
Advancing Regenerative Cellular Therapies in Non-Scarring Alopecia

 ,
,  ,
,  ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
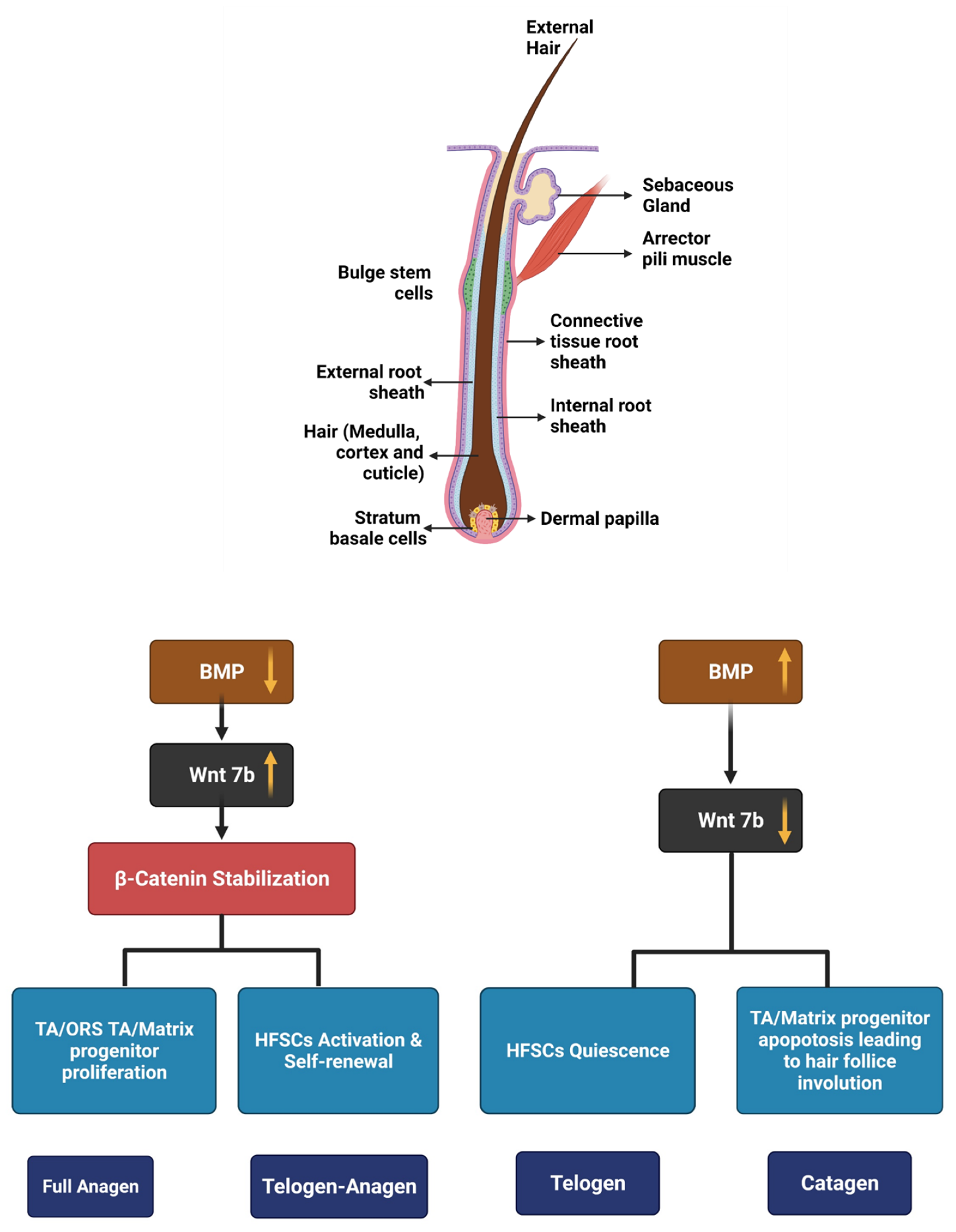
2. Hair Growth Cycle and Its Regulators
3. Treatment of Non-Scarring Alopecia and Adverse Effects
- They have to be continued lifelong, which decreases the compliance;
- The regrowth ceases with the discontinuation of therapy;
- They are associated with several adverse effects leading to temporary morbidity.
- Have unpredictable outcome;
- Have no permanent cure;
- Are associated with significant adverse reactions.
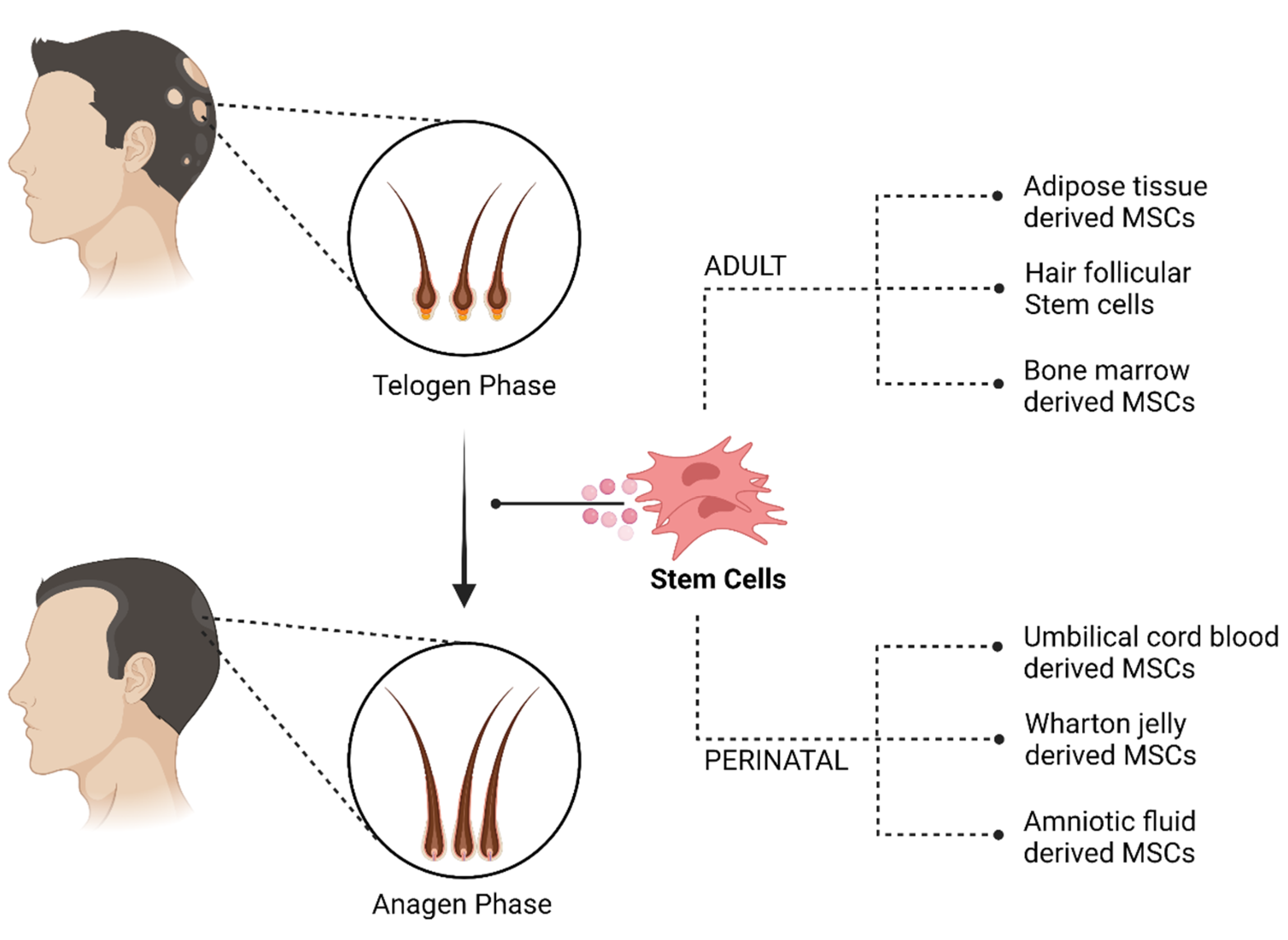
4. Cellular Therapy
4.1. Adult Stem Cells
- MSC must be plastic adherent when maintained under standard culture conditions.
