
Physiological Response and Comprehensive Resistance Evaluation of East African Endemic Aeollanthus repens under Water and Heat Stress

Abstract
1. Introduction
2. Results
2.1. Effects of High Temperature, Moisture, and Combined Stress on the Photosynthesis and Morphology of A. repens
2.2. Effects of High-Temperature and Water Stress on A. repens’ Cell Membranes
2.3. Effect of High-Temperature and Water Stress on Antioxidant Enzyme Activities of A. repens
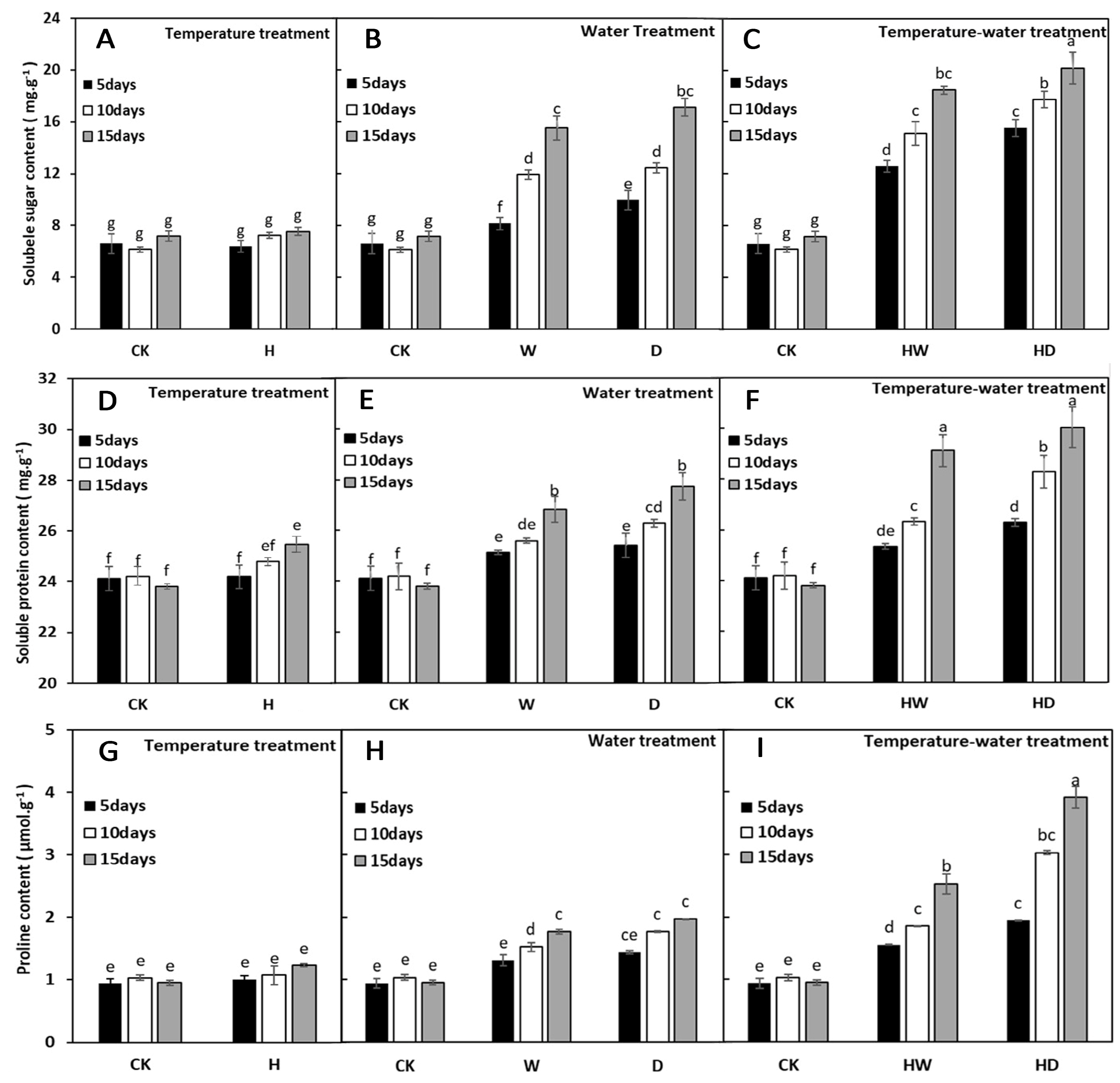
2.4. Effects of High-Temperature, Water, and Combined Stress on the Osmotic Regulation of A. repens
2.5. Comprehensive Evaluation of the Tolerance of A. repens to Different Growing Environments
2.6. Correlation Analysis
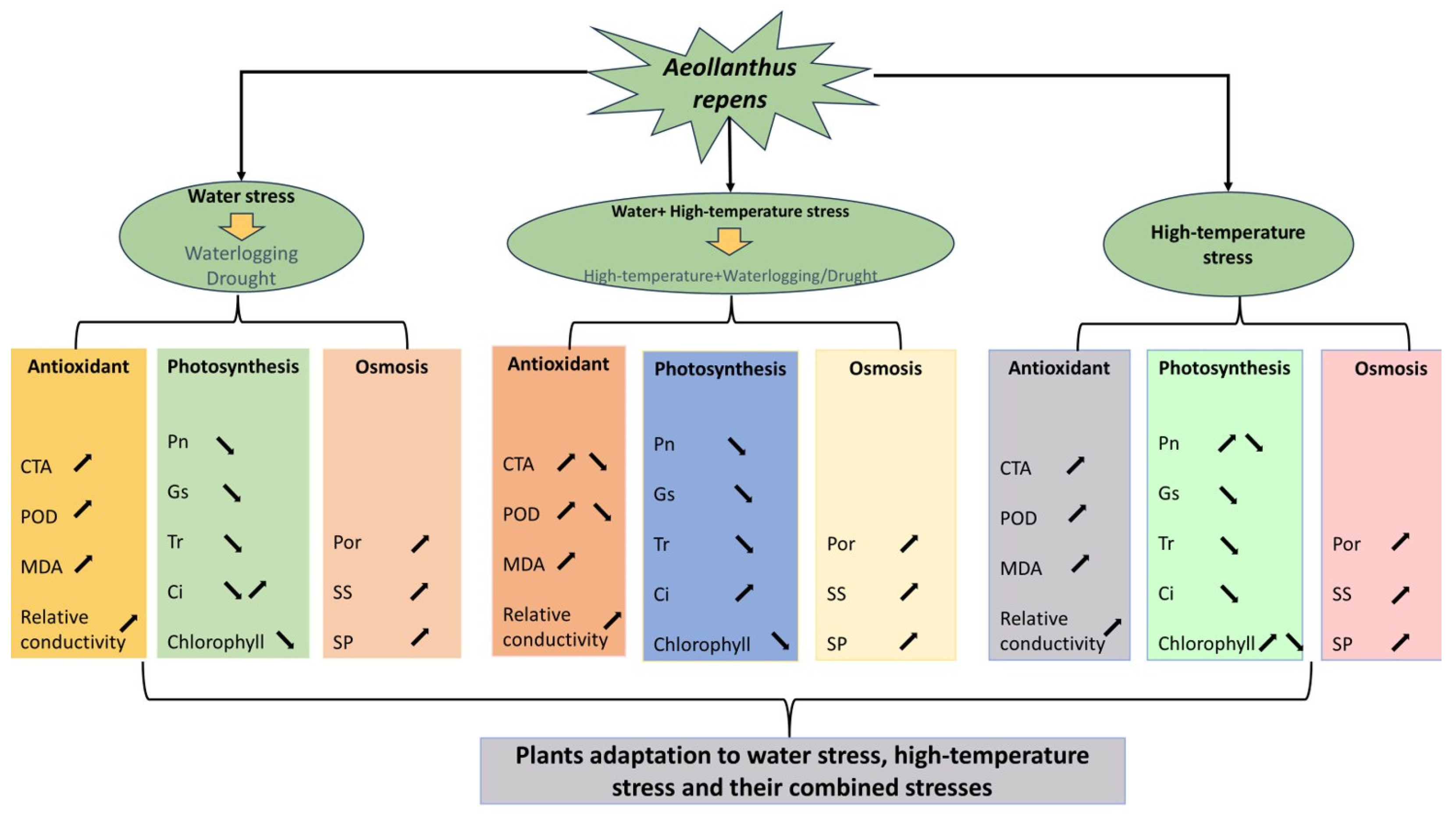
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Experimental Treatments
4.3. Measurement Indicators and Methods
4.3.1. Measurement of Indicators of the Photosynthetic Properties
4.3.2. Measurement of the Physiological Indicators
4.3.3. Comprehensive Evaluation of Resistance to High-Temperature Waterlogging Stress Conditions
4.4. Data Analysis
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.; Ding, Y.Y.; Li, Y.Y.; Guo, D.Y. Analysis of syatus qou and problems of water conservation in China in new situation. South North Water Transf. Water Sci. Technol. 2019, 17, 202–208. [Google Scholar]
- Fawzy, S.; Osman, I.A.; Doran, J.; David, W.R. Strategies for mitigation of climate change: A review. Environ. Chem. Lett. 2020, 18, 1–26. [Google Scholar] [CrossRef]
- van Veen, H.; Mustroph, A.; Barding, G.A.; Vergeer-van Eijk, M.; Welschen-Evertman, R.A.; Pedersen, O.; Sasidharan, R. Two Rumex species from contrasting hydrological niches regulate flooding tolerance through distinct mechanisms. Plant Cell 2013, 25, 4691–4707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yao, Y.B.; Wang, Y. Risk characteristics control technology measures of drought disaster in Southern China. Acta Ecol. Sin. 2017, 37, 7206–7218. [Google Scholar]
- Ji, R.X.; Yu, X.; Chang, Y.; Shen, C.; Bai, X.K.; Xia, X.L.; Yi, W.L. Geographical provenance variation of leaf anatomical structure of Caryopteris mongholica and its significance in response to environmental changes. Chin. J. Plant Ecol. 2020, 44, 277–286. [Google Scholar] [CrossRef]
