
Salt Tolerance and Ion Accumulation in Several Halophytic Plant Species Depending on the Type of Anion



Abstract
:
1. Introduction
2. Materials and Methods
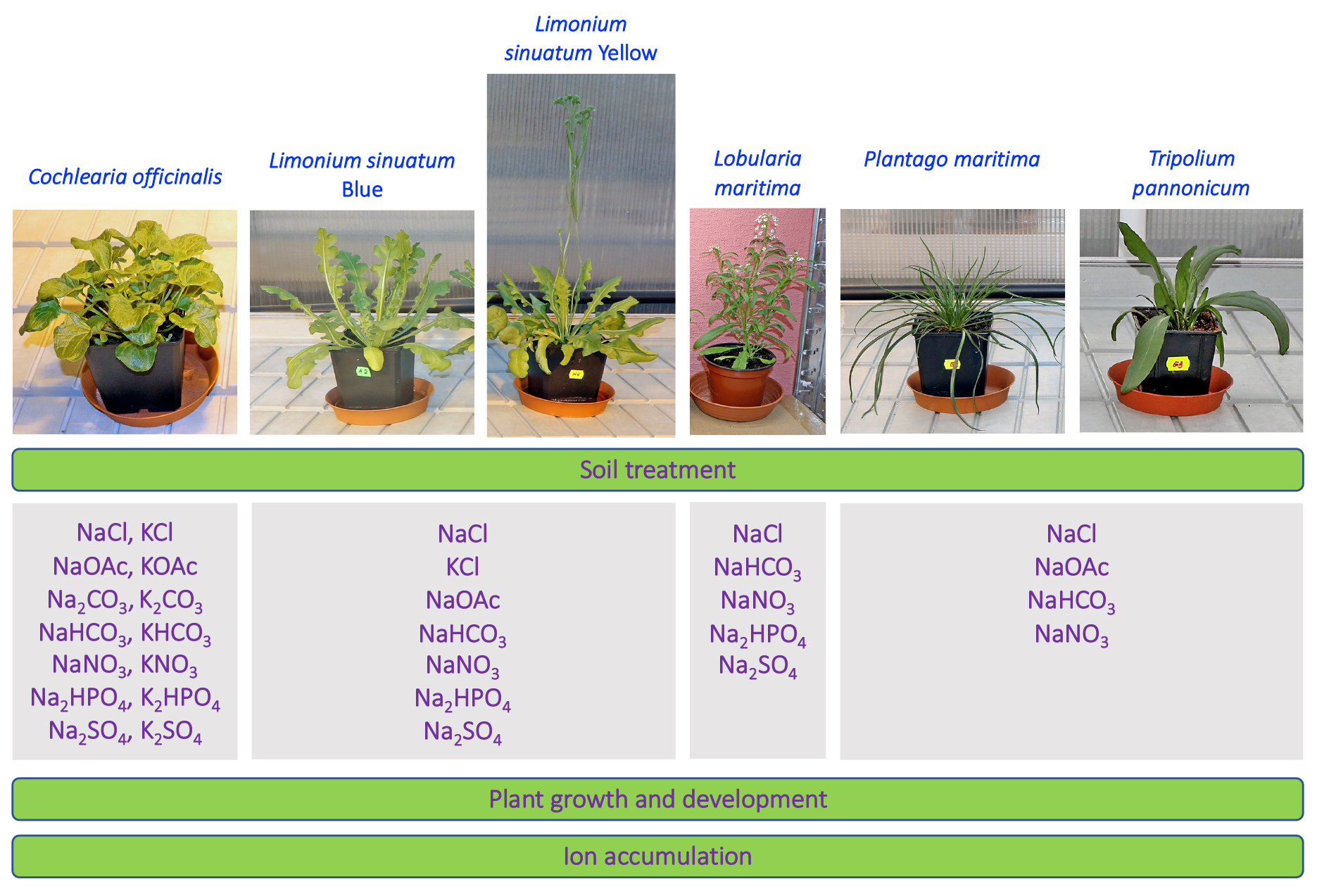
2.1. Plant Material
2.2. Plant Establishment and Cultivation Conditions
2.3. Treatments
2.4. Physiological Measurements
2.5. Plant Harvest and Measurements
2.6. Data Analysis
3. Results
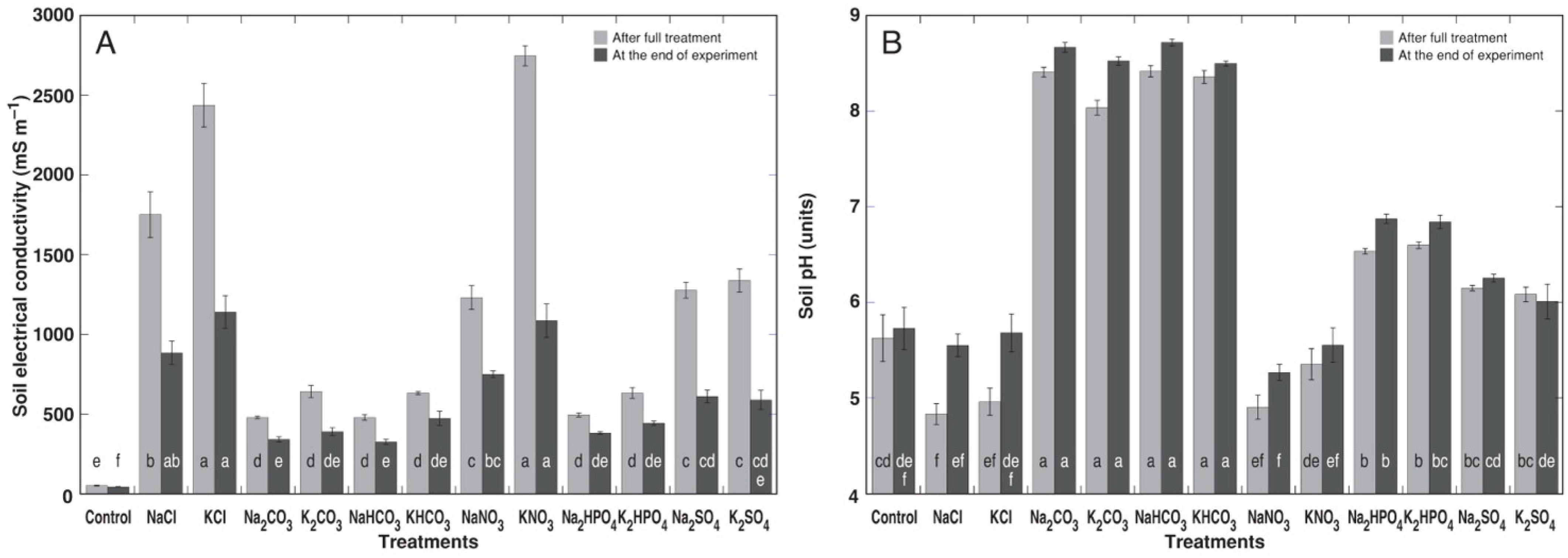
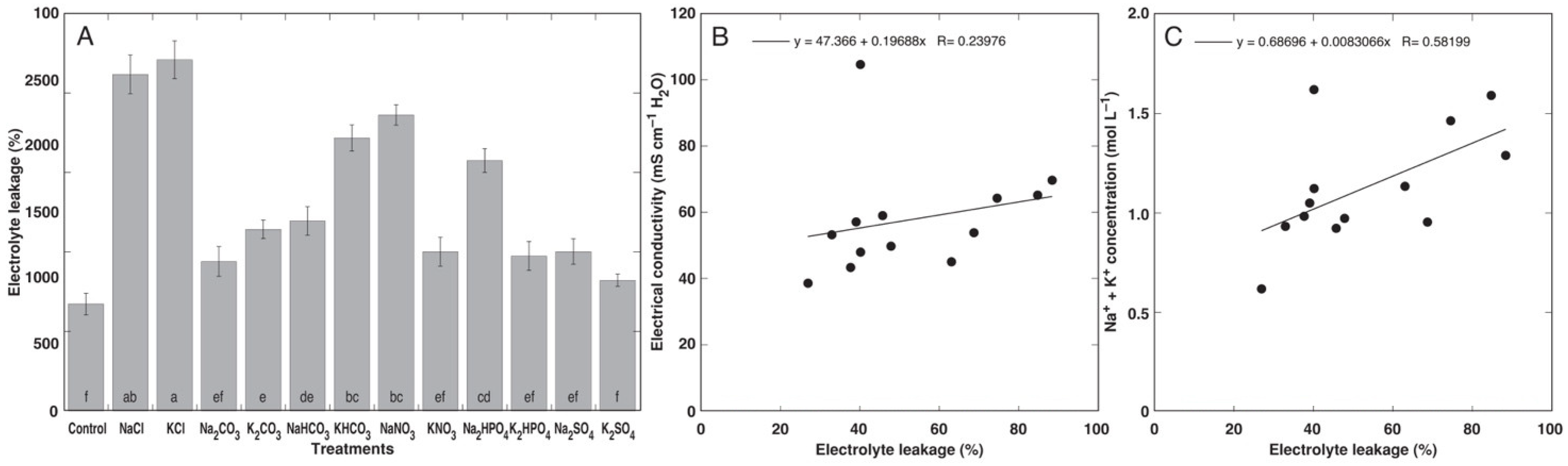
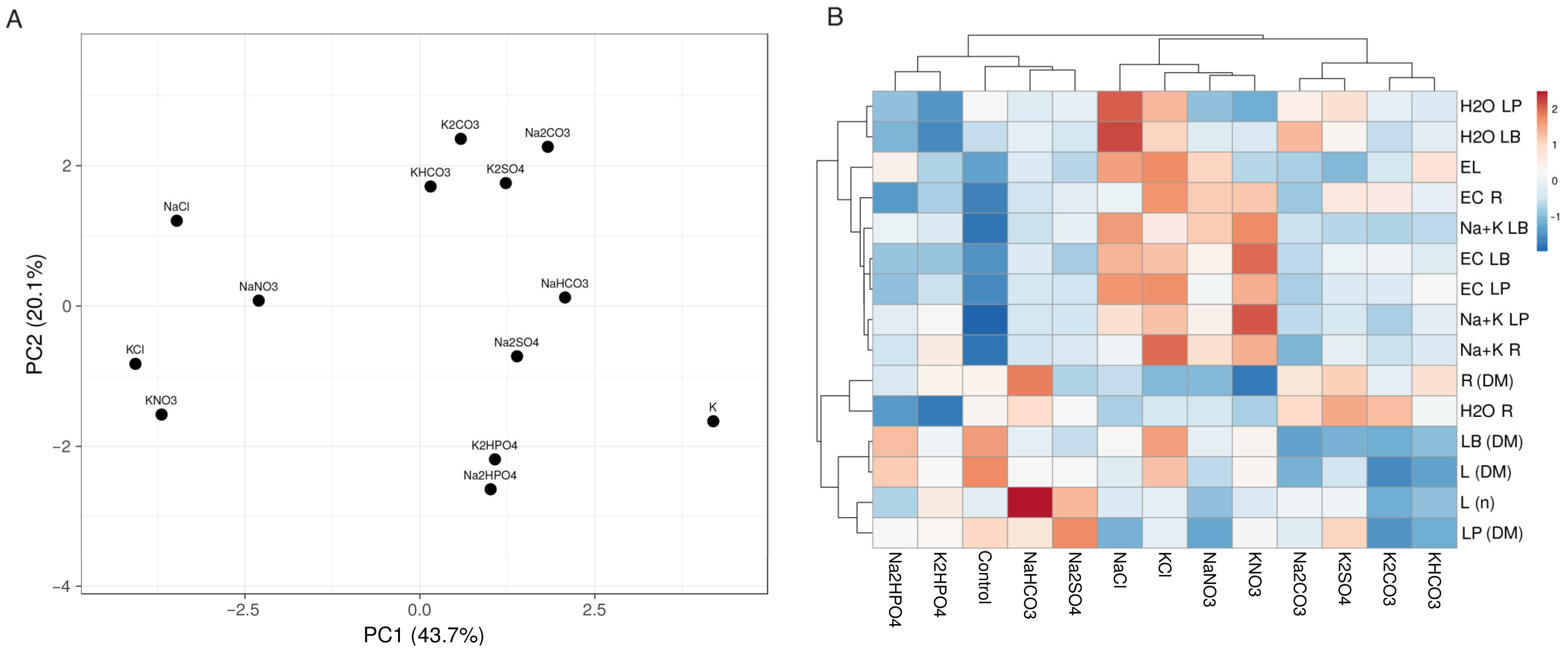
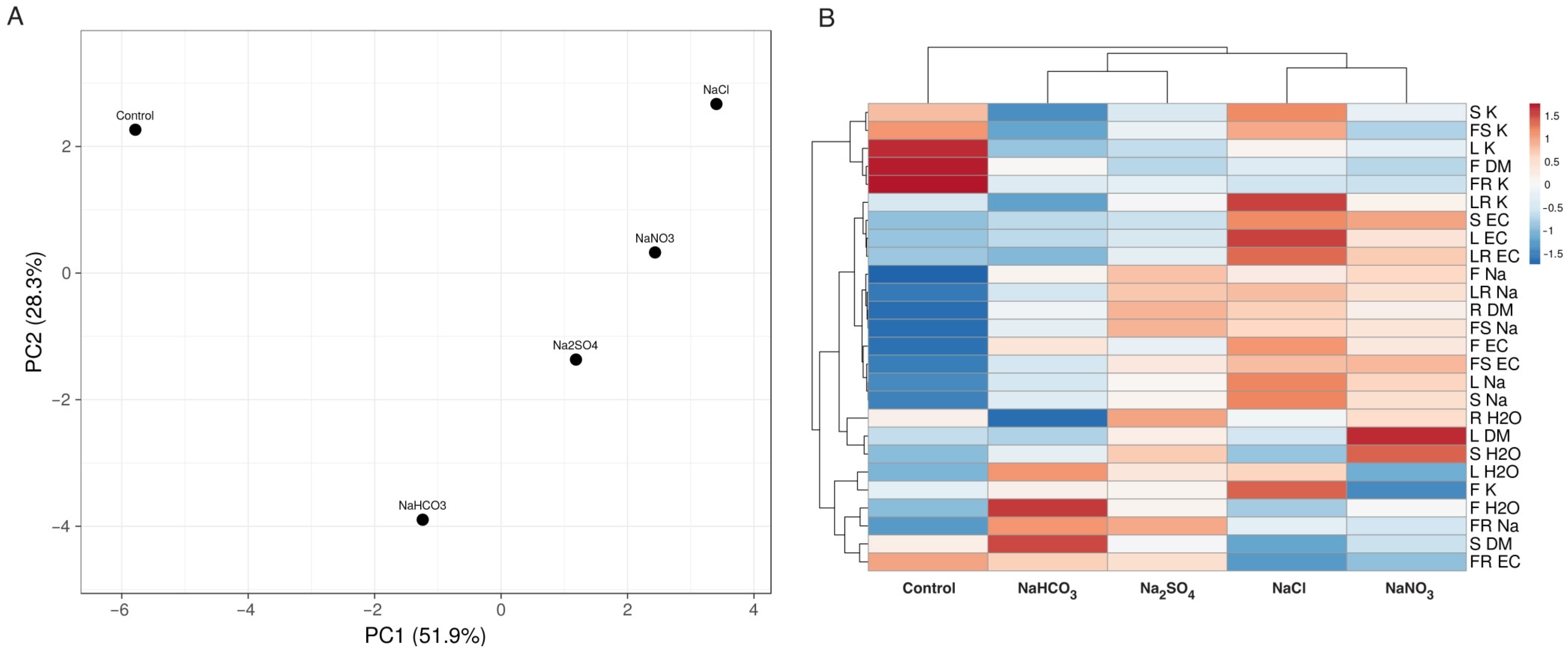
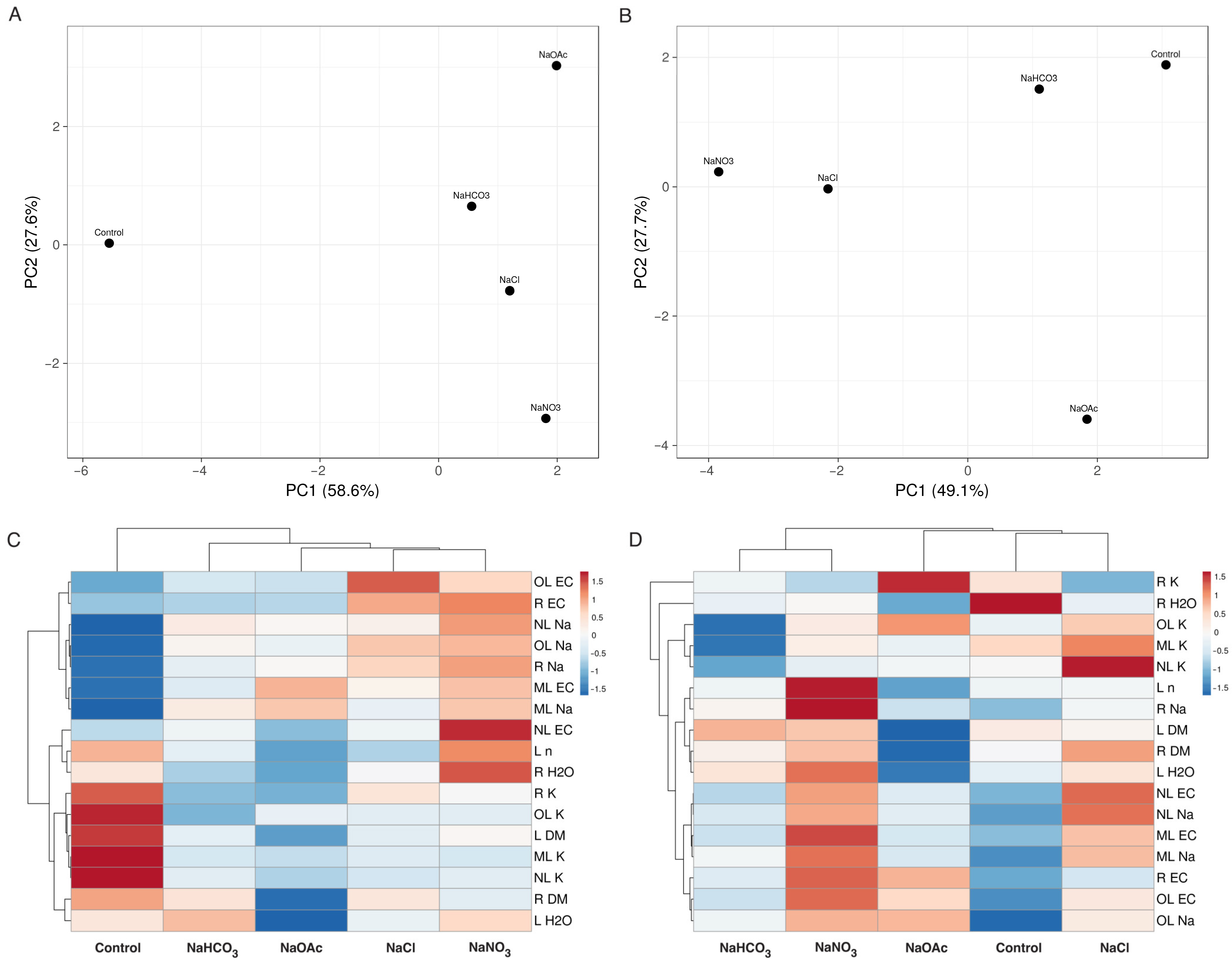
3.1. Experiment with Cochlearia officinalis
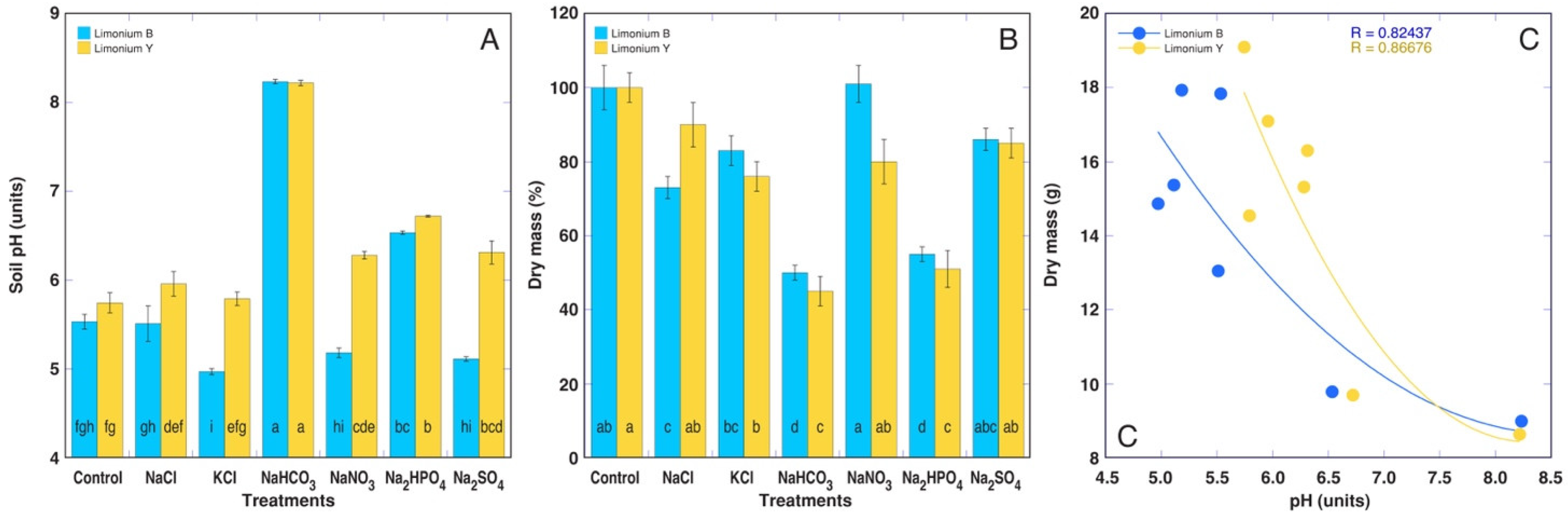
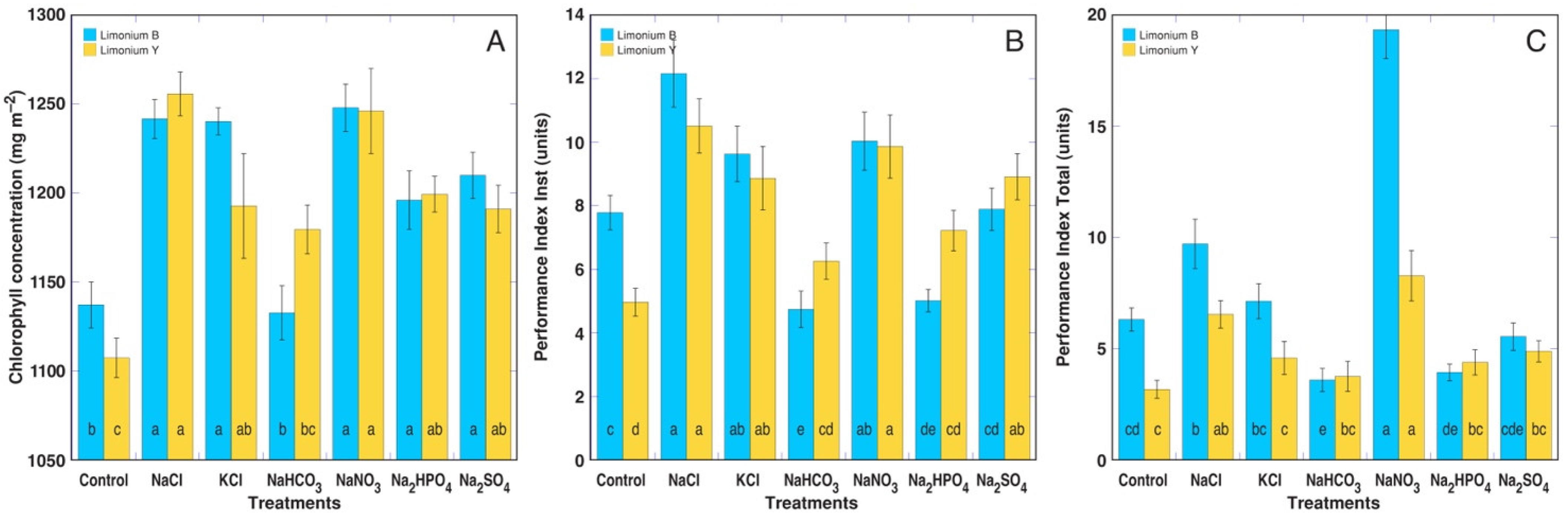
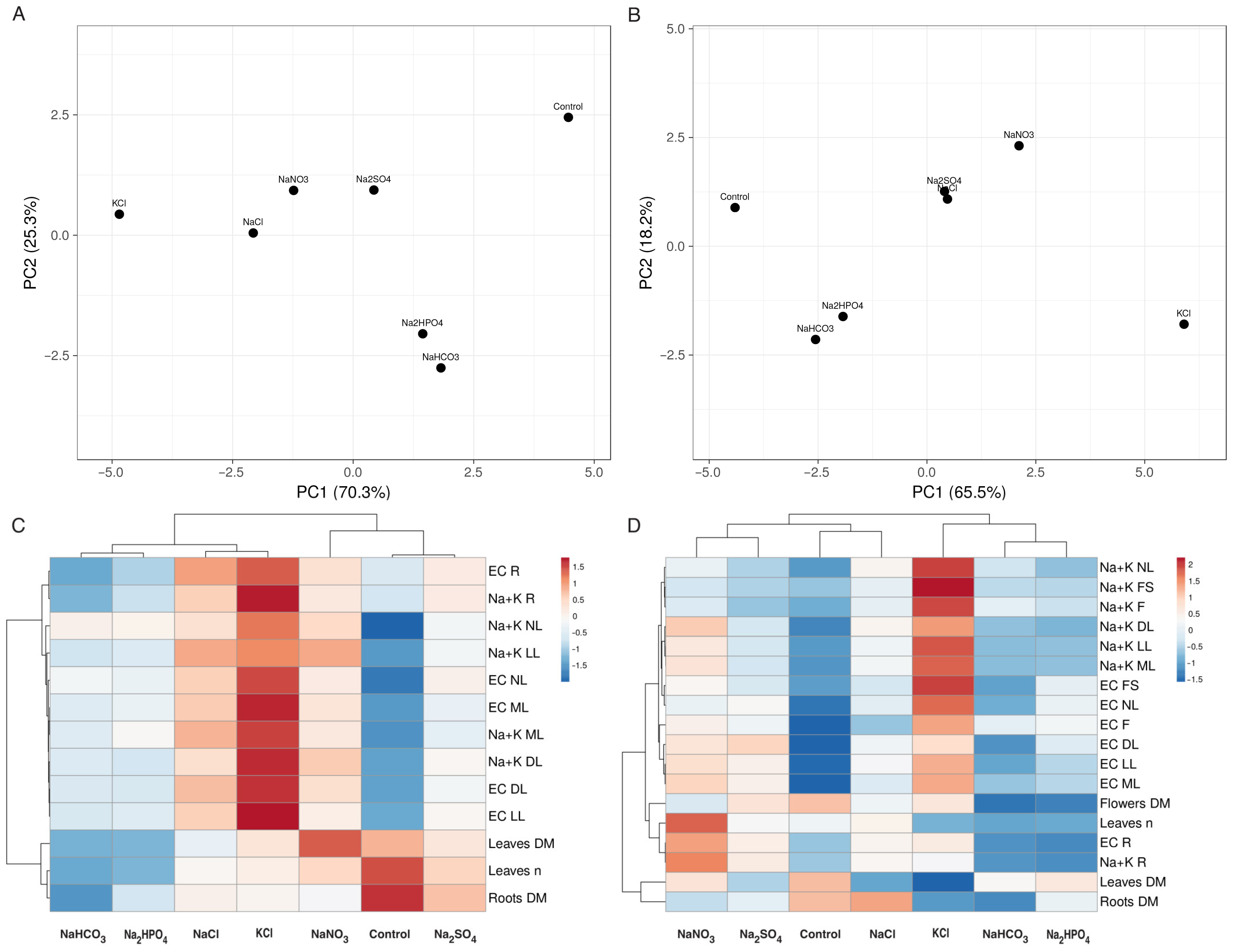
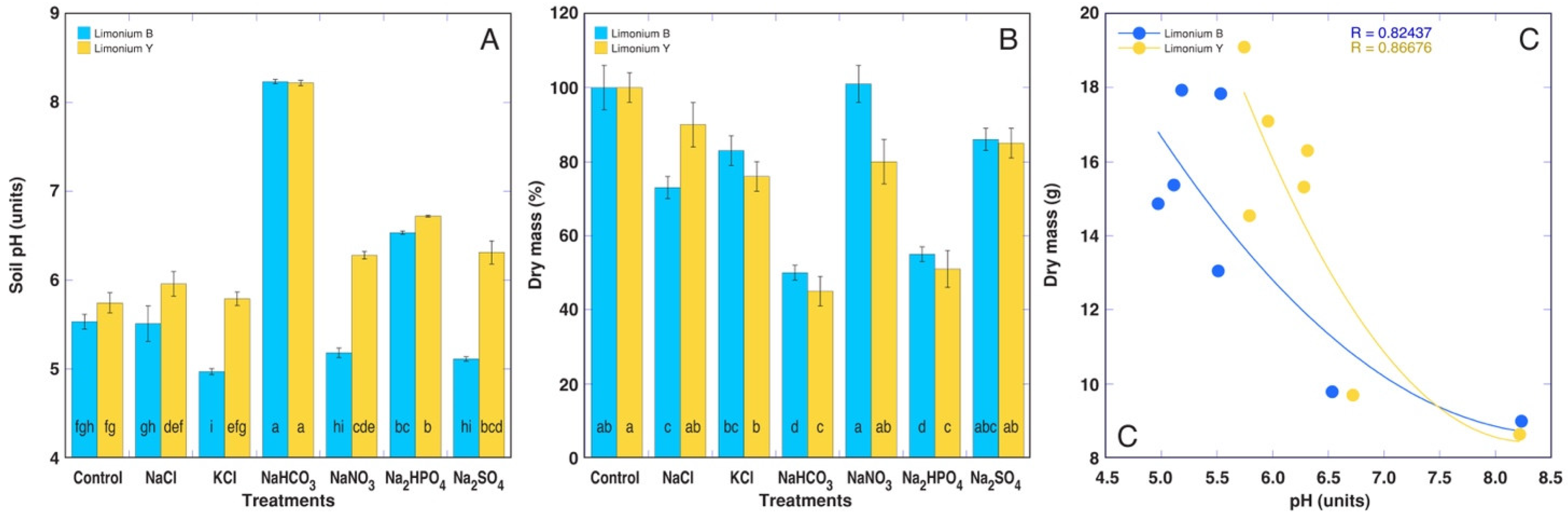
3.2. Experiment with Limonium sinuatum
3.3. Experiment with Lobularia maritima
3.4. Experiment with Plantago maritima and Tripolium pannonicum
4. Discussion
4.1. General Aspects
4.2. Salinity Tolerance as Affected by Anion Type
4.3. Ion Accumulation Capacity
4.4. Genotype-Related Differences
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ondrasek, G.; Rengel, Z. Environmental salinization processes: Detection, implications & solutions. Sci. Total Environ. 2021, 754, 142432. [Google Scholar]
- Uri, N. Cropland salinization and associated hydrology: Trends, processes and examples. Water 2018, 10, 1030. [Google Scholar] [CrossRef]
- Hammer, U.T. The saline lakes of Saskatchewan. III. Chemical characterization. Hydrobiology 1978, 63, 311–335. [Google Scholar] [CrossRef]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ response mechanisms to salinity stress. Plants 2023, 12, 2253. [Google Scholar] [CrossRef]
- Carden, D.E.; Walker, D.J.; Flowers, T.J.; Miller, A.J. Single-cell measurements of the contribution of cytosolic Na+ and K+ to salt tolerance. Plant Physiol. 2003, 131, 676–683. [Google Scholar] [CrossRef]
- Flowers, T.J.; Läuchli, A. Sodium versus potassium: Substitution and compartmentation. In Inorganic Plant Nutrition; Läuchli, A., Bieleski, R.L., Eds.; Springer: Berlin/Heidelberg, Germany, 1983; Volume 15B, pp. 651–681. [Google Scholar]
- Subbarao, G.V.; Ito, O.; Berry, W.L.; Wheeler, R.M. Sodium: A functional plant nutrient. Crit. Rev. Plant Sci. 2003, 22, 391–416. [Google Scholar]
- Rodriguez-Navarro, A. High-affinity potassium and sodium transport systems in plants. J. Exp. Bot. 2006, 57, 1149–1160. [Google Scholar] [CrossRef]
- Colmenero-Flores, J.M.; Franco-Navarro, J.D.; Cubero-Font, P.; Peinado-Torrubia, P.; Rosales, M.A. Chloride as a beneficial macronutrient in higher plants: New roles and regulation. Int. J. Mol. Sci. 2019, 20, 4686. [Google Scholar] [CrossRef]
- Bazihizina, N.; Colmer, T.D.; Cuin, T.A.; Mancuso, S.; Shabala, S. Friend or foe? Chloride patterning in halophytes. Trends Plant Sci. 2018, 24, 142–151. [Google Scholar] [CrossRef]
- Chen, S.; Xing, J.; Lan, H. Comparative effects of neutral salt and alkaline salt stress on seed germination, early seedling growth and physiological response of a halophyte species Chenopodium glaucum. Afr. J. Biotechnol. 2012, 11, 9572–9581. [Google Scholar]
