
Micellar Curcumin Substantially Increases the Antineoplastic Activity of the Alkylphosphocholine Erufosine against TWIST1 Positive Cutaneous T Cell Lymphoma Cell Lines


 ,
,  , ,
, , 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Drugs and Chemicals
2.2. Cell Lines and Cultivation Procedure
2.3. Bacterial Strains and Growth Conditions
2.4. Preparation and Characterization of Curcumin Loaded Micelles
2.5. Cell Viability Test
2.6. Mathematical Modeling of Cytotoxic Effects after Single Drug Treatment
2.7. Mathematical Modelling of Drug-Drug Interactions in Cell Lines
2.8. Determination of Extracellular Curcumin Levels
2.9. Colony Forming Units (CFU) Assay
2.10. Determination of Reduced Glutathione (GSH) Content
2.11. Determination of Cytosolic Mono- and Oligonucleosomes
2.12. Caspase-3 Activity Assay
2.13. Detection of Apoptosis with Hoechst Staining
2.14. Western Blot Analysis for Protein Expression
2.15. NF-κB p65 ELISA
2.16. Biofilm Formation Assay
2.17. Statistical Evaluation
3. Results
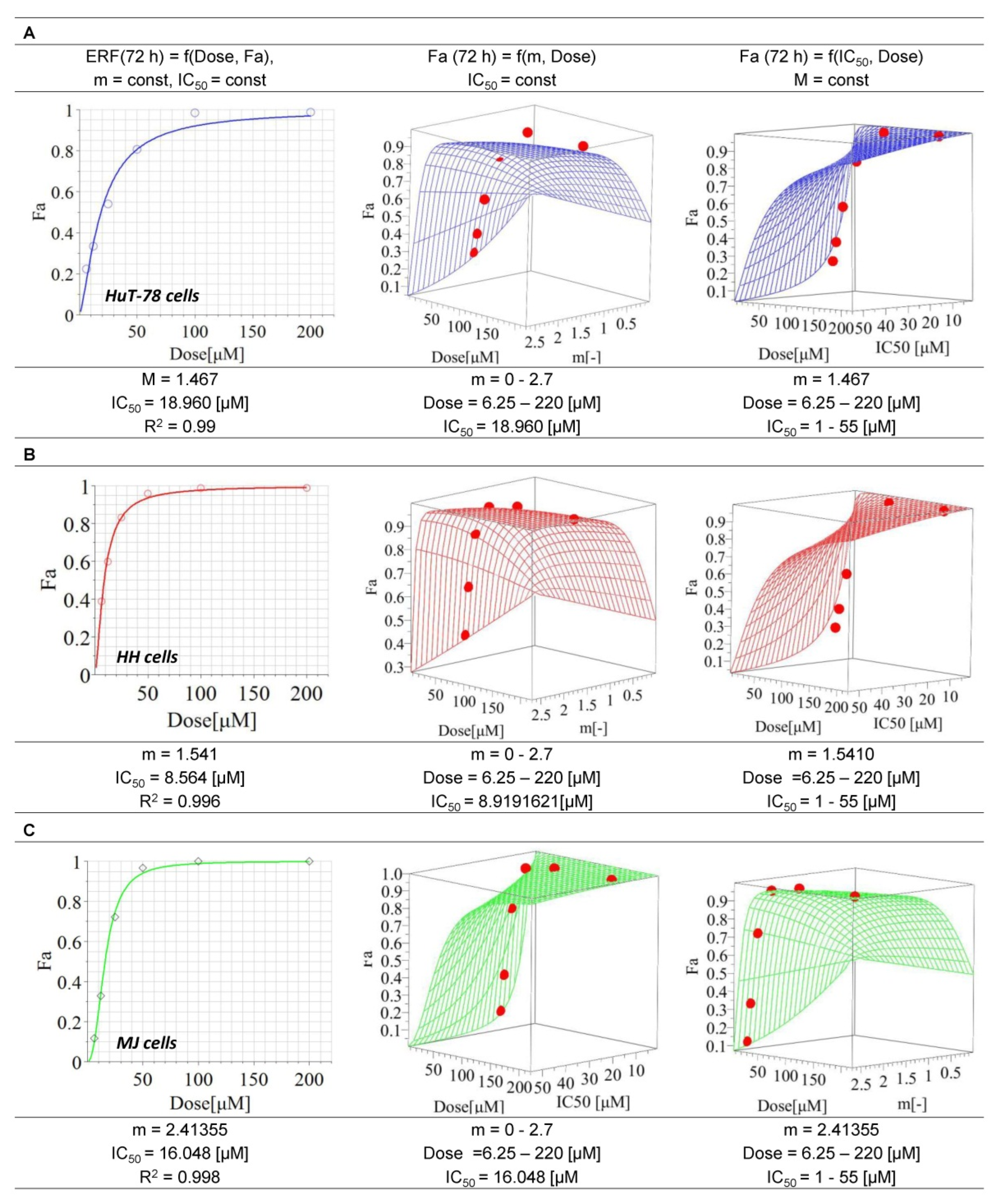
3.1. Cytotoxic Effects of Erufosine on CTCL Cell Lines
3.2. Production and Internalization Rate of MCRM
3.3. Micellar Curcumin Potentiates the Antiproliferative and Anticlonogenic Effect of Erufosine in TWIST1 Expressing CTCL Cell Lines
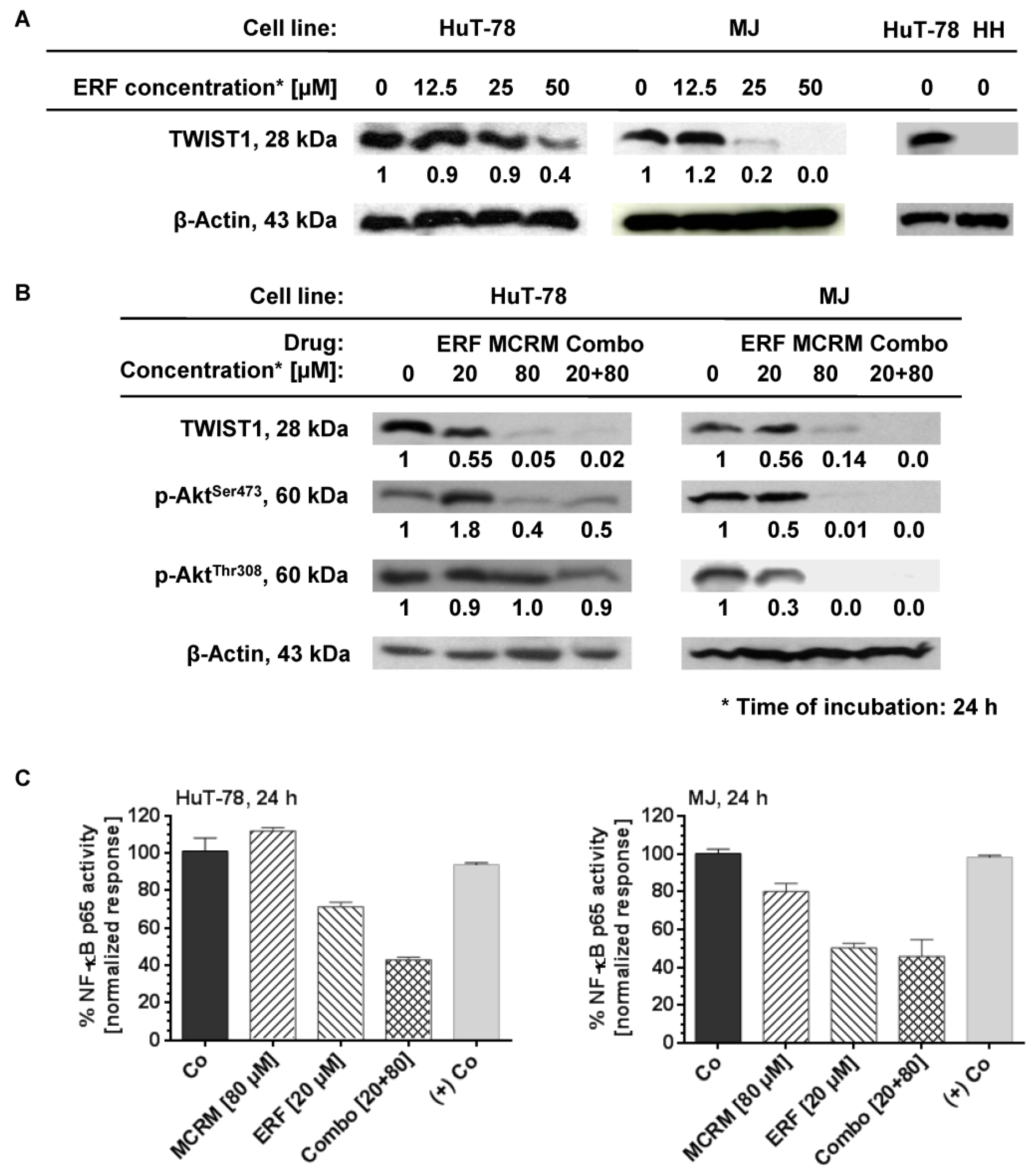
3.4. MCRM Increases the Inhibitory Effect of ERF on TWIST1, PKB/Akt and NF-κB in TWIST1 Expressing CTCL Cell Lines
3.5. MCRM Potentiates the Anti-Apoptotic Effect of ERF in TWIST1 Expressing CTCL Cell Lines
3.6. Effects of the Combinations between ERF and MCRM on Total GSH Levels in TWIST1 Expressing CTCL Cell Lines
3.7. The Combination Ratio of 4:1 between MCRM and ERF Inhibits Biofilm Formation of MRSA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CheckOrphan. 2016. Available online: http://www.checkorphan.org/diseases/cutaneous-t-cell-lymphoma (accessed on 15 May 2021).