- Webber, H. Canopy temperature for simulation of heat stress in irrigated wheat in a semi-arid environment: A multi-model comparison. Field Crop. Res. 2017, 202, 21–35. [Google Scholar] [CrossRef]
- Ahamed, G.J.; Li, X.; Wan, H.J.; Zhou, G.; Cheng, Y. SlWRKY81 reduces drought tolerance by attenuating proline biosynthesis in tomato. Sci. Hortic. 2020, 270, 109444. [Google Scholar] [CrossRef]
- Xu, L.X.; Tian, R.N. Advances in Research on Ornamental Plant Heat Tolerance. Subtrop. Plant Sci. 2019, 48, 96–102. [Google Scholar]
- Prasad, P.V.; Pisipati, S.R.; Momcilovic, I.; Ristic, Z. Independent and combined effects of high temperature and drought stress during grain filling on plant yield and chloroplast EF-Tu expression in spring wheat. J. Agron. Crop Sci. 2011, 197, 430–441. [Google Scholar] [CrossRef]
- Singh, R.; Singh, S.; Parihar, P.; Mishra, R.K.; Prasad, S.M. Reactive oxygen species(ROS): Beneficial companions of plants developmental processes. Front. Plant Sci. 2016, 7, 1299. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Hu, W.; Snider, J.L.; Zhou, Z. Short-term soil-waterlogging contributes to cotton cross tolerance to chronic elevated temperature by influencing ROS metabolism in the subtending leaf. Plant Physiol. Biochem. 2019, 139, 64–78. [Google Scholar] [CrossRef] [PubMed]
- Hossain, Z.; López-Climent, M.F.; Arbona, V.; Pérez-Clemente, R.M.; Gómez-Cadenas, A. Modulation of the antioxidant system in citrus under waterlogging and subsequent drainage. J. Plant Physiol. 2009, 166, 1391–1404. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dong, K.H. Study on the physological characteristics of Bothriochloa ischaemum under drought stress and re-watering treatments. Grassl. Turf. 2011, 31, 74–78. [Google Scholar]
- Mi, B.B.; Wu, F.F.; Xie, L.L.; Dai, X.Z.; Zhou, H.Q. Differences in morphological structure and physiological response of wax gourd seedlings under waterlogging stress. Nanfang Nongye Xuebao (J. South. Agric.) 2018, 49, 2419–2424. [Google Scholar]
