
An Efficient Peptidomics Screening for Exogenous Substrates and Inhibitory Peptides of the Dipeptidase ACE from Milk Hydrolysate




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Peptide Libraries Generated by Enzymatic Hydrolysis of Skimmed Milk Powders
2.3. ACE Incubation and Stable-Isotope Dimethyl Labeling
2.4. Identification of Substrate Candidates Using LC-MS/MS
2.5. Synthesis of Substrate Candidates and Confirmation of Their Reactivities towards ACE
2.6. Monitoring the ACE Hydrolysis of Substrate Candidates at Different Incubation Times
2.7. ACE Inhibitory Assay of Identified Peptide Substrates
2.8. ACE Hydrolysis towards the Angiotensin I Co-Incubated with Peptide Substrates
2.9. Statistical Analysis
3. Results and Discussion
3.1. Identification of ACE Substrate Candidates from Milk Hydrolysates
3.2. In Vitro Reactivity Confirmation of the ACE’s Substrate Candidates Using Synthetic Peptides
3.3. The Sequence Conservation of ACE’s Exogenous Substrates
3.4. Determination of The ACE Inhibitory (ACEI) Effect and Types for the Identified Substrates
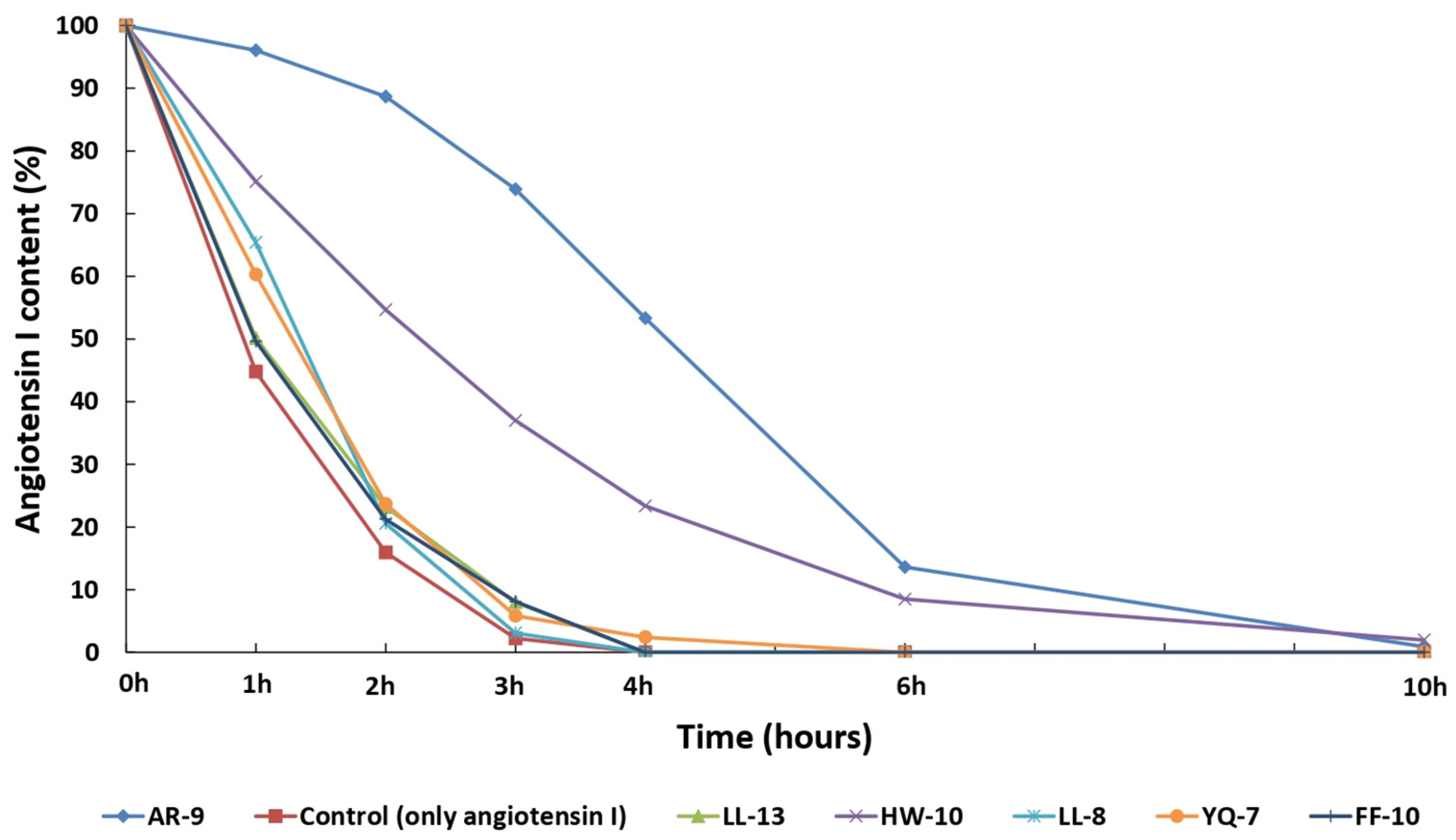
3.5. The Substrates’ Protective Effects on ACE-Mediated Hydrolysis of Angiotensin I
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Overall, C.M.; Blobel, C.P. In search of partners: Linking extracellular proteases to substrates. Nat. Rev. Mol. Cell Biol. 2007, 8, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Yates, N.A.; Deyanova, E.G.; Geissler, W.; Wiener, M.C.; Sachs, J.R.; Wong, K.K.; Thornberry, N.A.; Roy, R.S.; Settlage, R.E.; Hendrickson, R.C. Identification of peptidase substrates in human plasma by FTMS based differential mass spectrometry. Int. J. Mass Spectrom. 2007, 259, 174–183. [Google Scholar] [CrossRef]
- Doupis, J.; Veves, A. DPP4 inhibitors: A new approach in diabetes treatment. Adv. Ther. 2008, 25, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Linz, W.; Wiemer, G.; Gohlke, P.; Unger, T.; Schölkens, B. Contribution of kinins to the cardiovascular actions of angiotensin-converting enzyme inhibitors. Pharmacol. Rev. 1995, 47, 25–49. [Google Scholar] [PubMed]
- Dallas, D.C.; Guerrero, A.; Parker, E.A.; Robinson, R.C.; Gan, J.; German, J.B.; Barile, D.; Lebrilla, C.B. Current peptidomics: Applications, purification, identification, quantification, and functional analysis. Proteomics 2015, 15, 1026–1038. [Google Scholar] [CrossRef] [PubMed]
