Solid-in-Oil Nanodispersions for Transcutaneous Immunotherapy of Japanese Cedar Pollinosis

 ,
, 
Abstract
1. Introduction
2. Overview of Antigens Developed for Antigen-specific Immunotherapy (AIT) of Japanese Cedar Pollinosis (JCP)
2.1. Cry j Allergens and PE
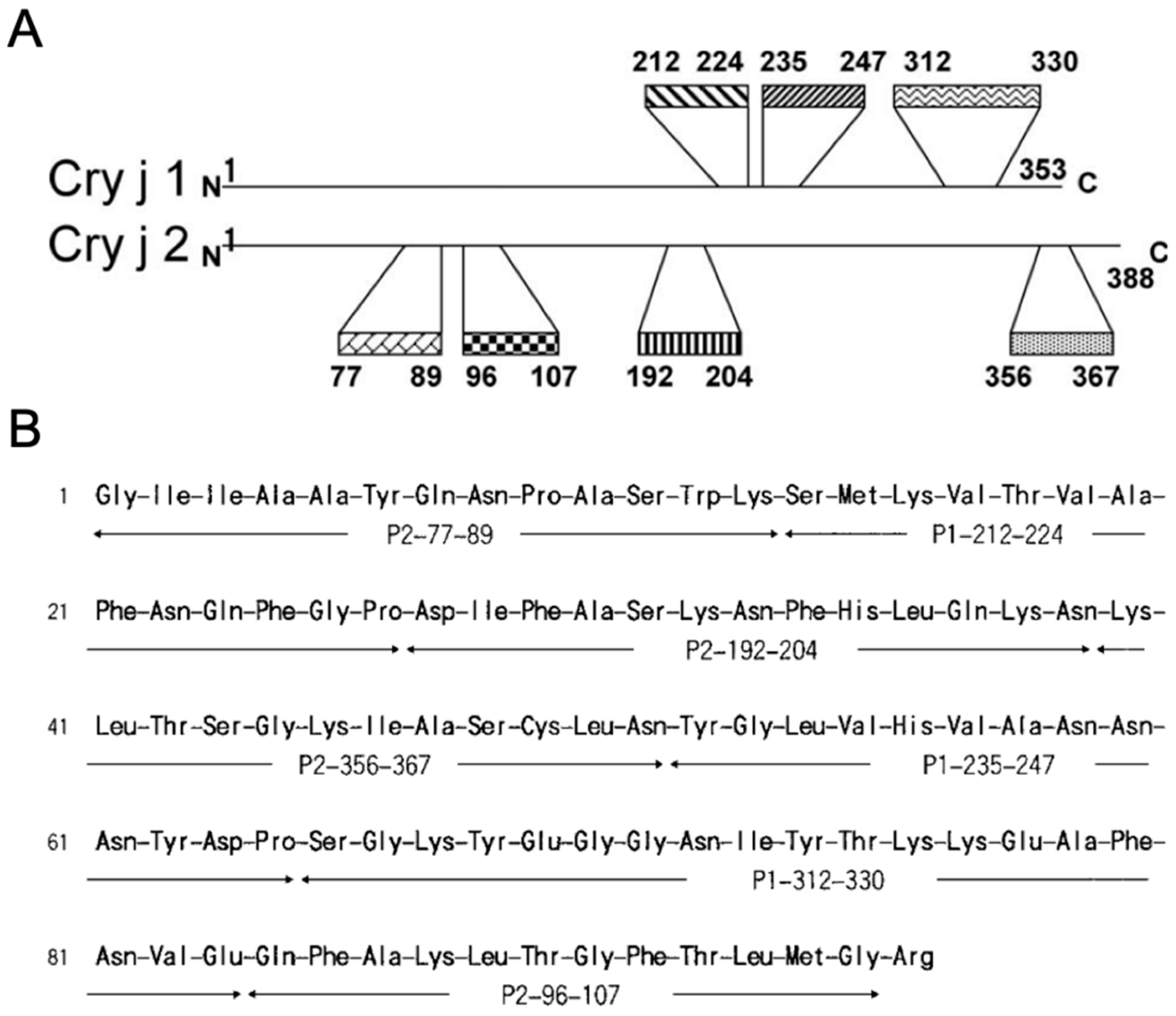
2.2. T Cell Epitope Peptide
2.3. Modified Allergen
2.4. DNA Vaccine
2.5. Adjuvant Conjugation
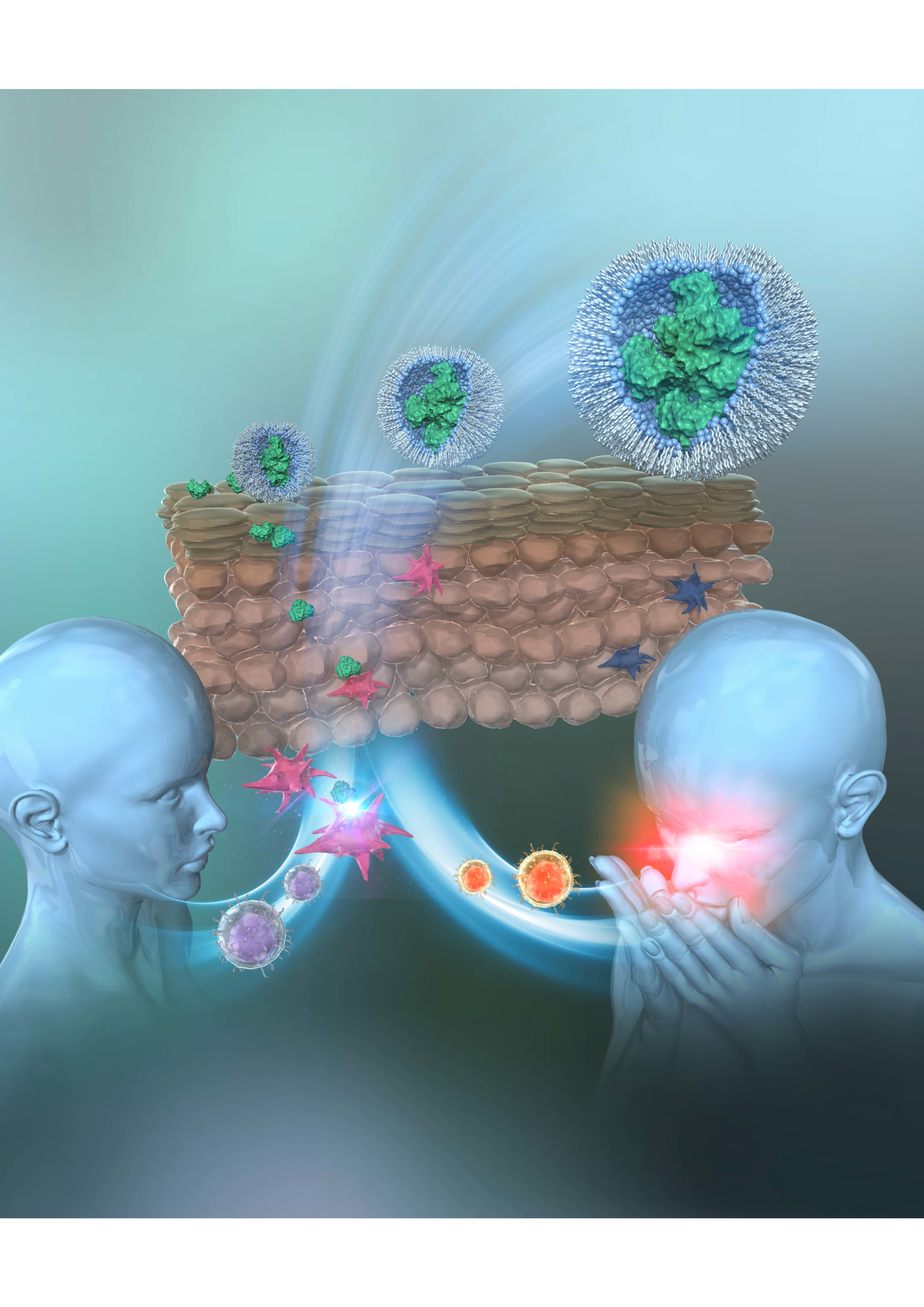
3. S/O Nanodispersions Developed for Transcutaneous Immunotherapy (TCIT) of JCP
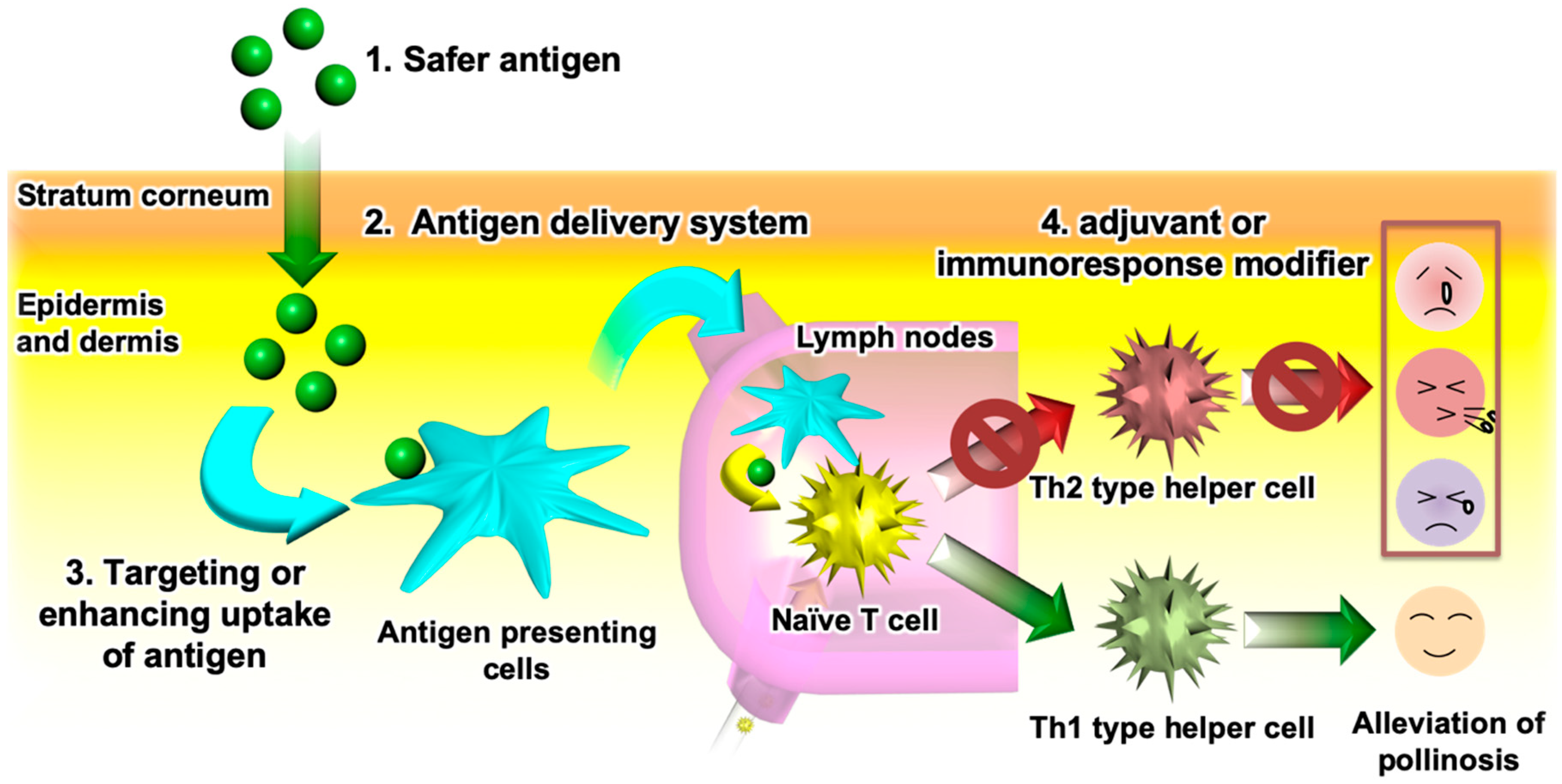
3.1. Background of Transcutaneous Antigen Delivery
3.2. Development and Application of S/O Nanodispersions for Transcutaneous Antigen Delivery
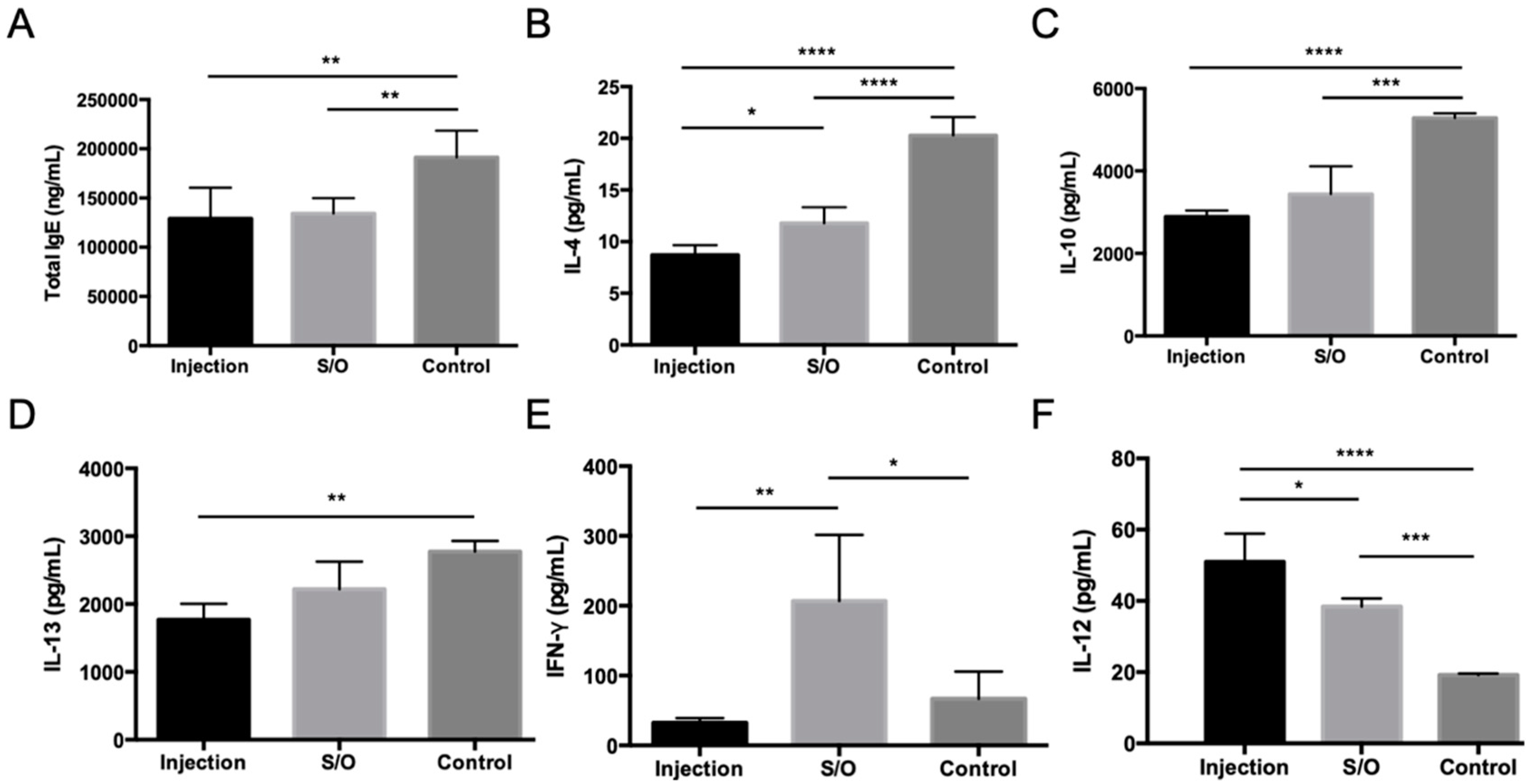
3.3. Applications of S/O Nanodispersions for TCIT of JCP
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nakamura, S.; Tsunoda, S.; Sakaida, H.; Masuda, S.; Said, A.S.; Takeuchi, K. Analysis of factors associated with cedar pollen sensitization and development of pollinosis in a young Japanese adult population. Allergol. Int. 2019, 68, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Saito, H.; Fujieda, S. Present state of Japanese cedar pollinosis: The national affliction. J. Allergy Clin. Immunol. 2014, 133, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Ricketti, P.A.; Alandijani, S.; Lin, C.H.; Casale, T.B. Investigational new drugs for allergic rhinitis. Expert Opin. Investig. Drugs 2017, 26, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Romeu-Bonilla, E.; Heiland, T. Next generation immunotherapy for tree pollen allergies. Hum. Vaccin. Immunother. 2017, 13, 2402–2415. [Google Scholar] [CrossRef] [PubMed]
- Kiel, M.A.; Röder, E.; Gerth van Wijk, R.; Al, M.J.; Hop, W.C.J.; Rutten-van Mölken, M.P.M.H. Real-life compliance and persistence among users of subcutaneous and sublingual allergen immunotherapy. J. Allergy Clin. Immunol. 2013, 132, 353–360. [Google Scholar] [CrossRef]
- Masuyama, K.; Matsuoka, T.; Kamijo, A. Current status of sublingual immunotherapy for allergic rhinitis in Japan. Allergol. Int. 2018, 67, 320–325. [Google Scholar] [CrossRef]
- Heath, W.R.; Carbone, F.R. The skin-resident and migratory immune system in steady state and memory: Innate lymphocytes, dendritic cells and T cells. Nat. Immunol. 2013, 14, 978–985. [Google Scholar] [CrossRef]
- Tay, S.S.; Roediger, B.; Tong, P.L.; Tikoo, S.; Weninger, W. The Skin-Resident Immune Network. Curr. Dermatol. Rep. 2014, 3, 13–22. [Google Scholar] [CrossRef]
- Langlois, A.; Graham, F.; Bégin, P. Epicutaneous peanut patch device for the treatment of peanut allergy. Expert Rev. Clin. Immunol. 2019, 15, 449–460. [Google Scholar] [CrossRef]
- Gunawardana, N.C.; Durham, S.R. New approaches to allergen immunotherapy. Ann. Allergyasthma Immunol. 2018, 121, 293–305. [Google Scholar] [CrossRef]