- Rare Disease Registries in Europe. Orphanet Report Series: Rare Diseases Collection. 2016. Available online: www.orpha.net (accessed on 15 May 2021).
- Bagherani, N.; Smoller, B.R. An overview of cutaneous T cell lymphomas. F1000Research 2016, 5, 1882. [Google Scholar] [CrossRef] [PubMed]
- Cieza-Díaz, D.E.; Conde-Montero, E.; Menarguez-Palanca, J.; Longo-Imedio, I. Epidemiological and clinical features of patients diagnosed with cutaneous T-cell lymphomas in a spanish tertiary care hospital. J. Eur. Acad. Dermatol. Venereol. 2016, 31, e150–e153. [Google Scholar] [CrossRef] [PubMed]
- Zinzani, P.L.; Bonthapally, V.; Huebner, D.; Lutes, R.; Chi, A.; Pileri, S. Panoptic clinical review of the current and future treatment of relapsed/refractory T-cell lymphomas: Cutaneous T-cell lymphomas. Crit. Rev. Oncol. 2016, 99, 228–240. [Google Scholar] [CrossRef] [PubMed]
- Thestrup-Pedersen, K. Cutaneous T-Cell Lymphoma. A hypothesis on disease pathophysiology involving deficiency in DNA repair. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1682–1685. [Google Scholar] [CrossRef]
- Duvic, M. Histone Deacetylase Inhibitors for Cutaneous T-Cell Lymphoma. Dermatol. Clin. 2015, 33, 757–764. [Google Scholar] [CrossRef]
- Willerslev-Olsen, A.; Krejsgaard, T.; Lindahl, L.M.; Bonefeld, C.M.; Wasik, M.A.; Koralov, S.B.; Geisler, C.; Kilian, M.; Iversen, L.; Woetmann, A.; et al. Bacterial toxins fuel disease progression in cutaneous T-cell lymphoma. Toxins 2013, 5, 1402–1421. [Google Scholar] [CrossRef]
- Axelrod, P.I.; Lorber, B.; Vonderheid, E.C. Infections complicating mycosis fungoides and Sézary syndrome. JAMA 1992, 267, 1354–1358. [Google Scholar] [CrossRef]
- Brunner, P.M.; Jonak, C.; Knobler, R. Recent advances in understanding and managing cutaneous T-cell lymphomas. F1000Research 2020, 9, 331. [Google Scholar] [CrossRef]
- Damsky, W.E.; Choi, J. Genetics of Cutaneous T Cell Lymphoma: From Bench to Bedside. Curr. Treat. Options Oncol. 2016, 17, 33. [Google Scholar] [CrossRef]
- Nicolay, J.P.; Felcht, M.; Schledzewski, K.; Goerdt, S.; Geraud, C. Sezary syndrome: Old enigmas, new targets. J. Dtsch. Dermatol. Ges. 2016, 14, 256–264. [Google Scholar] [CrossRef]
- Phyo, Z.; Shanbhag, S.; Rozati, S. Update on Biology of Cutaneous T-Cell Lymphoma. Front. Oncol. 2020, 10, 765. [Google Scholar] [CrossRef]
- Khan, M.A.; Chen, H.-c.; Zhang, D.; Fu, J. Twist: A molecular target in cancer therapeutics. Tumor Biol. 2013, 34, 2497–2506. [Google Scholar] [CrossRef] [PubMed]
- van Doorn, R.; Dijkman, R.; Vermeer, M.H.; Out-Luiting, J.J.; van der Raaij-Helmer, E.M.H.; Willemze, R.; Tensen, T.P. Aberrant Expression of the Tyrosine Kinase Receptor EphA4 and the Transcription Factor Twist in SΓ©zary Syndrome Identified by Gene Expression Analysis. Cancer Res. 2004, 64, 5578–5586. [Google Scholar] [CrossRef] [PubMed]
- Goswami, M.; Duvic, M.; Dougherty, A.; Ni, X. Increased Twist expression in advanced stage of mycosis fungoides and Sézary syndrome. J. Cutan. Pathol. 2012, 39, 500–507. [Google Scholar] [CrossRef]
- Kempf, W.; Kazakov, D.V.; Kerl, K. Cutaneous lymphomas: An update. Part 1: T-cell and natural killer/t-cell lymphomas and related conditions. Am. J. Dermatopathol. 2014, 36, 105–123. [Google Scholar] [CrossRef]
- Spaccarelli, N.; Rook, A.H. The Use of Interferons in the Treatment of Cutaneous T-Cell Lymphoma. Dermatol. Clin. 2015, 33, 731–745. [Google Scholar] [CrossRef]