- Biniossek, M.L.; Niemer, M.; Maksimchuk, K.; Mayer, B.; Fuchs, J.; Huesgen, P.F.; McCafferty, D.G.; Turk, B.; Fritz, G.; Mayer, J. Identification of protease specificity by combining proteome-derived peptide libraries and quantitative proteomics. Mol. Cell. Proteom. 2016, 15, 2515–2524. [Google Scholar] [CrossRef]
- Lone, A.M.; Kim, Y.-G.; Saghatelian, A. Peptidomics methods for the identification of peptidase–substrate interactions. Curr. Opin. Chem. Biol. 2013, 17, 83–89. [Google Scholar] [CrossRef]
- Lone, A.M.; Nolte, W.M.; Tinoco, A.D.; Saghatelian, A. Peptidomics of the prolyl peptidases. AAPS J. 2010, 12, 483–491. [Google Scholar] [CrossRef]
- Wilson, C.H.; Zhang, H.E.; Gorrell, M.D.; Abbott, C.A. Dipeptidyl peptidase 9 substrates and their discovery: Current progress and the application of mass spectrometry-based approaches. Biol. Chem. 2016, 397, 837–856. [Google Scholar] [CrossRef]
- Tinoco, A.D.; Tagore, D.M.; Saghatelian, A. Expanding the dipeptidyl peptidase 4-regulated peptidome via an optimized peptidomics platform. J. Am. Chem. Soc. 2010, 132, 3819–3830. [Google Scholar] [CrossRef]
- Benowitz, N.L. Antihypertensive agents. Basic Clin. Pharmacol. 1998, 39, 153–171. [Google Scholar]
- Skidgel, R.A.; Erdös, E.G. The broad substrate specificity of human angiotensin I converting enzyme. Clin. Exp. Hypertens. A 1987, 9, 243–259. [Google Scholar] [CrossRef]
- Skidgel, R.A.; Engelbrecht, S.; Johnson, A.R.; Erdös, E.G. Hydrolysis of substance P and neurotensin by converting enzyme and neutral endopeptidase. Peptides 1984, 5, 769–776. [Google Scholar] [CrossRef]
- Hemming, M.L.; Selkoe, D.J. Amyloid β-protein is degraded by cellular angiotensin-converting enzyme (ACE) and elevated by an ACE inhibitor. J. Biol. Chem. 2005, 280, 37644–37650. [Google Scholar] [CrossRef]
- Aluko, R. Bioactive peptides. In Functional Foods and Nutraceuticals; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–61. [Google Scholar]
- Hong, S.; Wang, F.L.; Ma, O.; Ef, S.; Dw, C. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme: Importance of the COOH-terminal dipeptide sequence. J. Biol. Chem. 1980, 255, 401–407. [Google Scholar]
- Rohrbach, M.; Williams, E., Jr.; Rolstad, R. Purification and substrate specificity of bovine angiotensin-converting enzyme. J. Biol. Chem. 1981, 256, 225–230. [Google Scholar] [CrossRef]
