
Colombian Fungal Diversity: Untapped Potential for Diverse Applications

 , , and
, , and 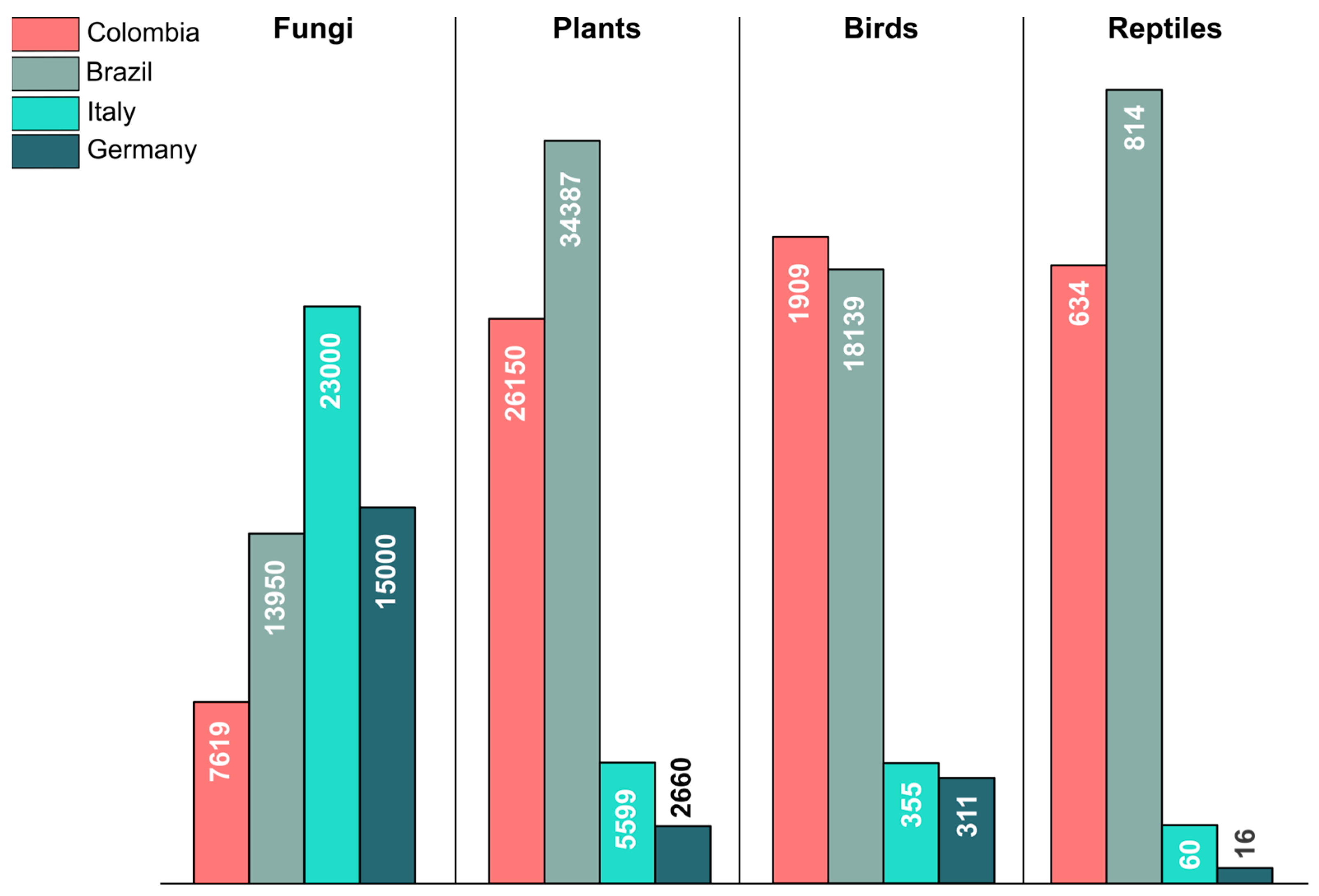{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
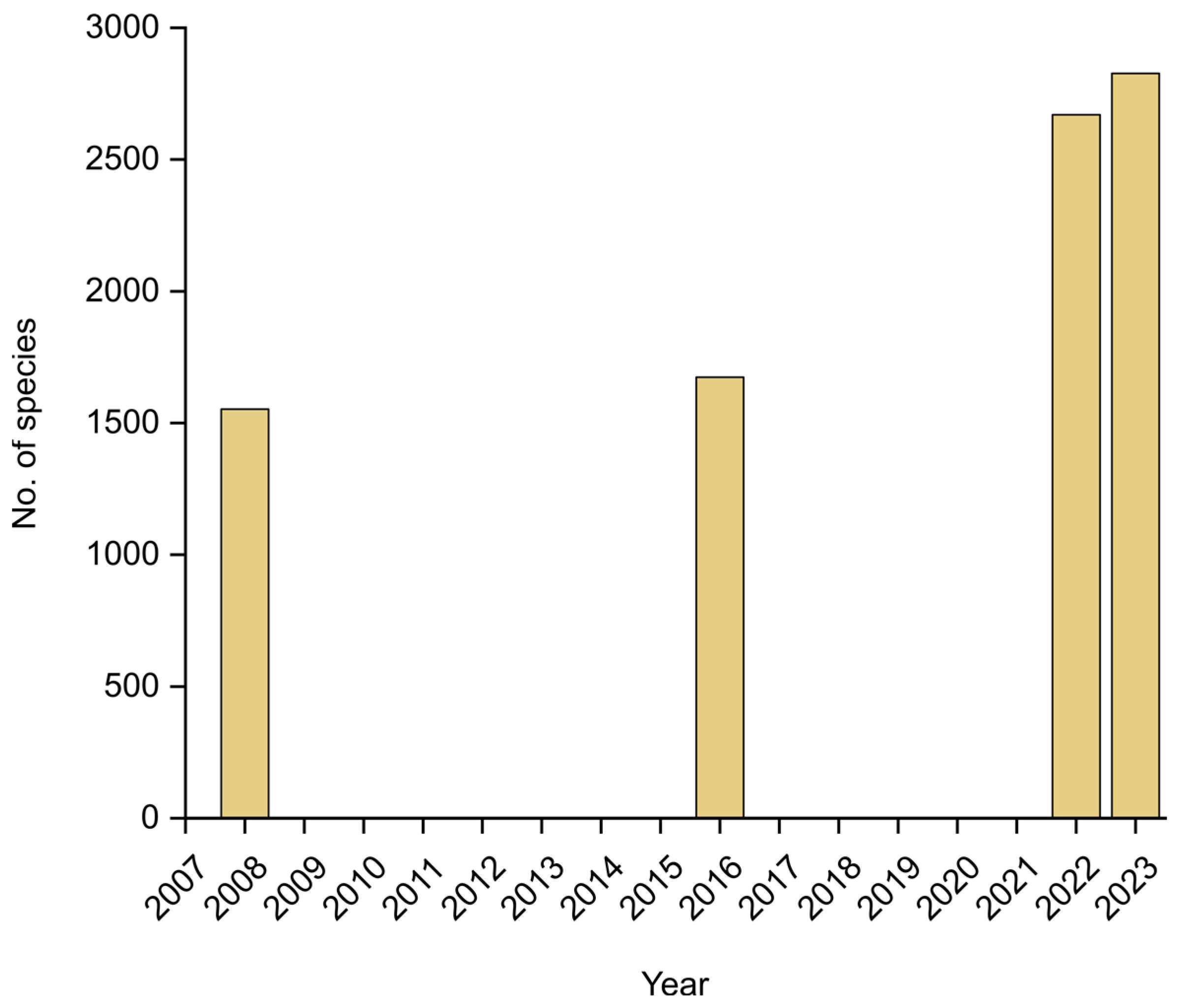
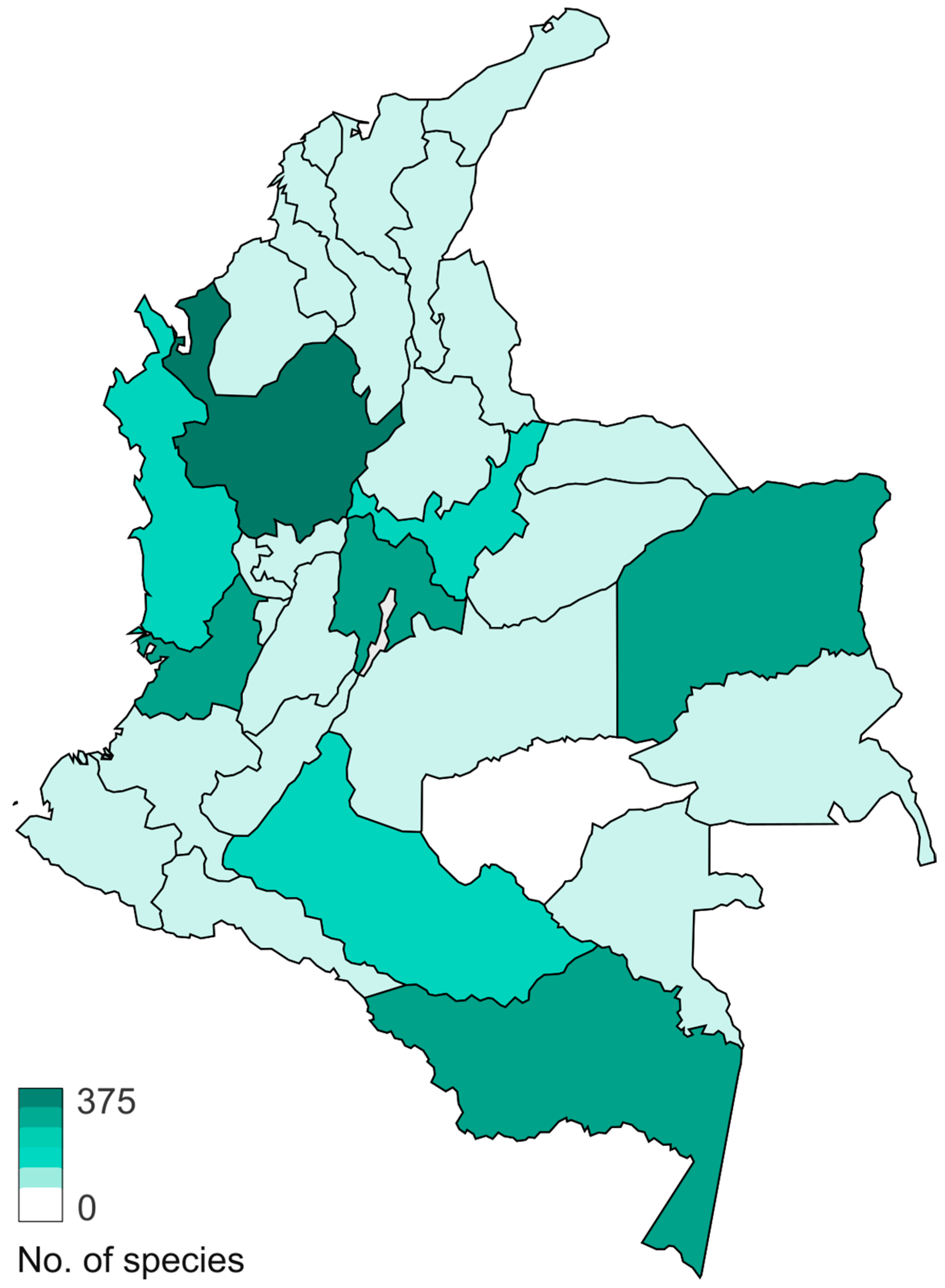
2. Fungal Diversity in Colombia
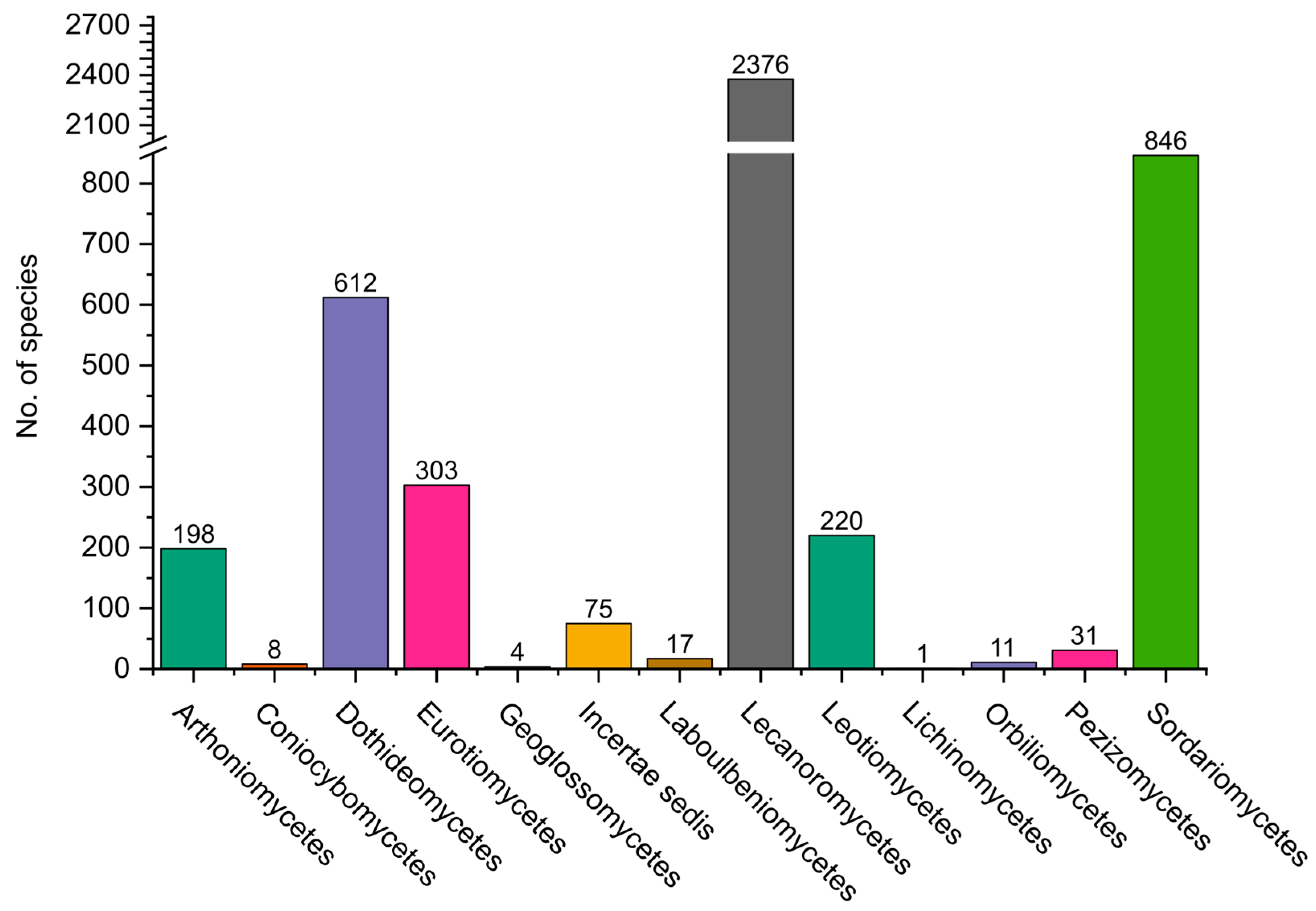
2.1. Ascomycota
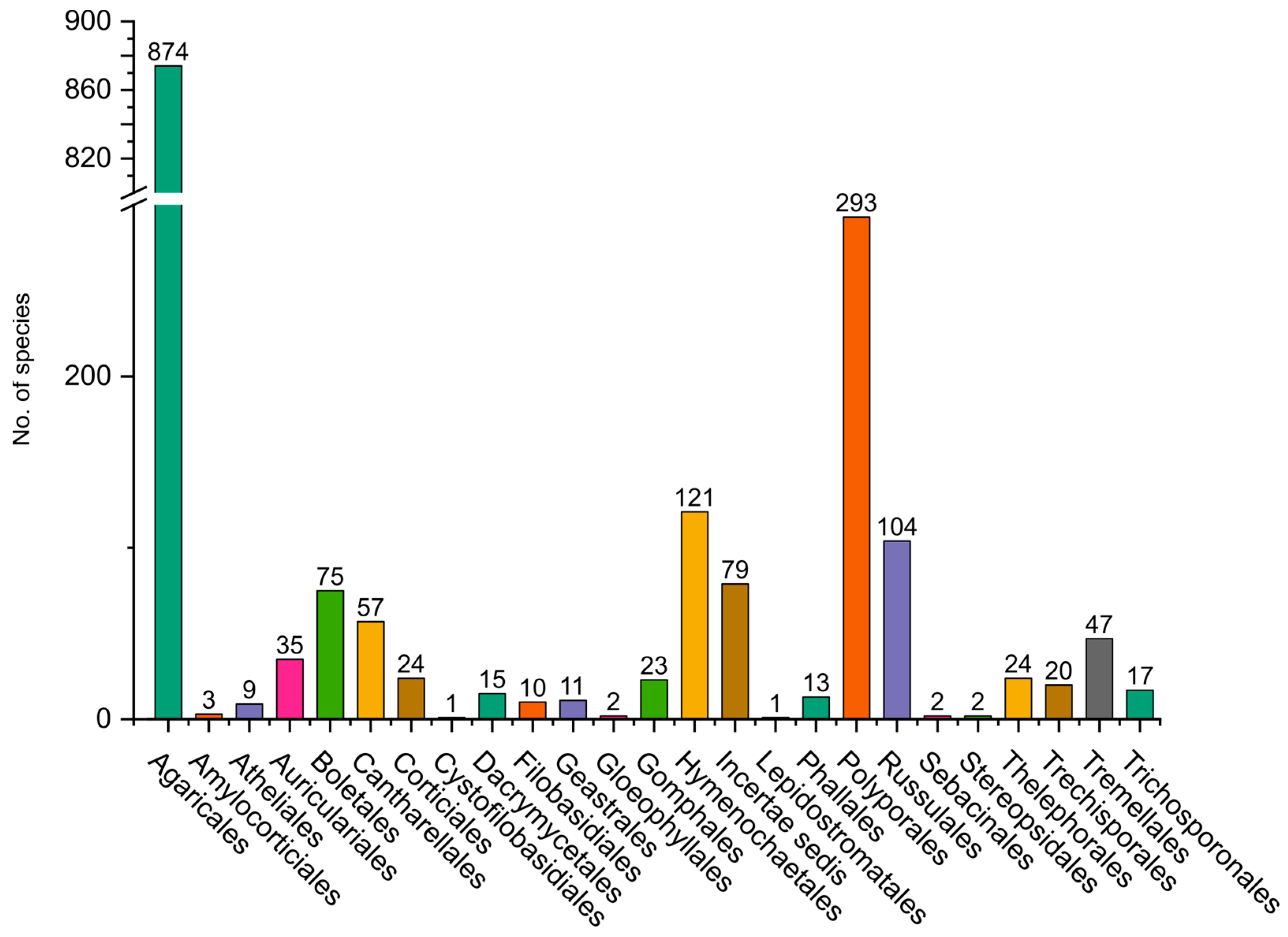
2.2. Basidiomycota
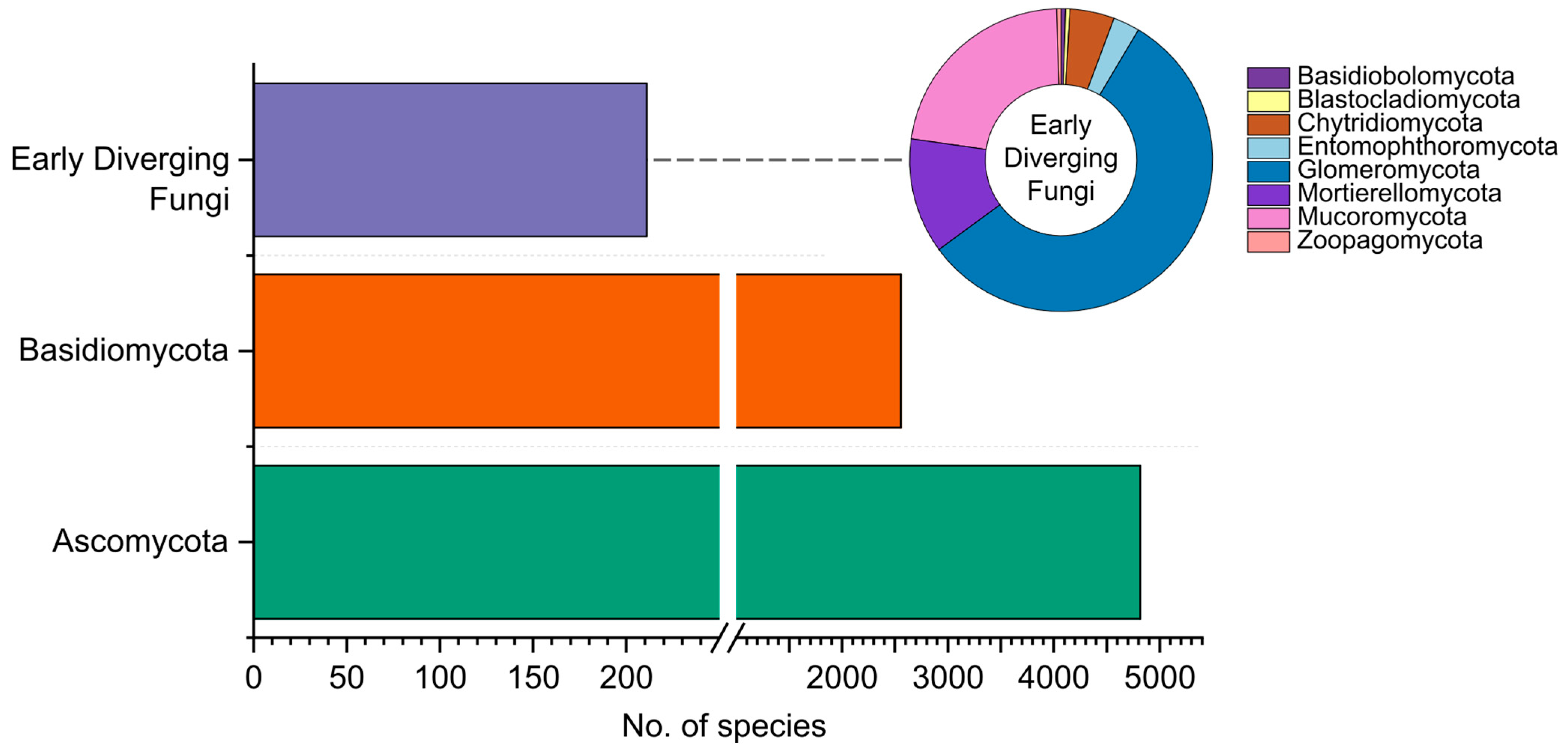
2.3. Early Diverging Fungi
3. Applications of Colombian Fungal Biodiversity
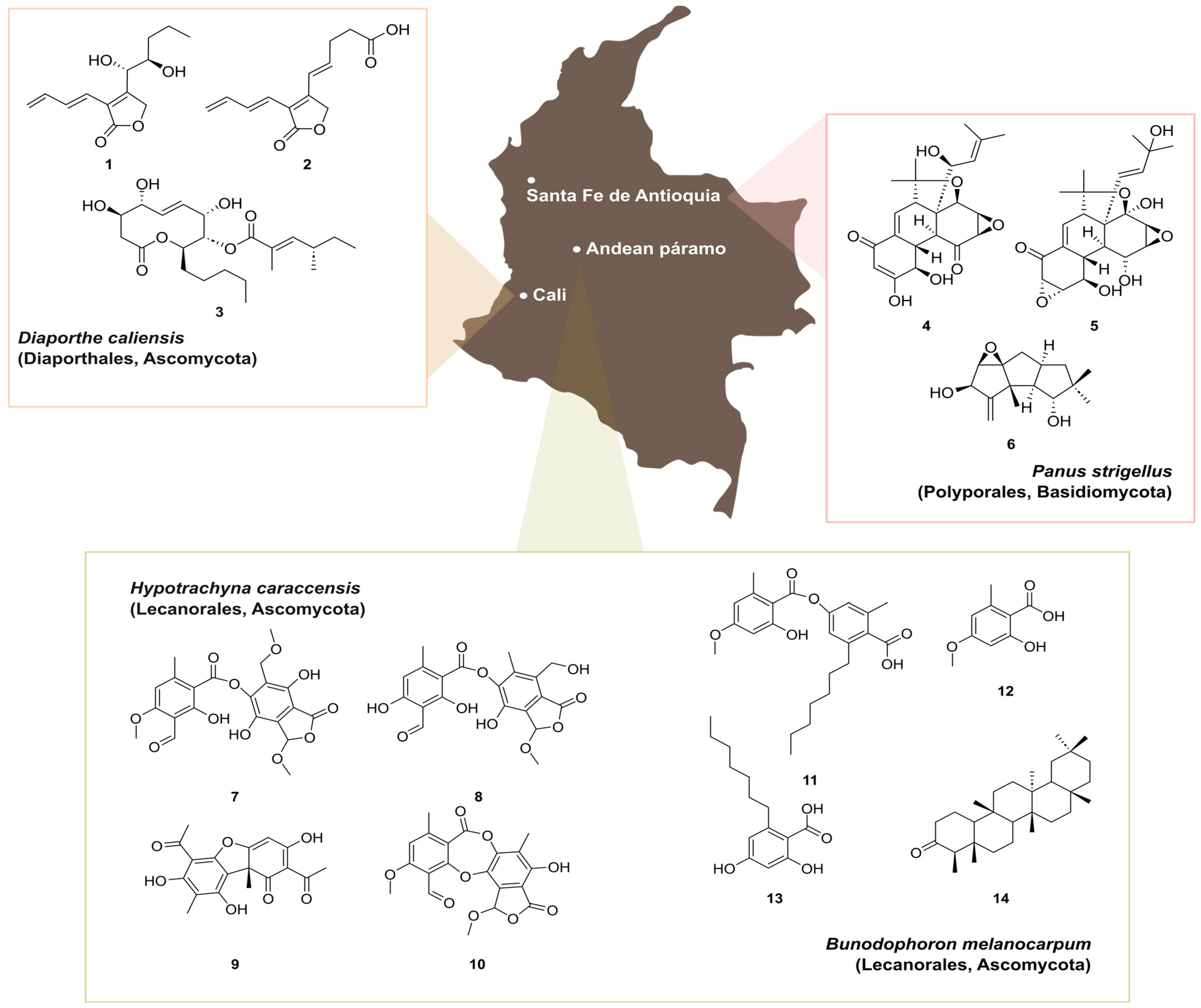
3.1. Natural Products from Colombian Fungi
3.2. Biocontrol Strategies
3.3. Food and Beverages from Fungi
3.4. Other Biotechnological Applications
4. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grace Niego, A.T.; Lambert, C.; Mortimer, P.; Thongklang, N.; Rapior, S.; Grosse, M.; Schrey, H.; Charria-Girón, E.; Hyde, K.D.; Stadler, M. The Contribution of Fungi to the Global Economy. Fungal Divers. 2023, 121, 95–137. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; Hassan, K.; Kemkuignou, B.M.; Čmoková, A.; Surup, F.; Kuhnert, E.; Paomephan, P.; Cheng, T.; de Hoog, S.; et al. Ten Decadal Advances in Fungal Biology Leading towards Human Well-Being. Fungal Divers. 2022, 116, 547–614. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The Amazing Potential of Fungi: 50 Ways We Can Exploit Fungi Industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Gaya, E.; Vasco-Palacios, A.M.; Vargas, N.; Lücking, R.; Carretero, J.; Sanjuan, T.; Moncada, B.; Allkin, B.; Bolaños-Rojas, A.C.; Castellanos-Castro, C.; et al. ColFungi: Colombian Resources for Fungi Made Accessible; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2021; p. 36. [Google Scholar] [CrossRef]
- Butler, R. The Top 10 Most Biodiverse Countries. 2016. Available online: https://news.mongabay.com/2016/05/top-10-biodiverse-countries (accessed on 1 October 2023).
- Butler, R. Countries with the Highest Biodiversity. 2019. Available online: https://rainforests.mongabay.com/03highest_biodiversity.htm (accessed on 1 October 2023).
- de Almeida, R.F.; Lücking, R.; Vasco-Palacios, A.M.; Gaya, E.; Diazgranados, M. (Eds.) Catalogue of Fungi of Colombia; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; p. 541. ISBN 978-1-84246-790-9. [Google Scholar]
