
A Real-Time Sensing System for Monitoring Neural Network Degeneration in an Alzheimer’s Disease-on-a-Chip Model




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Cortical NSCs
2.2. Biochip Fabrication, Neural Network Formation, and AD-on-a-Chip Establishment
2.3. Cell Viability and Cytotoxicity Assay (WST1 and Lactate Dehydrogenase (LDH) Assay)
2.4. Live/Dead Assay
2.5. Scanning Electron Microscopy (SEM)
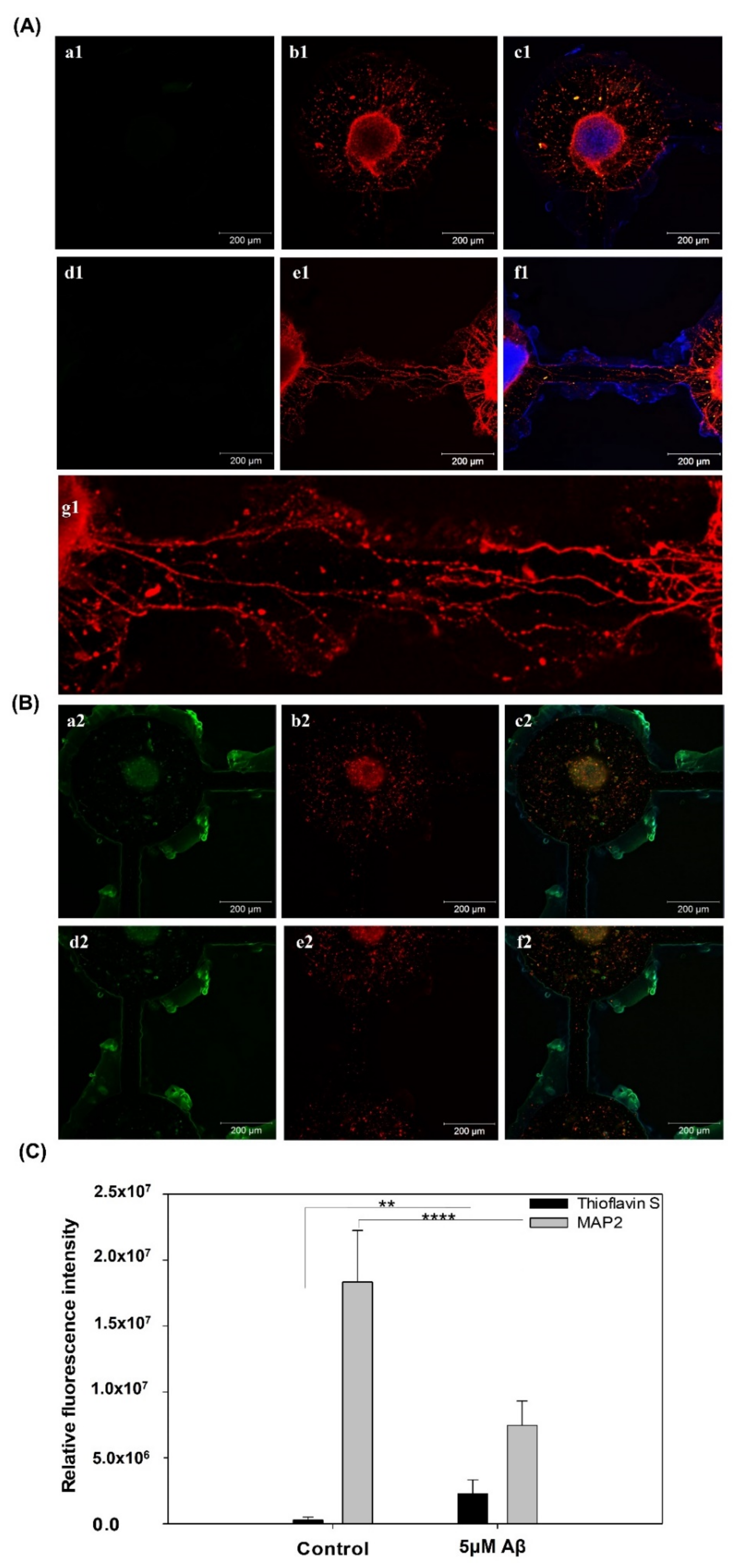
2.6. Immunocytochemistry
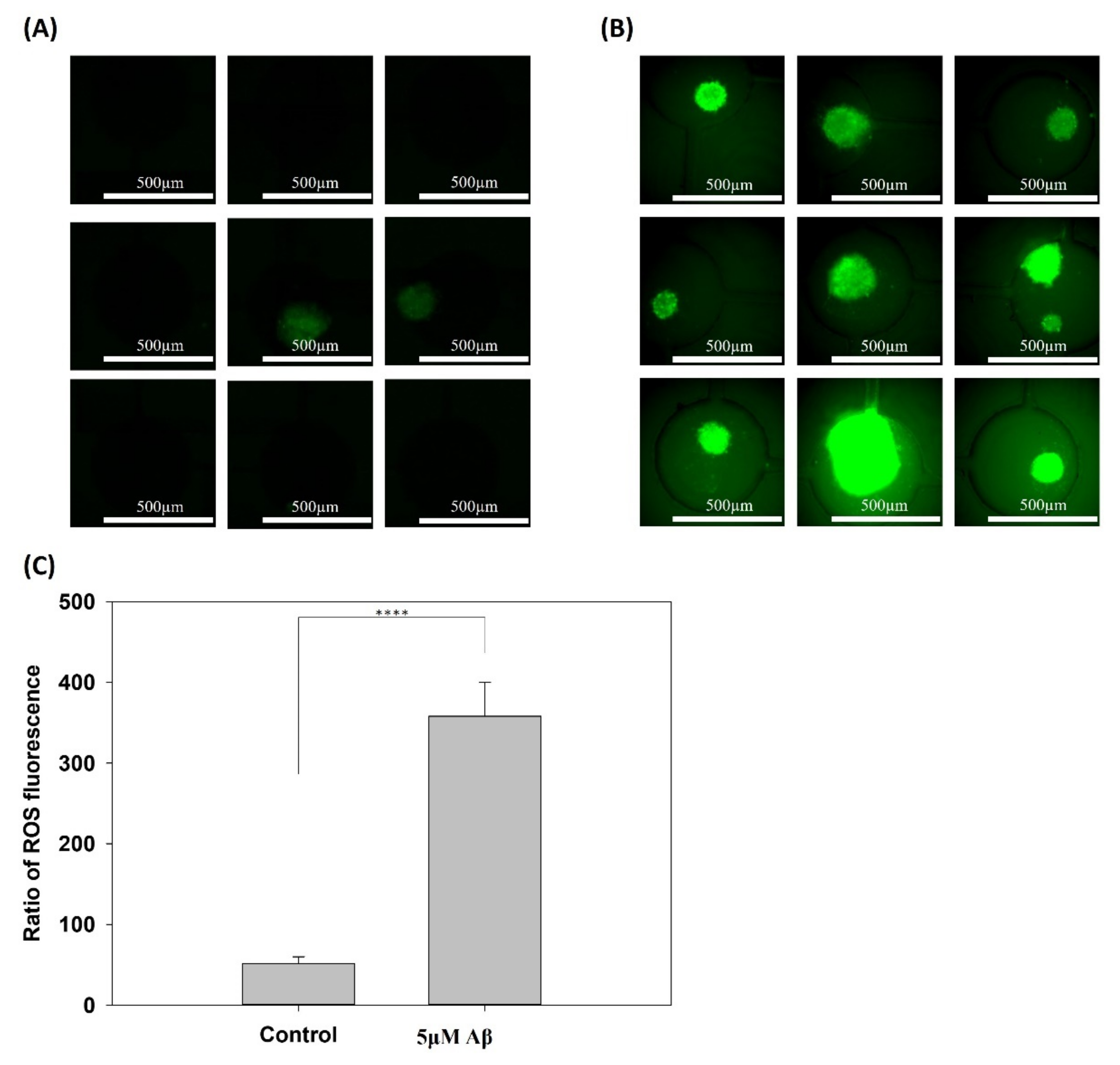
2.7. ROS Detection
2.8. Impedance Measurement of the Neural Network Connections
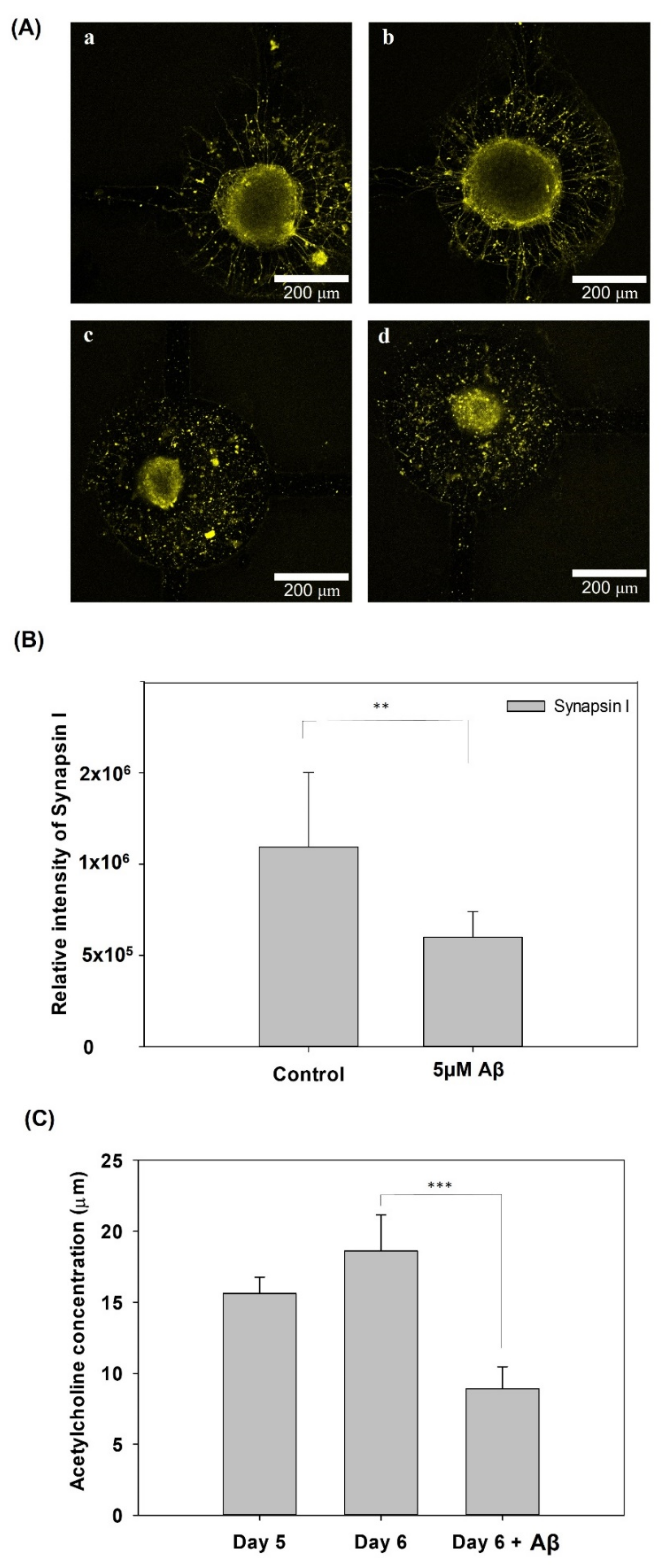
2.9. ACh Assay
2.10. Statistical Analysis
3. Results and Discussion
3.1. Effect of the Aβ Concentration on NSCs
3.2. The Addition of Aβ to the Biochip to Generate an In Vitro AD Model
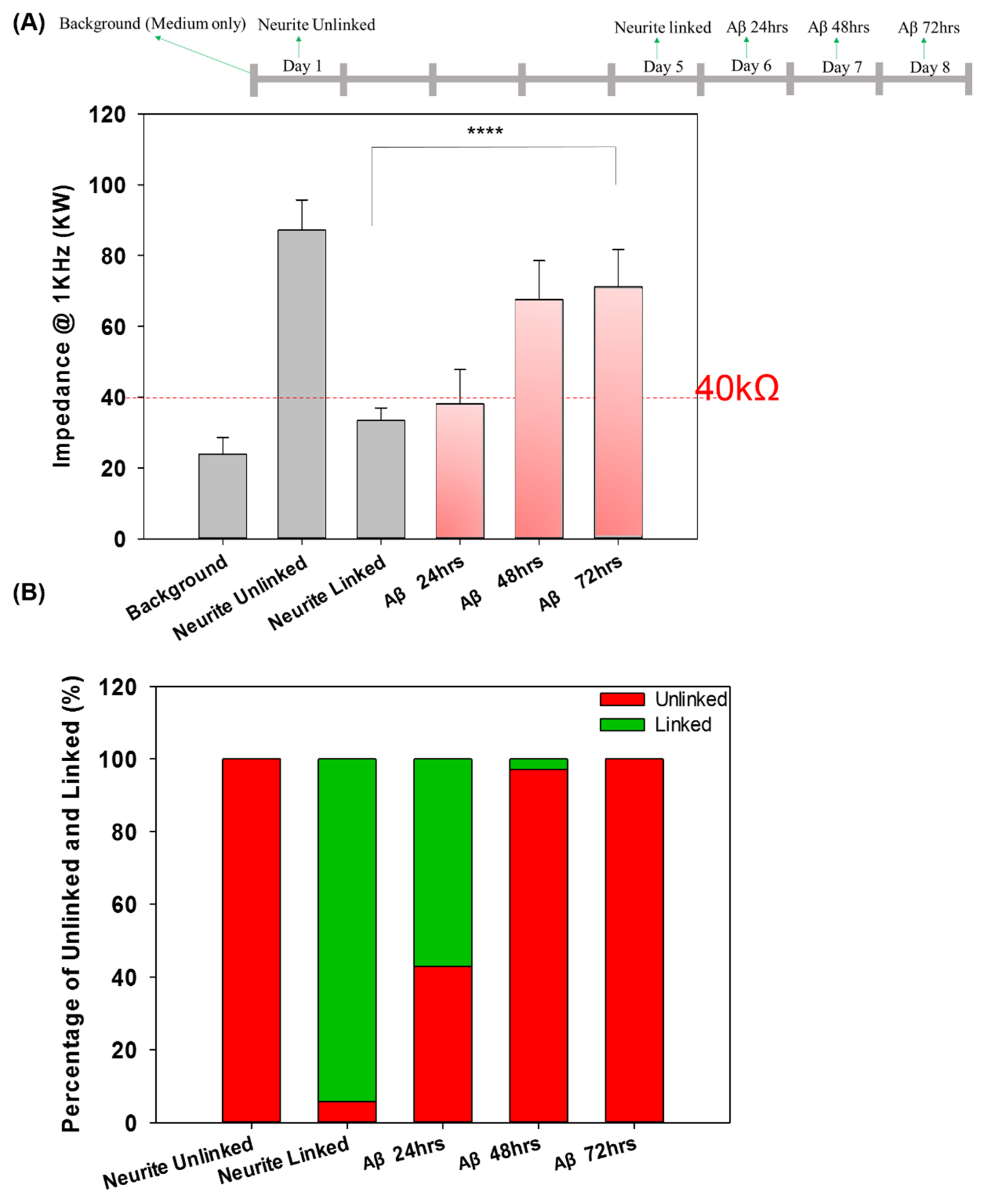
3.3. Real-Time Impedance Analysis Monitors Network Connection and Disconnection
3.4. Synapse Function Determination and ACh Determination
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. 2018 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2018, 14, 367–429. [Google Scholar] [CrossRef]
- Cummings, J.; Lee, G.; Mortsdorf, T.; Ritter, A.; Zhong, K. Alzheimer’s disease drug development pipeline: 2017. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2017, 3, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Estus, S.; Tucker, H.M.; van Rooyen, C.; Wright, S.; Brigham, E.F.; Wogulis, M.; Rydel, R.E. Aggregated Amyloid-β Protein Induces Cortical Neuronal Apoptosis and Concomitant “Apoptotic” Pattern of Gene Induction. J. Neurosci. 1997, 17, 7736. [Google Scholar] [CrossRef] [Green Version]