- Hashimoto, S.; Kawata, M.; Mino, N.; Ogino, S.; Takeda, N.; Kawashima, K.; Takagi, H.; Wakasa, Y.; Takano, M.; Tohda, Y.; et al. Clinical trials of Cry j 1 and Cry j 2 T-cell epitope peptide-expressing rice in patients with Japanese cedar pollinosis. J. Asian Pac. J. Allergy Immunol. 2019. Available online: http://apjai-journal.org/wp-content/uploads/2019/10/AP-050619-0575.pdf (accessed on 6 March 2020).
- Fujimura, T.; Kawamoto, S. Spectrum of allergens for Japanese cedar pollinosis and impact of component-resolved diagnosis on allergen-specific immunotherapy. Allergol. Int. 2015, 64, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Yasueda, H.; Yui, Y.; Shimizu, T.; Shida, T. Isolation and partial characterization of the major allergen from Japanese cedar (Cryptomeria japonica) pollen. J. Allergy Clin. Immunol. 1983, 71, 77–86. [Google Scholar] [CrossRef]
- Hashimoto, M.; Nigi, H.; Sakaguchi, M.; Inouye, S.; Imaoka, K.; Miyazawa, H.; Taniguchi, Y.; Kurimoto, M.; Yasueda, H.; Ogawa, T. Sensitivity to two major allergens (Cry j I and Cry j II) in patients with Japanese cedar (Cryptomeria japonica) pollinosis. Clin. Exp. Allergy 1995, 25, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Futamura, N.; Midoro-Horiuti, T.; Togawa, A.; Goldblum, R.M.; Yasueda, H.; Saito, A.; Shinohara, K.; Masuda, K.; Kurata, K.; et al. Isolation and characterization of native Cry j 3 from Japanese cedar (Cryptomeria japonica) pollen. Allergy 2007, 62, 547–553. [Google Scholar] [CrossRef]
- Osada, T.; Tanaka, Y.; Yamada, A.; Sasaki, E.; Utsugi, T. Identification of Cha o 3 homolog Cry j 4 from Cryptomeria japonica (Japanese cedar) pollen: Limitation of the present Japanese cedar–specific ASIT. Allergol. Int. 2018, 67, 467–474. [Google Scholar] [CrossRef]
- Sone, T.; Morikubo, K.; Miyahara, M.; Komiyama, N.; Shimizu, K.; Hsunoo, H.; Kino, K. T Cell Epitopes in Japanese Cedar (Cryptomeria japonica) Pollen Allergens: Choice of Major T Cell Epitopes in Cry j 1 and Cry j 2 Toward Design of the Peptide-Based Immunotherapeutics for the Management of Japanese Cedar Pollinosis. J. Immunol. 1998, 161, 448–457. [Google Scholar]
- O’Hehir, R.E.; Prickett, S.R.; Rolland, J.M. T Cell Epitope Peptide Therapy for Allergic Diseases. Curr. Allergy Asthma Rep. 2016, 16, 1–9. [Google Scholar] [CrossRef]
- Saito, S.; Hirahara, K.; Kawaguchi, J.; Serizawa, N.; Hino, K.; Taniguchi, Y. Identification of T cell determinants in Cry j 1 and Cry j 2 of size suitable for immunotherapy against Japanese cedar pollinosis. Annu Rep Sankyo Res Lab 2000, 52, 49–58. [Google Scholar]
- Hirahara, K.; Saito, S.; Serizawa, N.; Sasaki, R.; Sakaguchi, M.; Inouye, S.; Taniguchi, Y.; Kaminogawa, S.; Shiraishi, A. Oral administration of a dominant T-cell determinant peptide inhibits allergen-specific TH1 and TH2 cell responses in Cry j 2-primed mice. J. Allergy Clin. Immunol. 1998, 102, 961–967. [Google Scholar] [CrossRef]