- Expression of CD105, CD73 and CD90, and lack expression of CD45, CD34, CD14, or CD11b, CD79a, or CD19 and HLA-DR surface molecules.
- They must differentiate to osteoblasts, adipocytes and chondroblasts in vitro [62].
4.1.1. Adipose Tissue-Derived Cells
4.1.2. Hair Follicular Stem Cells
4.1.3. Bone Marrow-Derived Cells
4.2. Perinatal MSCs
4.2.1. Umbilical Cord Blood-Derived Cell
4.2.2. Wharton Jelly Derived Cell
4.2.3. Amniotic Fluid-Derived Cell
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, L.; Cook, D.K. Female pattern hair loss. Aust. J. Gen. Pract. 2018, 47, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Aso, T.; Ono, J.; Hosoi, R.; Kaneko, T. Androgenetic Alopecia Treatment in Asian Men. J. Clin. Aesthetic Dermatol. 2018, 11, 32–35. [Google Scholar]
- Jang, W.S.; Son, I.P.; Yeo, I.K.; Park, K.Y.; Li, K.; Kim, B.J.; Seo, S.J.; Kim, M.N.; Hong, C.K. The Annual Changes of Clinical Manifestation of Androgenetic Alopecia Clinic in Korean Males and Females: A Outpatient-Based Study. Ann. Dermatol. 2013, 25, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-C.; Hsieh, F.-N.; Lin, L.-Y.; Hsu, C.-K.; Sheu, H.-M.; Chen, W. Higher body mass index is associated with greater severity of alopecia in men with male-pattern androgenetic alopecia in Taiwan: A cross-sectional study. J. Am. Acad. Dermatol. 2014, 70, 297–302.e1. [Google Scholar] [CrossRef]
- Sinclair, R.; Torkamani, N.; Jones, L. Androgenetic alopecia: New insights into the pathogenesis and mechanism of hair loss. F1000Research 2015, 4, 585. [Google Scholar] [CrossRef]
- Di Loreto, C.; La Marra, F.; Mazzon, G.; Belgrano, E.; Trombetta, C.; Cauci, S. Immunohistochemical Evaluation of Androgen Receptor and Nerve Structure Density in Human Prepuce from Patients with Persistent Sexual Side Effects after Finasteride Use for Androgenetic Alopecia. PLoS ONE 2014, 9, e100237. [Google Scholar] [CrossRef]
- Male Androgenetic Alopecia. Available online: https://Pubmed.Ncbi.Nlm.Nih.Gov/25905192/ (accessed on 8 January 2022).
- Wang, T.; Zhou, C.; Shen, Y.; Wang, X.; Ding, X.; Tian, S.; Liu, Y.; Peng, G.; Xue, S.; Zhou, J.; et al. Prevalence of androgenetic alopecia in China: A community-based study in six cities. Br. J. Dermatol. 2010, 162, 843–847. [Google Scholar] [CrossRef]
- Monselise, A.; Bar-On, R.; Chan, L.; Leibushor, N.; McElwee, K.; Shapiro, J. Examining the Relationship between Alopecia Areata, Androgenetic Alopecia, and Emotional Intelligence. J. Cutan. Med. Surg. 2013, 17, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-H.; Byun, J.-W.; Lee, W.-S.; Kang, H.; Kye, Y.-C.; Kim, K.-H.; Kim, D.-W.; Kim, M.-B.; Kim, S.-J.; Kim, H.-O.; et al. Quality of Life Assessment in Male Patients with Androgenetic Alopecia: Result of a Prospective, Multicenter Study. Ann. Dermatol. 2012, 24, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, S.; Fischer, T.; Chren, M.; Strauss, B.; Elsner, P. Strategies of coping and quality of life in women with alopecia. Br. J. Dermatol. 2001, 144, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, G.H. Review of Human Hair Follicle Biology: Dynamics of Niches and Stem Cell Regulation for Possible Therapeutic Hair Stimulation for Plastic Surgeons. Aesthetic Plast. Surg. 2018, 43, 253–266. [Google Scholar] [CrossRef]
- Neuhaus, K.; Schiestl, C.; Adelsberger, R.; Weibel, L.; Meuli, M.; Böttcher-Haberzeth, S. Bold to do—Bald to be? Outcomes decades after harvesting the scalp in burned children. Burn. 2018, 45, 543–553. [Google Scholar] [CrossRef]
- Almohanna, H.M.; Perper, M.; Tosti, A. Safety concerns when using novel medications to treat alopecia. Expert Opin. Drug Saf. 2018, 17, 1115–1128. [Google Scholar] [CrossRef]
- Morris, R.J.; Liu, Y.; Marles, L.; Yang, Z.; Trempus, C.; Li, S.; Lin, J.; Sawicki, J.; Cotsarelis, G. Capturing and profiling adult hair follicle stem cells. Nat. Biotechnol. 2004, 22, 411–417. [Google Scholar] [CrossRef]
- Tumbar, T.; Guasch, G.; Greco, V.; Blanpain, C.; Lowry, W.E.; Rendl, M.; Fuchs, E. Defining the Epithelial Stem Cell Niche in Skin. Science 2004, 303, 359–363. [Google Scholar] [CrossRef] [Green Version]
- Blanpain, C.; Fuchs, E. Epidermal Stem Cells of the Skin. Annu. Rev. Cell Dev. Biol. 2006, 22, 339–373. [Google Scholar] [CrossRef] [Green Version]
- Blanpain, C.; Lowry, W.E.; Geoghegan, A.; Polak, L.; Fuchs, E. Self-Renewal, Multipotency, and the Existence of Two Cell Populations within an Epithelial Stem Cell Niche. Cell 2004, 118, 635–648. [Google Scholar] [CrossRef]
- Blanpain, C.; Fuchs, E. Epidermal homeostasis: A balancing act of stem cells in the skin. Nat. Rev. Mol. Cell Biol. 2009, 10, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Lowry, W.E.; Blanpain, C.; Nowak, J.A.; Guasch, G.; Lewis, L.; Fuchs, E. Defining the impact of β-catenin/Tcf transactivation on epithelial stem cells. Genes Dev. 2005, 19, 1596–1611. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, R.; Fuchs, E. Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation. Development 1999, 126, 4557–4568. [Google Scholar] [CrossRef]
- Reddy, S.; Andl, T.; Bagasra, A.; Lu, M.M.; Epstein, D.J.; Morrisey, E.E.; Millar, S.E. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech. Dev. 2001, 107, 69–82. [Google Scholar] [CrossRef]
- Blessing, M.; Nanney, L.B.; King, L.; Jones, C.M.; Hogan, B.L. Transgenic mice as a model to study the role of TGF-beta-related molecules in hair follicles. Genes Dev. 1993, 7, 204–215. [Google Scholar] [CrossRef] [Green Version]
- Botchkarev, V.; Botchkareva, N.V.; Roth, W.; Nakamura, M.; Chen, L.-H.; Herzog, W.; Lindner, G.; McMahon, J.A.; Peters, C.; Lauster, R.; et al. Noggin is a mesenchymally derived stimulator of hair-follicle induction. Nat. Cell Biol. 1999, 1, 158–164. [Google Scholar] [CrossRef]
- Kobielak, K.; Stokes, N.; de la Cruz, J.; Polak, L.; Fuchs, E. Loss of a quiescent niche but not follicle stem cells in the absence of bone morphogenetic protein signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 10063–10068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, Y.; Blanco, A.; Tosti, A. Androgenetic Alopecia: An Update of Treatment Options. Drugs 2016, 76, 1349–1364. [Google Scholar] [CrossRef] [PubMed]
- Mella, J.M.; Perret, M.C.; Manzotti, M.; Catalano, H.N.; Guyatt, G. Efficacy and Safety of Finasteride Therapy for Androgenetic Alopecia. Arch. Dermatol. 2010, 146, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Rahimi-Ardabili, B.; Pourandarjani, R.; Habibollahi, P.; Mualeki, A. Finasteride induced depression: A prospective study. BMC Clin. Pharmacol. 2006, 6, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amico, A.V.; Roehrborn, C.G. Effect of 1 mg/day finasteride on concentrations of serum prostate-specific antigen in men with androgenic alopecia: A randomised controlled trial. Lancet Oncol. 2007, 8, 21–25. [Google Scholar] [CrossRef]
- Lachgar, S.; Charveron, M.; Gall, Y.; Bonafe, J.L. Minoxidil upregulates the expression of vascular endothelial growth factor in human hair dermal papilla cells. Br. J. Dermatol. 1998, 138, 407–411. [Google Scholar] [CrossRef]
- Marubayashi, A.; Nakaya, Y.; Fukui, K.; Li, M.; Arase, S. Minoxidil-Induced Hair Growth is Mediated by Adenosine in Cultured Dermal Papilla Cells: Possible Involvement of Sulfonylurea Receptor 2B as a Target of Minoxidil. J. Investig. Dermatol. 2001, 117, 1594–1600. [Google Scholar] [CrossRef] [Green Version]
- FDA. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/97/20834_rogaine%20extra%20strength%20for%20men%205%25_medr.pdf (accessed on 8 January 2022).