- Kar, S.; Slowikowski, S.P.M.; Westaway, D.; Mount, H.T.J. Interactions between beta-amyloid and central cholinergic neurons: Implications for Alzheimer’s disease. J. Psychiatry Neurosci. 2004, 29, 427–441. [Google Scholar]
- Desai, N.; Alex, A.; AbdelHafez, F.; Calabro, A.; Goldfarb, J.; Fleischman, A.; Falcone, T. Three-dimensional in vitro follicle growth: Overview of culture models, biomaterials, design parameters and future directions. Reprod. Biol. Endocrinol. RBE 2010, 8, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamir, E.R.; Ewald, A.J. Three-dimensional organotypic culture: Experimental models of mammalian biology and disease. Nat. Reviews. Mol. Cell Biol. 2014, 15, 647–664. [Google Scholar] [CrossRef]
- Matsusaki, M.; Case, C.P.; Akashi, M. Three-dimensional cell culture technique and pathophysiology. Adv. Drug Deliv. Rev. 2014, 74, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Jorfi, M.; d’Avanzo, C.; Kim, D.Y.; Irimia, D. Three-Dimensional Models of the Human Brain Development and Diseases. Adv. Healthc. Mater. 2018, 7, 1700723. [Google Scholar] [CrossRef] [Green Version]
- Luni, C.; Serena, E.; Elvassore, N. Human-on-chip for therapy development and fundamental science. Curr. Opin. Biotechnol. 2014, 25, 45–50. [Google Scholar] [CrossRef]