- Murasugi, T.; Nakagami, Y.; Yoshitomi, T.; Hirahara, K.; Yamashita, M.; Taniguchi, Y.; Sakaguchi, M.; Ito, K. Oral administration of a T cell epitope inhibits symptoms and reactions of allergic rhinitis in Japanese cedar pollen allergen-sensitized mice. Eur. J. Pharmacol. 2005, 510, 143–148. [Google Scholar] [CrossRef]
- Yoshitomi, T.; Hirahara, K.; Kawaguchi, J.; Serizawa, N.; Taniguchi, Y.; Saito, S.; Sakaguchi, M.; Inouye, S.; Shiraishi, A. Three T-cell determinants of Cry j 1 and Cry j 2, the major Japanese cedar pollen antigens, retain their immunogenicity and tolerogenicity in a linked peptide. Immunology 2002, 107, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Takaiwa, F.; Yang, L. Development of a rice-based peptide vaccine for Japanese cedar and cypress pollen allergies. Transgenic Res. 2014, 23, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Saito, S.; Yang, L.; Nagasaka, S.; Nishizawa, N.; Takaiwa, F. Oral immunotherapy against a pollen allergy using a seed-based peptide vaccine. Plant Biotechnol. J. 2005, 3, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Kawabe, Y.; Hayashida, Y.; Numata, K.; Harada, S.; Hayashida, Y.; Ito, A.; Kamihira, M. Oral Immunotherapy for Pollen Allergy Using T-Cell Epitope-Containing Egg White Derived from Genetically Manipulated Chickens. PLoS ONE 2012, 7, e48512. [Google Scholar] [CrossRef]
- Tsunematsu, M.; Yamaji, T.; Kozutsumi, D.; Murakami, R.; Kimura, S.; Kino, K. Effect of Cry-consensus peptide, a novel recombinant peptide for immunotherapy of Japanese cedar pollinosis, on an experimental allergic rhinitis model in B10.S mice. Allergol. Int. 2007, 56, 465–472. [Google Scholar] [CrossRef]
- Yamanaka, K.; Yuta, A.; Kakeda, M.; Sasaki, R.; Kitagawa, H.; Gabazza, E.C.; Okubo, K.; Kurokawa, I.; Mizutani, H. Induction of IL-10–producing regulatory T cells with TCR diversity by epitope-specific immunotherapy in pollinosis. J. Allergy Clin. Immunol. 2009, 124, 842–845. [Google Scholar] [CrossRef]
- Kitaoka, M.; Shin, Y.; Kamiya, N.; Kawabe, Y.; Kamihira, M.; Goto, M. Transcutaneous Peptide Immunotherapy of Japanese Cedar Pollinosis Using Solid-in-Oil Nanodispersion Technology. Aaps Pharmscitech 2015, 16, 1418–1424. [Google Scholar] [CrossRef]
- Kong, Q.; Kitaoka, M.; Wakabayashi, R.; Kamiya, N.; Goto, M. Transcutaneous immunotherapy of pollinosis using solid-in-oil nanodispersions loaded with T cell epitope peptides. Int. J. Pharm. 2017, 529, 401–409. [Google Scholar] [CrossRef]
- Murakami, D.; Kubo, K.; Sawatsubashi, M.; Kikkawa, S.; Ejima, M.; Saito, A.; Kato, A.; Komune, S. Phase I/II study of oral immunotherapy with Cry j1-galactomannan conjugate for Japanese cedar pollinosis. Auris Nasus Larynx 2014, 41, 350–358. [Google Scholar] [CrossRef]