- Yang, C.W.; Wang, P.; Li, C.Y.; Shi, D.C.; Wang, D.L. Comparison of effects of salt and alkali stresses on the growth and photosynthesis of wheat. Photosynthetica 2008, 46, 107–114. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.X.; Ding, X.M.; Hu, Y.; Tian, S.Y.; Yan, D.F.; Shao, S.; Gao, Y.; Liu, R.; Yang, Y.F. Effects of saline and alkaline stress on germination, seedling growth, and ion balance in wheat. Agron. J. 2010, 102, 1252–1260. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Z.; Chen, Y.; Yang, C.; Shi, D. Effects of salt and alkali stresses on growth and ion balance in rice (Oryza sativa L.). Plant Soil Environ. 2011, 57, 286–294. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.X.; Yan, C.; Zhong, X.; Gu, F.X.; Liu, Q.; Xa, X.; Li, H. Ionomic and metabolic responses to neutral salt or alkaline salt stresses in maize (Zea mays L.) seedlings. BMC Plant Biol. 2017, 17, 41. [Google Scholar] [CrossRef]
- Li, J.; Hussain, T.; Feng, X.; Guo, K.; Chen, H.; Yang, C.; Liu, X. Comparative study on the resistance of Suaeda glauca and Suaeda salsa to drought, salt, and alkali stresses. Ecol. Eng. 2019, 140, 105593. [Google Scholar] [CrossRef]
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859–868. [Google Scholar]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wiren, N. Three functional transporters for constitutive, diurnally regulated, and starvation-induced uptake of ammonium into Arabidopsis roots. Plant Cell 1999, 11, 937–948. [Google Scholar] [CrossRef]
- Raddatz, N.; de los Rios, L.M.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated transport of nitrate, potassium, and sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Coskun, D.; Britto, D.T.; Kronzucker, H.J. The nitrogen-potassium intersection: Membranes, metabolism, and mechanism. Plant Cell Environ. 2017, 40, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Ievinsh, G. Type of anion largely determines salinity tolerance in four Rumex species. Plants 2023, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Colmer, T.D. Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 2015, 115, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Ievinsh, G.; Ieviņa, S.; Andersone-Ozola, U.; Samsone, I. Leaf sodium, potassium and electrolyte accumulation capacity of plant species from salt-affected coastal habitats of the Baltic Sea: Towards a definition of Na hyperaccumulation. Flora 2021, 274, 151748. [Google Scholar] [CrossRef]
- Pegtel, D.M. Effect of ploidy level on fruit morphology, seed germination and juvenile growth in scurvy grass (Cochlearia officinalis L. s.l., Brassicaceae). Plant Species Biol. 1999, 14, 201–215. [Google Scholar] [CrossRef]
- Lepping, O.; Daniels, F.J.A. Phytosociology of beach and salt marsh vegetation in Northern West Greenland. Polarforschung 2006, 76, 95–108. [Google Scholar]
- de Vos, A.C.; Broekman, R.; de Almeida Guerra, C.C.; van Rijsselberghe, M.; Rozema, J. Developing and testing new halophyte crops: A case study of salt tolerance of two species of the Brassicaceae, Diplotaxis tenuifolia and Cochlearia officinalis. Environ. Exp. Bot. 2013, 92, 154–164. [Google Scholar] [CrossRef]
- Le Saos, J. Effets du NaCl et du CaCl2 sur la croissance du Cochlearia anglica. Bull. Soc. Bot. France Act. Bot. 1978, 3–4, 53–59. [Google Scholar] [CrossRef]
- Malekmohammadi, M.; Akhani, H.; Borsch, T. Phylogenetic relationships of Limonium (Plumbaginaceae) inferred from multiple chloroplast and nuclear loci. Taxon 2017, 66, 1128–1146. [Google Scholar] [CrossRef]
- Koutroumpa, K.; Theodoridis, S.; Warren, B.H.; Jiménez, A.; Celep, F.; Doğan, M.; Romeiras, M.M.; Santos-Guerra, A.; Fernández-Palacios, J.M.; Caujapé-Castells, J.; et al. An expanded molecular phylogeny of Plumbaginaceae, with emphasis on Limonium (sea lavenders): Taxonomic implications and biogeographic considerations. Ecol. Evol. 2018, 8, 12397–12424. [Google Scholar] [CrossRef]