- Lucky, A.W.; Piacquadio, D.J.; Ditre, C.M.; Dunlap, F.; Kantor, I.; Pandya, A.G.; Savin, R.C.; Tharp, M.D. A randomized, placebo-controlled trial of 5% and 2% topical minoxidil solutions in the treatment of female pattern hair loss. J. Am. Acad. Dermatol. 2004, 50, 541–553. [Google Scholar] [CrossRef]
- Pratt, C.H.; King, L.E.; Messenger, A.G.; Christiano, A.M.; Sundberg, J.P. Alopecia areata. Nat. Rev. Dis. Prim. 2017, 3, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Walker, S.A.; Rothman, S. A Statistical Study and Consideration of Endocrine Influences. J. Investig. Dermatol. 1950, 14, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Alsantali, A. Alopecia areata: A new treatment plan. Clin. Cosmet. Investig. Dermatol. 2011, 4, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Gentile, P.; Garcovich, S. Autologous activated platelet-rich plasma (AA-PRP) and non-activated (A-PRP) in hair growth: A retrospective, blinded, randomized evaluation in androgenetic alopecia. Expert Opin. Biol. Ther. 2020, 20, 327–337. [Google Scholar] [CrossRef]
- Totey, S.M.; Dhurat, R.S.; Kadam, P.P.; Sevilla, G.P.; Shetty, G. Safety and efficacy of growth factor concentrate in the treatment of nasolabial fold correction: Split face pilot study. Indian J. Dermatol. 2015, 60, 520. [Google Scholar] [CrossRef]
- Fukuoka, H.; Narita, K.; Suga, H. Hair Regeneration Therapy: Application of Adipose-Derived Stem Cells. Curr. Stem Cell Res. Ther. 2017, 12, 531–534. [Google Scholar] [CrossRef]
- Oh, H.A.; Kwak, J.; Kim, B.J.; Jin, H.J.; Park, W.S.; Choi, S.J.; Oh, W.; Um, S. Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth. Cells 2020, 9, 1344. [Google Scholar] [CrossRef]
- Yoon, B.S.; Moon, J.-H.; Jun, E.K.; Kim, J.; Maeng, I.; Kim, J.S.; Lee, J.H.; Baik, C.S.; Kim, A.; Cho, K.S.; et al. Secretory Profiles and Wound Healing Effects of Human Amniotic Fluid–Derived Mesenchymal Stem Cells. Stem Cells Dev. 2010, 19, 887–902. [Google Scholar] [CrossRef]
- Ou, K.-L.; Kuo, Y.-W.; Wu, C.-Y.; Huang, B.-H.; Pai, F.-T.; Chou, H.-H.; Saito, T.; Ueno, T.; Cho, Y.-C.; Huang, M.-S. The Potential of a Hair Follicle Mesenchymal Stem Cell-Conditioned Medium for Wound Healing and Hair Follicle Regeneration. Appl. Sci. 2020, 10, 2646. [Google Scholar] [CrossRef] [Green Version]
- Payushina, O.V.; Butorina, N.N.; Sheveleva, O.N.; Domaratskaya, E.I. Effect of Mesenchymal Stromal Cells and Conditioned Media on Healing of Skin Wound. Bull. Exp. Biol. Med. 2018, 165, 572–575. [Google Scholar] [CrossRef]
- Carrasco, E.; Soto-Heredero, G.; Mittelbrunn, M. The Role of Extracellular Vesicles in Cutaneous Remodeling and Hair Follicle Dynamics. Int. J. Mol. Sci. 2019, 20, 2758. [Google Scholar] [CrossRef] [Green Version]
- Somuncu Özge, S.; Taşlı, P.N.; Şişli, H.B.; Somuncu, S.; Şahin, F. Characterization and Differentiation of Stem Cells Isolated from Human Newborn Foreskin Tissue. Appl. Biochem. Biotechnol. 2015, 177, 1040–1054. [Google Scholar] [CrossRef]
- Yan, H.; Gao, Y.; Ding, Q.; Liu, J.; Li, Y.; Jin, M.; Xu, H.; Ma, S.; Wang, X.; Zeng, W.; et al. Exosomal Micro RNAs Derived from Dermal Papilla Cells Mediate Hair Follicle Stem Cell Proliferation and Differentiation. Int. J. Biol. Sci. 2019, 15, 1368–1382. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.L.; Gangadaran, P.; Bak, S.S.; Oh, J.M.; Kalimuthu, S.; Lee, H.W.; Baek, S.H.; Zhu, L.; Sung, Y.K.; Jeong, S.Y.; et al. Extracellular vesicles derived from MSCs activates dermal papilla cell in vitro and promotes hair follicle conversion from telogen to anagen in mice. Sci. Rep. 2017, 7, 15560. [Google Scholar] [CrossRef] [Green Version]
- Choi, B.Y. Hair-Growth Potential of Ginseng and Its Major Metabolites: A Review on Its Molecular Mechanisms. Int. J. Mol. Sci. 2018, 19, 2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, T.-R.; Oh, C.T.; Choi, E.J.; Park, H.M.; Han, H.J.; Ji, H.J.; Kim, B.J. Human placental extract exerts hair growth-promoting effects through the GSK-3β signaling pathway in human dermal papilla cells. Int. J. Mol. Med. 2015, 36, 1088–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestita, M.; Filoni, A.; Bonamonte, D.; Elia, R.; Giudice, G. Abstract: The Use of Nanofat in Androgenic Alopecia. a Prospective Blinded Study. Plast. Reconstr. Surg.—Glob. Open 2017, 5, 90. [Google Scholar] [CrossRef]
- Epstein, G.K.; Epstein, J.S. Mesenchymal Stem Cells and Stromal Vascular Fraction for Hair Loss: Current Status. Facial Plast. Surg. Clin. N. Am. 2018, 26, 503–511. [Google Scholar] [CrossRef]
- Gentile, P. Autologous Cellular Method Using Micrografts of Human Adipose Tissue Derived Follicle Stem Cells in Androgenic Alopecia. Int. J. Mol. Sci. 2019, 20, 3446. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Liu, X.-M.; Liu, Z.-N.; Wang, Y.; Han, X.; Lian, A.-B.; Mu, Y.; Jin, M.-H.; Liu, J.-Y. Human hair follicle-derived mesenchymal stem cells: Isolation, expansion, and differentiation. World J. Stem Cells 2020, 12, 462–470. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Tsutsui, K.; Taguchi, T.; Sanzen, N.; Nakagawa, A.; Kakiguchi, K.; Yonemura, S.; Tanegashima, C.; Keeley, S.D.; Kiyonari, H.; et al. Hair follicle epidermal stem cells define a niche for tactile sensation. eLife 2018, 7, e38883. [Google Scholar] [CrossRef]
- Chacón-Martínez, C.A.; Klose, M.; Niemann, C.; Glauche, I.; Wickström, S. Hair follicle stem cell cultures reveal self-organizing plasticity of stem cells and their progeny. EMBO J. 2016, 36, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Elmaadawi, I.H.; Mohamed, B.M.; Ibrahim, Z.A.S.; Abdou, S.M.; El Attar, Y.A.; Youssef, A.; Shamloula, M.M.; Taha, A.; Metwally, H.G.; El Afandy, M.M.; et al. Stem cell therapy as a novel therapeutic intervention for resistant cases of alopecia areata and androgenetic alopecia. J. Dermatol. Treat. 2018, 29, 431–440. [Google Scholar] [CrossRef]
- Bak, D.H.; Choi, M.J.; Kim, S.R.; Lee, B.C.; Kim, J.M.; Jeon, E.S.; Oh, W.; Lim, E.S.; Park, B.C.; Kim, M.J.; et al. Human umbilical cord blood mesenchymal stem cells engineered to overexpress growth factors accelerate outcomes in hair growth. Korean J. Physiol. Pharmacol. 2018, 22, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Czarnecka, A.; Odziomek, A.; Murzyn, M.; Dubis, J.; Bagłaj-Oleszczuk, M.; Anita, H.-G. Wharton’s jelly-derived mesenchymal stem cells in the treatment of four patients with alopecia areata. Adv. Clin. Exp. Med. 2021, 30, 211–218. [Google Scholar] [CrossRef]
- Cananzi, M.; Atala, A.; De Coppi, P. Stem cells derived from amniotic fluid: New potentials in regenerative medicine. Reprod. Biomed. Online 2009, 18 (Suppl. 1), 17–27. [Google Scholar] [CrossRef]
- Abd-Allah, S.H.; El-Shal, A.S.; Shalaby, S.M.; Abd-Elbary, E.; Mazen, N.F.; Kader, R.R.A. The role of placenta-derived mesenchymal stem cells in healing of induced full-thickness skin wound in a mouse model. IUBMB Life 2015, 67, 701–709. [Google Scholar] [CrossRef]