- Suzuki, N.; Bassil, E.; Hamilton, J.S.; Inupakutika, M.A.; Zandalinas, S.I.; Tripathy, D.; Luo, Y.; Dion, E.; Fukui, G.; Kumazaki, A.; et al. ABA is required for plant acclimation to a combination of salt and heat stress. PLoS ONE 2016, 11, e0147625. [Google Scholar] [CrossRef] [PubMed]
- Fang, M.Y.; Wang, Y.P.; Zhao, Y.; Reziya, A.; Shen, Y.H.; Zhang, T.J. Morphological and physiolobical responses of eight alfalfa cultivars to combined stress of cold and drought. Acta Agrestia Sin. 2022, 30, 2967–2974. [Google Scholar]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Wang, Q.L.; Zhou, G.S.; Ma, X.Y. Responses of summer maize leaf water content and photosynthetic characteristics to consecutive drought with different intensities. Chin. J. Ecol. 2015, 34, 3111–3117. [Google Scholar]
- Tian, L.; Xie, X.Y.; Bao, Y.X.; Hu, J.C.; Yu, L.; Wang, J.X. Effect of moisture stress on photosynthetic characteristics of summer maize leaf during growth stages. Chin. J. Agrometeorol. 2013, 34, 655–660. [Google Scholar]
- Yu, S.Y.; Zhou, Y.F.; Hang, W.; Wang, X.; Gao, X.Q.; Fu, B.Z. Effect of drought, salt, and drought-salt combined stress on photosynthetic and physiological characteristics of Agropyron mongolicum. Acta Agrestia Sin. 2021, 29, 2399–2406. [Google Scholar]
- Ni, X.; Zhou, B.Z.; Cao, Y.H.; Lu, X.Z. Impact of drought stress on plant photosynthetic physiology: A review. J. Jiangsu For. Sci. Technol. 2017, 44, 34–39+52. [Google Scholar]
- Gu, X.B.; Xue, L.; Lu, L.H.; Xiao, J.P.; Song, G.H.; Xie, M.; Zhang, H.Q. Melatonin enhances the waterlogging tolerance of Prunus persica by modulating antioxidant metabolism and anaerobic respiration. J. Plant Growth Regul. 2020, 40, 2178–2190. [Google Scholar] [CrossRef]
- Rahmati, M.; Miras-Avalos, J.M.; Valsesia, P.; Lescourret, F.; Genard, M.; Davarynejad, G.H.; Bannayan, M.; Azizi, M.; Vercambre, G. Disentangling the effects of water stress on carbon acquisition, vegetative growth, and fruit quality of peach trees by means of the qualitree model. Front. Plant Sci. 2018, 9, 3. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.Q.; Wang, C.Y.; Song, H.D.; Bai, Y.X.; Zhang, H.X.; Hu, Z.H.; Wang, L.; Shi, K.; Zheng, X.D.; Yan, T.C.; et al. The MdMEK2-MdMPK6-MdWRKY17 pathway stabilizes chlorophyll levels by directly regulating MdSUFB in apple under drought stress. Plant 2021, 108, 814–828. [Google Scholar] [CrossRef]