- Bobrowicz, M.; Fassnacht, C.; Ignatova, D.; Chang, Y.-T.; Dimitriou, F.; Guenova, E. Pathogenesis and Therapy of Primary Cutaneous T-Cell Lymphoma: Collegium Internationale Allergologicum (CIA) Update 2020. Int. Arch. Allergy Immunol. 2020, 181, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Zic, J.A. Extracorporeal Photopheresis in the Treatment of Mycosis Fungoides and Sézary Syndrome. Dermatol. Clin. 2015, 33, 765–776. [Google Scholar] [CrossRef]
- Vieyra-Garcia, P.A.; Wolf, P. Extracorporeal Photopheresis: A Case of Immunotherapy Ahead of Its Time. Transfus. Med. Hemotherapy 2020, 47, 226–235. [Google Scholar] [CrossRef]
- Wilcox, R.A. Cutaneous T-cell lymphoma: 2016 update on diagnosis, risk-stratification, and management. Am. J. Hematol. 2016, 91, 151–165. [Google Scholar] [CrossRef]
- Chung, C.G.; Poligone, B. Other Chemotherapeutic Agents in Cutaneous T-Cell Lymphoma. Dermatol. Clin. 2016, 33, 787–805. [Google Scholar] [CrossRef] [PubMed]
- Eibl, H.; Kaufmann-Kolle, P. Medical Application of Synthetic Phospholipids as Liposomes and Drugs. J. Liposome Res. 1995, 5, 131–148. [Google Scholar] [CrossRef]
- Kaufmann-Kolle, P.; Koetting, J.; Drevs, J.; Berger, M.; Unger, C.; Eibl, H. Intravenous application of alkylphosphocholines: Comparison of different homologues in lamellar structures. J. Cancer Res. Clin. Oncol. 1992, 120, R14. [Google Scholar]
- Konstantinov, S.M.; Eibl, H.; Berger, M.R. BCR-ABL influences the antileukaemic efficacy of alkylphosphocholines. Br. J. Haematol. 1999, 107, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, M.C.; Konstantinov, S.M.; Topashka-Ancheva, M.; Berger, M.R. Combination effects of alkylphosphocholines and gemcitabine in malignant and normal hematopoietic cells. Cancer Lett. 2002, 182, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.M.; Papa, V.; Tazzari, P.L.; Ricci, F.; Evangelisti, C.; Chiarini, F.; Grimaldi, C.; Cappellini, A.; Martinelli, G.; Ottaviani, E.; et al. Erucylphosphohomocholine, the first intravenously applicable alkylphosphocholine, is cytotoxic to acute myelogenous leukemia cells through JNK- and PP2A-dependent mechanisms. Leukemia 2010, 24, 687–698. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zaharieva, M.M.; Kroumov, A.D.; Dimitrova, L.; Tsvetkova, I.; Trochopoulos, A.; Konstantinov, S.M.; Berger, M.R.; Momchilova, M.; Yoncheva, K.; Najdenski, H.M. Micellar curcumin improves the antibacterial activity of the alkylphosphocholines erufosine and miltefosine against pathogenic Staphyloccocus aureus strains. Biotechnol. Biotechnol. Equip. 2019, 33, 38–53. [Google Scholar] [CrossRef]
- Fleer, E.A.; Unger, C.; Kim, D.J.; Eibl, H. Metabolism of ether phospholipids and analogs in neoplastic cells. Lipids 1987, 22, 856–861. [Google Scholar] [CrossRef]
- Fiegl, M.; Lindner, L.H.; Juergens, M.; Eibl, H.; Hiddemann, W.; Braess, J. Erufosine, a novel alkylphosphocholine, in acute myeloid leukemia: Single activity and combination with other antileukemic drugs. Cancer Chemother. Pharmacol. 2007, 62, 321–329. [Google Scholar] [CrossRef]
- Berger, M.R.; Sobottka, S.B.; Konstantinov, S.; Eibl, H. Erucylphosphocholine is the prototype of i.v. injectable alkylphosphocholines. Drugs Today 1998, 34, 73–81. [Google Scholar]
- Königs, S.K.; Pallasch, C.P.; Lindner, L.H.; Schwamb, J.; Schulz, A.; Brinker, R.; Claasen, J.; Veldurthy, A.; Eibl, H.; Hallek, M.; et al. Erufosine, a novel alkylphosphocholine, induces apoptosis in CLL through a caspase-dependent pathway. Leuk. Res. 2010, 34, 1064–1069. [Google Scholar] [CrossRef] [PubMed]
