
Serological Evidence of Zika Virus Infections in Sudan

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cohort Selection and Reference Sera for ZIKV Antibodies
2.2. Dengue Virus IgG Antibody Test
2.3. Flavivirus Neutralization Tests
2.4. Calibration Curve for ZIKV Neutralization Units
2.5. Statistical Analyses
3. Results
3.1. Identification of ZIKV-Neutralizing Sera
3.2. Comparison of the Neutralizing ZIKV Antibody Response with That of Standard Sera
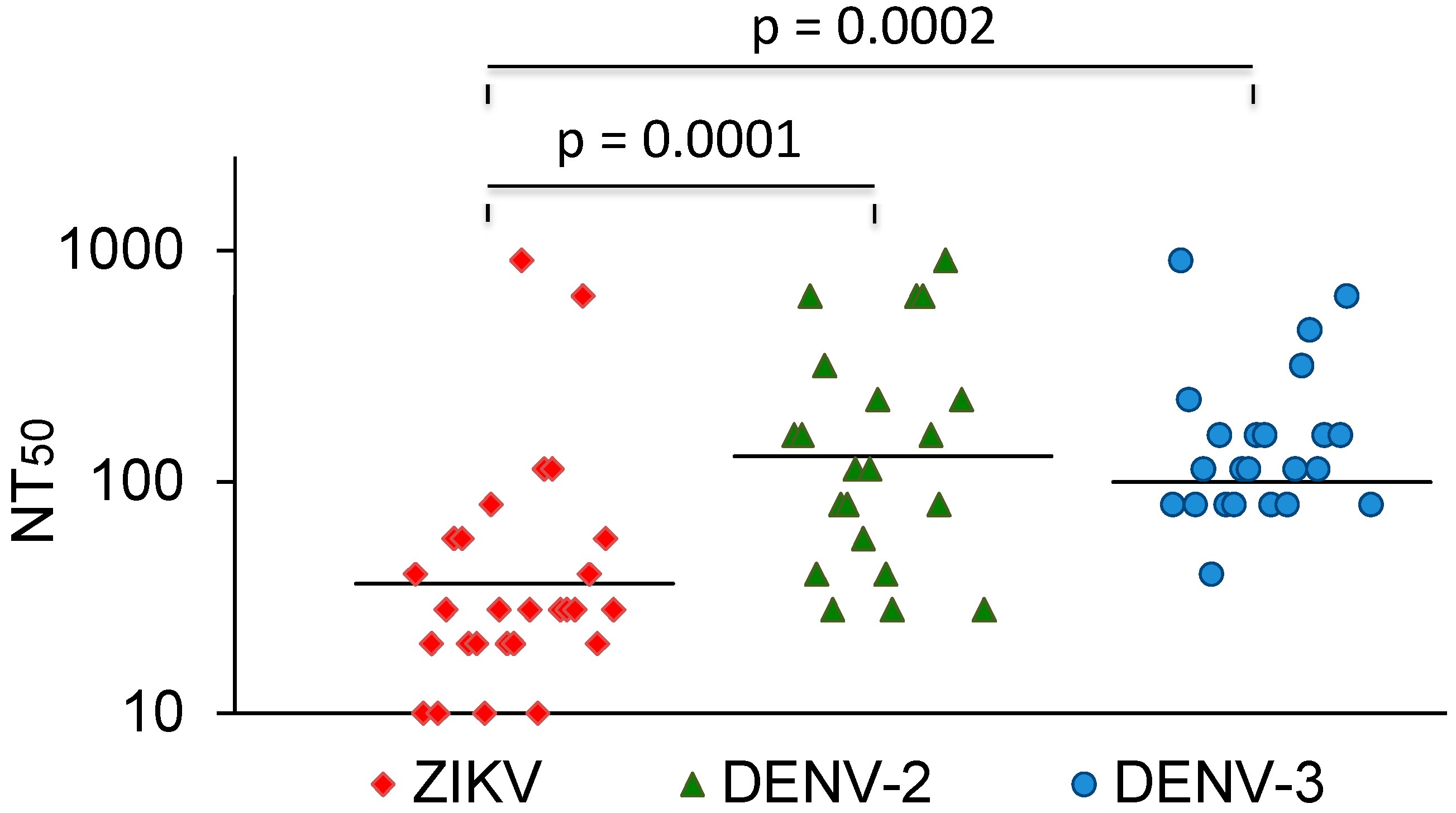
3.3. Comparison with DENV and YFV Neutralization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chong, H.Y.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Flavivirus infection-A review of immunopathogenesis, immunological response, and immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef] [PubMed]
- Mead, P.S.; Duggal, N.K.; Hook, S.A.; Delorey, M.; Fischer, M.; Olzenak McGuire, D.; Becksted, H.; Max, R.J.; Anishchenko, M.; Schwartz, A.M.; et al. Zika Virus Shedding in Semen of Symptomatic Infected Men. N. Engl. J. Med. 2018, 378, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef]
- Musso, D.; Ko, A.I.; Baud, D. Zika Virus Infection—After the Pandemic. N. Engl. J. Med. 2019, 381, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Countries and Territories with Current or Previous Zika Virus Transmission, 1 by WHO Regional Office; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Soghaier, M.A.; Abdelgadir, D.M.; Abdelkhalig, S.M.; Kafi, H.; Zarroug, I.M.A.; Sall, A.A.; Eldegai, M.H.; Elageb, R.M.; Osman, M.M.; Khogali, H. Evidence of pre-existing active Zika virus circulation in Sudan prior to 2012. BMC Res. Notes 2018, 11, 906. [Google Scholar] [CrossRef] [PubMed]