- Zhu, J.X.; Wu, Y.B.; Liu, X. The combined stress of elevated temperature and drought on the leaf economy spectrum traits of poplar seedlings. J. Fujian Agric. For. Univ. (Nutural Sci. Ed.) 2018, 47, 554–560. [Google Scholar]
- Zhang, Z.M.; Dai, L.X.; Song, W.W.; Ding, H.; Ci, D.W.; Kang, T.; Ming, T.Y.; Wan, S.B. Effect of drought stresses at different growth stages on peanut leaf protective enzyme activities and osmoregulation substances content. Acta Agron. Sin. 2013, 39, 133–141. [Google Scholar] [CrossRef]
- Cohen, I.; Zandalinas, S.I.; Huck, C.; Fritschi, F.B.; Mittler, R. Meta-analysis of drought and heat stress combination impact on crop yield and yield components. Physiol. Plant 2021, 171, 66–76. [Google Scholar] [CrossRef]
- Yang, H.L.; Zhou, Y.; Zhang, D.Y. The research of transgenic ALDH cotton on drought and salt tolerance. Xinjiang Agric. Sci. 2015, 52, 1177–1182. [Google Scholar]
- Han, Y.L.; Wang, X.C.; Xu, X.L.; Gao, Y.; Wen, G.S.; Zhang, G.S.; Zhang, R.M.; Wang, Y.K. Responses of anti-oxidant enzymes and the ascorbate-glutathione cycle to heat, drought, and synergistic stress in Phyllostachys edulis seedings. J. Zhejiang A F Univ. 2018, 35, 268–276. [Google Scholar]
- Wang, Y.T.; Wang, Z.Z.; Zhao, T.; Hui, Z.M. Effect of 24-Epibrassinolide pretreatments on the antioxidant system and osmotic adjustment substance in Vitis vinifera seedlings under drought stress. Acta Bot. Boreali Occident. Sin. 2019, 39, 489–497. [Google Scholar]
- Abrams, D.; Kubiske, E.; Steiner, C. Drought adaptations and responses in five genotypes of Fraxinus pennsylvanica Marsh.: Photosynthesis, water relations and leaf morphology. Tree Physiol. 1990, 6, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Shirinpour, M.; Asghari, A.; Aharizad, S.; Rasoulzadeh, A.; Khorasani, S.K. Genetic interaction and inheritance of biochemical traits can predict tolerance of hybrid maize cv.SC704 to drought. Acta Physiol. Plant. 2020, 42, 124–137. [Google Scholar] [CrossRef]