- Brown, J.H. Why Are There so Many Species in the Tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Gaya, E.; Motato-Vásquez, V.; Lücking, R. Diversity of Fungi of Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Cossu, T.; Lücking, R.; Vasco-Palacios, A.M.; Moncada, B.; Kirk, P.M.; de Almeida, R.; Gaya, E.; Coca, L.F.; De Souza, J.; Diaz Escandon, D. Annotated Checklist of Fungi of Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Vasco-Palacios, A.M.; Bahram, M.; Boekhout, T.; Tedersoo, L. Carbon Content and PH as Important Drivers of Fungal Community Structure in Three Amazon Forests. Plant Soil 2020, 450, 111–131. [Google Scholar] [CrossRef]
- Vasco-Palacios, A.M.; Hernandez, J.; Peñuela-Mora, M.C.; Franco-Molano, A.E.; Boekhout, T. Ectomycorrhizal Fungi Diversity in a White Sand Forest in Western Amazonia. Fungal Ecol. 2018, 31, 9–18. [Google Scholar] [CrossRef]
- Vasco-Palacios, A.M.; Boekhout, T. Pseudomonotes tropenbosii, an Endemic Dipterocarp Tree from a Neotropical Terra-Firme Forest in Colombian Amazonia that Hosts Ectomycorrhizal Fungi. In Mycorrhizal Fungi in South America. Fungal Biology; Lugo, M.A., Pagano, M.C., Eds.; Springer: Cham, Switzerland, 2022. [Google Scholar] [CrossRef]
- Osorio-Cadavid, E.; Chaves-Lopez, C.; Tofalo, R.; Paparella, A.; Suzzi, G. Detection and Identification of Wild Yeasts in Champús, a Fermented Colombian Maize Beverage. Food Microbiol. 2008, 25, 771–777. [Google Scholar] [CrossRef]
- Salazar Alzate, B.C.; Cortés Rodríguez, M.; Montoya Campuzano, O. Identification of Some Kefir Microorganisms and Optimization of Their Production in Sugarcane Juice. Rev. Fac. Nac. Agron. Medellín 2016, 69, 7935–7943. [Google Scholar] [CrossRef]
- Ordoñez-Burbano, D.E.; Abella-Medina, C.A.; Echeverry-Tamayo, A.; Paz-Lasprilla, L.M.; Benitez-Campo, N. Biodegradación de hidrocarburos alifáticos saturados por microorganismos aislados de suelo contaminado con derivados del petroleo. Rev. Ciencias 2018, 22, 33–44. [Google Scholar] [CrossRef]
- Ramírez-Castrillón, M.; Barona-Colorado, A.; Bados-Lopez, M.C.; Bolaños-Burbano, D. Diversity of Environmental Yeasts of Colombia: A Systematic Review. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Moncada, B.; Coca, L.F.; Díaz-Escandón, D.; Jaramillo-Ciro, M.; Simijaca-Salcedo, D.; Soto-Medina, E.A.; Lücking, R. Diversity, Ecogeography, and Importance of Lichens of Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Correa-Ochoa, M.A.; Vélez-Monsalve, L.C.; Saldarriaga-Molina, J.C. Spatial Distribution of Lichen Communities and Air Pollution Mapping in a Tropical City: Medellín, Colombia. Rev. Biol. Trop. 2021, 69, 1107–1123. [Google Scholar] [CrossRef]