- Murakami, D.; Sawatsubashi, M.; Kikkawa, S.; Ejima, M.; Saito, A.; Kato, A.; Komune, S. Safety and efficacy of a new regimen of short-term oral immunotherapy with Cry j 1-galactomannan conjugate for Japanese cedar pollinosis: A prospective, randomized, open-label study. Allergol. Int. 2015, 64, 161–168. [Google Scholar] [CrossRef]
- Murakami, D.; Sawatsubashi, M.; Kikkawa, S.; Ejima, M.; Saito, A.; Kato, A.; Komune, S. Effect of short-term oral immunotherapy with Cry j1-galactomannan conjugate on quality of life in Japanese cedar pollinosis patients: A prospective, randomized, open-label study. Auris Nasus Larynx 2016, 43, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Murakami, D.; Sawatsubashi, M.; Omori, H.; Saito, A.; Kato, A.; Komune, S.; Nakagawa, T. Safety and efficacy of short-term oral immunotherapy with Cry j 1-galactomannan conjugate for Japanese cedar pollinosis: A randomized controlled trial. Sci. Rep. 2017, 7, 46142. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Higasijima, K.; Wakabayashi, R.; Tahara, Y.; Kitaoka, M.; Obayashi, H.; Hou, Y.; Kamiya, N.; Goto, M. Transcutaneous Delivery of Immunomodulating Pollen Extract-Galactomannan Conjugate by Solid-in-Oil Nanodispersions for Pollinosis Immunotherapy. Pharmaceutics 2019, 11, 563. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Fujinami, K.; Ishikawa, R.; Tateno, M.; Tahara, Y.; Okumura, Y.; Ohta, H.; Miyazaki, H.; Taniguchi, M. Recombinant Fusion Allergens, Cry j 1 and Cry j 2 from Japanese Cedar Pollen, Conjugated with Polyethylene Glycol Potentiate the Attenuation of Cry j 1-Specific IgE Production in Cry j 1-Sensitized Mice and Japanese Cedar Pollen Allergen-Sensitized Monkey. Int. Arch. Allergy Immunol. 2015, 168, 32–43. [Google Scholar] [CrossRef]
- Wakasa, Y.; Takagi, H.; Hirose, S.; Yang, L.; Saeki, M.; Nishimura, T.; Kaminuma, O.; Hiroi, T.; Takaiwa, F. Oral immunotherapy with transgenic rice seed containing destructed Japanese cedar pollen allergens, Cry j 1 and Cry j 2, against Japanese cedar pollinosis. Plant Biotechnol. J. 2013, 11, 66–76. [Google Scholar] [CrossRef]
- Wakasa, Y.; Takagi, H.; Watanabe, N.; Kitamura, N.; Fujiwara, Y.; Ogo, Y.; Hayashi, S.; Yang, L.; Ohta, M.; Tin, W.W.T.; et al. Concentrated protein body product derived from rice endosperm as an oral tolerogen for allergen-specific immunotherapy-a new mucosal vaccine formulation against Japanese cedar pollen allergy. PLoS ONE 2015, 10, 1–22. [Google Scholar] [CrossRef]
- Toda, M.; Sato, H.; Takebe, Y.; Taniguchi, Y.; Saito, S.; Inouye, S.; Takemori, T.; Sakaguchi, M. Inhibition of immunoglobulin E response to Japanese cedar pollen allergen (Cry j 1) in mice by DNA immunization: Different outcomes dependent on the plasmid DNA inoculation method. Immunology 2000, 99, 179–186. [Google Scholar] [CrossRef]