- Adam, A.; Schuttoff, T.; Reiche, S.; Jassoy, C. High seroprevalence of dengue virus indicates that dengue virus infections are frequent in central and eastern Sudan. Trop. Med. Int. Health 2018, 23, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Beier, J.; Adam, A.; Jassoy, C. West Nile Virus Seroprevalence and Cross-Neutralization in Sera from Eastern and Central Sudan. Vector Borne Zoonotic Dis. 2022, 22, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Rabe, I.B.; Staples, J.E.; Villanueva, J.; Hummel, K.B.; Johnson, J.A.; Rose, L.; Mts; Hills, S.; Wasley, A.; Fischer, M.; et al. Interim Guidance for Interpretation of Zika Virus Antibody Test Results. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Maeda, J. Review of diagnostic plaque reduction neutralization tests for flavivirus infection. Vet. J. 2013, 195, 33–40. [Google Scholar] [CrossRef]
- Collins, M.H.; McGowan, E.; Jadi, R.; Young, E.; Lopez, C.A.; Baric, R.S.; Lazear, H.M.; de Silva, A.M. Lack of Durable Cross-Neutralizing Antibodies Against Zika Virus from Dengue Virus Infection. Emerg. Infect. Dis. 2017, 23, 773–781. [Google Scholar] [CrossRef]
- Montoya, M.; Collins, M.; Dejnirattisai, W.; Katzelnick, L.C.; Puerta-Guardo, H.; Jadi, R.; Schildhauer, S.; Supasa, P.; Vasanawathana, S.; Malasit, P.; et al. Longitudinal Analysis of Antibody Cross-neutralization Following Zika Virus and Dengue Virus Infection in Asia and the Americas. J. Infect. Dis. 2018, 218, 536–545. [Google Scholar] [CrossRef]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.H.; Onlamoon, N.; Sewatanon, J.; Edupuganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.J.; Wilson, P.C.; et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus. Proc. Natl. Acad. Sci. USA 2016, 113, 7852–7857. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Supasa, P.; Wongwiwat, W.; Rouvinski, A.; Barba-Spaeth, G.; Duangchinda, T.; Sakuntabhai, A.; Cao-Lormeau, V.M.; Malasit, P.; Rey, F.A.; et al. Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 2016, 17, 1102–1108. [Google Scholar] [CrossRef]
- PAHO. Guidelines for Surveillance of Zika Virus Disease and Its Complications, 2018th ed.; PAHO: Washington, DC, USA, 2018. [Google Scholar]