- Wang, L.B.; Zu, W.; Dong, S.K.; Li, L.J.; Xu, Y.H. Effect of drought stresses and times on compensation effect after re-watering in soybean. Trans. Chin. Soc. Agric. Eng. 2015, 31, 150–156. [Google Scholar]
- Li, J.; Zhao, S.; Yu, X.; Du, W.; Li, H.; Sun, Y.; Sun, H.; Ruan, C. Role of Xanthoceras sorbifolium MYB44 in tolerance to combined drought and heat stress via modulation of stomatal closure and ROS homeostasis. Plant Physiol. Biochem. 2021, 162, 410–420. [Google Scholar] [CrossRef]
- Ramachandra, R.A.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Chlorophyll Content/[SPAD] | Net Photosynthetic Rate /[μmol·(m2·s)−1] | Stomatal Conductance /[mol·(m2·s)−1] | Carbon Dioxide Concentration/[μmol·mol−1] | Stomatal Conductance /[mmol·(m2·s)−1] | |
|---|---|---|---|---|---|---|
| CK | 5 d | 38.1 ± 0.5 b | 8.68 ± 0.56 a | 0.132 ± 0.011 a | 1240 ± 17 b | 2500 ± 27 a |
| 10 d | 38.3 ± 0.6 b | 10.21 ± 0.48 a | 0.168 ± 0.009 a | 1241 ± 29 b | 3100 ± 31 a | |
| 15 d | 38.3 ± 0.3 b | 10.34 ± 0.32 a | 0.143 ± 0.007 a | 1240 ± 57 b | 2300 ± 25 a | |
| W | 5 d | 37.0 ± 1.7 b | 6.75 ± 0.12 ab | 0.081 ± 0.001 b | 1194 ± 89 c | 1600 ± 47 b |
| 10 d | 33.1 ± 2.4 c | 5.25 ± 0.31 c | 0.070 ± 0.004 b | 1034 ± 46 d | 900 ± 36 b | |
| 15 d | 26.2 ± 0.4 d | 2.90 ± 0.19 d | 0.036 ± 0.002 bc | 1235 ± 35 c | 800 ± 23 b | |
| D | 5 d | 30.4 ± 0.9 d | 6.11 ± 0.48 bc | 0.100 ± 0.010 ab | 1213 ± 48 b | 1500 ± 19 b |
| 10 d | 28.2 ± 1.5 d | 4.18 ± 0.21 c | 0.053 ± 0.002 bc | 1097 ± 57 cd | 1200 ± 13 b | |
| 15 d | 17.5 ± 1.7 e | 1.58 ± 0.03 d | 0.033 ± 0.002 bc | 1254 ± 46 b | 700 ± 27 b | |
| H | 5 d | 38.9 ± 0.2 b | 8.82 ± 0.04 a | 0.143 ± 0.003 a | 1258 ± 49.4 b | 3000 ± 109 a |
| 10 d | 42.4 ± 1.1 a | 9.99 ± 0.32 a | 0.145 ± 0.009 a | 1248 ± 86 b | 2600 ± 69 a | |
| 15 d | 36.5 ± 3.0 b | 6.42 ± 0.26 b | 0.109 ± 0.006 ab | 1234 ± 54 b | 2300 ± 181 a | |
| HW | 5 d | 33.8 ± 1.3 c | 5.53 ± 0.11 bc | 0.080 ± 0.003 b | 1043 ± 68 cd | 1400 ± 79 b |
| 10 d | 25.6 ± 0.5 d | 2.93 ± 0.03 cd | 0.032 ± 0.001 bc | 1294 ± 88 b | 1200 ± 67 b | |
| 15 d | 14.3 ± 1.3 f | 1.64 ± 0.07 d | 0.027 ± 0.001 c | 1270 ± 99 b | 800 ± 70 b | |
| HD | 5 d | 28.8 ± 2.0 d | 4.18 ± 0.19 c | 0.078 ± 0.005 b | 1027 ± 37 d | 1100 ± 78 b |
| 10 d | 12.3 ± 0.1 f | 1.44 ± 0.07 d | 0.027 ± 0.001 c | 1238 ± 49 b | 1500 ± 46 b | |