- Toda, M.; Kasai, M.; Hosokawa, H.; Nakano, N.; Taniguchi, Y.; Inouye, S.; Kaminogawa, S.; Takemori, T.; Sakaguchi, M. DNA vaccine using invariant chain gene for delivery of CD4+T cell epitope peptide derived from Japanese cedar pollen allergen inhibits allergen-specific IgE response. Eur. J. Immunol. 2002, 32, 1631–1639. [Google Scholar] [CrossRef]
- Su, Y.; Connolly, M.; Marketon, A.; Heiland, T. CryJ-LAMP DNA Vaccines for Japanese Red Cedar Allergy Induce Robust Th1-Type Immune Responses in Murine Model. J. Immunol. Res. 2016, 2016, 4857869. [Google Scholar] [CrossRef]
- Su, Y.; Romeu-Bonilla, E.; Anagnostou, A.; Fitz-Patrick, D.; Hearl, W.; Heiland, T. Safety and long-term immunological effects of CryJ2-LAMP plasmid vaccine in Japanese red cedar atopic subjects: A phase I study. Hum. Vaccin. Immunother. 2017, 13, 2804–2813. [Google Scholar] [CrossRef]
- Suzuki, M.; Ohta, N.; Min, W.P.; Matsumoto, T.; Min, R.; Zhang, X.; Toida, K.; Murakami, S. Immunotherapy with CpG DNA conjugated with T-cell epitope peptide of an allergenic Cry j 2 protein is useful for control of allergic conditions in mice. Int. Immunopharmacol. 2007, 7, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Kaburaki, Y.; Fujimura, T.; Kurata, K.; Masuda, K.; Toda, M.; Yasueda, H.; Chida, K.; Kawarai, S.; Sakaguchi, M. Induction of Th1 immune responses to Japanese cedar pollen allergen (Cry j 1) in mice immunized with Cry j 1 conjugated with CpG oligodeoxynucleotide. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Van Hoang, V.; Zou, Y.; Kurata, K.; Enomoto, K. Expression of recombinant T-cell epitopes of major Japanese cedar pollen allergens fused with cholera toxin B subunit in Escherichia coli. Protein Expr. Purif. 2015, 109, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Hiroi, T.; Yang, L.; Takamura, K.; Ishimitsu, R.; Kawauchi, H.; Takaiwa, F. Efficient induction of oral tolerance by fusing cholera toxin B subunit with allergen-specific T-cell epitopes accumulated in rice seed. Vaccine 2008, 26, 6027–6030. [Google Scholar] [CrossRef] [PubMed]
- Ishii, M.; Koyama, A.; Iseki, H.; Narumi, H.; Yokoyama, N.; Kojima, N. Anti-allergic potential of oligomannose-coated liposome-entrapped Cry j 1 as immunotherapy for Japanese cedar pollinosis in mice. Int. Immunopharmacol. 2010, 10, 1041–1046. [Google Scholar] [CrossRef]
- Kitaoka, M.; Naritomi, A.; Kawabe, Y.; Kamihira, M.; Kamiya, N.; Goto, M. Transcutaneous pollinosis immunotherapy using a solid-in-oil nanodispersion system carrying T cell epitope peptide and R848. Bioeng. Transl. Med. 2017, 2, 102–108. [Google Scholar] [CrossRef]
- Immonen, A.; Farci, S.; Taivainen, A.; Partanen, J.; Pouvelle-Moratille, S.; Närvänen, A.; Kinnunen, T.; Saarelainen, S.; Rytkönen-Nissinen, M.; Maillere, B.; et al. T Cell Epitope-Containing Peptides of the Major Dog Allergen Can f 1 as Candidates for Allergen Immunotherapy. J. Immunol. 2005, 175, 3614–3620. [Google Scholar] [CrossRef]