- Williams, A.R.; Hare, J.M. Mesenchymal Stem Cells: Biology, Pathophysiology, Translational Findings, and Therapeutic Implications for Cardiac Disease. Circ. Res. 2011, 109, 923–940. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Qin, X.; Jiang, Q.; Nagano, K.; Moriishi, T.; Miyazaki, T.; Komori, H.; Ito, K.; Von Der Mark, K.; Sakane, C.; Kaneko, H.; et al. Runx2 is essential for the transdifferentiation of chondrocytes into osteoblasts. PLoS Genet. 2020, 16, e1009169. [Google Scholar] [CrossRef]
- Caplan, H.; Olson, S.D.; Kumar, A.; George, M.; Prabhakara, K.S.; Wenzel, P.; Bedi, S.; Toledano-Furman, N.E.; Triolo, F.; Kamhieh-Milz, J.; et al. Mesenchymal Stromal Cell Therapeutic Delivery: Translational Challenges to Clinical Application. Front. Immunol. 2019, 10, 1645. [Google Scholar] [CrossRef]
- Asakawa, K.; Toyoshima, K.-E.; Ishibashi, N.; Tobe, H.; Iwadate, A.; Kanayama, T.; Hasegawa, T.; Nakao, K.; Toki, H.; Noguchi, S.; et al. Hair organ regeneration via the bioengineered hair follicular unit transplantation. Sci. Rep. 2012, 2, 424. [Google Scholar] [CrossRef] [Green Version]
- Balañá, M.E. Epidermal stem cells and skin tissue engineering in hair follicle regeneration. World J. Stem Cells 2015, 7, 711–727. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Scioli, M.G.; Bielli, A.; Orlandi, A.; Cervelli, V. Stem cells from human hair follicles: First mechanical isolation for immediate autologous clinical use in androgenetic alopecia and hair loss. Stem Cell Investig. 2017, 4, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquez-Curtis, L.A.; Janowska-Wieczorek, A.; McGann, L.E.; Elliott, J.A. Mesenchymal stromal cells derived from various tissues: Biological, clinical and cryopreservation aspects. Cryobiology 2015, 71, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheykhhasan, M.; Qomi, R.T.; Ghiasi, M. Fibrin Scaffolds Designing in order to Human Adipose-derived Mesenchymal Stem Cells Differentiation to Chondrocytes in the Presence of TGF-β. Int. J. Stem Cells 2015, 8, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, A.; Pham, A.; Cress, M.J.; Pincelli, T.; TerKonda, S.P.; Bruce, A.J.; Zubair, A.C.; Wolfram, J.; Shapiro, S.A. Adipose-derived cellular and cell-derived regenerative therapies in dermatology and aesthetic rejuvenation. Ageing Res. Rev. 2019, 54, 100933. [Google Scholar] [CrossRef]
- Bourin, P.; Bunnell, B.A.; Casteilla, L.; Dominici, M.; Katz, A.J.; March, K.L.; Redl, H.; Rubin, J.P.; Yoshimura, K.; Gimble, J.M. Stromal cells from the adipose tissue-derived stromal vascular fraction and culture expanded adipose tissue-derived stromal/stem cells: A joint statement of the International Federation for Adipose Therapeutics and Science (IFATS) and the International Society for Cellular Therapy (ISCT). Cytotherapy 2013, 15, 641–648. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, E.; Fujita, D.; Takahashi, M.; Oba, S.; Nishimatsu, H. Adipose tissue-derived stem cells as a therapeutic tool for cardiovascular disease. World J. Cardiol. 2015, 7, 454–465. [Google Scholar] [CrossRef]
- Traktuev, D.O.; Prater, D.N.; Merfeld-Clauss, S.; Sanjeevaiah, A.R.; Saadatzadeh, M.R.; Murphy, M.; Johnstone, B.H.; Ingram, D.A.; March, K.L. Robust functional vascular network formation in vivo by cooperation of adipose progenitor and endothelial cells. Circ. Res. 2009, 104, 1410–1420. [Google Scholar] [CrossRef]
- Traktuev, D.O.; Merfeld-Clauss, S.; Li, J.; Kolonin, M.; Arap, W.; Pasqualini, R.; Johnstone, B.H.; March, K.L. A Population of Multipotent CD34-Positive Adipose Stromal Cells Share Pericyte and Mesenchymal Surface Markers, Reside in a Periendothelial Location, and Stabilize Endothelial Networks. Circ. Res. 2008, 102, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Rehman, J.; Traktuev, D.; Li, J.; Merfeld-Clauss, S.; Temm-Grove, C.J.; Bovenkerk, J.E.; Pell, C.L.; Johnstone, B.H.; Considine, R.V.; March, K.L. Secretion of Angiogenic and Antiapoptotic Factors by Human Adipose Stromal Cells. Circulation 2004, 109, 1292–1298. [Google Scholar] [CrossRef]
- Kinnaird, T.; Stabile, E.; Burnett, M.; Lee, C.; Barr, S.; Fuchs, S.; Epstein, S. Marrow-Derived Stromal Cells Express Genes Encoding a Broad Spectrum of Arteriogenic Cytokines and Promote In Vitro and In Vivo Arteriogenesis Through Paracrine Mechanisms. Circ. Res. 2004, 94, 678–685. [Google Scholar] [CrossRef] [Green Version]
- Whiting, D.A. Scalp Biopsy as a Diagnostic and Prognostic Tool in Androgenetic Alopecia. Dermatol. Ther. 1998, 8, 24–33. [Google Scholar]
- Festa, E.; Fretz, J.; Berry, R.; Schmidt, B.; Rodeheffer, M.; Horowitz, M.; Horsley, V. Adipocyte Lineage Cells Contribute to the Skin Stem Cell Niche to Drive Hair Cycling. Cell 2011, 146, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Hausman, G.J.; Martin, R.J. The Development of Adipocytes Located around Hair Follicles in the Fetal Pig. J. Anim. Sci. 1982, 54, 1286–1296. [Google Scholar] [CrossRef]
- Szöke, K.; Brinchmann, J.E. Concise Review: Therapeutic Potential of Adipose Tissue-Derived Angiogenic Cells. Stem Cells Transl. Med. 2012, 1, 658–667. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A Perivascular Origin for Mesenchymal Stem Cells in Multiple Human Organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Perez-Meza, D.; Ziering, C.; Sforza, M.; Krishnan, G.; Ball, E.; Daniels, E. Hair follicle growth by stromal vascular fraction-enhanced adipose transplantation in baldness. Stem Cells Cloning Adv. Appl. 2017, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Anderi, R.; Makdissy, N.; Azar, A.; Rizk, F.; Hamade, A. Cellular therapy with human autologous adipose-derived adult cells of stromal vascular fraction for alopecia areata. Stem Cell Res. Ther. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Aronowitz, J.A.; Lockhart, R.A.; Birnbaum, Z.E.; Hakakian, C.S.; Daniels, E.; Washenik, K. Abstract: Stromal Vascular Fraction Enhanced Adipose Transplantation in Hair Loss Early Experience & Active Phase II FDA Investigation. Plast. Reconstr. Surg. Glob. Open 2016, 4, 50–55. [Google Scholar] [CrossRef]
- Shin, H.; Ryu, H.H.; Kwon, O.; Park, B.-S.; Jo, S.J. Clinical use of conditioned media of adipose tissue-derived stem cells in female pattern hair loss: A retrospective case series study. Int. J. Dermatol. 2015, 54, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Won, C.H.; Chung, W.-K.; Park, B.-S. Up-to-date Clinical Trials of Hair Regeneration Using Conditioned Media of Adipose-Derived Stem Cells in Male and Female Pattern Hair Loss. Curr. Stem Cell Res. Ther. 2017, 12, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Fukuoka, H.; Suga, H. Hair Regeneration Treatment Using Adipose-Derived Stem Cell Conditioned Medium: Follow-up with Trichograms. Eplasty 2015, 15, e10. [Google Scholar] [PubMed]