- Langerak, T.; Brinkman, T.; Mumtaz, N.; Arron, G.; Hermelijn, S.; Baldewsingh, G.; Wongsokarijo, M.; Resida, L.; Rockx, B.; Koopmans, M.P.G.; et al. Zika Virus Seroprevalence in Urban and Rural Areas of Suriname, 2017. J. Infect. Dis. 2019, 220, 28–31. [Google Scholar] [CrossRef]
- Nurtop, E.; Moyen, N.; Dzia-Lepfoundzou, A.; Dimi, Y.; Ninove, L.; Drexler, J.F.; Gallian, P.; de Lamballerie, X.; Priet, S. A Report of Zika Virus Seroprevalence in Republic of the Congo. Vector Borne Zoonotic Dis. 2020, 20, 40–42. [Google Scholar] [CrossRef]
- Pastorino, B.; Sengvilaipaseuth, O.; Chanthongthip, A.; Vongsouvath, M.; Souksakhone, C.; Mayxay, M.; Thirion, L.; Newton, P.N.; de Lamballerie, X.; Dubot-Peres, A. Low Zika Virus Seroprevalence in Vientiane, Laos, 2003–2015. Am. J. Trop. Med. Hyg. 2019, 100, 639–642. [Google Scholar] [CrossRef]
- Sam, I.C.; Montoya, M.; Chua, C.L.; Chan, Y.F.; Pastor, A.; Harris, E. Low seroprevalence rates of Zika virus in Kuala Lumpur, Malaysia. Trans. R Soc. Trop. Med. Hyg. 2019, 113, 678–684. [Google Scholar] [CrossRef]
- Page, M.; Mattiuzzo, G.; Hassall, M.; Ashall, J.; Myhill, S.; Faulkner, V.; Hockley, J.; Atkinson, E.; Wilkinson, D.; Rigsby, P.; et al. WHO Collaborative Study to Assess the Suitability of the 1st International Standard for Antibody to Zika Virus; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Mattiuzzo, G.; Knezevic, I.; Hassall, M.; Ashall, J.; Myhill, S.; Faulkner, V.; Hockley, J.; Rigsby, P.; Wilkinson, D.E.; Page, M.; et al. Harmonization of Zika neutralization assays by using the WHO International Standard for anti-Zika virus antibody. NPJ Vaccines 2019, 4, 42. [Google Scholar] [CrossRef]
- Dick, G.W.; Kitchen, S.F.; Haddow, A.J. Zika virus. I. Isolations and serological specificity. Trans. R Soc. Trop. Med. Hyg. 1952, 46, 509–520. [Google Scholar] [CrossRef]
- Prevention, C.f.D.C.a. Zika Travel Information. Available online: https://wwwnc.cdc.gov/travel/page/zika-travel-information (accessed on 12 January 2024).