- Abril, M.A.Q.; Ospina, D.M.R.; Rave, M.I.D.; Valencia, J.L. Lichens as biosensors for the evaluation of urban and sub-urban air pollution in a tropical mountain valley, Rionegro, Antioquia. Rev. Bionatura 2021, 6, 1501–1509. [Google Scholar] [CrossRef]
- Albornoz, L.; Torres-Benítez, A.; Moreno-Palacios, M.; Simirgiotis, M.J.; Montoya-Serrano, S.A.; Sepulveda, B.; Stashenko, E.; García-Beltrán, O.; Areche, C. Phylogenetic Studies and Metabolite Analysis of Sticta Species from Colombia and Chile by Ultra-High Performance Liquid Chromatography-High Resolution-Q-Orbitrap-Mass Spectrometry. Metabolites 2022, 12, 156. [Google Scholar] [CrossRef] [PubMed]
- Soto-Medina, E.A.; Díaz, D.; Montaño, J. Biogeography and richness of lichens in Colombia [Biogeografía y riqueza de los líquenes de Colombia]. Rev. Acad. Colomb. Cienc. Exactas Fís. Nat. 2021, 45, 122–135. [Google Scholar] [CrossRef]
- Lasso, E.; Matheus-Arbeláez, P.; Gallery, R.E.; Garzón-López, C.; Cruz, M.; Leon-Garcia, I.V.; Aragón, L.; Ayarza-Páez, A.; Curiel Yuste, J. Homeostatic Response to Three Years of Experimental Warming Suggests High Intrinsic Natural Resistance in the Páramos to Warming in the Short Term. Front. Ecol. Evol. 2021, 9, 615006. [Google Scholar] [CrossRef]
- Moncada, B.; Sipman, H.; Lücking, R. Testing DNA Barcoding in Usnea (Parmeliaceae) in Colombia Using the Internal Transcribed Spacer (ITS). Plant Fungal Syst. 2020, 65, 358–385. [Google Scholar] [CrossRef]
- Moncada, B.; Rincón-Murillo, D.; Lücking, R. Three New Lobarioid Lichens (Lichenized Ascomycota: Peltigeraceae) from Colombia in Memory of Enrique Forero. Rev. Acad. Colomb. Cienc. Exactas Fís. Nat. 2023, 47, 619–640. [Google Scholar] [CrossRef]
- Wilk, K. Calogaya miniata comb. nov., Huneckia crocina comb. nov., and New Neotropical Records of Wetmoreana Brouardii. Mycotaxon 2021, 136, 387–400. [Google Scholar] [CrossRef]
- Simijaca, D.; Lücking, R.; Moncada, B. Two New Species of Astrothelium (Trypetheliaceae) with Amyloid Ascospores Inhabiting the Canopy of Quercus humboldtii Trees in Colombia. Phytotaxa 2021, 508, 229–234. [Google Scholar] [CrossRef]
- Soto-Medina, E.A.; Aptroot, A.; Lücking, R. New Species of Lichen for Colombia Tropical Dry Forest. Cryptogam. Mycol. 2023, 44, 104–107. [Google Scholar] [CrossRef]
