. [Google Scholar] [CrossRef]
- Liu, L.; Ma, X. Integrated river basin management in rapidly urbanizing areas: A case of Shenzhen, China. Front. Environ. Sci. Eng. China 2011, 5, 243–254. [Google Scholar] [CrossRef]
- Qian, J.L.; Fu, L.M. Using persulfate oxidation method for the determination of total nitrogen and total phosphorus in water. Environ. Sci. 1987, 8, 81–84. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Gomez-Rubio, V. Generalized Additive Models: An Introduction with R (2nd Edition). J. Stat. Softw. 2018, 86, 1–5. [Google Scholar] [CrossRef]
- Wood, S.N.; Pya, N.; Säfken, B. Smoothing parameter and model selection for general smooth models. J. Am. Stat. Assoc. 2016, 111, 1548–1563. [Google Scholar] [CrossRef]
- Line, D.E. Effect of development on water quality for seven streams in North Carolina. Environ. Monit. Assess. 2013, 185, 6277–6289. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, D.M.; Hawkins, C.P. Land use and the structure of western US stream invertebrate assemblages: Predictive models and ecological traits. J. N. Am. Benthol. Soc. 2008, 27, 986–999. [Google Scholar] [CrossRef]
- Chinnayakanahalli, K.J.; Hawkins, C.P.; Tarboton, D.G.; Hill, R.A. Natural flow regime, temperature and the composition and richness of invertebrate assemblages in streams of the western United States. Freshw. Biol. 2011, 56, 1248–1265. [Google Scholar] [CrossRef]
- Murphy, J.F.; Giller, P.S. Seasonal dynamics of macroinvertebrate assemblages in the benthos and associated with detritus packs in two low-order streams with different riparian vegetation. Freshw. Biol. 2000, 43, 617–631. [Google Scholar] [CrossRef]
- Johnson, R.C.; Carreiro, M.M.; Jin, H.; Jack, J.D. Within-year temporal variation and life-cycle seasonality affect stream macroinvertebrate community structure and biotic metrics. Ecol. Indic. 2012, 13, 206–214. [Google Scholar] [CrossRef]
- Helms, B.S.; Schoonover, J.E.; Feminella, J.W. Seasonal variability of landuse impacts on macroinvertebrate assemblages in streams of western Georgia, USA. J. N. Am. Benthol. Soc. 2009, 28, 991–1006. [Google Scholar] [CrossRef]
- Ortiz, J.D.; Puig, M.A. Point source effects on density, biomass and diversity of benthic macroinvertebrates in a Mediterranean stream. River Res. Appl. 2007, 23, 155–170. [Google Scholar] [CrossRef]
- Shieh, S.; Kondratieff, B.C.; Ward, J.V. Longitudinal changes in benthic organic matter and macroinvertebrates in a polluted Colorado plains stream. Hydrobiologia 1999, 411, 191–209. [Google Scholar] [CrossRef]
- Angradi, T.R.; Bolgrien, D.W.; Jicha, T.M.; Moffett, M.F. Macroinvertebrate assemblage response to urbanization in three mid-continent USA great rivers. Fund. Appl. Limnol. 2010, 176, 183–198. [Google Scholar] [CrossRef]
- Miserendino, M.L.; Masi, C.I. The effects of land use on environmental features and functional organization of macroinvertebrate communities in Patagonian low order streams. Ecol. Indic. 2010, 10, 311–319. [Google Scholar] [CrossRef]
- Gadawski, P.; Riss, H.W.; Płóciennik, M.; Meyer, E.I. City Channel Chironomids-Benthic Diversity in Urban Conditions. River Res. Appl. 2016, 32, 1978–1988. [Google Scholar] [CrossRef]
- Stepenuck, K.F.; Crunkilton, R.L.; Wang, L. Impacts of urban landuse on macroinvertebrate communities in southeastern Wisconsin streams 1. JAWRA J. Am. Water Resour. Assoc. 2002, 38, 1041–1051. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.D.; Lu, J. Urban River Pollution Control and Remediation. Procedia Environ. Sci. 2012, 13, 1856–1862. [Google Scholar] [CrossRef]