- Hsu, J.-L.; Huang, S.-Y.; Chow, N.-H.; Chen, S.-H. Stable-isotope dimethyl labeling for quantitative proteomics. Anal. Chem. 2003, 75, 6843–6852. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Shih, Y.-H.; Chen, F.-A.; Wang, L.-F.; Hsu, J.-L. Discovery and study of novel antihypertensive peptides derived from Cassia obtusifolia seeds. J. Agric. Food Chem. 2019, 67, 7810–7820. [Google Scholar] [CrossRef]
- Forghani, B.; Zarei, M.; Ebrahimpour, A.; Philip, R.; Bakar, J.; Hamid, A.A.; Saari, N. Purification and characterization of angiotensin converting enzyme-inhibitory peptides derived from Stichopus horrens: Stability study against the ACE and inhibition kinetics. J. Funct. Foods 2016, 20, 276–290. [Google Scholar] [CrossRef]
- Bersanetti, P.A.; Andrade, M.C.C.; Casarini, D.E.; Juliano, M.A.; Nchinda, A.T.; Sturrock, E.D.; Juliano, L.; Carmona, A.K. Positional-scanning combinatorial libraries of fluorescence resonance energy transfer peptides for defining substrate specificity of the angiotensin I-converting enzyme and development of selective C-domain substrates. Biochemistry 2004, 43, 15729–15736. [Google Scholar] [CrossRef] [PubMed]
- Corvol, P.; Eyries, M.; Soubrier, F. Peptidyl-dipeptidase A/angiotensin I-converting enzyme. In Handbook of Proteolytic Enzymes; Academic Press: Cambridge, MA, USA, 2004; pp. 332–346. [Google Scholar]
- Tu, M.; Wang, C.; Chen, C.; Zhang, R.; Liu, H.; Lu, W.; Jiang, L.; Du, M. Identification of a novel ACE-inhibitory peptide from casein and evaluation of the inhibitory mechanisms. Food Chem. 2018, 256, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Zarei, M.; Abidin, N.B.Z.; Auwal, S.M.; Chay, S.Y.; Abdul Haiyee, Z.; Md Sikin, A.; Saari, N. Angiotensin converting enzyme (ACE)-peptide interactions: Inhibition kinetics, in silico molecular docking and stability study of three novel peptides generated from palm kernel cake proteins. Biomolecules 2019, 9, 569. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | D/H Ratio | Product | Protein |
|---|---|---|---|
| AYFYPELFR | 0.19 | AYFYPEL | CASA1_BOVIN |
| FFVAPFPEVF | 0.87 | FFVAPFPE | |
| HQGLPQEVLNENLLR | 0.76 | HQGLPQEVLNENL | |
| VAPFPEVFG | 0.83 | VAPFPEV | |
| YVPLGTQ | 0.33 | YVPLG | |
| NAVPITPTLNR | 0.61 | NAVPITP | CASA2_BOVIN |
| NAVPITPTL | |||
| HQPHQPLPPTVM | 0.40 | HQPHQPLPPT | CASB_BOVIN |