- Choi, Y.J.; Park, J.; Lee, S.-H. Size-controllable networked neurospheres as a 3D neuronal tissue model for Alzheimer’s disease studies. Biomaterials 2013, 34, 2938–2946. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Pocernich, C.B.; Drake, J. Elevated glutathione as a therapeutic strategy in Alzheimer’s disease. Drug Dev. Res. 2002, 56, 428–437. [Google Scholar] [CrossRef]
- Murphy, R.M. Peptide Aggregation in Neurodegenerative Disease. Annu. Rev. Biomed. Eng. 2002, 4, 155–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitan, G.; Kirkitadze, M.D.; Lomakin, A.; Vollers, S.S.; Benedek, G.B.; Teplow, D.B. Amyloid β-protein (Aβ) assembly: Aβ40 and Aβ42 oligomerize through distinct pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 330–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.-L.; Shim, S.; Kim, D.-H.; Won, S.-H.; Joo, S.; Kim, S.; Jeon, N.L.; Yoon, S.-Y. β-Amyloid is transmitted via neuronal connections along axonal membranes. Ann. Neurol. 2014, 75, 88–97. [Google Scholar] [CrossRef]
- Kunze, A.; Meissner, R.; Brando, S.; Renaud, P. Co-pathological connected primary neurons in a microfluidic device for alzheimer studies. Biotechnol. Bioeng. 2011, 108, 2241–2245. [Google Scholar] [CrossRef]
- Park, J.; Lee, B.K.; Jeong, G.S.; Hyun, J.K.; Lee, C.J.; Lee, S.-H. Three-dimensional brain-on-a-chip with an interstitial level of flow and its application as an in vitro model of Alzheimer’s disease. Lab Chip 2015, 15, 141–150. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; Zhao, P.; Nandakumar, K.; Wang, L.; Song, Y. Microfluidics-Based Systems in Diagnosis of Alzheimer’s Disease and Biomimetic Modeling. Micromachines 2020, 11, 787. [Google Scholar] [CrossRef]