- Kim, S.J.; Kim, M.J.; Lee, Y.J.; Lee, J.C.; Kim, J.H.; Kim, D.H.; Do, Y.H.; Choi, J.W.; Chung, S.I.; Do, B.-R. Innovative method of alopecia treatment by autologous adipose-derived SVF. Stem Cell Res. Ther. 2021, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Stevens, H.P.; Donners, B.S.; De Bruijn, B.J. Introducing Platelet-Rich Stroma: Platelet-Rich Plasma (PRP) and Stromal Vascular Fraction (SVF) Combined for the Treatment of Androgenetic Alopecia. Aesthetic Surg. J. 2018, 38, 811–822. [Google Scholar] [CrossRef]
- Cho, I.J.; Lui, P.; Obajdin, J.; Riccio, F.; Stroukov, W.; Willis, T.; Spagnoli, F.; Watt, F.M. Mechanisms, Hallmarks, and Implications of Stem Cell Quiescence. Stem Cell Rep. 2019, 12, 1190–1200. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Qian, G.; Bigliardi, P.L.; Ho, B.S.-Y.; Xie, X.-M.; Bigliardi-Qi, M. Aging in hair follicle stem cells and niche microenvironment. J. Dermatol. 2017, 44, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Rompolas, P.; Greco, V. Stem cell dynamics in the hair follicle niche. Semin. Cell Dev. Biol. 2014, 25–26, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Watt, F.M.; Hogan, B.L.M. Out of Eden: Stem Cells and Their Niches. Science 2000, 287, 1427–1430. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Fang, D.; Kumar, S.M.; Li, L.; Nguyen, T.K.; Acs, G.; Herlyn, M.; Xu, X. Isolation of a Novel Population of Multipotent Adult Stem Cells from Human Hair Follicles. Am. J. Pathol. 2006, 168, 1879–1888. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Chovatiya, G.; Tumbar, T. Epigenetic control in skin development, homeostasis and injury repair. Exp. Dermatol. 2019, 28, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Waghmare, S.K.; Bansal, R.; Lee, J.; Zhang, Y.V.; McDermitt, D.J.; Tumbar, T. Quantitative proliferation dynamics and random chromosome segregation of hair follicle stem cells. EMBO J. 2008, 27, 1309–1320. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.V.; Cheong, J.; Ciapurin, N.; McDermitt, D.J.; Tumbar, T. Distinct Self-Renewal and Differentiation Phases in the Niche of Infrequently Dividing Hair Follicle Stem Cells. Cell Stem Cell 2009, 5, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Tumbar, T. Hairy tale of signaling in hair follicle development and cycling. Semin. Cell Dev. Biol. 2012, 23, 906–916. [Google Scholar] [CrossRef] [Green Version]
- Gentile, P.; Scioli, M.G.; Cervelli, V.; Orlandi, A.; Garcovich, S. Autologous Micrografts from Scalp Tissue: Trichoscopic and Long-Term Clinical Evaluation in Male and Female Androgenetic Alopecia. BioMed Res. Int. 2020, 2020, 7397162. [Google Scholar] [CrossRef] [Green Version]
- Ito, M.; Liu, Y.; Yang, Z.; Nguyen, J.; Liang, F.; Morris, R.J.; Cotsarelis, G. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat. Med. 2005, 11, 1351–1354. [Google Scholar] [CrossRef]
- Plikus, M.V.; Mayer, J.A.; De La Cruz, D.; Baker, R.E.; Maini, P.K.; Maxson, R.; Chuong, C.-M. Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration. Nature 2008, 451, 340–344. [Google Scholar] [CrossRef] [Green Version]
- Garza, L.A.; Yang, C.-C.; Zhao, T.; Blatt, H.B.; Lee, M.; He, H.; Stanton, D.C.; Carrasco, L.; Spiegel, J.H.; Tobias, J.W.; et al. Bald scalp in men with androgenetic alopecia retains hair follicle stem cells but lacks CD200-rich and CD34-positive hair follicle progenitor cells. J. Clin. Investig. 2011, 121, 613–622. [Google Scholar] [CrossRef] [Green Version]
- Gentile, P.; Scioli, M.G.; Bielli, A.; De Angelis, B.; De Sio, C.; De Fazio, D.; Ceccarelli, G.; Trivisonno, A.; Orlandi, A.; Cervelli, V.; et al. Platelet-Rich Plasma and Micrografts Enriched with Autologous Human Follicle Mesenchymal Stem Cells Improve Hair Re-Growth in Androgenetic Alopecia. Biomolecular Pathway Analysis and Clinical Evaluation. Biomedicines 2019, 7, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Hao, H.; Xia, L.; Liu, J.; Ti, D.; Tong, C.; Hou, Q.; Han, Q.; Zhao, Y.; Liu, H.; et al. Treatment of MSCs with Wnt1a-conditioned medium activates DP cells and promotes hair follicle regrowth. Sci. Rep. 2015, 4, 5432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, G.-I. Bone marrow-derived stem/stromal cells and adipose tissue-derived stem/stromal cells: Their comparative efficacies and synergistic effects. J. Biomed. Mater. Res. Part A 2017, 105, 2640–2648. [Google Scholar] [CrossRef] [PubMed]
- Chahla, J.; Mannava, S.; Cinque, M.E.; Geeslin, A.G.; Codina, D.; LaPrade, R.F. Bone Marrow Aspirate Concentrate Harvesting and Processing Technique. Arthrosc. Tech. 2017, 6, e441–e445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imam, M.A.; Mahmoud, S.S.; Holton, J.; Abouelmaati, D.; Elsherbini, Y.; Snow, M. A systematic review of the concept and clinical applications of Bone Marrow Aspirate Concentrate in Orthopaedics. SICOT J. 2017, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.B.; Seo, M.-S.; Park, W.T.; Lee, G.W. Bone Marrow Aspirate Concentrate: Its Uses in Osteoarthritis. Int. J. Mol. Sci. 2020, 21, 3224. [Google Scholar] [CrossRef]
- Yoo, B.-Y.; Shin, Y.-H.; Yoon, H.-H.; Seo, Y.-K.; Song, K.-Y.; Park, J.-K. Application of mesenchymal stem cells derived from bone marrow and umbilical cord in human hair multiplication. J. Dermatol. Sci. 2010, 60, 74–83. [Google Scholar] [CrossRef]
- Chung, J.Y.; Song, M.; Ha, C.-W.; Kim, J.-A.; Lee, C.-H.; Park, Y.-B. Comparison of articular cartilage repair with different hydrogel-human umbilical cord blood-derived mesenchymal stem cell composites in a rat model. Stem Cell Res. Ther. 2014, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- El-Badri, N.S.; Hakki, A.; Saporta, S.; Liang, X.; Madhusodanan, S.; Willing, A.E.; Sanberg, C.D.; Sanberg, P.R. Cord Blood Mesenchymal Stem Cells: Potential Use in Neurological Disorders. Stem Cells Dev. 2006, 15, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y. Umbilical cord blood mesenchymal stem cells protect amyloid-β42 neurotoxicityviaparacrine. World J. Stem Cells 2012, 4, 110–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-Y.; Zheng, Z.-H.; Li, X.-Y.; Guo, J.; Zhang, Y.; Li, H.; Wang, Y.-W.; Ren, J.; Wu, Z.-B. Treatment of Foot Disease in Patients with Type 2 Diabetes Mellitus using Human Umbilical Cord Blood Mesenchymal Stem Cells: Response and Correction of Immunological Anomalies. Curr. Pharm. Des. 2013, 19, 4893–4899. [Google Scholar] [CrossRef]
- Seo, Y.; Yang, S.-R.; Jee, M.K.; Joo, E.K.; Roh, K.-H.; Seo, M.-S.; Han, T.H.; Lee, S.Y.; Ryu, P.D.; Jung, J.-W.; et al. Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Protect against Neuronal Cell Death and Ameliorate Motor Deficits in Niemann Pick Type C1 Mice. Cell Transplant. 2011, 20, 1033–1047. [Google Scholar] [CrossRef]