| IPPLTQT | 0.47 | IPPLT | |
| MFPPQSVL | 0.21 | MFPPQS | |
| VVPPFLQPEVM | 0.80 | VVPPFLQPE | |
| YPVEPFTESQSL | 0.76 | YPVEPFTE | |
| YPVEPFTESQ | |||
| LVYPFPGPIHNSL | 0.26 | LVYPFPGPIHN | |
| INNQFLPYPYY | 0.33 | INNQFLPYP | CASK_BOVIN |
| SRYPSYGLN | 0.81 | SRYPSYG | |
| SAYPGQITSN | 0.65 | SAYPGQIT | TRY1_BOVIN |
| QLDAYPSGAW | 0.47 | QLDAYPSG | CASA1_BOVIN |
| FPQYLQY | 0.12 | FPQYL | CASA2_BOVIN |
| KVIPYVRY | 0.66 | KVIPYV | |
| FALPQYLK | 0.94 | FALPQY | |
| HLPLPLLQSW | 0.12 | HLPLPL | CASB_BOVIN |
| HLPLPLLQ | |||
| AVPYPQR | 0.44 | AVPYP | |
| DMPIQAF | 0.55 | DMPIQ | |
| DMPIQAFL | 0.59 | DMPIQA | |
| LHLPLPLL | 0.74 | LHLP | |
| LHLPLP | |||
| ALPMHIR | 0.15 | ALPMH | LACB_BOVIN |
| HPHPHLSF | 0.15 | HPHPHL | CASK_BOVIN |
| LRPVAAEIY | 0.65 | LRPVAAE | TRFL_BOVIN |
| TTMPLW | 0.15 | TTMP | CASA1_BOVIN |
| ALNEINQFY | 0.84 | ALNEINQ | CASA2_BOVIN |
| QVSLNSGY | 0.56 | QVSLNS | TRY1_BOVIN |
| SIVHPSY | 0.18 | SIVHP |
| Substrate | IC50 (μM) (Regular Assay) | IC50 (μM) (Pre-Incubation) | Type |
|---|---|---|---|
| AR-9 | 6.8 ± 0.23 | 25.3 ± 0.96 | Real substrate |
| HW-10 | 10.6 ± 0.17 | 17.8 ± 0.65 | |
| LL-8 | 12.3 ± 0.37 | 71.9 ± 2.19 | |
| LL-13 | 38.93 ± 2.05 | 27.29 ± 2.16 | Pro-drug |
| FF-10 | 104.4 ± 2.47 | 44.63 ± 3.06 | |
| YQ-7 | 249.6 ± 7.9 | 189.3 ± 9.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.-H.; Nong, N.T.P.; Hsu, J.-L. An Efficient Peptidomics Screening for Exogenous Substrates and Inhibitory Peptides of the Dipeptidase ACE from Milk Hydrolysate. Pharmaceutics 2023, 15, 425. https://doi.org/10.3390/pharmaceutics15020425
Huang J-H, Nong NTP, Hsu J-L. An Efficient Peptidomics Screening for Exogenous Substrates and Inhibitory Peptides of the Dipeptidase ACE from Milk Hydrolysate. Pharmaceutics. 2023; 15(2):425. https://doi.org/10.3390/pharmaceutics15020425
Chicago/Turabian StyleHuang, Ju-Hsuan, Nhung Thi Phuong Nong, and Jue-Liang Hsu. 2023. "An Efficient Peptidomics Screening for Exogenous Substrates and Inhibitory Peptides of the Dipeptidase ACE from Milk Hydrolysate" Pharmaceutics 15, no. 2: 425. https://doi.org/10.3390/pharmaceutics15020425
APA StyleHuang, J.-H., Nong, N. T. P., & Hsu, J.-L. (2023). An Efficient Peptidomics Screening for Exogenous Substrates and Inhibitory Peptides of the Dipeptidase ACE from Milk Hydrolysate. Pharmaceutics, 15(2), 425. https://doi.org/10.3390/pharmaceutics15020425