- Yosifov, D.Y.; Konstantinov, S.M.; Berger, M.R. Erucylphospho-N,N,N-trimethylpropylammonium Shows Substantial Cytotoxicity in Multiple Myeloma Cells. Ann. N.Y. Acad. Sci. 2009, 1171, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Zaharieva, M.; Konstantinov, S.; Pilicheva, B.; Karaivanova, M.; Berger, M. Erufosine: A Membrane Targeting Antineoplastic Agent with Signal Transduction Modulating Effects. Ann. N.Y. Acad. Sci. 2007, 1095, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, V.; Zaharieva, M.M.; Das, S.N.; Berger, M.R. Erufosine simultaneously induces apoptosis and autophagy by modulating the Akt-mTOR signaling pathway in oral squamous cell carcinoma. Cancer Lett. 2012, 319, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Dineva, I.K.; Zaharieva, M.M.; Konstantinov, S.M.; Eibl, H.; Berger, M.R. Erufosine suppresses breast cancer in vitro and in vivo for its activity on PI3K, c-Raf and Akt proteins. J. Cancer Res. Clin. Oncol. 2012, 138, 1909–1917. [Google Scholar] [CrossRef]
- Yosifov, D.Y.; Todorov, P.T.; Zaharieva, M.; Georgiev, K.D.; Pilicheva, B.A.; Konstantinov, S.M.; Berger, M.R. Erucylphospho-N,N,N-trimethylpropylammonium (erufosine) is a potential antimyeloma drug devoid of myelotoxicity. Cancer Chemother. Pharmacol. 2010, 67, 13–25. [Google Scholar] [CrossRef]
- Rudner, J.; Ruiner, C.-E.; Handrick, R.; Eibl, H.-J.; Belka, C.; Jendrossek, V. The Akt-inhibitor Erufosine induces apoptotic cell death in prostate cancer cells and increases the short term effects of ionizing radiation. Radiat. Oncol. 2010, 5, 108. [Google Scholar] [CrossRef]
- Zaharieva, M.; Kirilov, M.; Chai, M.; Berger, S.M.; Konstantinov, S.; Berger, M.R. Reduced Expression of the Retinoblastoma Protein Shows That the Related Signaling Pathway Is Essential for Mediating the Antineoplastic Activity of Erufosine. PLoS ONE 2014, 9, e100950. [Google Scholar] [CrossRef]
- Yosifov, D.Y.; Kaloyanov, K.A.; Guenova, M.L.; Prisadashka, K.; Balabanova, M.B.; Berger, M.R.; Konstantinov, S.M. Alkylphosphocholines and curcumin induce programmed cell death in cutaneous T-cell lymphoma cell lines. Leuk. Res. 2014, 38, 49–56. [Google Scholar] [CrossRef]
- Chometon, G.; Cappuccini, F.; Raducanu, A.; Aumailley, M.; Jendrossek, V. The Membrane-targeted Alkylphosphocholine Erufosine Interferes with Survival Signals from the Extracellular Matrix. Anti-Cancer Agents Med. Chem. 2014, 14, 578–591. [Google Scholar] [CrossRef]
- Ruiter, G.A.; Zerp, S.F.; Bartelink, H.; van Blitterswijk, W.J.; Verheij, M. Anti-cancer alkyl-lysophospholipids inhibit the phosphatidylinositol 3-kinase–Akt/PKB survival pathway. Anti-Cancer Drugs 2003, 14, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Konstantinov, S.M.; Topashka-Ancheva, M.; Benner, A.; Berger, M.R. Alkylphosphocholines: Effects on human leukemic cell lines and normal bone marrow cells. Int. J. Cancer 1998, 77, 778–786. [Google Scholar] [CrossRef]
- Bagley, R.G.; Kurtzberg, L.; Rouleau, C.; Yao, M.; Teicher, B.A. Erufosine, an alkylphosphocholine, with differential toxicity to human cancer cells and bone marrow cells. Cancer Chemother. Pharmacol. 2011, 68, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Dummer, R.; Krasovec, M.; Röger, J.; Sindermann, H.; Burg, G. Topical administration of hexadecylphosphocholine in patients with cutaneous lymphomas: Results of a phase I/II study. J. Am. Acad. Dermatol. 1993, 29, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Dummer, R.; Vogt, T.; Hefner, H.; Sindermann, H.; Burg, G. Topical Application of Hexadecylphosphocholine in Patients with Cutaneous Lymphomas. Prog. Exp. Tumor Res. 1992, 34, 160–169. [Google Scholar] [CrossRef]