- Soto-Medina, E.A. New Records and a Key for Species of Synarthonia (Lichenized Scomycota: Arthoniaceae) in Colombia. Rev. Acad. Colomb. Cienc. Exactas Fís. Nat. 2022, 46, 165–168. [Google Scholar] [CrossRef]
- Ossowska, E.A.; Moncada, B.; Kukwa, M.; Flakus, A.; Rodriguez-Flakus, P.; Olszewska, S.; Lücking, R. New Species of Sticta (Lichenised Ascomycota, Lobarioid Peltigeraceae) from Bolivia Suggest a High Level of Endemism in the Central Andes. MycoKeys 2022, 92, 131–160. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Álvaro-Alba, W.R.; Moncada, B.; Marín-Canchala, N.L.; Tunjano, S.S.; Cárdenas-López, D. Lichens from the Colombian Amazon: 666 Taxa Including 28 New Species and 157 New Country Records Document an Extraordinary Diversity. Bryologist 2023, 126, 242–303. [Google Scholar] [CrossRef]
- Diederich, P.; Millanes, A.M.; Wedin, M.; Lawrey, J. Flora of Lichenicolous Fungi 1; National Museum of Natural History: Luxembourg, 2022. [Google Scholar]
- Sipman, H.J.M.; Hekking, W.; Aguirre, C.J. Checklist of Lichenised and Lichenicolous Fungi from Colombia; Universidad Nacional de Colombia: Bogotá, Colombia, 2008; 242p. [Google Scholar]
- Sipman, H.J.M.; Aguirre, C.; Líquenes, J. Catálogo de Plantas y Líquenes de Colombia; Bernal, R., Gradstein, S.R., Celis, M., Eds.; Universidad Nacional de Colombia, Facultad de Ciencias, Instituto de Ciencias Naturales: Bogotá, Colombia, 2016; Volume 1, pp. 159–281. [Google Scholar]
- Lücking, R.; Rivas Plata, E.; Chaves, J.L.; Umaña, L.; Sipman, H.J.M. How many tropical lichens are there... really? Bibl. Lichenol. 2009, 100, 399–418. [Google Scholar]
- Coca, L.F.; Lumbsch, T.H.; Mercado-Díaz, J.A.; Widhelm, T.J.; Goffinet, B.; Kirika, P.; Lücking, R. Diversity, Phylogeny, and Historical Biogeography of the Genus Coccocarpia (Lichenized Ascomycota: Peltigerales) in the Tropics; Elsevier: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Vasco-Palacios, A.M.; Moncada, B. Two Centuries of Mycological History in Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Traversa, G.; Sosa, A.J.; Chantre, G.R.; Bianchinotti, M.V. Biological Studies of Puccinia Lantanae, a Potential Biocontrol Agent of “Lippia” (Phyla Nodiflora Var. Minor). Agron. Colomb. 2022, 40, 383–394. [Google Scholar] [CrossRef]
- Álvarez-Morales, L.C.; Morales-Osorio, J.G.; Salazar-Yepes, M. First report of Puccinia lagenophorae Cooke on Senecio spp. in Colombia. Can. J. Plant Pathol. 2022, 44, 689–694. [Google Scholar] [CrossRef]