- Pawitan, J.A. Prospect of Stem Cell Conditioned Medium in Regenerative Medicine. BioMed Res. Int. 2014, 2014, 965849. [Google Scholar] [CrossRef] [Green Version]
- Di Santo, S.; Yang, Z.; Von Ballmoos, M.W.; Voelzmann, J.; Diehm, N.; Baumgartner, I.; Kalka, C. Novel Cell-Free Strategy for Therapeutic Angiogenesis: In Vitro Generated Conditioned Medium Can Replace Progenitor Cell Transplantation. PLoS ONE 2009, 4, e5643. [Google Scholar] [CrossRef] [Green Version]
- Park, B.-S.; Kim, W.-S.; Choi, J.-S.; Kim, H.-K.; Won, J.-H.; Ohkubo, F.; Fukuoka, H. Hair growth stimulated by conditioned medium of adipose-derived stem cells is enhanced by hypoxia: Evidence of increased growth factor secretion. Biomed. Res. 2010, 31, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Arck, P.C.; Handjiski, B.; Hagen, E.; Joachim, R.; Klapp, B.F.; Paus, R. Indications for a brain-hair follicle axis: Inhibition of keratinocyte proliferation and up-regulation of keratinocyte apoptosis in telogen hair follicles by stress and substance P. FASEB J. 2001, 15, 2536–2538. [Google Scholar] [CrossRef]
- Ito, N.; Ito, T.; Kromminga, A.; Bettermann, A.; Takigawa, M.; Kees, F.; Straub, R.H.; Paus, R. Human hair follicles display a functional equivalent of the hypothalamic-pituitary-adrenal (HPA) axis and synthesize cortisol. FASEB J. 2005, 19, 1332–1334. [Google Scholar] [CrossRef]
- Slominski, A.; Wortsman, J.; Tuckey, R.; Paus, R. Differential expression of HPA axis homolog in the skin. Mol. Cell. Endocrinol. 2007, 265–266, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yu, M.; Yu, W.; Weinberg, J.; Shapiro, J.; McElwee, K.J. Development of Alopecia Areata Is Associated with Higher Central and Peripheral Hypothalamic–Pituitary–Adrenal Tone in the Skin Graft Induced C3H/He, J Mouse Model. J. Investig. Dermatol. 2009, 129, 1527–1538. [Google Scholar] [CrossRef] [Green Version]
- De Miguel, M.P.; Montiel, F.A.; Iglesias, P.L.; Blázquez-Martínez, A.; Nistal, M. Epiblast-derived stem cells in embryonic and adult tissues. Int. J. Dev. Biol. 2009, 53, 1529–1540. [Google Scholar] [CrossRef]
- Edmondson, S.R.; Thumiger, S.P.; Werther, G.A.; Wraight, C.J. Epidermal Homeostasis: The Role of the Growth Hormone and Insulin-Like Growth Factor Systems. Endocr. Rev. 2003, 24, 737–764. [Google Scholar] [CrossRef] [PubMed]
- Pappa, K.; Anagnou, N.P. Novel sources of fetal stem cells: Where do they fit on the developmental continuum? Regen. Med. 2009, 4, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalitin, N.; Schlesinger, H.; Levy, M.J.; Kessler, E.; Kessler-Icekson, G. Expression of procollagen C-proteinase enhancer in cultured rat heart fibroblasts: Evidence for co-regulation with type I collagen. J. Cell. Biochem. x2003, 90, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Sobolewski, K.; Małkowski, A.; Bańkowski, E.; Jaworski, S. Wharton’s jelly as a reservoir of peptide growth factors. Placenta 2005, 26, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Wang, F.; Jin, C.; Huang, X.; Miller, D.L.; Basilico, C.; McKeehan, W.L. Role of Fibroblast Growth Factor Type 1 and 2 in Carbon Tetrachloride-Induced Hepatic Injury and Fibrogenesis. Am. J. Pathol. x2003, 163, 1653–1662. [Google Scholar] [CrossRef] [Green Version]
- Karahuseyinoglu, S.; Cinar, O.; Kilic, E.; Kara, F.; Akay, G.G.; Demiralp, D.Ö.; Tukun, A.; Uckan, D.; Can, A. Biology of Stem Cells in Human Umbilical Cord Stroma: In Situ and In Vitro Surveys. Stem Cells 2006, 25, 319–331. [Google Scholar] [CrossRef]
- Lund, R.D.; Wang, S.; Lu, B.; Girman, S.; Holmes, T.; Sauvé, Y.; Messina, D.J.; Harris, I.R.; Kihm, A.J.; Harmon, A.M.; et al. Cells Isolated from Umbilical Cord Tissue Rescue Photoreceptors and Visual Functions in a Rodent Model of Retinal Disease. Stem Cells 2009, 25, 602–611. [Google Scholar] [CrossRef]
- Nanaev, A.; Kohnen, G.; Milovanov, A.; Domogatsky, S.; Kaufmann, P. Stromal differentiation and architecture of the human umbilical cord. Placenta 1997, 18, 53–64. [Google Scholar] [CrossRef]
- Nekanti, U.; Rao, V.B.; Bahirvani, A.G.; Jan, M.; Totey, S.; Ta, M. Long-Term Expansion and Pluripotent Marker Array Analysis of Wharton’s Jelly-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2010, 19, 117–130. [Google Scholar] [CrossRef]
- Prasanna, S.J.; Gopalakrishnan, D.; Shankar, S.R.; Vasandan, A.B. Pro-Inflammatory Cytokines, IFNγ and TNFα, Influence Immune Properties of Human Bone Marrow and Wharton Jelly Mesenchymal Stem Cells Differentially. PLoS ONE 2010, 5, e9016. [Google Scholar] [CrossRef]
- Troyer, D.L.; Weiss, M.L. Concise Review: Wharton’s Jelly-Derived Cells Are a Primitive Stromal Cell Population. Stem Cells 2008, 26, 591–599. [Google Scholar] [CrossRef] [Green Version]
- Fong, C.-Y.; Chak, L.-L.; Biswas, A.; Tan, J.-H.; Gauthaman, K.; Chan, W.-K.; Bongso, A. Human Wharton’s Jelly Stem Cells Have Unique Transcriptome Profiles Compared to Human Embryonic Stem Cells and Other Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2011, 7, 1–16. [Google Scholar] [CrossRef]
- Can, A.; Balci, D. Isolation, Culture, and Characterization of Human Umbilical Cord Stroma-derived Mesenchymal Stem Cells. Methods Pharmacol. Toxicol. 2011, 698, 51–62. [Google Scholar] [CrossRef]
- Aljitawi, O.S.; Xiao, Y.; Zhang, D.; Stehno-Bittel, L.; Garimella, R.; Hopkins, R.A.; Detamore, M.S. Generating CK19-Positive Cells with Hair-Like Structures from Wharton’s Jelly Mesenchymal Stromal Cells. Stem Cells Dev. 2013, 22, 18–26. [Google Scholar] [CrossRef]
- Wang, L.; Singh, M.; Bonewald, L.F.; Detamore, M.S. Signalling strategies for osteogenic differentiation of human umbilical cord mesenchymal stromal cells for 3D bone tissue engineering. J. Tissue Eng. Regen. Med. 2009, 3, 398–404. [Google Scholar] [CrossRef] [Green Version]
- Jadalannagari, S.; Aljitawi, O.S. Ectodermal Differentiation of Wharton’s Jelly Mesenchymal Stem Cells for Tissue Engineering and Regenerative Medicine Applications. Tissue Eng. Part B Rev. 2015, 21, 314–322. [Google Scholar] [CrossRef]
- Sabapathy, V.; Sundaram, B.; VM, S.; Mankuzhy, P.; Kumar, S. Human Wharton’s Jelly Mesenchymal Stem Cells Plasticity Augments Scar-Free Skin Wound Healing with Hair Growth. PLoS ONE 2014, 9, e93726. [Google Scholar] [CrossRef]
- Yoo, B.Y.; Shin, Y.H.; Yoon, H.H.; Kim, Y.J.; Seo, Y.K.; Song, K.Y.; Park, J.K. Evaluation of Human Umbilical Cord-Derived Mesenchymal Stem Cells on in Vivo Hair Inducing Activity. J. Tissue Eng. Regen. Med. 2009, 6, 15–22. [Google Scholar]
- Yoo, B.-Y.; Shin, Y.-H.; Yoon, H.-H.; Seo, Y.-K.; Song, K.-Y.; Park, J.-K. Optimization of the reconstruction of dermal papilla like tissues employing umbilical cord mesenchymal stem cells. Biotechnol. Bioprocess Eng. 2010, 15, 182–190. [Google Scholar] [CrossRef]
- Calvin, S.E.; Oyen, M.L. Microstructure and Mechanics of the Chorioamnion Membrane with an Emphasis on Fracture Properties. Ann. N. Y. Acad. Sci. 2007, 1101, 166–185. [Google Scholar] [CrossRef]