- Grieve, C.; Poss, J.A. Productivity and mineral nutrition of Limonium species irrigated with saline wastewaters. HortScience 2005, 40, 654–658. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Y.; Mi, P.; Wang, B.; Yuan, F. Salt-tolerance screening in Limonium sinuatum varieties with different flower colors. Sci. Rep. 2021, 11, 14562. [Google Scholar] [CrossRef] [PubMed]
- Popova, O.V.; Golldack, D. In the halotolerant Lobularia maritima (Brassicaceae) salt adaptation correlates with activation of the vacuolar H+-ATPase and the vacuolar Na+/H+ antiporter. J. Plant Physiol. 2007, 164, 1278–1288. [Google Scholar] [CrossRef] [PubMed]
- Popova, O.V.; Yang, O.; Dietz, K.-J.; Golldack, D. Differential transcript regulation in Arabidopsis thaliana and the halotolerant Lobularia maritima indicates genes with potential function in plant salt adaptation. Gene 2008, 423, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Ben Hsouna, A.; Michalak, M.; Kukula-Koch, W.; Ben Saad, R.; ben Romdhane, W.; Zeljković, S.Ć.; Mnif, W. Evaluation of halophyte biopotential as an unused natural resource: The case of Lobularia maritima. Biomolecules 2022, 12, 1583. [Google Scholar] [CrossRef] [PubMed]
- Ben Hsouna, A.; Ghneim-Herrera, T.; Ben Romdhane, W.; Dabbous, A.; Ben Saad, R.; Brini, F.; Abdelly, C.; Ben Hamed, K. Early effects of salt stress on the physiological and oxidative status of the halophyte Lobularia maritima. Funct. Plant Biol. 2020, 47, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Ben Saad, R.; Ben Romdhane, W.; Baazaoui, N.; Bouteraa, M.; Ben Hsouna, A.; Mishra, A.; Zelković, B.Ć. Assessment of the cadmium and copper phytoremediation potential of the Lobularia maritima thioredoxin 2 gene using genetically engineered tobacco. Agronomy 2023, 13, 399. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Makowski, W. Assessment of shoot priming efficiency to counteract complex metal stress in halotolerant Lobularia maritima. Plants 2023, 12, 1440. [Google Scholar] [CrossRef]
- Sleimi, N.; Guerfali, S.; Bankaji, I. Biochemical indicators of salt stress in Plantago maritima: Implications for environmental stress assessment. Ecol. Indic. 2015, 48, 570–577. [Google Scholar] [CrossRef]
- Ievinsh, G.; Andersone-Ozola, U.; Jēkabsone, A. Similar responses of relatively salt tolerant plants to Na and K during chloride salinity: Comparison of growth, water content and ion accumulation. Life 2022, 12, 1577. [Google Scholar] [CrossRef]
- Andersone, U.; Samsone, I.; Ievinsh, G. Protection of photosynthesis in coastal marsh plants Aster tripolium and Hydrocotyle vulgaris in conditions of increased soil salinity. Environ. Exp. Biol. 2012, 10, 89–97. [Google Scholar]
- Ludwiczak, A.; Ciarkowska, A.; Dehnavi, A.R.; Cárdenas-Pérez, S.; Piernik, A. Growth stage-, organ- and time-dependent salt tolerance of halophyte Tripolium pannonicum (Jacq.) Dobrocz. Life 2023, 13, 462. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Yu, B.; Liu, Y. Differential sensitivity to chloride and sodium ions in seedlings of Glycine max and G. soja under NaCl stress. J. Plant Physiol. 2005, 162, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucl. Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Boros, E.; Kolpakova, M. A review of the defining chemical properties of soda lakes and pans: An assessment on a large geographic scale of Eurasian inland saline surface waters. PLoS ONE 2018, 13, 0202205. [Google Scholar] [CrossRef] [PubMed]