- Dumontet, C.; Thomas, L.; Berard, F.; Gimonet, J.F.; Coiffier, B. A phase II trial of miltefosine in patients with cutaneous T-cell lymphoma. Bull. Cancer 2006, 93, E115–E118. [Google Scholar] [PubMed]
- Pronk, L.; Planting, A.; Oosterom, R.; Drogendijk, T.; Stoter, G.; Verweij, J. Increases in leucocyte and platelet counts induced by the alkyl phospholipid hexadecylphosphocholine. Eur. J. Cancer 1994, 30, 1019–1022. [Google Scholar] [CrossRef] [PubMed]
- Berkovic, D.; Bensch, M.; Bertram, J.; Wille, T.; Haase, D.; Binder, C.; Fleer, E. Effects of hexadecylphosphocholine on thrombocytopoiesis. Eur. J. Cancer 2001, 37, 503–511. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Sung, B. Pharmacological basis for the role of curcumin in chronic diseases: An age-old spice with modern targets. Trends Pharmacol. Sci. 2009, 30, 85–94. [Google Scholar] [CrossRef]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin. Exp. Pharmacol. Physiol. 2012, 39, 283–299. [Google Scholar] [CrossRef]
- Duvoix, A.; Blasius, R.; Delhalle, S.; Schnekenburger, M.; Morceau, F.; Henry, E.; Dicato, M.; Diederich, M. Chemopreventive and therapeutic effects of curcumin. Cancer Lett. 2005, 223, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Syng-Ai, C.; Kumari, A.L.; Khar, A. Effect of curcumin on normal and tumor cells: Role of glutathione and bcl-2. Mol. Cancer Ther. 2004, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, B.; Zhang, X.; Hazarika, P.; Aggarwal, B.B.; Duvic, M. Curcumin selectively induces apoptosis in cutaneous T-cell lymphoma cell lines and patients’ PBMCs: Potential role for STAT-3 and NF-kappaB signaling. J. Investig. Dermatol. 2010, 130, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Gahlot, S.; Majumdar, S. Oxidative Stress Induced by Curcumin Promotes the Death of Cutaneous T-cell Lymphoma (HuT-78) by Disrupting the Function of Several Molecular Targets. Mol. Cancer Ther. 2012, 11, 1873–1883. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, A.; Barik, A.; Mishra, B.; Rathinasamy, K.I.; Pandey, R.; Priyadarsini, K. Quantitative cellular uptake, localization and cytotoxicity of curcumin in normal and tumor cells. Biochim. Biophys. Acta 2008, 1780, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Lapteva, M.; Mondon, K.; Möller, M.; Gurny, R.; Kalia, Y.N. Polymeric Micelle Nanocarriers for the Cutaneous Delivery of Tacrolimus: A Targeted Approach for the Treatment of Psoriasis. Mol. Pharm. 2014, 11, 2989–3001. [Google Scholar] [CrossRef]
- Xue, B.; Wang, Y.; Tang, X.; Xie, P.; Wang, Y.; Luo, F.; Wu, C.; Qian, Z. Biodegradable Self-Assembled MPEG-PCL Micelles for Hydrophobic Oridonin Delivery In Vitro. J. Biomed. Nanotechnol. 2012, 8, 80–89. [Google Scholar] [CrossRef]
- Lapteva, M.; Möller, M.; Gurny, R.; Kalia, Y.N. Self-assembled polymeric nanocarriers for the targeted delivery of retinoic acid to the hair follicle. Nanoscale 2015, 7, 18651–18662. [Google Scholar] [CrossRef]
- Eibl, H.; Hilgard, C.; Unger, C. Alkylphosphocholines: New Drugs in Cancer Therapy; Eibl, H., Hilgard, C., Unger, C., Eds.; Karger: Basel, Switzerland, 1992; 173p. [Google Scholar]
- ISO 10993-5:2009; Biological Evaluation of Medical Devices—Part 5: Tests for In Vitro Cytotoxicity. ISO: Geneva, Switzerland, 2017.