- Lotgering, F.K.; Wallenburg, H.C. Mechanisms of production and clearance of amniotic fluid. Semin. Perinatol. 1986, 10, 10. [Google Scholar]
- Gosden, C.; Brock, D.J. Combined use of alphafetoprotein and amniotic fluid cell morphology in early prenatal diagnosis of fetal abnormalities. J. Med. Genet. 1978, 15, 262–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoehn, H.; Bryant, E.M.; Karp, L.; Martin, G.M. Cultivated cells from diagnostic amniocentesis in second trimester pregnancies. II. Cytogenetic parameters as functions of clonal type and preparative technique. Clin. Genet. 1975, 7, 29–36. [Google Scholar] [CrossRef] [PubMed]
- De Coppi, P.; Bartsch, G.; Siddiqui, M.; Xu, T.; Santos, C.C.; Perin, L.; Mostoslavsky, G.; Serre, A.C.; Snyder, E.Y.; Yoo, J.J.; et al. Isolation of amniotic stem cell lines with potential for therapy. Nat. Biotechnol. 2007, 25, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.; Lee, J.; Chang, Y.; Hwang, S. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum. Reprod. 2004, 19, 1450–1456. [Google Scholar] [CrossRef]
- Kaviani, A.; Guleserian, K.; Perry, T.; Jennings, R.W.; Ziegler, M.M.; Fauza, D. Fetal tissue engineering from amniotic fluid. J. Am. Coll. Surg. x2003, 196, 592–597. [Google Scholar] [CrossRef]
- Moschidou, D.; Mukherjee, S.; Blundell, M.P.; Jones, G.N.; Atala, A.J.; Thrasher, A.J.; Fisk, N.M.; De Coppi, P.; Guillot, P.V. Human Mid-Trimester Amniotic Fluid Stem Cells Cultured Under Embryonic Stem Cell Conditions with Valproic Acid Acquire Pluripotent Characteristics. Stem Cells Dev. 2013, 22, 444–458. [Google Scholar] [CrossRef]
- Prusa, A.; Marton, E.; Rosner, M.; Bernaschek, G.; Hengstschläger, M. Oct-4-expressing cells in human amniotic fluid: A new source for stem cell research? Hum. Reprod. x2003, 18, 1489–1493. [Google Scholar] [CrossRef] [Green Version]
- In’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Noort, W.A.; Claas, F.H.J.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H.H. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood x2003, 102, 1548–1549. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, X.-H.; Liang, M.-Y.; Ren, M.-H. Prenatal cytogenetic diagnosis study of 2782 cases of high-risk pregnant women. Chin. Med. J. 2010, 123, 423–430. [Google Scholar]
- Moschidou, D.; Mukherjee, S.; Blundell, M.; Drews, K.; Jones, G.N.; Abdulrazzak, H.; Nowakowska, B.; Phoolchund, A.; Lay, K.; Ramasamy, T.S.; et al. Valproic Acid Confers Functional Pluripotency to Human Amniotic Fluid Stem Cells in a Transgene-free Approach. Mol. Ther. 2012, 20, 1953–1967. [Google Scholar] [CrossRef] [Green Version]
- You, Q.; Cai, L.; Zheng, J.; Tong, X.; Zhang, D.; Zhang, Y. Isolation of human mesenchymal stem cells from third-trimester amniotic fluid. Int. J. Gynecol. Obstet. 2008, 103, 149–152. [Google Scholar] [CrossRef]
- Gucciardo, L.; Ochsenbein-Kölble, N.; Ozog, Y.; Verbist, G.; Van Duppen, V.; Fryns, J.; Lories, R.; Deprest, J. A Comparative Study on Culture Conditions and Routine Expansion of Amniotic Fluid-Derived Mesenchymal Progenitor Cells. Fetal Diagn. Ther. 2013, 34, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Roubelakis, M.G.; Bitsika, V.; Zagoura, D.; Trohatou, O.; Pappa, K.I.; Makridakis, M.; Antsaklis, A.; Vlahou, A.; Anagnou, N.P. In vitro and in vivo properties of distinct populations of amniotic fluid mesenchymal progenitor cells. J. Cell. Mol. Med. 2011, 15, 1896–1913. [Google Scholar] [CrossRef] [Green Version]
- Mirabella, T.; Cilli, M.; Carlone, S.; Cancedda, R.; Gentili, C. Amniotic liquid derived stem cells as reservoir of secreted angiogenic factors capable of stimulating neo-arteriogenesis in an ischemic model. Biomaterials 2011, 32, 3689–3699. [Google Scholar] [CrossRef]
- Moorefield, E.C.; McKee, E.E.; Solchaga, L.; Orlando, G.; Yoo, J.J.; Walker, S.; Furth, M.E.; Bishop, C.E. Cloned, CD117 Selected Human Amniotic Fluid Stem Cells Are Capable of Modulating the Immune Response. PLoS ONE 2011, 6, e26535. [Google Scholar] [CrossRef]
- Park, J.; Jun, E.K.; Son, D.; Hong, W.; Jang, J.; Yun, W.; Yoon, B.S.; Song, G.; Kim, I.Y.; You, S. Overexpression of Nanog in amniotic fluid–derived mesenchymal stem cells accelerates dermal papilla cell activity and promotes hair follicle regeneration. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Berebichez-Fridman, R.; Montero-Olvera, P.R. Sources and Clinical Applications of Mesenchymal Stem Cells: State-of-the-art review. Sultan Qaboos Univ. Med. J. 2018, 18, e264–e277. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Stem Cell-Rich | Growth Factor-Rich |
|---|---|
| I. Adult stem cells A. Adipose derived stem cell (ADSC) 1. Nanofat 2. Stromal vascular fraction (SVF) B. Hair follicular stem cell (HFSCs) 1. Autologous micro grafts [Human intra and extra dermal adipose tissue derived hair follicle stem cells (HD-AFSCs)] 2. Cultured HFSCs i. Hair follicle-derived MSCs (HF-MSC) ii. Hair follicle epidermal stem cells (HF-ESC) C. Bone marrow derived 1. Bone marrow mononuclear cells (BMMC) 2. Bone marrow aspirate concentrate (BMAC) II. Perinatal stem cells A. Umbilical cord blood derived B. Wharton jelly MSCs (WJ-MSC)/Umbilical cord MSCs (UC-MSC) 1. Sub amniotic. 2. Perivascular. 3. Intervascular. C. Amniotic fluid derived D. Placental MSC | I. Platelet-rich plasma (PRP) A. Autologous activated PRP (AA-PRP) B. Autologous non-activated PRP (A-PRP) II. Autologous growth factor concentrate (GFC) III. Conditioned medium (Secretomes) A. ADSC-CM (AAPE) B. hUCB-MSC-CM C. AF-MSC-CM D. HF-MSC-CM E. BM-MSC-CM (Genetically engineered) IV. Extracellular vesicles A. Exosomes i. New-born foreskin stem cell ii. DPC iii. BM-MSC B. Exosome-like (Ginseng) C. Microvesicles V. Placental extract |
| Title | Study Type | Sample Size | Active Agent | Results | Remarks |
|---|---|---|---|---|---|
| Adipose tissue derived stem cells (ADSC) | |||||
| Hair follicle growth by stromal vascular fraction enhancedadipose transplantation in baldness [82] | Pilot case series in humans (MPHL—Grade 2 to 6 and FPHL—Grade 1 to 3) | 9 | Adipose tissue enriched SVF | when fat + SVF was used, mean increase was 31 hair/cm2. Whereas, for fat alone, it was 14 hairs/cm2. | 6 month follow up was available for only 6 patients |
| Cellular therapy with human autologous adipose-derived adult cells of stromal vascular fraction for alopecia areata [83] | Human clinical study (Alopecia areata: Grade 1 or 2) | 20 | Adipose derived stromal vascular cells | It was noted that there was statistically significant increase in hair density, hair diameter and pull test value. | - |