- Pengasamy, P. Soil chemistry factors confounding crop salinity tolerance—A review. Agronomy 2016, 6, 53. [Google Scholar] [CrossRef]
- Fang, S.; Hou, X.; Liang, X. Response mechanisms of plants under saline-alkali stress. Front. Plant Sci. 2021, 12, 667458. [Google Scholar] [CrossRef]
- Yang, C.W.; Jianaer, A.; Li, C.Y.; Shi, D.C.; Wang, D.L. Comparison of the effects of salt-stress and alkali-stress on photosynthesis and energy storage of an alkali-resistant halophyte Chloris virgata. Photosynthetica 2008, 46, 273–278. [Google Scholar] [CrossRef]
- Csitári, B.; Bedics, A.; Felföldi, T.; Boros, E.; Nagy, H.; Máthé, I.; Szkely, A.J. Anion-type modulates the effect of salt stress on saline lake bacteria. Extremophiles 2022, 26, 12. [Google Scholar] [CrossRef]
- Faget, M.; Blossfeld, S.; von Gillhaussen, P.; Schurr, U.; Temperton, V.M. Disentangling who is who during rhizosphere acidification in root interactions: Combining fluorescence with optode techniques. Front. Plant Sci. 2013, 4, 392. [Google Scholar] [CrossRef]
- Martin, P.K.; Koebner, R.M.D. Sodium and chloride ions contribute synergistically to salt toxicity in wheat. Biol. Plant. 1995, 37, 265–271. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Renagasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl− ions in soil have simultaneous detrimental effect on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Magen, H.; Tarchitzky, J.; Kafkafi, U. Advances in chloride nutrition of plants. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2000; Volume 68, pp. 97–150. [Google Scholar]
- Turner, J.E.; Greville, K.; Murphy, E.C.; Hooks, M.A. Characterization of Arabidopsis fluoroacetate-resistant mutants reveals the principal mechanism of acetate activation for entry into the glyoxylate cycle. J. Biol. Chem. 2005, 280, 2780–2787. [Google Scholar] [CrossRef]
- Lin, M.; Oliver, D.J. The role of acetyl-coenzyme A synthetase in Arabidopsis. Plant Physiol. 2008, 147, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Yang, H.; Pangestu, F.; Nikolau, B.J. Failure to maintain acetate homeostasis by acetate-activating enzymes impacts plant development. Plant Physiol. 2020, 182, 1256–1271. [Google Scholar] [CrossRef] [PubMed]
- Krotkov, G.; Barker, H.A. Utilization of acetate by tobacco leaves, as determined with C14. Amer. J. Bot. 1948, 35, 12–15. [Google Scholar] [CrossRef]
- Jardine, K.J.; McDowell, N. Fermentation-mediated growth, signaling, and defense in plants. New Phytol. 2023, 239, 839–851. [Google Scholar] [CrossRef]
- Headley, F.B.; Curtis, E.W.; Scofield, C.S. Effect on plant growth of sodium salts in the soil. J. Agric. Res. 1916, 6, 857–869. [Google Scholar]
- Geng, G.; Wang, G.; Stevanato, P.; Lv, C.; Wang, Q.; Yu, L.; Wang, Y. Physiological and proteomic analysis of different molecular mechanisms of sugar beet response to acidic and alkaline pH environment. Front. Plant Sci. 2021, 12, 682799. [Google Scholar] [CrossRef]
- Cole, J.J.; Prairie, Y.T. Dissolved CO2 in freshwater systems. In Encyclopedia of Inland Waters; Elsevier: Amsterdam, The Netherlands, 2009; pp. 30–34. [Google Scholar]