- Cruz-O’Byrne, R.; Gamez-Guzman, A.; Piraneque-Gambasica, N.; Aguirre-Forero, S. Genomic sequencing in Colombian coffee fermentation reveals new records of yeast species. Food Biosci. 2023, 52, 102415. [Google Scholar] [CrossRef]
- Gómez-Montoya, N.; Ríos-Sarmiento, C.; Zora-Vergara, B.; Benjumea-Aristizabal, C.; Santa-Santa, D.J.; Zuluaga-Moreno, M.; Franco-Molano, A.E. Diversidad de macrohongos (Basidiomycota) de Colombia: Listado de especies. Actual. Biol. 2022, 44, 116. [Google Scholar] [CrossRef]
- Pinzón-Osorio, C.A.; Schuster, M.H.B.; Carrero-Torres, C.C. Nuevos registros del género Geastrum (Agaricomycetes, Basidiomycota) para Colombia. Hoehnea 2022, 49, e242021. [Google Scholar] [CrossRef]
- Ramírez-Castrillón, M.; Barona-Colorado, A.; Bados-Lopez, M.C.; Bolaños-Burbano, D. Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Salazar-Yepes, M.; Piepenbring, M. Diversity of Rust and Smut Fungi of Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Peña-Venegas, C.P.; Vasco-Palacios, A.M. Endo- and Ectomycorrhizas in Tropical Ecosystems of Colombia. In Mycorrhizal Fungi in South America. Fungal Biology; Pagano, M., Lugo, M., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Vasco-Palacios, A.M.; Lücking, R.; Moncada, B.; Palacio, M.; Motato-Vásquez, V. A Critical Assessment of Biogeographic Distribution Patterns of Colombian Fungi. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Simijaca, D.; Mueller, G.; Vasco-Palacios, A.M. Fungal Conservation in Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- López-Quintero, C.A.; Straatsma, G.; Franco-Molano, A.E.; Boekhout, T. Macrofungal Diversity in Colombian Amazon Forests Varies with Regions and Regimes of Disturbance. Biodivers. Conserv. 2012, 21, 2221–2243. [Google Scholar] [CrossRef]
- Vasco-Palacios, A.M.; Franco-Molano, A.E.; López-Quintero, C.A.; Boekhout, T. Macromycetes (Ascomycota, Basidiomycota) de la región del medio Caquetá, departamentos de Caquetá y Amazonas (Colombia). Biota Colomb. 2005, 6, 127–140. [Google Scholar]
- Corrales, A.; Benjumea, C.; Gomez-Montoya, N. Diversity, Functional Groups, and Community Structure of Fungi of Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Vargas, C.J.I.; Salazar, E.C.A. Biodiversidad y Mariposas en una Región del Alto Chocó, San José del Palmar, Colombia. Bol. Científico Cent. Mus. Mus. Hist. Nat. 2014, 18, 259–284. Available online: https://revistasojs.ucaldas.edu.co/index.php/boletincientifico/article/view/4480 (accessed on 1 October 2023).