- Choi, Y.J.; Chae, S.; Kim, J.H.; Barald, K.F.; Park, J.Y.; Lee, S.-H. Neurotoxic amyloid beta oligomeric assemblies recreated in microfluidic platform with interstitial level of slow flow. Sci. Rep. 2013, 3, 1921. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.S.; Ryu, J.; Park, C.B. High-Throughput Analysis of Alzheimer’s β-Amyloid Aggregation Using a Microfluidic Self-Assembly of Monomersf. Anal. Chem. 2009, 81, 2751–2759. [Google Scholar] [CrossRef]
- Mai, T.D.; Ferraro, D.; Aboud, N.; Renault, R.; Serra, M.; Tran, N.T.; Viovy, J.-L.; Smadja, C.; Descroix, S.; Taverna, M. Single-step immunoassays and microfluidic droplet operation: Towards a versatile approach for detection of amyloid-β peptide-based biomarkers of Alzheimer’s disease. Sens. Actuators B Chem. 2018, 255, 2126–2135. [Google Scholar] [CrossRef]
- Preische, O.; Schultz, S.A.; Apel, A.; Kuhle, J.; Kaeser, S.A.; Barro, C.; Gräber, S.; Kuder-Buletta, E.; LaFougere, C.; Laske, C.; et al. Serum neurofilament dynamics predicts neurodegeneration and clinical progression in presymptomatic Alzheimer’s disease. Nat. Med. 2019, 25, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, V.D.; Qiu, L.; Tan, E.K.; Zeng, L.; Zhang, Y. Modelling Alzheimer’s disease: Insights from in vivo to in vitro three-dimensional culture platforms. J. Tissue Eng. Regen. Med. 2018, 12, 1944–1958. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-C.; Lee, I.C.; Lei, K.F. Toward the Development of an Artificial Brain on a Micropatterned and Material-Regulated Biochip by Guiding and Promoting the Differentiation and Neurite Outgrowth of Neural Stem/Progenitor Cells. ACS Appl. Mater. Interfaces 2018, 10, 5269–5277. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.C.; Wu, Y.C. Assembly of polyelectrolyte multilayer films on supported lipid bilayers to induce neural stem/progenitor cell differentiation into functional neurons. ACS Appl. Mater. Interfaces 2014, 6, 14439–14450. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, M.; Gearing, A.J.H.; Miller, K.M. A central role for astrocytes in the inflammatory response to β-amyloid; chemokines, cytokines and reactive oxygen species are produced. J. Neuroimmunol. 1999, 93, 182–193. [Google Scholar] [CrossRef]
- Multhaup, G.; Ruppert, T.; Schlicksupp, A.; Hesse, L.; Beher, D.; Masters, C.L.; Beyreuther, K. Reactive oxygen species and Alzheimer’s disease. Biochem. Pharmacol. 1997, 54, 533–539. [Google Scholar] [CrossRef]
- LeVine, H. [18] Quantification of β-sheet amyloid fibril structures with thioflavin T. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 309, pp. 274–284. [Google Scholar]
- Chevy, Q.; Kepecs, A. When Acetylcholine Unlocks Feedback Inhibition in Cortex. Neuron 2018, 97, 481–484. [Google Scholar] [CrossRef] [Green Version]
- Van der Zee, E.A.; Luiten, P.G.M. Muscarinic acetylcholine receptors in the hippocampus, neocortex and amygdala: A review of immunocytochemical localization in relation to learning and memory. Prog. Neurobiol. 1999, 58, 409–471. [Google Scholar] [CrossRef]