- Hirahara, K.; Tatsuta, T.; Takatori, T.; Ohtsuki, M.; Kirinaka, H.; Kawaguchi, J.; Serizawa, N.; Taniguchi, Y.; Saito, S.; Sakaguchi, M.; et al. Preclinical evaluation of an immunotherapeutic peptide comprising 7 T-cell determinants of Cry j 1 and Cry j 2, the major Japanese cedar pollen allergens. J. Allergy Clin. Immunol. 2001, 108, 94–100. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Hirahara, K.; Fujimura, T.; Toda, M. Approaches to immunotherapies for Japanese cedar pollinosis. Auris Nasus Larynx 2011, 38, 431–438. [Google Scholar] [CrossRef]
- Usui, M.; Saito, A.; Taniguchi, N.; Nishijima, N.; Azakami, H.; Kato, A. Reduction of antigenicity of Cry j I, major allergen of Japanese cedar pollen, by the attachment of polysaccharides. Biosci. Biotechnol. Biochem. 2003, 67, 2425–2430. [Google Scholar] [CrossRef]
- Aoki, R.; Saito, A.; Azakami, H.; Kato, A. Effects of Various Saccharides on the Masking of Epitope Sites and Uptake in the Gut of Cedar Allergen Cry j 1−Saccharide Conjugates by a Naturally Occurring Maillard Reaction. J. Agric. Food Chem. 2010, 58, 7986–7990. [Google Scholar] [CrossRef] [PubMed]
- Kojima, N.; Biao, L.; Nakayama, T.; Ishii, M.; Ikehara, Y.; Tsujimura, K. Oligomannose-coated liposomes as a therapeutic antigen-delivery and an adjuvant vehicle for induction of in vivo tumor immunity. J. Control. Release 2008, 129, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ukidve, A.; Dasgupta, A.; Mitragotri, S. Transdermal immunomodulation: Principles, advances and perspectives. Adv. Drug Deliv. Rev. 2018, 127, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Bos, J.D.; Meinardi, M. The 500 Dalton rule for the skin penetration of chemical compounds and drugs. Exp. Dermatol. 2000, 9, 165–169. [Google Scholar] [CrossRef]
- Madison, K.C. Barrier function of the skin: “La Raison d’Être” of the epidermis. J. Invest. Dermatol. 2003, 121, 231–241. [Google Scholar] [CrossRef]
- Lee, H.; Song, C.; Baik, S.; Kim, D.; Hyeon, T.; Kim, D.H. Device-assisted transdermal drug delivery. Adv. Drug Deliv. Rev. 2018, 127, 35–45. [Google Scholar] [CrossRef]
- Lane, M.E. Skin penetration enhancers. Int. J. Pharm. 2013, 447, 12–21. [Google Scholar] [CrossRef]
- Jain, S.; Patel, N.; Shah, M.K.; Khatri, P.; Vora, N. Recent Advances in Lipid-Based Vesicles and Particulate Carriers for Topical and Transdermal Application. J. Pharm. Sci. 2017, 106, 423–445. [Google Scholar] [CrossRef]
- Kitaoka, M.; Wakabayashi, R.; Kamiya, N.; Goto, M. Solid-in-oil nanodispersions for transdermal drug delivery systems. Biotechnol. J. 2016, 11, 1375–1385. [Google Scholar] [CrossRef]