| Stromal Vascular Fraction Enhanced Adipose Transplantation in Hair Loss: Early Experience & Active Phase II FDA Investigation [84] | Prospective, single blinded human clinical trial (AGA; Grade 1 to 4) | 9 | Adipose tissue enriched SVF | Compared to baseline, there was a 14 percent increase in the number of hairs (p = 0.01), with a mean difference of 28 hairs and a 34 percent increase in the anagen percentage (p = 0.09). | Only 6 patients were analyzed at 6 months as 3 lost follow up. |
| Clinical use of conditioned media of adipose tissue-derived stem cells in female pattern hair loss: a retrospective case series study [85] | Retrospective, observational human study (FPHL) | 27 | ADSC-CM (AAPE) | The density of the hair rose from 105.4 to 122.7 hairs/cm2 (p = 0.001). The thickness of the hair increased from 57.5 μ to 64.0 μ (p = 0.001). | Dose: Once per week for 12 weeks |
| Hair Regeneration Treatment Using Adipose-Derived Stem Cell Conditioned Medium: Follow-up With trichograms [87] | Prospective human study on alopecia | 22 (Half side comparison: 10) | ADSC-CM (AAPE) |
| Dose: 6 sessions every 3–5 weeks |
| Innovative method of alopecia treatment by autologous adipose-derived SVF [88] | Clinical human study(AGA) | 9 | Autologous SVF |
| A single dose of Autologous SVF was administered |
| Introducing Platelet-Rich Stroma: Platelet Rich Plasma (PRP) and Stromal Vascular Fraction (SVF) Combined for the Treatment of Androgenetic Alopecia [89] | Clinical human study (AGA) | 10 | PRP + SVF (Platelet rich stroma) |
| A single dose of Autologous PRP + SVF was administered |
| Hair follicular stem cells (HFSC) | |||||
| Stem cells from human hair follicles: first mechanical isolation for immediate autologous clinical use in androgenetic alopecia and hair loss [67] | Prospective human study (AGA) | 11 | HFSC obtained by mechanical centrifugation of punch biopsy from hair follicle |
| Primary outcomes were microscopic identification and counting of HFSCs. |
| Autologous Cellular Method Using micrografts of Human Adipose Tissue Derived Follicle Stem Cells in Androgenic Alopecia [52] | Retrospective observational case-series, randomized, evaluator-blinded, placebo controlled, half-head group study in human (AGA) | 33 | Autologous cell biological technique (A-CBT) based on micro-grafts containing Hair Follicle Mesenchymal Stem Cells (HF-MSCs) |
| Dose: 3 injections at 45 days interval |
| Platelet-Rich Plasma and Micrografts Enriched with Autologous Human Follicle Mesenchymal Stem Cells Improve Hair Re-Growth in Androgenetic Alopecia. Biomolecular Pathway Analysis and Clinical Evaluation [103] | Retrospective observational case-series in humans (AGA) | 21 (HF-MSC) 57 (A-PRP) | HF-MSC And A-PRP |
|
|
| Autologous Micrografts from Scalp Tissue: Trichoscopic and Long-Term Clinical Evaluation in Male and Female Androgenetic Alopecia [99] | Placebo controlled, randomized, evaluator-blinded, half-head group study in humans (AGA) | 27 | Micrografts enriched with HF-MSCs |
| Six patients exhibited dynamic hair loss after 26 months. |
| Bone marrow derived stem cells (BMSC) | |||||
| Stem cell therapy as a novel therapeutic intervention for resistant cases of alopecia areata and androgenetic alopecia [56] | Double randomized clinical human study (AGA and AA) | 40 (20 AGA and 20 AA) | Autologous bone marrow derived mononuclear cells (BMMCs or autologous follicular stem cells (FSC). |
| No adverse effects were noted |
| Application of mesenchymal stem cells derived from bone marrow and umbilical cord in human hair multiplication [109] | Experimental study in vitro and on athymic mice | - | Culture expanded MSCs from bone marrow and umbilical cord of human beings | The DPLTs created in this technique were identical in size, shape and expression to actual DP. | MSCs were preconditioned in dermal papilla formation medium (DPFM) before being subcultured to create self-aggregated DPLTs. |
| Umbilical cord blood derived cells | |||||
| Human umbilical cord blood mesenchymal stem cells engineered to overexpress growth factors accelerate outcomes in hair growth [57] | Experimental study on C3H/HeJ mice | - | hUCB-MSCs |
| IGFBP-1 had a beneficial influence on cell survival, VEGF secretion, alkaline phosphatase (ALP), CD133, and b-catenin expression, and the formation of 3D spheroids of hDPCs via colocalization of an IGF-1 and IGFBP-1. |
| Migration Inhibitory Factor in Conditioned Medium from Human Umbilical Cord Blood-Derived Mesenchymal Stromal Cells Stimulates Hair Growth [40] | Double-blind placebo-controlled clinical trial (AGA) | 30 | hUCB-MSC-CM |
|
|
| Wharton jelly stem cells | |||||
| Human Wharton’s Jelly Mesenchymal Stem Cells Plasticity Augments Scar-Free Skin Wound Healing with Hair Growth [139] | Experimental study on black SCID mice | - | Human WJ-MSCs |
|
|
| Wharton’s jelly-derived mesenchymal stem cells in the treatment of four patients with alopecia areata [58] | Experimental case series on humans | 4 | Human WJ-MSC |
| This intervention was determined to be risk-free, with no negative side effects. |
| Amniotic fluid Mesenchymal stem cells (AF-MSC) | |||||
| Secretory Profiles and Wound Healing Effects of Human Amniotic Fluid–Derived Mesenchymal Stem Cells [41] | Experimental study in vitro and on ICR mice | - | AF-MSCs |
| AF-MSC-CM could be a potential therapeutic for improving the efficacy of tissue repair. |
| Overexpression of Nanog in amniotic fluid–derived mesenchymal stem cells accelerates dermal papilla cell activity and promotes hair follicle regeneration [159] | Experimental study on C57BL/6 mice | - | AF-N-MSCs |
| The secretome of autologous MSCs genetically modified to overexpress Nanog could be an attractive option for treatment of alopecia as a potent anagen inducer and hair growth stimulator. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anudeep, T.C.; Jeyaraman, M.; Muthu, S.; Rajendran, R.L.; Gangadaran, P.; Mishra, P.C.; Sharma, S.; Jha, S.K.; Ahn, B.-C. Advancing Regenerative Cellular Therapies in Non-Scarring Alopecia. Pharmaceutics 2022, 14, 612. https://doi.org/10.3390/pharmaceutics14030612
Anudeep TC, Jeyaraman M, Muthu S, Rajendran RL, Gangadaran P, Mishra PC, Sharma S, Jha SK, Ahn B-C. Advancing Regenerative Cellular Therapies in Non-Scarring Alopecia. Pharmaceutics. 2022; 14(3):612. https://doi.org/10.3390/pharmaceutics14030612
Chicago/Turabian StyleAnudeep, Talagavadi Channaiah, Madhan Jeyaraman, Sathish Muthu, Ramya Lakshmi Rajendran, Prakash Gangadaran, Prabhu Chandra Mishra, Shilpa Sharma, Saurabh Kumar Jha, and Byeong-Cheol Ahn. 2022. "Advancing Regenerative Cellular Therapies in Non-Scarring Alopecia" Pharmaceutics 14, no. 3: 612. https://doi.org/10.3390/pharmaceutics14030612
APA StyleAnudeep, T. C., Jeyaraman, M., Muthu, S., Rajendran, R. L., Gangadaran, P., Mishra, P. C., Sharma, S., Jha, S. K., & Ahn, B.-C. (2022). Advancing Regenerative Cellular Therapies in Non-Scarring Alopecia. Pharmaceutics, 14(3), 612. https://doi.org/10.3390/pharmaceutics14030612