- Moreira, F.T.C.; Sale, M.G.F.; di Lorenzo, M. Towards timely Alzheimer diagnosis: A self-powered amperometric biosensor for the neurotransmitter acetylcholine. Biosens. Bioelectron. 2017, 87, 607–614. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.M. Acetylcholine and Choline Amperometric Enzyme Sensors Characterized in Vitro and in Vivo. Anal. Chem. 2004, 76, 1098–1106. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, N.-C.; Liang, C.-C.; Li, Y.-C.E.; Lee, I.-C. A Real-Time Sensing System for Monitoring Neural Network Degeneration in an Alzheimer’s Disease-on-a-Chip Model. Pharmaceutics 2022, 14, 1022. https://doi.org/10.3390/pharmaceutics14051022
Liu N-C, Liang C-C, Li Y-CE, Lee I-C. A Real-Time Sensing System for Monitoring Neural Network Degeneration in an Alzheimer’s Disease-on-a-Chip Model. Pharmaceutics. 2022; 14(5):1022. https://doi.org/10.3390/pharmaceutics14051022
Chicago/Turabian StyleLiu, Nien-Che, Chu-Chun Liang, Yi-Chen Ethan Li, and I-Chi Lee. 2022. "A Real-Time Sensing System for Monitoring Neural Network Degeneration in an Alzheimer’s Disease-on-a-Chip Model" Pharmaceutics 14, no. 5: 1022. https://doi.org/10.3390/pharmaceutics14051022
APA StyleLiu, N.-C., Liang, C.-C., Li, Y.-C. E., & Lee, I.-C. (2022). A Real-Time Sensing System for Monitoring Neural Network Degeneration in an Alzheimer’s Disease-on-a-Chip Model. Pharmaceutics, 14(5), 1022. https://doi.org/10.3390/pharmaceutics14051022