- Ievinsh, G.; Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A. Salinity and heavy metal tolerance, and phytoextraction potential of Ranunculus sceleratus plants from a sandy coastal beach. Life 2022, 12, 1959. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Fernández, J.A.; Rubio, L.; Pérez, L.; Terés, J.; Barcélo, J. Transport and use of bicarbonate in plants: Current knowledge and challenges ahead. Int. J. Mol. Sci. 2018, 19, 1352. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Hu, H.; Ries, A.; Merilo, E.; Kollist, H.; Schroeder, J.I. Central functions of bicarbonate in S-type anion channel activation and OST1 protein kinase in CO2 signal transduction guard cell. EMBO J. 2011, 30, 1645–1658. [Google Scholar] [CrossRef]
- Devinar, G.; Llanes, A.; Masciarelli, O.; Luna, V. Different relative humidity conditions combined with chloride and sulfate salinity treatments modify abscisic acid and salicylic acid levels in the halophyte Prosopis strombulifera. Plant Growth Reg. 2013, 70, 247–256. [Google Scholar] [CrossRef]
- Reginato, M.; Sosa, L.; Llanes, A.; Hampp, E.; Vettorazzi, N.; Reinoso, H.; Luna, V. Growth responses and ion accumulation in the halophytic legume Prosopis strombulifera are determined by Na2SO4 and NaCl. Plant Biol. 2014, 16, 97–106. [Google Scholar] [CrossRef]
- Assareh, M.H.; Rasouli, B.; Amiri, B. Effects of NaCl and Na2SO4 on germination and initial growth phase of Halostachys caspica. Desert 2010, 15, 119–125. [Google Scholar]
- Reich, M.; Aghajanzadeh, T.; Helm, J.; Parmar, S.; Hawkesford, M.J.; De Kok, L.J. Chloride and sulfate salinity differently affect biomass, mineral nutrient composition and expression of sulfate transport and assimilation genes in Brassica rapa. Plant Soil 2017, 411, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Rennenberg, H. The fate of excess sulfur in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 121–153. [Google Scholar] [CrossRef]
- Curtin, D.; Wen, G. Plant cation-anion balance as affected by the ionic composition of the growing medium. Plant Soil 2004, 267, 109–115. [Google Scholar] [CrossRef]
- Yuan, J.-F.; Feng, G.; Ma, H.-Y.; Tian, C.-Y. Effect of nitrate on root development and nitrogen uptake of Suaeda physophora under NaCl salinity. Pedosphere 2010, 20, 536–544. [Google Scholar] [CrossRef]
- Ushakova, S.A.; Kovaleva, N.P.; Tikhomirova, N.A.; Gribovskaya, I.V.; Kolmakova, A.A. Effect of photosynthetically active radiation, salinization, and type of nitrogen nutrition on growth of Salicornia europaea. Russ. J. Plant Physiol. 2006, 53, 785–793. [Google Scholar] [CrossRef]
- Wang, D.; Wang, H.; Han, B.; Wang, B.; Guo, A.; Zheng, D.; Liu, C.; Chang, L.; Peng, M.; Wang, X. Sodium instead of potassium and chloride is an important macronutrient to improve leaf succulence and shoot development for halophyte Sesuvium portulacastrum. Plant Physiol. Biochem. 2012, 51, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E.; Heck, U.; Wiemken, A. Vacuoles as storage compartments for nitrate in barley leaves. Nature 1981, 289, 292–294. [Google Scholar] [CrossRef]
- Liu, T.; Ren, T.; White, P.J.; Cong, R.; Lu, J. Storage nitrogen co-ordinates leaf expansion and photosynthetic capacity in winter oilseed rape. J. Exp. Bot. 2018, 69, 2995–3007. [Google Scholar] [CrossRef] [PubMed]
- Młodzińska, E.; Zboińska, M. Phosphate uptake and allocation—A closer look at Arabidopsis thaliana L. and Oryza sativa L. Front. Plant Sci. 2016, 7, 198. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Rinehart, C.A.; Sahi, S.V. Comprehensive study of excess phosphate response reveals ethylene mediated signaling that negatively regulates plant growth and development. Sci. Rep. 2017, 7, 3074. [Google Scholar] [CrossRef]