- Li, F.; Bae, M.; Kwon, Y.; Chung, N.; Hwang, S.; Park, S.; Park, H.; Kong, D.S.; Park, Y. Ecological exergy as an indicator of land-use impacts on functional guilds in river ecosystems. Ecol. Model. 2013, 252, 53–62. [Google Scholar] [CrossRef]
- Brönmark, C.; Hansson, L. The Biology of Lakes and Ponds; Oxford University Press: Oxford, UK, 2017; ISBN 0191022543. [Google Scholar]
- Usseglio Polatera, P.; Beisel, J.N. Longitudinal changes in macroinvertebrate assemblages in the Meuse River: Anthropogenic effects versus natural change. River Res. Appl. 2002, 18, 197–211. [Google Scholar] [CrossRef]
- Habersack, H.; Hein, T.; Stanica, A.; Liska, I.; Mair, R.; Jäger, E.; Hauer, C.; Bradley, C. Challenges of river basin management: Current status of, and prospects for, the River Danube from a river engineering perspective. Sci. Total Environ. 2016, 543, 828–845. [Google Scholar] [CrossRef]
- Nakano, D.; Nagayama, S.; Kawaguchi, Y.; Nakamura, F. Significance of the stable foundations provided and created by large wood for benthic fauna in the Shibetsu River, Japan. Ecol. Eng. 2018, 120, 249–259. [Google Scholar] [CrossRef]
- Whitcraft, C.R.; Levin, L.A. Regulation of benthic algal and animal communities by salt marsh plants: Impact of shading. Ecology 2007, 88, 904–917. [Google Scholar] [CrossRef]
- Giller, P.S.; Giller, P. The Biology of Streams and Rivers; Oxford University Press: Oxford, UK, 1998; ISBN 0198549776. [Google Scholar]
- Al-Shami, S.A.; Rawi, C.S.M.; Ahmad, A.H.; Hamid, S.A.; Nor, S.A.M. Influence of agricultural, industrial, and anthropogenic stresses on the distribution and diversity of macroinvertebrates in Juru River Basin, Penang, Malaysia. Ecotoxicol. Environ. Saf. 2011, 74, 1195–1202. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, J.; Shi, W.; Wang, M.; Chen, K.; Wang, L. Priority pollutants in water and sediments of a river for control basing on benthic macroinvertebrate community structure. Water 2019, 11, 1267. [Google Scholar] [CrossRef]
- Townsend, C.R.; Scarsbrook, M.R.; Dolédec, S. The intermediate disturbance hypothesis, refugia, and biodiversity in streams. Limnol. Oceanogr. 1997, 42, 938–949. [Google Scholar] [CrossRef]
- Eötvös, C.B.; Lövei, G.L.; Magura, T. Predation pressure on sentinel insect prey along a riverside urbanization gradient in Hungary. Insects 2020, 11, 97. [Google Scholar] [CrossRef]
- Peterson, C.H.; Summerson, H.C.; Thomson, E.; Lenihan, H.S.; Grabowski, J.; Manning, L.; Micheli, F.; Johnson, G. Synthesis of linkages between benthic and fish communities as a key to protecting essential fish habitat. Bull. Mar. Sci. 2000, 66, 759–774. [Google Scholar]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef]
| Wet Density | Dry Density | Z Adjusted | p−Value | ||
|---|---|---|---|---|---|
| Clitellata | Glossiphonia complanata | 0.33 ± 0.33 | 0.67 ± 0.46 | 0.563 | 0.573 |
| Helobdella stagnalis | 0.67 ± 0.46 | 0.67 ± 0.67 | −0.503 | 0.615 | |
| Glossiphonia lata | 1 ± 0.55 | 0.33 ± 0.33 | −1.015 | 0.310 | |
| Erpobdella octoculata | 2.67 ± 2.34 | 0.67 ± 0.46 | −0.026 | 0.979 | |
| Oligochaeta | Rhyacodrilus sinicus | 4.33 ± 2.33 | 1.67 ± 1.17 | −0.893 | 0.372 |