| 15 d | 7.6 ± 0.1 g | −2.13 ± 0.08 e | 0.021 ± 0.001 c | 1591 ± 65 a | 300 ± 13 c | |
| Processing | a | b | c | d | e | f | g | h | i | j | k | l | m | Position |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK1 | 0.88 | 0.95 | 0.97 | 1.00 | 0.00 | 0.21 | 1.00 | 0.87 | 0.13 | 0.75 | 0.38 | 0.78 | 0.68 | 1 |
| CK2 | 0.88 | 0.94 | 1.00 | 0.97 | 0.00 | 0.20 | 0.98 | 0.99 | 0.09 | 1.00 | 0.38 | 1.00 | ||
| CK3 | 0.88 | 1.00 | 0.93 | 0.99 | 0.00 | 0.21 | 0.98 | 1.00 | 0.11 | 0.83 | 0.38 | 0.72 | ||
| W1 | 0.85 | 0.79 | 0.86 | 0.88 | 0.11 | 0.42 | 0.79 | 0.71 | 0.17 | 0.41 | 0.30 | 0.45 | 0.48 | 3 |
| W2 | 0.73 | 0.71 | 0.59 | 0.80 | 0.21 | 0.49 | 0.62 | 0.59 | 0.27 | 0.34 | 0.01 | 0.21 | ||
| W3 | 0.53 | 0.52 | 0.33 | 0.72 | 0.42 | 0.30 | 0.55 | 0.40 | 0.60 | 0.10 | 0.37 | 0.18 | ||
| D1 | 0.66 | 0.74 | 0.73 | 0.83 | 0.16 | 0.61 | 0.59 | 0.66 | 0.20 | 0.54 | 0.33 | 0.43 | 0.44 | 4 |
| D2 | 0.59 | 0.60 | 0.55 | 0.72 | 0.34 | 0.34 | 0.53 | 0.51 | 0.41 | 0.22 | 0.12 | 0.31 | ||
| D3 | 0.28 | 0.37 | 0.22 | 0.66 | 0.53 | 0.15 | 0.41 | 0.30 | 0.70 | 0.08 | 0.40 | 0.14 | ||
| H1 | 0.90 | 0.94 | 0.98 | 0.98 | 0.19 | 0.32 | 0.99 | 0.88 | 0.00 | 0.83 | 0.41 | 0.94 | 0.70 | 2 |
| H2 | 1.00 | 0.84 | 0.92 | 0.96 | 0.33 | 0.42 | 0.95 | 0.97 | 0.10 | 0.84 | 0.39 | 0.81 | ||
| H3 | 0.83 | 0.73 | 0.90 | 0.90 | 0.60 | 0.75 | 0.93 | 0.69 | 0.26 | 0.60 | 0.37 | 0.71 | ||
| HW1 | 0.75 | 0.75 | 0.54 | 0.79 | 0.52 | 1.00 | 0.58 | 0.61 | 0.29 | 0.40 | 0.03 | 0.38 | 0.45 | 5 |
| HW2 | 0.52 | 0.60 | 0.36 | 0.69 | 0.65 | 0.46 | 0.49 | 0.41 | 0.59 | 0.08 | 0.47 | 0.38 | ||
| HW3 | 0.19 | 0.15 | 0.12 | 0.47 | 0.77 | 0.22 | 0.32 | 0.30 | 0.87 | 0.04 | 0.43 | 0.17 | ||
| HD1 | 0.61 | 0.60 | 0.33 | 0.66 | 0.50 | 0.92 | 0.50 | 0.51 | 0.29 | 0.39 | 0.00 | 0.27 | 0.34 | 6 |
| HD2 | 0.13 | 0.28 | 0.17 | 0.63 | 1.00 | 0.15 | 0.29 | 0.29 | 0.81 | 0.04 | 0.37 | 0.43 | ||
| HD3 | 0.00 | 0.00 | 0.00 | 0.00 | 0.12 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 |
| a | b | c | d | e | f | g | h | i | j | k | l | m | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a | 1.00 | ||||||||||||
| b | −0.96 ** | 1.00 | |||||||||||
| c | −0.94 ** | 0.94 ** | 1.00 | ||||||||||