- Bernal, R.; Gradstein, S.R.; Celis, M. (Eds.) Catálogo de Plantas y Líquenes de Colombia; Instituto de Ciencias Naturales, Universidad Nacional de Colombia: Bogotá, Colombia, 2019; Available online: https://catalogoplantasdecolombia.unal.edu.co (accessed on 1 October 2023).
- Pérez-Escobar, O.A.; Lucas, E.; Jaramillo, C.; Monro, A.; Morris, S.K.; Bogarín, D.; Greer, D.; Dodsworth, S.; Aguilar-Cano, J.; Sanchez Meseguer, A.; et al. The Origin and Diversification of the Hyperdiverse Flora in the Chocó Biogeographic Region. Front. Plant. Sci. 2019, 10, 1328. [Google Scholar] [CrossRef] [PubMed]
- Sanjuan, T.; Brothers, K. Diversity of Non-Lichenised Macro-Ascomycota of Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Vasco-Palacios, A.M.; Franco-Molano, A.E. Diversity of Colombian macrofungi (Ascomycota-Basidiomycota). Mycotaxon 2013, 121, 100–158. [Google Scholar]
- Clerici, N.; Armenteras, D.; Kareiva, P.; Botero, R.; Ramírez-Delgado, J.P.; Forero-Medina, G.; Ochoa, J.; Pedraza, C.; Schneider, L.; Lora, C.; et al. Deforestation in Colombian Protected Areas Increased during Post-Conflict Periods. Sci. Rep. 2020, 10, 4971. [Google Scholar] [CrossRef] [PubMed]
- IUCN. The IUCN Red List of Threatened Species, Version 2022-2. 2022. Available online: https://www.iucnredlist.org (accessed on 1 October 2023).
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Doring, M.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Pawłowska, J.; Letcher, P.M.; Kirk, P.M.; Humber, R.A.; Schüßler, A.; Wrzosek, M.; Muszewska, A.; Okrasińska, A.; Istel, Ł.; et al. Notes for Genera: Basal Clades of Fungi (Including Aphelidiomycota, Basidiobolomycota, Blastocladiomycota, Calcarisporiellomycota, Caulochytriomycota, Chytridiomycota, Entomophthoromycota, Glomeromycota, Kickxellomycota, Monoblepharomycota, Mortierellomyc. Fungal Divers. 2018, 92, 43–129. [Google Scholar] [CrossRef]
- Barta, M.; Cagáň, L. Aphid-Pathogenic Entomophthorales (Their Taxonomy, Biology and Ecology). Biologia 2006, 61, S543–S616. [Google Scholar] [CrossRef]
- Morales-López, S.; Ceballos-Garzón, A.; Parra-Giraldo, C.M. Zygomycete Fungi Infection in Colombia: Literature Review. Curr. Fungal Infect. Rep. 2018, 12, 149–154. [Google Scholar] [CrossRef]
- Méndez Puentes, C.A.; Camacho Suarez, J.G.; Echeverry Hernandez, S. Identificación de Bacterias y Hongos en El Aire de Neiva, Colombia. Rev. Salud Pública 2016, 17, 728–737. [Google Scholar] [CrossRef]
- Grossart, H.-P.; Van den Wyngaert, S.; Kagami, M.; Wurzbacher, C.; Cunliffe, M.; Rojas-Jimenez, K. Fungi in Aquatic Ecosystems. Nat. Rev. Microbiol. 2019, 17, 339–354. [Google Scholar] [CrossRef]
- van de Vossenberg, B.T.L.H.; Warris, S.; Nguyen, H.D.T.; van Gent-Pelzer, M.P.E.; Joly, D.L.; van de Geest, H.C.; Bonants, P.J.M.; Smith, D.S.; Lévesque, C.A.; van der Lee, T.A.J. Comparative Genomics of Chytrid Fungi Reveal Insights into the Obligate Biotrophic and Pathogenic Lifestyle of Synchytrium endobioticum. Sci. Rep. 2019, 9, 8672. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Garner, T.W.J. Chytrid Fungi and Global Amphibian Declines. Nat. Rev. Microbiol. 2020, 18, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Flechas, S.V.; Paz, A.; Crawford, A.J.; Sarmiento, C.; Acevedo, A.A.; Arboleda, A.; Bolívar-García, W.; Echeverry-Sandoval, C.L.; Franco, R.; Mojica, C.; et al. Current and Predicted Distribution of the Pathogenic Fungus Batrachochytrium dendrobatidis in Colombia, a Hotspot of Amphibian Biodiversity. Biotropica 2017, 49, 685–694. [Google Scholar] [CrossRef]
- Radek, R.; Wellmanns, D.; Wolf, A. Two New Species of Nephridiophaga (Zygomycota) in the Malpighian Tubules of Cockroaches. Parasitol. Res. 2011, 109, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Gleason, F.H.; Marano, A.V.; Johnson, P.; Martin, W.W. Blastocladian Parasites of Invertebrates. Fungal Biol. Rev. 2010, 24, 56–67. [Google Scholar] [CrossRef]
- Karling, J.S. Zoosporic soil fungi of La Guajiera, Colombia. Nova Hedwig. 1984, 40, 329–340. [Google Scholar]
- Matthews, V.D. A new genus of the Blastocladiaceae. J. Elisha Mitchell Sci. Soc. 1937, 53, 191–195. [Google Scholar]
- Landinez-Torres, A.; Panelli, S.; Picco, A.M.; Comandatore, F.; Tosi, S.; Capelli, E. A Meta-Barcoding Analysis of Soil Mycobiota of the Upper Andean Colombian Agro-Environment. Sci. Rep. 2019, 9, 10085. [Google Scholar] [CrossRef]
- Veerkamp, J.; Gams, W. Los Hongos de Colombia-VIII some new species of soil fungi from Colombia. Caldasia 1983, 13, 709–717. [Google Scholar]
- Gualdrón-Arenas, C.; Suárez-Navarro, A.L.; Valencia-Zapata, H. Hongos del suelo aislados de zonas de vegetación natural del páramo de Chisacá, Colombia. Caldasia 1997, 19, 235–245. [Google Scholar]
- Kirk, P.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Landínez-Torres, A.Y.; Becerra Abril, J.L.; Tosi, S.; Nicola, L. Soil Microfungi of the Colombian Natural Regions. Int. J. Environ. Res. Public Health 2020, 17, 8311. [Google Scholar] [CrossRef] [PubMed]
- Schüßler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef]
- Walker, C.; Schüßler, A.; Vincent, B.; Cranenbrouck, S.; Declerck, S. Anchoring the Species Rhizophagus intraradices (Formerly Glomus Intraradices). Fungal Syst. Evol. 2021, 8, 179–201. [Google Scholar] [CrossRef] [PubMed]
- Charria-Girón, E.; Espinosa, M.C.; Zapata-Montoya, A.; Méndez, M.J.; Caicedo, J.P.; Dávalos, A.F.; Ferro, B.E.; Vasco-Palacios, A.M.; Caicedo, N.H. Evaluation of the Antibacterial Activity of Crude Extracts Obtained from Cultivation of Native Endophytic Fungi Belonging to a Tropical Montane Rainforest in Colombia. Front. Microbiol. 2021, 12, 716523. [Google Scholar] [CrossRef] [PubMed]