- Chou, T.C.; Martin, N. (Eds.) CompuSyn for Drug Combinations: PC Software and User’s Guide: A Computer Program for Quantitation of Synergism and Antagonism in Drug Combinations, and the Determination of IC50and ED50 and LD50 Values; ComboSyn Inc: Paramus, NJ, USA, 2005. [Google Scholar]
- Zaharieva, M.; Trochopoulos, A.; Dimitrova, L.; Berger, M.; Najdenski, H.; Konstantinov, S.; Kroumov, A. New Insights in Routine Procedure for Mathematical Evaluation of in vitro Cytotoxicity Data from Cancer Cell Lines. Int. J. Bioautomation 2018, 22, 87–106. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Chazotte, B. Labeling Nuclear DNA with Hoechst 33342. Cold Spring Harb. Protoc. 2011, 2011, pdb.prot5557. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Hola, V.; DI Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Wong, H.K. STAT Assays with a TWIST: Differentiating Sézary Syndrome from Erythrodermic Inflammatory Dermatitis. J. Investig. Dermatol. 2016, 136, 1313–1315. [Google Scholar] [CrossRef][Green Version]
- Merindol, N.; Riquet, A.; Szablewski, V.; Eliaou, J.-F.; Puisieux, A.; Bonnefoy, N. The emerging role of Twist proteins in hematopoietic cells and hematological malignancies. Blood Cancer J. 2014, 4, e206. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Restuccia, D.F.; Lan, Q.; Hynx, D.; Dirnhofer, S.; Hess, D.; Rüegg, C.; Hemmings, B.A. Akt/PKB-Mediated Phosphorylation of Twist1 Promotes Tumor Metastasis via Mediating Cross-Talk between PI3K/Akt and TGF-OI Signaling Axes. Cancer Discov. 2012, 2, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Rahman, M.A.; Chen, Z.G.; Shin, D.M. Multiple biological functions of Twist1 in various cancers. Oncotarget 2017, 8, 20380–20393. [Google Scholar] [CrossRef] [PubMed]
- Kaleağasıoğlu, F.; Zaharieva, M.; Konstantinov, S.M.; Berger, M.R. Alkylphospholipids are Signal Transduction Modulators with Potential for Anticancer Therapy. Anti-Cancer Agents Med. Chem. 2019, 19, 66–91. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Kendall, S.E.; Raices, R.; Finlay, J.; Covarrubias, M.; Liu, Z.; Lowe, G.; Lin, Y.; Teh, Y.H.; Leigh, V.; et al. TWIST1 associates with NF-kB subunit RELA via carboxyl-terminal WR domain to promote cell autonomous invasion through IL8 production. BMC Biol. 2012, 10, 73. [Google Scholar] [CrossRef]
- Ilieva, Y.; Kaloyanov, K.; Yosifov, D.; Robev, B.; Zhelezova, I.; Genova, M.; Mihova, A.; Balatzenko, G.; Zaharieva, M.; Berger, M.R.; et al. Antineoplastic potential of curcumin (cooperative study in Bulgaria and Germany). Phytochem. Rev. 2014, 13, 459–469. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Kumar, A.; Bharti, A.C. Anticancer potential of curcumin: Preclinical and clinical studies. Anticancer Res. 2003, 23, 363–398. [Google Scholar] [PubMed]
- Shishodia, S.; Amin, H.M.; Lai, R.; Aggarwal, B.B. Curcumin (diferuloylmethane) inhibits constitutive NF-kappaB activation, induces G1/S arrest, suppresses proliferation, and induces apoptosis in mantle cell lymphoma. Biochem. Pharmacol. 2005, 70, 700–713. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, A.; Kunwar, A.; Narang, H.; Priyadarsini, K.I.; Krishna, M.; Pandey, R.; Sainis, K.B. Delayed activation of PKCdelta and NFkappaB and higher radioprotection in splenic lymphocytes by copper (II)-Curcumin (1) complex as compared to curcumin. J. Cell. Biochem. 2007, 102, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Gupta, S.C.; Tyagi, A.K.; Aggarwal, B.B. Curcumin, a component of golden spice: From bedside to bench and back. Biotechnol. Adv. 2014, 32, 1053–1064. [Google Scholar] [CrossRef]
- Obando, D.; Widmer, F.; Wright, L.C.; Sorrell, T.C.; Jolliffe, K.A. Synthesis, antifungal and antimicrobial activity of alkylphospholipids. Bioorg. Med. Chem. 2007, 15, 5158–5165. [Google Scholar] [CrossRef]