| d | −0.90 ** | 0.93 ** | 0.88 ** | 1.00 | |||||||||
| e | −0.93 ** | 0.91 ** | 0.95 ** | 0.78 ** | 1.00 | ||||||||
| f | −0.92 ** | 0.89 ** | 0.90 ** | 0.71 ** | 0.96 ** | 1.00 | |||||||
| g | −0.94 ** | 0.94 ** | 0.96 ** | 0.92 ** | 0.92 ** | 0.86 ** | 1.00 | ||||||
| h | −0.97 ** | 0.97 ** | 0.94 ** | 0.88 ** | 0.93 ** | 0.91 ** | 0.91 ** | 1.00 | |||||
| i | 0.94 ** | −0.95 ** | −0.96 ** | −0.91 ** | −0.92 ** | −0.87 ** | −0.96 ** | −0.94 ** | 1.00 | ||||
| j | 0.87 ** | −0.88 ** | −0.93 ** | −0.78 ** | −0.93 ** | −0.88 ** | −0.91 ** | −0.90 ** | 0.95 ** | 1.000 | |||
| k | −0.45 * | 0.44 * | 0.24 | 0.53 * | 0.21 | 0.19 | 0.29 | 0.47 * | −0.33 | −0.17 | 1.00 | ||
| l | 0.76 ** | −0.80 ** | −0.88 ** | −0.79 ** | −0.78 ** | −0.73 ** | −0.88 ** | −0.77 ** | 0.88 ** | 0.91 ** | −0.05 | 1.00 | |
| m | 0.93 ** | −0.93 ** | −0.95 ** | −0.92 ** | −0.88 ** | −0.85 ** | −0.97 ** | −0.92 ** | 0.96 ** | 0.92 ** | −0.31 | 0.92 ** | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, Y.; Li, J.; Liao, Y.; Mbuni, Y.M.; Li, X.; Khong, Q. Physiological Response and Comprehensive Resistance Evaluation of East African Endemic Aeollanthus repens under Water and Heat Stress. Int. J. Plant Biol. 2024, 15, 397-411. https://doi.org/10.3390/ijpb15020031
Tu Y, Li J, Liao Y, Mbuni YM, Li X, Khong Q. Physiological Response and Comprehensive Resistance Evaluation of East African Endemic Aeollanthus repens under Water and Heat Stress. International Journal of Plant Biology. 2024; 15(2):397-411. https://doi.org/10.3390/ijpb15020031
Chicago/Turabian StyleTu, Yingying, Jitao Li, Yiying Liao, Yuvenalis Morara Mbuni, Xiaoning Li, and Qiyan Khong. 2024. "Physiological Response and Comprehensive Resistance Evaluation of East African Endemic Aeollanthus repens under Water and Heat Stress" International Journal of Plant Biology 15, no. 2: 397-411. https://doi.org/10.3390/ijpb15020031
APA StyleTu, Y., Li, J., Liao, Y., Mbuni, Y. M., Li, X., & Khong, Q. (2024). Physiological Response and Comprehensive Resistance Evaluation of East African Endemic Aeollanthus repens under Water and Heat Stress. International Journal of Plant Biology, 15(2), 397-411. https://doi.org/10.3390/ijpb15020031