- Hoyos, L.V.; Chaves, A.; Grandezz, D.; Medina, A.; Correa, J.; Ramirez-castrillon, M.; Valencia, D.; Caicedo-ortega, N.H. Systematic Screening Strategy for Fungal Laccase Activity of Endophytes from Otoba gracilipes with Bioremediation Potential. Fungal Biol. 2023, 127, 1298–1311. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Carpintero, R.; Romero-Perdomo, F.; Martínez, J.F.; Lewandowski, I. A Review of the Knowledge Base for the Development of Natural Ingredients Value Chains for a Sustainable Biobased Economy in Colombia. Discov. Sustain. 2023, 4, 33. [Google Scholar] [CrossRef]
- Huddart, J.E.A.; Crawford, A.J.; Luna-Tapia, A.L.; Restrepo, S.; Di Palma, F. EBP-Colombia and the Bioeconomy: Genomics in the Service of Biodiversity Conservation and Sustainable Development. Proc. Natl. Acad. Sci. USA 2022, 119, e2115641119. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, A. The Road Ahead: Narratives and Imaginaries of the Value of Biodiversity in Shaping Bioeconomy Policy in Colombia. Tapuya Lat. Am. Sci. Technol. Soc. 2022, 5, 2059137. [Google Scholar] [CrossRef]
- Charria-Girón, E.; Marin-Felixa, Y.; Beutling, U.; Franke, R.; Brönstrup, M.; Vasco-Palacios, A.M.; Caicedo, N.H.; Surup, F. Metabolomics insights into the polyketide-lactones produced by Diaporthe caliensis sp. nov., an endophyte of the medicinal plant Otoba gracilipes. Microbiol. Spectr. 2023, e0274323. [Google Scholar] [CrossRef]
- Llanos-López, N.A.; Ebada, S.S.; Vasco-Palacios, A.M.; Sánchez-Giraldo, L.M.; López, L.; Rojas, L.F.; Mándi, A.; Kurtán, T.; Marin-Felix, Y. Panapophenanthrin, a Rare Oligocyclic Diterpene from Panus strigellus. Metabolites 2023, 13, 848. [Google Scholar] [CrossRef]
- Sekizawa, R.; Ikeno, S.; Nakamura, H.; Naganawa, H.; Matsui, S.; Iinuma, H.; Takeuchi, T. Panepophenanthrin, from a Mushroom Strain, a Novel Inhibitor of the Ubiquitin-Activating Enzyme. J. Nat. Prod. 2002, 65, 1491–1493. [Google Scholar] [CrossRef] [PubMed]
- Moses, J.E.; Commeiras, L.; Baldwin, J.E.; Adlington, R.M. Total Synthesis of Panepophenanthrin. Org. Lett. 2003, 5, 2987–2988. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Johnson, R.P.; Porco, J.A. Total Synthesis of the Ubiquitin-Activating Enzyme Inhibitor (+)-Panepophenanthrin. Angew. Chem. Int. Ed. 2003, 42, 3913–3917. [Google Scholar] [CrossRef] [PubMed]
- Calcott, M.J.; Ackerley, D.F.; Knight, A.; Keyzers, R.A.; Owen, J.G. Secondary Metabolism in the Lichen Symbiosis. Chem. Soc. Rev. 2018, 47, 1730–1760. [Google Scholar] [CrossRef] [PubMed]
- Leal, A.; Rojas, J.L.; Valencia-Islas, N.A.; Castellanos, L. New β-Orcinol Depsides from Hypotrachyna caraccensis, a Lichen from the Páramo Ecosystem and Their Free Radical Scavenging Activity. Nat. Prod. Res. 2018, 32, 1375–1382. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Islas, N.A.; Arguello, J.J.; Rojas, J.L. Antioxidant and Photoprotective Metabolites of Bunodophoron melanocarpum, A Lichen from the Andean Páramo. Pharm. Sci. 2020, 27, 281–290. [Google Scholar] [CrossRef]
- Salgado, F.; Albornoz, L.; Cortéz, C.; Stashenko, E.; Urrea-Vallejo, K.; Nagles, E.; Galicia-Virviescas, C.; Cornejo, A.; Ardiles, A.; Simirgiotis, M.; et al. Secondary Metabolite Profiling of Species of the Genus Usnea by UHPLC-ESI-OT-MS-MS. Molecules 2017, 23, 54. [Google Scholar] [CrossRef]
- Izquierdo-García, L.F.; González-Almario, A.; Cotes, A.M.; Moreno-Velandia, C.A. Trichoderma virens Gl006 and Bacillus velezensis Bs006: A Compatible Interaction Controlling Fusarium Wilt of Cape Gooseberry. Sci. Rep. 2020, 10, 6857. [Google Scholar] [CrossRef]
- García, D.; González-Almario, A.; Cotes, A.M. Controlling Fusarium Wilt of Cape Gooseberry by Microbial Consortia. Lett. Appl. Microbiol. 2023, 76, 72. [Google Scholar] [CrossRef]
- Wang, W.; de Silva, D.D.; Moslemi, A.; Edwards, J.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum Species Causing Anthracnose of Citrus in Australia. J. Fungi 2021, 7, 47. [Google Scholar] [CrossRef]
- Forcelini, B.B.; Peres, N.A. Widespread Resistance to QoI Fungicides of Colletotrichum acutatum from Strawberry Nurseries and Production Fields. Plant Health Prog. 2018, 19, 338–341. [Google Scholar] [CrossRef]
- Muñoz-Guerrero, J.; Guerra-Sierra, B.E.; Alvarez, J.C. Fungal Endophytes of Tahiti Lime (Citrus Citrus × Latifolia) and Their Potential for Control of Colletotrichum acutatum J. H. Simmonds Causing Anthracnose. Front. Bioeng. Biotechnol. 2021, 9, 650351. [Google Scholar] [CrossRef] [PubMed]
- Cardona, N.L.; Franco-Sierra, N.D.; Correa Alvarez, J. Complete Mitogenome of the Biocontroller Fungus Purpureocillium sp. (Ascomycota, Ophiocordycipitaceae, Hypocreales). Mitochondrial DNA Resour. 2018, 3, 1158–1160. [Google Scholar] [CrossRef] [PubMed]
- Robey, M.T.; Caesar, L.K.; Drott, M.T.; Keller, N.P.; Kelleher, N.L. An Interpreted Atlas of Biosynthetic Gene Clusters from 1000 Fungal Genomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2020230118. [Google Scholar] [CrossRef]
- Mira Taborda, Y.D.; Castañeda Sánchez, D.A.; Morales Osorio, J.G.; Patiño Hoyos, L.F. Ecological, phytosanitary, and agronomic aspects of target weeds for biological control studies in Antioquia, Colombia. Acta Agronóm. 2022, 71, 195–206. [Google Scholar] [CrossRef]
- López-Arboleda, W.A.; Ramírez-Castrillón, M. Diversidad de Levaduras Asociadas a Chichas Tradicionales de Colombia. Rev. Colomb. Biotecnol. 2010, 12, 176–186. [Google Scholar]