| Limnodrilus hoffmeisteri | 320.33 ± 63.86 | 211.67 ± 51.69 | −1.408 | 0.159 | |
| Branchiura sowerbyi | 69.33 ± 23.03 | 32.33 ± 12.06 | −0.477 | 0.633 | |
| Limnodrilus grandisetosus | 2 ± 1.46 | 0.67 ± 0.67 | −0.592 | 0.554 | |
| Teneridrilus mastix | 3.33 ± 2.14 | 1 ± 0.73 | −0.494 | 0.621 | |
| Polychaeta | Nephtys oligobranchia | 1 ± 0.55 | 2 ± 0.85 | 0.817 | 0.414 |
| Gastropoda | Semisulcospira libertina | 1.33 ± 1.04 | 2.67 ± 2.07 | 0.026 | 0.979 |
| Pomacea canaliculata | 0.33 ± 0.33 | 0.67 ± 0.46 | 0.563 | 0.573 | |
| Parafossarulus eximius | 6 ± 4.38 | 0.33 ± 0.33 | −0.622 | 0.534 | |
| Bellamya purificata | 1.33 ± 1.04 | 0.67 ± 0.46 | −0.026 | 0.979 | |
| Melanoides tuberculata | 9.33 ± 5.1 | 4 ± 3.04 | −0.813 | 0.416 | |
| Bellamya aeruginosa | 0.33 ± 0.33 | 0 ± 0 | −0.950 | 0.342 | |
| Gyraulus convexiusculus | 0.33 ± 0.33 | 0 ± 0 | −0.950 | 0.342 | |
| Radix swinhoei | 0 ± 0 | 0.33 ± 0.33 | 0.950 | 0.342 | |
| Tarebia granifera | 1.33 ± 0.78 | 0.33 ± 0.33 | −1.040 | 0.298 | |
| Bivalvia | Corbicula fluminea | 5 ± 3.74 | 0.33 ± 0.33 | −1.065 | 0.287 |
| Limnoperna fortunei | 5.67 ± 4.39 | 0 ± 0 | −1.747 | 0.081 | |
| Crustacea | Exopalaemon modestus | 1.67 ± 1.17 | 0.33 ± 0.33 | −0.622 | 0.534 |
| Palaemonetes sinensis | 0.33 ± 0.33 | 0 ± 0 | −0.950 | 0.342 | |
| Insecta | Orthocladius rivulorum | 18.33 ± 8.5 | 9 ± 6.24 | −0.871 | 0.384 |
| Propsilocerus akamusi | 43.67 ± 18.39 | 6.67 ± 6.67 | −1.785 | 0.074 | |
| Procladius choreus | 16.33 ± 10.42 | 3 ± 3 | −1.039 | 0.299 | |
| Orthetrum sp. | 0.67 ± 0.67 | 0.33 ± 0.33 | 0.000 | 1.000 | |
| Pericoma sp. | 0.33 ± 0.33 | 0.67 ± 0.46 | 0.563 | 0.573 | |
| Labrogomphus sp. | 1.67 ± 1.07 | 1.67 ± 0.82 | 0.307 | 0.759 | |
| Microchironomus tener | 13 ± 11.08 | 0 ± 0 | −1.396 | 0.163 | |
| Dicrotendipes tritomus | 40.67 ± 29.23 | 56.33 ± 40.14 | −0.265 | 0.791 | |
| Baetis sp. | 4.33 ± 2.75 | 1 ± 0.55 | −0.131 | 0.896 | |
| Copera sp. | 4.67 ± 2.56 | 0 ± 0 | −2.052 | 0.040 | |
| Chaoborus sp. | 7.33 ± 7.33 | 0 ± 0 | −0.950 | 0.342 | |
| Chironomus plumosus | 484 ± 168.82 | 950.67 ± 204.15 | 2.027 | 0.043 | |
| Tanypus chinensis | 7.67 ± 6.03 | 8 ± 8 | −0.935 | 0.350 | |
| Glyptotendipes barbipes | 25 ± 14.22 | 9.67 ± 7.82 | −0.518 | 0.605 | |
| Total denstiy | 1105.67 ± 181.45 | 1308.34 ± 233.16 | 0.473 | 0.636 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, Q.; Xu, W.; Chen, L.; Wang, L.; Wang, Y.; Liu, Y.; Xie, L. Effect of Urban River Morphology on the Structure of Macroinvertebrate Communities in a Subtropical Urban River. Sustainability 2022, 14, 10046. https://doi.org/10.3390/su141610046
Sheng Q, Xu W, Chen L, Wang L, Wang Y, Liu Y, Xie L. Effect of Urban River Morphology on the Structure of Macroinvertebrate Communities in a Subtropical Urban River. Sustainability. 2022; 14(16):10046. https://doi.org/10.3390/su141610046
Chicago/Turabian StyleSheng, Qiang, Wang Xu, Long Chen, Lei Wang, Yudong Wang, Yihong Liu, and Linshen Xie. 2022. "Effect of Urban River Morphology on the Structure of Macroinvertebrate Communities in a Subtropical Urban River" Sustainability 14, no. 16: 10046. https://doi.org/10.3390/su141610046
APA StyleSheng, Q., Xu, W., Chen, L., Wang, L., Wang, Y., Liu, Y., & Xie, L. (2022). Effect of Urban River Morphology on the Structure of Macroinvertebrate Communities in a Subtropical Urban River. Sustainability, 14(16), 10046. https://doi.org/10.3390/su141610046