- Gong, C.; Wu, Q.; Wang, Y.; Zhang, D.; Luo, F.; Zhao, X.; Wei, Y.; Qian, Z. A biodegradable hydrogel system containing curcumin encapsulated in micelles for cutaneous wound healing. Biomaterials 2013, 34, 6377–6387. [Google Scholar] [CrossRef]
- Caon, T.; Mazzarino, L.; Simões, C.M.O.; Senna, E.L.; Silva, M.A.S. Lipid- and Polymer-Based Nanostructures for Cutaneous Delivery of Curcumin. AAPS PharmSciTech 2016, 18, 920–925. [Google Scholar] [CrossRef]
- Trochopoulos, A.G.; Zaharieva, M.M.; Marinova, M.H.; Yoncheva, K.; Tibi, I.P.; Berger, M.R.; Konstantinov, S.M. Antineoplastic effect of a novel nanosized curcumin on cutaneous T cell lymphoma. Oncol. Lett. 2020, 20, 304. [Google Scholar] [CrossRef]
- Kennedy, L.; Sandhu, J.K.; Harper, M.-E.; Cuperlovic-Culf, M. Role of Glutathione in Cancer: From Mechanisms to Therapies. Biomolecules 2020, 10, 1429. [Google Scholar] [CrossRef]
- Bansal, A.; Celeste Simon, M. Glutathione metabolism in cancer progression and treatment resistance. J. Cell Biol. 2018, 217, 2291–2298. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Incubation Time [h] | Absorption (AU) +/− SD | Asample − Ablank (AU) | Quantity of the Substance (mmol) |
|---|---|---|---|---|
| Control | 0 | 0.05748 +/− 0.01 | - | - |
| CRM | 0.47531 +/− 0.01 | 0.41783 | 8.0290 × 10−5 | |
| MCRM | 0.25410 +/− 0.01 | 0.19662 | 3.3212 × 10−5 | |
| Control | 1 | 0.12833 +/− 0.02 | - | - |
| CRM | 0.38940 +/− 0.02 | 0.26107 | 4.4100 × 10−5 | |
| MCRM | 0.27864 +/− 0.02 | 0.15033 | 2.5394 × 10−5 | |
| Control | 2 | 0.10574 +/− 0.02 | - | - |
| CRM | 0.37878 +/− 0.02 | 0.27304 | 4.612 × 10−5 | |
| MCRM | 0.23807 +/− 0.02 | 0.13233 | 2.2354 × 10−5 | |
| Control | 3 | 0.15630 +/− 0.01 | - | - |
| CRM | 0.43492 +/− 0.01 | 0.27862 | 4.7064 × 10−5 | |
| MCRM | 0.23947 +/− 0.01 | 0.08317 | 1.4050 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trochopoulos, A.G.X.; Ilieva, Y.; Kroumov, A.D.; Dimitrova, L.L.; Pencheva-El Tibi, I.; Philipov, S.; Berger, M.R.; Najdenski, H.M.; Yoncheva, K.; Konstantinov, S.M.; et al. Micellar Curcumin Substantially Increases the Antineoplastic Activity of the Alkylphosphocholine Erufosine against TWIST1 Positive Cutaneous T Cell Lymphoma Cell Lines. Pharmaceutics 2022, 14, 2688. https://doi.org/10.3390/pharmaceutics14122688
Trochopoulos AGX, Ilieva Y, Kroumov AD, Dimitrova LL, Pencheva-El Tibi I, Philipov S, Berger MR, Najdenski HM, Yoncheva K, Konstantinov SM, et al. Micellar Curcumin Substantially Increases the Antineoplastic Activity of the Alkylphosphocholine Erufosine against TWIST1 Positive Cutaneous T Cell Lymphoma Cell Lines. Pharmaceutics. 2022; 14(12):2688. https://doi.org/10.3390/pharmaceutics14122688
Chicago/Turabian StyleTrochopoulos, Antonios G. X., Yana Ilieva, Alexander D. Kroumov, Lyudmila L. Dimitrova, Ivanka Pencheva-El Tibi, Stanislav Philipov, Martin R. Berger, Hristo M. Najdenski, Krassimira Yoncheva, Spiro M. Konstantinov, and et al. 2022. "Micellar Curcumin Substantially Increases the Antineoplastic Activity of the Alkylphosphocholine Erufosine against TWIST1 Positive Cutaneous T Cell Lymphoma Cell Lines" Pharmaceutics 14, no. 12: 2688. https://doi.org/10.3390/pharmaceutics14122688
APA StyleTrochopoulos, A. G. X., Ilieva, Y., Kroumov, A. D., Dimitrova, L. L., Pencheva-El Tibi, I., Philipov, S., Berger, M. R., Najdenski, H. M., Yoncheva, K., Konstantinov, S. M., & Zaharieva, M. M. (2022). Micellar Curcumin Substantially Increases the Antineoplastic Activity of the Alkylphosphocholine Erufosine against TWIST1 Positive Cutaneous T Cell Lymphoma Cell Lines. Pharmaceutics, 14(12), 2688. https://doi.org/10.3390/pharmaceutics14122688