- Chaves-López, C.; Serio, A.; Grande-Tovar, C.D.; Cuervo-Mulet, R.; Delgado-Ospina, J.; Paparella, A. Traditional Fermented Foods and Beverages from a Microbiological and Nutritional Perspective: The Colombian Heritage. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1031–1048. [Google Scholar] [CrossRef]
- Sandoval-Lozano, C.J.; Caballero-Torres, D.; López-Giraldo, L.J. Screening Wild Yeast Isolated from Cocoa Bean Fermentation Using Volatile Compounds Profile. Molecules 2022, 27, 902. [Google Scholar] [CrossRef]
- Fernández-Niño, M.; Rodríguez-Cubillos, M.J.; Herrera-Rocha, F.; Anzola, J.M.; Cepeda-Hernández, M.L.; Aguirre Mejía, J.L.; Chica, M.J.; Olarte, H.H.; Rodríguez-López, C.; Calderón, D.; et al. Dissecting Industrial Fermentations of Fine Flavour Cocoa through Metagenomic Analysis. Sci. Rep. 2021, 11, 8638. [Google Scholar] [CrossRef]
- Vargas, N.; Gómez-Montoya, N.; Peña-Cañón, R.; Torres-Morales, G. Useful Fungi of Colombia. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]
- Franco–Molano, A.E.; Vasco-Palacios, A.M.; López-Quintero, C.A.; Boekhout, T. Macrohongos de la Región del Medio Caquetá, Colombia; Guía de Campo Multimpresos: Medellín, Colombia, 2005. [Google Scholar]
- Moreno-Bayona, D.A.; Gómez-Méndez, L.D.; Blanco-Vargas, A.; Castillo-Toro, A.; Herrera-Carlosama, L.; Poutou-Piñales, R.A.; Salcedo-Reyes, J.C.; Díaz-Ariza, L.A.; Castillo-Carvajal, L.C.; Rojas-Higuera, N.S.; et al. Simultaneous Bioconversion of Lignocellulosic Residues and Oxodegradable Polyethylene by Pleurotus ostreatus for Biochar Production, Enriched with Phosphate Solubilizing Bacteria for Agricultural Use. PLoS ONE 2019, 14, e0217100. [Google Scholar] [CrossRef]
- Copete-Pertuz, L.S.; Alandete-Novoa, F.; Plácido, J.; Correa-Londoño, G.A.; Mora-Martínez, A.L. Enhancement of Ligninolytic Enzymes Production and Decolourising Activity in Leptosphaerulina sp. by Co–Cultivation with Trichoderma viride and Aspergillus terreus. Sci. Total Environ. 2019, 646, 1536–1545. [Google Scholar] [CrossRef] [PubMed]
- Mejía-Otálvaro, F.; Merino-Restrepo, A.; Hormaza-Anaguano, A. Evaluation of a Trametes pubescens Laccase Concentrated Extract on Allura Red AC Decolorization without the Addition of Synthetic Mediators. J. Environ. Manag. 2021, 285, 112117. [Google Scholar] [CrossRef] [PubMed]
- Merino-Restrepo, A.; Mejía-Otálvaro, F.; Velásquez-Quintero, C.; Hormaza-Anaguano, A. Evaluation of Several White-Rot Fungi for the Decolorization of a Binary Mixture of Anionic Dyes and Characterization of the Residual Biomass as Potential Organic Soil Amendment. J. Environ. Manag. 2020, 254, 109805. [Google Scholar] [CrossRef] [PubMed]
- Caicedo, N.H.; Davalos, A.F.; Puente, P.A.; Rodríguez, A.Y.; Caicedo, P.A. Antioxidant Activity of Exo-Metabolites Produced by Fusarium oxysporum: An Endophytic Fungus Isolated from Leaves of Otoba gracilipes. Microbiologyopen 2019, 8, e903. [Google Scholar] [CrossRef] [PubMed]
- Guerra Sierra, B.E.; Arteaga-Figueroa, L.A.; Sierra-Pelaéz, S.; Alvarez, J.C. Talaromyces santanderensis: A New Cadmium-Tolerant Fungus from Cacao Soils in Colombia. J. Fungi 2022, 8, 1042. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.J.; Caicedo, N.H.; Salamanca, C. Trametes elegans: A Fungal Endophytic Isolate from Otoba gracilipes as Biocatalyst for Natural Flavors Production. N. Biotechnol. 2018, 44, S75. [Google Scholar] [CrossRef]
- Jaramillo, D.A.; Méndez, M.J.; Vargas, G.; Stashenko, E.E.; Vasco-Palacios, A.-M.; Ceballos, A.; Caicedo, N.H. Biocatalytic Potential of Native Basidiomycetes from Colombia for Flavour/Aroma Production. Molecules 2020, 25, 4344. [Google Scholar] [CrossRef]
- Garcia-Cortes, A.; Garcia-Vásquez, J.A.; Aranguren, Y.; Ramirez-Castrillon, M. Pigment Production Improvement in Rhodotorula mucilaginosa AJB01 Using Design of Experiments. Microorganisms 2021, 9, 387. [Google Scholar] [CrossRef]
- Vargas-Sinisterra, A.F.; Ramírez-Castrillón, M. Yeast Carotenoids: Production and Activity as Antimicrobial Biomolecule. Arch. Microbiol. 2021, 203, 873–888. [Google Scholar] [CrossRef]
- Ortiz-Moreno, M.L.; Moncada, B.; Vasco-Palacios, A.M.; de Almeida, R.F.; Gaya, E. Fungi in Colombian and international biological collections. In Catalogue of Fungi of Colombia; De Almeida, R.F., Lücking, R., Vasco-Palacios, A.M., Gaya, E., Diazgranados, M., Eds.; Kew Publishing Royal Botanic Gardens: Richmond, UK, 2022; ISBN 978-1-84246-790-9. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charria-Girón, E.; Vasco-Palacios, A.M.; Moncada, B.; Marin-Felix, Y. Colombian Fungal Diversity: Untapped Potential for Diverse Applications. Microbiol. Res. 2023, 14, 2000-2021. https://doi.org/10.3390/microbiolres14040135
Charria-Girón E, Vasco-Palacios AM, Moncada B, Marin-Felix Y. Colombian Fungal Diversity: Untapped Potential for Diverse Applications. Microbiology Research. 2023; 14(4):2000-2021. https://doi.org/10.3390/microbiolres14040135
Chicago/Turabian StyleCharria-Girón, Esteban, Aída M. Vasco-Palacios, Bibiana Moncada, and Yasmina Marin-Felix. 2023. "Colombian Fungal Diversity: Untapped Potential for Diverse Applications" Microbiology Research 14, no. 4: 2000-2021. https://doi.org/10.3390/microbiolres14040135
APA StyleCharria-Girón, E., Vasco-Palacios, A. M., Moncada, B., & Marin-Felix, Y. (2023). Colombian Fungal Diversity: Untapped Potential for Diverse Applications. Microbiology Research, 14(4), 2000-2021. https://doi.org/10.3390/microbiolres14040135