- Breckle, S.-W. Salinity, halophytes and salt affected natural ecosystems. In Salinity: Environment–Plants–Molecules; Lauchli, A., Luttge, U., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 53–77. [Google Scholar]
- Ieviņa, S.; Karlsons, A.; Osvalde, A.; Andersone-Ozola, U.; Ievinsh, G. Coastal wetland species Rumex hydrolapathum: Tolerance against flooding, salinity and heavy metals for its potential use in phytoremediation and environmental restoration technologies. Life 2023, 13, 1604. [Google Scholar] [CrossRef]
- Mi, P.; Yuan, F.; Guo, J.; Han, G.; Wang, B. Salt glands play a pivotal role in the salt stress resistance of four recretohalophyte Limonium Mill. species. Plant Biol. 2021, 23, 1063–1073. [Google Scholar] [CrossRef]
- Ahmad, I.; Larher, F.; Stewart, G.R. Sorbitol, a compatible solute in Plantago maritima. New Phytol. 1979, 82, 671–678. [Google Scholar] [CrossRef]
- Ueda, A.; Kanechi, M.; Uno, Y.; Inagaki, N. Photosynthetic limitations of a halophyte sea aster (Aster tripolium L.) under water stress and NaCl stress. J. Plant Res. 2003, 11, 65–70. [Google Scholar] [CrossRef]
- Geissler, N.; Hussin, S.; Koyro, H.-W. Interactive effects of NaCl salinity and elevated atmospheric CO2 concentration on growth, photosynthesis, water relations and chemical composition of the potential cash crop halophyte Aster tripolium L. Environ. Exp. Bot. 2009, 65, 220–231. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Koźmińska, A.; Hanus-Fajerska, E.; Dziurka, K.; Dziurka, M. Insight into phytohormonal modulation of defense mechanisms to salt excess in a halophyte and a glycophyte from Asteraceae family. Plant Soil 2021, 463, 55–76. [Google Scholar] [CrossRef]
- Karlsons, A.; Osvalde, A.; Nečajeva, J.; Ievinsh, G. Changes in nutritional status of coastal plants Hydrocotyle vulgaris and Aster tripolium at elevated soil salinity. Acta Univ. Latv. 2008, 745, 165–177. [Google Scholar]
- Ramani, B.; Reeck, T.; Debez, A.; Stelzer, R.; Huchzermeyer, B.; Schmidt, A.; Papenbrock, J. Aster tripolium L. and Sesuvium portulacastrum L.: Two halophytes, two strategies to survive in saline habitats. Plant Physiol. Biochem. 2006, 44, 395–408. [Google Scholar] [CrossRef]
- Purmale, L.; Jēkabsone, A.; Andersone-Ozola, U.; Ievinsh, G. Salinity tolerance and ion accumulation potential in vitro and in planta of different Armeria maritima accessions from a dry coastal meadow. Plants 2022, 11, 2570. [Google Scholar] [CrossRef]
- Jēkabsone, A.; Andersone-Ozola, U.; Karlsons, A.; Romanovs, M.; Ievinsh, G. Effect of salinity on growth, ion accumulation and mineral nutrition of different accessions of a crop wild relative legume species, Trifolium fragiferum. Plants 2022, 11, 797. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Amtmann, A. K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Yamada, M.; Kuroda, C.; Fujiyama, H. Function of sodium and potassium in growth of sodium-loving Amaranthaceae species. Soil Sci. Plant Nutr. 2016, 62, 20–26. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salt | Type | Cochlearia officinalis | Limonium sinuatum Blue | Limonium sinuatum Yellow | Lobularia maritima | Plantago maritima | Tripolium pannonicum |
|---|---|---|---|---|---|---|---|
| NaCl | N | 200 | 250 | 250 | 200 | 300 | 300 |
| KCl | N | 200 | 250 | 250 | – | – | – |
| NaOAc | B | 200 | 250 | 250 | – | 300 | 300 |
| KOAc | B | 200 | – | – | – | – | – |
| Na2CO3 | B | 200 | – | – | – | – | – |