- Wakabayashi, R.; Sakuragi, M.; Kozaka, S.; Tahara, Y.; Kamiya, N.; Goto, M. Solid-in-Oil Peptide Nanocarriers for Transcutaneous Cancer Vaccine Delivery against Melanoma. Mol. Pharm. 2018, 15, 955–961. [Google Scholar] [CrossRef]
- Tahara, Y.; Honda, S.; Kamiya, N.; Piao, H.; Hirata, A.; Hayakawa, E.; Fujii, T.; Goto, M. A solid-in-oil nanodispersion for transcutaneous protein delivery. J. Control. Release 2008, 131, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Azoia, N.G.; Ribeiro, A.; Shimanovich, U.; Silva, C.; Cavaco-Paulo, A. In vitro and computational studies of transdermal perfusion of nanoformulations containing a large molecular weight protein. Colloids Surf. B Biointerfaces 2013, 108, 271–278. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Martins, M.; Azoia, N.; Carvalho, A.; Silva, C.; Matamá, T.; Gomes, A.; Cavaco-Paulo, A. Assessment of a Protease Inhibitor Peptide for Anti-Ageing. Protein Pept. Lett. 2015, 22, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, M.; Imamura, K.; Hirakawa, Y.; Tahara, Y.; Kamiya, N.; Goto, M. Sucrose laurate-enhanced transcutaneous immunization with a solid-in-oil nanodispersion. Med. Chem. Commun. 2014, 5, 20–24. [Google Scholar] [CrossRef]
- Piao, H.; Kamiya, N.; Hirata, A.; Fujii, T.; Goto, M. A novel solid-in-oil nanosuspension for transdermal delivery of diclofenac sodium. Pharm. Res. 2008, 25, 896–901. [Google Scholar] [CrossRef]
- Tahara, Y.; Namatsu, K.; Kamiya, N.; Hagimori, M.; Kamiya, S.; Arakawa, M.; Goto, M. Transcutaneous immunization by a solid-in-oil nanodispersion. Chem. Commun. 2010, 46, 9200–9202. [Google Scholar] [CrossRef]
- Kitaoka, M.; Naritomi, A.; Hirakawa, Y.; Kamiya, N.; Goto, M. Transdermal Immunization using Solid-in-oil Nanodispersion with CpG Oligodeoxynucleotide Adjuvants. Pharm. Res. 2015, 32, 1486–1492. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Wakabayashi, R.; Naritomi, A.; Sakuragi, M.; Kamiya, N.; Goto, M. Transcutaneous immunization against cancer using solid-in-oil nanodispersions. Medchemcomm 2015, 6, 1387–1392. [Google Scholar] [CrossRef]
- Kitaoka, M.; Xiao, W.; Kong, Q.; Tahara, Y.; Kamiya, N.; Goto, M. A Solid-in-Oil Nanodispersion System for Transcutaneous Immunotherapy of Cow’s Milk Allergies. Pharmaceutics 2020, 12, 205. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Allergen | Route | Mechanism | References |
|---|---|---|---|---|
| T cell epitope peptide | Epitope p246–259 of Cry j 2 | Oral | Suppression of Th1 and Th2 responses | [20,21] |
| Integrated peptide from three epitopes | Oral | Inhibition of lymph node cell proliferation to epitopes from Cry j 1/2 | [22] | |
| Rice seed containing peptides from fourteen epitopes | Oral | - | [23] | |
| Rice seed containing 7Crp from seven epitopes | Oral | Inhibition of T-cell proliferative response to Cry j 1 | [24] | |
| Chicken egg containing 7Crp from seven epitopes | Oral | Induction of oral tolerance | [25] | |