- Swanstrom, J.A.; Plante, J.A.; Plante, K.S.; Young, E.F.; McGowan, E.; Gallichotte, E.N.; Widman, D.G.; Heise, M.T.; de Silva, A.M.; Baric, R.S. Dengue Virus Envelope Dimer Epitope Monoclonal Antibodies Isolated from Dengue Patients Are Protective against Zika Virus. mBio 2016, 7, e01123-16. [Google Scholar] [CrossRef]
- Andrade, D.V.; Harris, E. Recent advances in understanding the adaptive immune response to Zika virus and the effect of previous flavivirus exposure. Virus Res. 2018, 254, 27–33. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Selection Criteria | No. Sera | ZIKV NT Pos. |
|---|---|---|
| DENV IgG ELISA-positive | 106 | 23 (21.7%) |
| DENV IgG ELISA-negative | 92 | 3 (3.2%) |
| Total | 198 | 26 |
| Neutralizing Titer (NT50) | ||||||
|---|---|---|---|---|---|---|
| Control Serum | ZIKV | DENV-1 | DENV-2 | DENV-3 | DENV-4 | YFV |
| 1st International Standard | 422 | 640 | 453 | 320 | 320 | 453 |
| Working Reagent | 1040 | >1280 | >1280 | >1280 | >1280 | 40 |
| DENV IgG | ZIKV | DENV-1 | DENV-2 | DENV-3 | DENV-4 | YFV | ||
|---|---|---|---|---|---|---|---|---|
| Serum | Panbio 1 | NT50 | IU/mL 2 | Screening 3 | NT50 4 | NT50 4 | Screening 3 | NT50 4 |
| RS1 | + | 40 | 64 | + | 160 | 80 | + | <20 |
| RS14 | + | 20 | 22 | + | 640 | 226 | + | <20 |
| RS21 | + | 28 | 39 | + | 320 | 113 | + | <20 |
| RS22 | + | 57 | 99 | + | 28 | 40 | + | <20 |
| RS25 | + | 57 | 99 | n. d. | 80 | 160 | + | <20 |
| RS26 | + | 20 | 22 | n. d. | 80 | 80 | + | <20 |
| RS36 | + | 20 | 22 | + | 113 | 80 | + | <20 |
| RS52 | + | 80 | 147 | + | 113 | 113 | + | <20 |
| RS82 | + | 28 | 39 | + | n. d. | n. d. | + | <20 |
| RSS86 | + | 28 | 39 | + | 226 | 160 | + | <20 |
| RS109 | + | 640 | 1310 | + | 226 | 160 | + | <20 |
| RS110 | + | 20 | 22 | + | 40 | 160 | + | n. d. |
| RS120 | + | 20 | 22 | + | 28 | 80 | + | <20 |
| RS125 | + | 40 | 64 | n. d. | n. d. | 640 | n. d. | <20 |
| NK9 | + | 28 | 39 | + | n. d. | 80 | + | 57 |
| NK41 | + | 905 | 1861 | neg. | <20 | <20 | + | <20 |
| NK61 | neg. | 226 | 450 | neg. | <20 | <20 | equiv. | 40 |
| NK77 | neg. | 320 | 645 | neg. | <20 | <20 | equiv. | <20 |
| NK109 | + | 28 | 39 | + | n. d. | 80 | + | 905 |
| K31 | + | 20 | 22 | equiv. | <20 | <20 | neg. | <20 |
| K39 | neg. | 226 | 450 | + | 160 | 113 | + | 80 |
| K72 | + | 57 | 99 | neg. | 28 | <20 | neg. | <20 |
| K198 | + | 113 | 215 | + | 640 | 320 | + | 226 |
| K222 | + | 113 | 215 | + | 160 | 453 | + | <20 |
| K224 | + | 28 | 39 | + | 80 | 113 | + | <20 |
| K228 | + | 28 | 39 | + | 905 | 160 | + | 28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adam, A.; Wenzel, R.; Unger, E.; Reiche, S.; Jassoy, C. Serological Evidence of Zika Virus Infections in Sudan. Viruses 2024, 16, 1045. https://doi.org/10.3390/v16071045
Adam A, Wenzel R, Unger E, Reiche S, Jassoy C. Serological Evidence of Zika Virus Infections in Sudan. Viruses. 2024; 16(7):1045. https://doi.org/10.3390/v16071045
Chicago/Turabian StyleAdam, Awadalkareem, Robert Wenzel, Elisabeth Unger, Sven Reiche, and Christian Jassoy. 2024. "Serological Evidence of Zika Virus Infections in Sudan" Viruses 16, no. 7: 1045. https://doi.org/10.3390/v16071045
APA StyleAdam, A., Wenzel, R., Unger, E., Reiche, S., & Jassoy, C. (2024). Serological Evidence of Zika Virus Infections in Sudan. Viruses, 16(7), 1045. https://doi.org/10.3390/v16071045