| K2CO3 | B | 200 | – | – | – | – | – |
| NaHCO3 | B | 200 | 250 | 250 | 200 | 300 | 300 |
| KHCO3 | B | 200 | – | – | – | – | – |
| NaNO3 | N | 200 | 250 | 250 | 200 | 300 | 300 |
| KNO3 | N | 200 | – | – | – | – | – |
| Na2HPO4 | B | 200 | 250 | 250 | – | – | – |
| K2HPO4 | B | 200 | – | – | – | – | – |
| Na2SO4 | N | 200 | 250 | 250 | 200 | – | – |
| K2SO4 | N | 200 | – | – | – | – | – |
| Treatment | Leaf Number (n) | Leaf Petiole DM (g plant−1) | Leaf Blade DM (g plant−1) | Root DM (g plant−1) | Leaf Petiole H2O (g g−1) | Leaf Blade H2O (g g−1 DM) | Root H2O (g g−1 DM) |
|---|---|---|---|---|---|---|---|
| Control | 65 ± 2 a | 1.5 ± 0.1 ab | 5.7 ± 0.8 a | 0.73 ± 0.15 a | 5.8 ± 0.2 abcd | 4.0 ± 0.3 bc | 6.1 ± 0.6 a |
| NaCl | 64 ± 8 a | 1.0 ± 0.1 ab | 5.0 ± 0.4 a | 0.58 ± 0.02 a | 7.3 ± 0.4 a | 5.9 ± 0.4 a | 5.2 ± 0.1 a |
| KCl | 66 ± 5 a | 1.2 ± 0.2 ab | 5.7 ± 0.6 a | 0.51 ± 0.08 a | 6.7 ± 0.4 ab | 5.1 ± 0.2 abc | 5.5 ± 0.2 a |
| Na2CO3 | 68 ± 10 a | 1.2 ± 0.1 ab | 4.3 ± 0.3 a | 0.78 ± 0.18 a | 6.0 ± 0.1 abcd | 5.3 ± 0.8 ab | 6.5 ± 0.1 a |
| K2CO3 | 52 ± 12 a | 0.9 ± 0.1 b | 4.3 ± 0.3 a | 0.66 ± 0.11 a | 5.5 ± 0.5 bcd | 4.0 ± 0.4 bc | 6.8 ± 0.4 a |
| NaHCO3 | 102 ± 17 a | 1.4 ± 0.2 ab | 4.9 ± 0.4 a | 0.96 ± 0.07 a | 5.5 ± 0.3 bcd | 4.3 ± 0.3 abc | 6.5 ± 0.9 a |
| KHCO3 | 55 ± 9 a | 0.9 ± 0.1 b | 4.4 ± 0.2 a | 0.80 ± 0.18 a | 5.4 ± 0.4 bcd | 4.3 ± 0.3 abc | 5.9 ± 0.7 a |
| NaNO3 | 55 ± 10 a | 0.9 ± 0.1 b | 4.9 ± 0.5 a | 0.51 ± 0.02 a | 4.9 ± 0.3 cd | 4.3 ± 0.4 abc | 5.5 ± 0.5 a |
| KNO3 | 63 ± 8 a | 1.3 ± 0.1 ab | 5.1 ± 0.6 a | 0.39 ± 0.03 a | 4.7 ± 0.3 cd | 4.2 ± 0.3 abc | 5.2 ± 0.6 a |
| Na2HPO4 | 58 ± 7 a | 1.3 ± 0.2 ab | 5.6 ± 0.9 a | 0.62 ± 0.20 a | 4.9 ± 0.1 cd | 3.7 ± 0.5 bc | 4.8 ± 0.2 a |
| K2HPO4 | 76 ± 12 a | 1.3 ± 0.2 ab | 5.0 ± 0.6 a | 0.74 ± 0.03 a | 4.5 ± 0.2 d | 3.4 ± 0.1 c | 4.6 ± 0.4 a |
| Na2SO4 | 85 ± 21 a | 1.7 ± 0.2 a | 4.6 ± 0.5 a | 0.55 ± 0.12 a | 5.6 ± 0.2 bcd | 4.2 ± 0.1 abc | 6.0 ± 0.2 a |
| K2SO4 | 68 ± 6 a | 1.5 ± 0.1 ab | 4.4 ± 0.4 a | 0.84 ± 0.23 a | 6.3 ± 0.7 abc | 4.7 ± 0.4 abc | 6.9 ± 1.2 a |
| Treatment | Leaf Chlorophyll Concentration (mg m−2) | Performance Index Total (Relative Units) | ||||
|---|---|---|---|---|---|---|
| Week 1 | Week 2 | Week 3 | Week 1 | Week 2 | Week 3 | |
| Control | 1216 ± 9 ab | 1191 ± 16 b | 1190 ± 17 c | 3.6 ± 0.3 d | 4.4 ± 0.4 e | 2.9 ± 0.2 e |
| NaCl | 1246 ± 11 ab | 1330 ± 15 a | 1319 ± 14 ab | 9.4 ± 0.5 a | 9.9 ± 0.7 a | 9.8 ± 0.5 a |
| KCl | 1245 ± 22 ab | 1281 ± 26 ab | 1248 ± 27 abc | 9.2 ± 0.5 a | 6.8 ± 0.6 bcd | 7.1 ± 0.3 bc |
| Na2CO3 | 1231 ± 9 ab | 1282 ± 16 ab | 1281 ± 14 abc | 6.7 ± 0.5 bc | 6.6 ± 0.6 bcde | 4.8 ± 0.5 d |
| K2CO3 | 1258 ± 15 ab | 1304 ± 29 a | 1282 ± 26 abc | 7.7 ± 0.7 b | 5.9 ± 0.4 bcde | 5.3 ± 0.4 cd |
| NaHCO3 | 1243 ± 21 ab | 1267 ± 13 ab | 1256 ± 11 abc | 5.3 ± 0.2 cd | 5.6 ± 0.3 cde | 5.3 ± 0.4 cd |
| KHCO3 | 1185 ± 33 b | 1257 ± 30 ab | 1281 ± 32 abc | 6.4 ± 0.5 bc | 4.9 ± 0.3 de | 4.9 ± 0.2 d |
| NaNO3 | 1274 ± 12 a | 1337 ± 10 a | 1291 ± 8 ab | 9.6 ± 0.5 a | 7.9 ± 0.5 ab | 10.0 ± 0.4 a |
| KNO3 | 1263 ± 25 ab | 1347 ± 31 a | 1328 ± 25 a | 7.5 ± 0.5 b | 7.7 ± 0.4 abc | 8.9 ± 0.6 ab |
| Na2HPO4 | 1241 ± 16 ab | 1280 ± 14 ab | 1259 ± 14 abc | 6.7 ± 0.5 bc | 6.6 ± 0.8 bcde | 5.6 ± 0.5 cd |
| K2HPO4 | 1214 ± 10 ab | 1262 ± 24 ab | 1228 ± 22 bc | 5.1 ± 0.4 cd | 5.2 ± 0.3 de | 4.8 ± 0.3 d |
| Na2SO4 | 1246 ± 12 ab | 1321 ± 19 a | 1302 ± 16 ab | 6.5 ± 0.5 bc | 4.7 ± 0.5 de | 4.9 ± 0.4 d |