| Cry-consensus from five or six epitopes | SC | Induction of IgG2a, suppression of ratios of IL-4/IFN-γ and IL-5/IFN-γ,Th1 deviation | [17,26] | |
| 7Crp from seven epitopes | SL | Induction of IL-10-producing regulatory T cells | [27] | |
| 7CrpR from seven epitopes | T | - | [28] | |
| Mixture of seven epitopes | T | Suppression of Th1 and Th2 responses | [29] | |
| Modified allergen | Cry j 1-glactomannan conjugate | Oral | Enhancement of antigen uptake, induction of IgG4 and IL-10 | [30,31,32,33] |
| PE-galactomannan conjugate | T | Enhancement of antigen uptake, suppression of Th2 responses, Th1 deviation | [34] | |
| Polyethylene glycol-modified Cry j 1/2 | SC | Induction of IgG and Th1 responses, suppression of Th2 responses | [35] | |
| Rice seed containing fusion protein from Cry j 1/2 | Oral | Suppression of Th1 and Th2 responses | [36,37] | |
| DNA vaccine | Vaccine encoding Cry j 1 gene | IM | Induction of IFN-γ, IgG 2a and Th1 responses, suppression of IgG 1 | [38] |
| Vaccine encoding T cell epitope peptide (p247-258) from Cry j 2 | IM | Induction of IgG 2a and Th1 responses | [39] | |
| Cry j 1-LAMP and Cry j 2-LAMP vaccine | IM | Induction of IFN-γ, IgG 2a and Th1 responses | [40,41] | |
| Adjuvant conjugation | Cry j 2 T cell epitope peptide-CpG conjugate | SC | Suppression of IL-4 and IL-5 | [42] |
| Cry j 1-CpG conjugate | SC | Induction of IFN-γ and IgG2a, Th1 deviation | [43] | |
| Cholera toxin B-fused T cell epitope peptides | Oral | Induction of oral tolerance | [44,45] | |
| Oligomannose-coated liposomes carrying Cry j 1 | ID | Induction of IFN-γ, suppression of IL-5/IFN-γ ratio, Th1 deviation | [46] | |
| 7CrpR from seven epitopes combined with R848 | T | Induction of IgG 2a/IgE ratio, Th1 deviation | [47] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, Q.; Kitaoka, M.; Wakabayashi, R.; Tahara, Y.; Kamiya, N.; Goto, M. Solid-in-Oil Nanodispersions for Transcutaneous Immunotherapy of Japanese Cedar Pollinosis. Pharmaceutics 2020, 12, 240. https://doi.org/10.3390/pharmaceutics12030240
Kong Q, Kitaoka M, Wakabayashi R, Tahara Y, Kamiya N, Goto M. Solid-in-Oil Nanodispersions for Transcutaneous Immunotherapy of Japanese Cedar Pollinosis. Pharmaceutics. 2020; 12(3):240. https://doi.org/10.3390/pharmaceutics12030240
Chicago/Turabian StyleKong, Qingliang, Momoko Kitaoka, Rie Wakabayashi, Yoshiro Tahara, Noriho Kamiya, and Masahiro Goto. 2020. "Solid-in-Oil Nanodispersions for Transcutaneous Immunotherapy of Japanese Cedar Pollinosis" Pharmaceutics 12, no. 3: 240. https://doi.org/10.3390/pharmaceutics12030240
APA StyleKong, Q., Kitaoka, M., Wakabayashi, R., Tahara, Y., Kamiya, N., & Goto, M. (2020). Solid-in-Oil Nanodispersions for Transcutaneous Immunotherapy of Japanese Cedar Pollinosis. Pharmaceutics, 12(3), 240. https://doi.org/10.3390/pharmaceutics12030240