| K2SO4 | 1236 ± 8 ab | 1305 ± 18 a | 1251 ± 16 abc | 5.2 ± 0.3 cd | 4.9 ± 0.2 de | 4.3 ± 0.3 de |
| Treatment | Leaf Number (n) | Leaf DM (g plant−1) | Root DM (g plant−1) | Flower DM (g plant–1) | Total DM (g plant−1) | Root H2O (g g−1) |
|---|---|---|---|---|---|---|
| Limonium sinuatum Blue | ||||||
| Control | 113 ± 25 a | 14.1 ± 0.8 ab | 3.8 ± 0.3 a | – | 17.8 ± 1.1 ab | 3.8 ± 0.1 a |
| NaCl | 74 ± 8 ab | 10.6 ± 0.5 c | 2.4 ± 0.2 bc | – | 13.1 ± 0.6 c | 4.0 ± 0.1 a |
| KCl | 77 ± 9 ab | 12.5 ± 0.5 bc | 2.4 ± 0.2 bc | – | 14.9 ± 0.6 bc | 4.0 ± 0.1 a |
| NaHCO3 | 40 ± 2 b | 7.9 ± 0.2 d | 1.1 ± 0.1 d | – | 9.0 ± 0.3 d | 2.7 ± 0.5 b |
| NaNO3 | 89 ± 15 ab | 15.7 ± 0.9 a | 2.2 ± 0.1 bc | – | 17.9 ± 0.9 a | 4.5 ± 0.1 a |
| Na2HPO4 | 44 ± 2 b | 8.0 ± 0.2 d | 1.8 ± 0.1 cd | – | 9.8 ± 0.3 d | 2.7 ± 0.1 b |
| Na2SO4 | 89 ± 13 ab | 12.4 ± 0.5 bc | 3.0 ± 0.2 ab | – | 15.4 ± 0.6 abc | 4.4 ± 0.2 a |
| Limonium sinuatum Yellow | ||||||
| Control | 52 ± 11 a | 6.9 ± 0.7 a | 1.8 ± 0.2 ab | 10.5 ± 1.1 a | 19.1 ± 0.7 a | 4.2 ± 0.2 ab |
| NaCl | 56 ± 10 a | 4.5 ± 0.4 a | 2.0 ± 0.3 a | 8.6 ± 0.7 a | 17.1 ± 1.1 ab | 4.7 ± 0.1 a |
| KCl | 38 ± 3 a | 3.8 ± 0.4 a | 0.8 ± 0.1 c | 9.8 ± 0.4 a | 14.5 ± 0.8 b | 4.6 ± 0.9 a |
| NaHCO3 | 36 ± 6 a | 6.0 ± 1.0 a | 0.7 ± 0.1 c | 1.7 ± 0.3 b | 8.6 ± 0.8 c | 2.8 ± 0.3 b |
| NaNO3 | 75 ± 16 a | 6.4 ± 0.9 a | 1.1 ± 0.2 c | 8.0 ± 0.7 a | 15.3 ± 1.2 ab | 4.0 ± 0.1 ab |
| Na2HPO4 | 36 ± 4 a | 6.3 ± 1.1 a | 1.4 ± 0.1 b | 2.6 ± 0.5 b | 9.7 ± 0.9 c | 3.0 ± 0.1 b |
| Na2SO4 | 54 ± 10 a | 5.0 ± 0.5 a | 1.3 ± 0.2 bc | 10.0 ± 1.2 a | 16.3 ± 0.7 ab | 3.6 ± 0.2 ab |
| Treatment | Leaf DM (g plant−1) | Stem DM (g plant−1) | Flower DM (g plant−1) | Root DM (g plant−1) | Total DM (g plant−1) | Leaf H2O (g g−1) |
|---|---|---|---|---|---|---|
| Control | 0.75 ± 0.07 a | 0.69 ± 0.07 a | 0.65 ± 0.12 a | 0.15 ± 0.01 a | 2.24 ± 0.14 a | 9.2 ± 0.4 a |
| NaCl | 0.77 ± 0.12 a | 0.56 ± 0.03 a | 0.33 ± 0.04 b | 0.24 ±0.06 a | 1.89 ± 0.10 a | 10.0 ± 0.6 a |
| NaNO3 | 0.98 ± 0.06 a | 0.61 ± 0.05 a | 0.28 ± 0.03 b | 0.22 ± 0.06 a | 2.08 ± 0.07 a | 9.2 ± 0.9 a |
| NaHCO3 | 0.74 ± 0.05 a | 0.81 ± 0.16 a | 0.39 ± 0.06 b | 0.21 ± 0.02 a | 2.14 ± 0.23 a | 10.2 ± 1.4 a |
| Na2SO4 | 0.84 ± 0.06 a | 0.66 ± 0.13 a | 0.28 ± 0.04 b | 0.24 ± 0.02 a | 2.02 ± 0.24 a | 9.8 ± 0.8 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jēkabsone, A.; Kuļika, J.; Romanovs, M.; Andersone-Ozola, U.; Ievinsh, G. Salt Tolerance and Ion Accumulation in Several Halophytic Plant Species Depending on the Type of Anion. Int. J. Plant Biol. 2023, 14, 1131-1154. https://doi.org/10.3390/ijpb14040083
Jēkabsone A, Kuļika J, Romanovs M, Andersone-Ozola U, Ievinsh G. Salt Tolerance and Ion Accumulation in Several Halophytic Plant Species Depending on the Type of Anion. International Journal of Plant Biology. 2023; 14(4):1131-1154. https://doi.org/10.3390/ijpb14040083
Chicago/Turabian StyleJēkabsone, Astra, Jekaterina Kuļika, Māris Romanovs, Una Andersone-Ozola, and Gederts Ievinsh. 2023. "Salt Tolerance and Ion Accumulation in Several Halophytic Plant Species Depending on the Type of Anion" International Journal of Plant Biology 14, no. 4: 1131-1154. https://doi.org/10.3390/ijpb14040083
APA StyleJēkabsone, A., Kuļika, J., Romanovs, M., Andersone-Ozola, U., & Ievinsh, G. (2023). Salt Tolerance and Ion Accumulation in Several Halophytic Plant Species Depending on the Type of Anion. International Journal of Plant Biology, 14(4), 1131-1154. https://doi.org/10.3390/ijpb14040083