
Factors Associated with Neutralizing Antibody Responses following 2-Dose and 3rd Booster Monovalent COVID-19 Vaccination in Japanese People Living with HIV
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Purification of Polyclonal IgG Fractions
2.3. Production of Pseudotyped Viruses
2.4. Target Cell Preparation
2.5. Polyclonal IgG Neutralization Assay
2.6. Statistical Analysis
3. Results
3.1. Cohort Characteristics
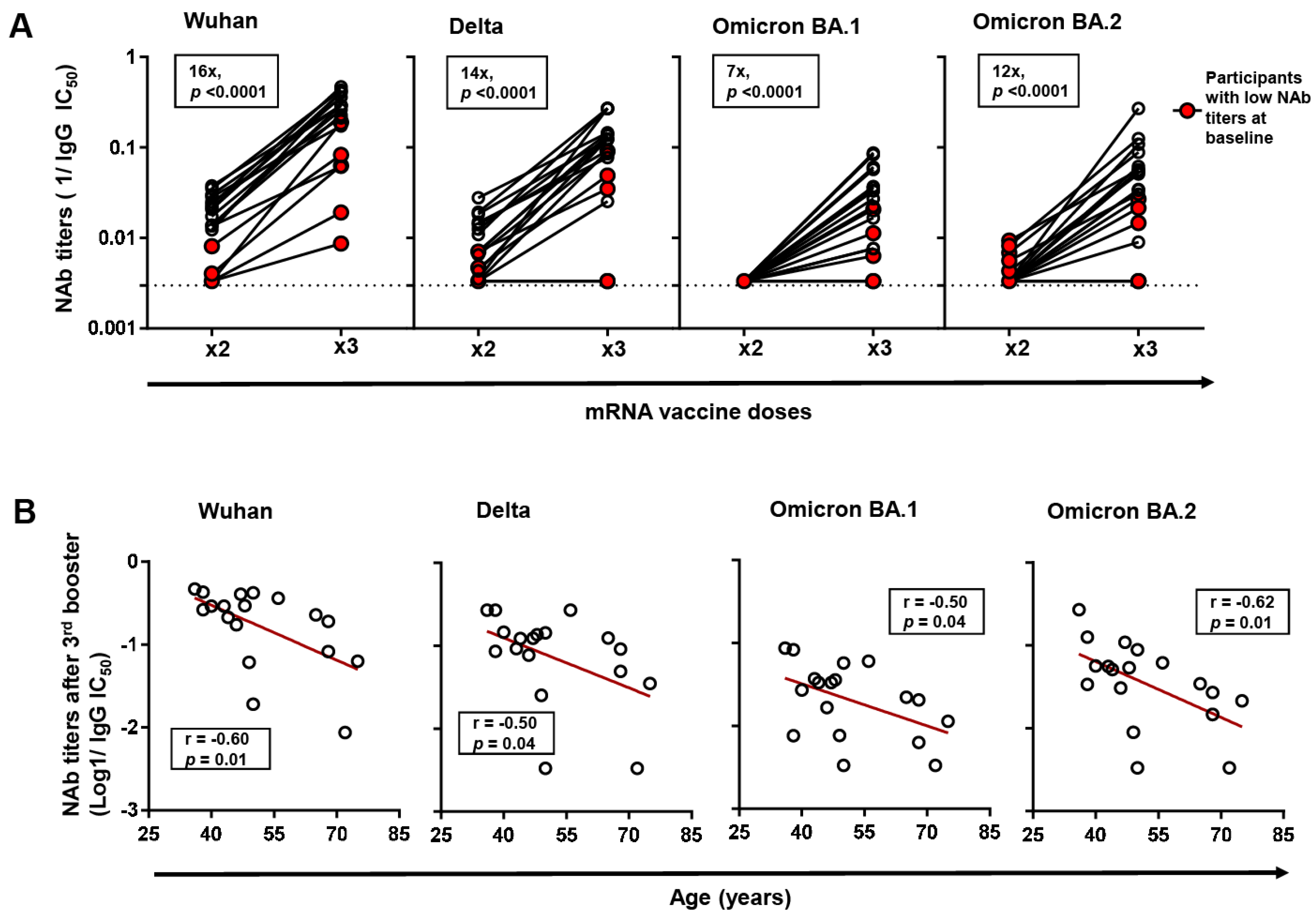
3.2. Neutralizing Antibody Titers against SARS-CoV-2 following 2-Dose Vaccination in PLWH
3.3. Neutralizing Antibody Titers against SARS-CoV-2 and Variants of Concern following Monovalent 3rd Booster Vaccination in PLWH
3.4. Factors Associated with Neutralizing Antibody Titers following COVID-19 Vaccination in PLWH
3.5. Factors Associated with Neutralizing Antibody Titers following COVID-19 Vaccination in Different Age Subgroups of PLWH
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Lapointe, H.R.; Mwimanzi, F.; Cheung, P.K.; Sang, Y.; Yaseen, F.; Umviligihozo, G.; Kalikawe, R.; Speckmaier, S.; Moran-Garcia, N.; Datwani, S.; et al. People with Human Immunodeficiency Virus Receiving Suppressive Antiretroviral Therapy Show Typical Antibody Durability after Dual Coronavirus Disease 2019 Vaccination and Strong Third Dose Responses. J. Infect. Dis. 2023, 227, 838–849. [Google Scholar] [CrossRef]
- Brumme, Z.L.; Mwimanzi, F.; Lapointe, H.R.; Cheung, P.K.; Sang, Y.; Duncan, M.C.; Yaseen, F.; Agafitei, O.; Ennis, S.; Ng, K.; et al. Humoral Immune Responses to COVID-19 Vaccination in People Living with HIV Receiving Suppressive Antiretroviral Therapy. npj Vaccines 2022, 7, 28. [Google Scholar] [CrossRef]
- Nault, L.; Marchitto, L.; Goyette, G.; Tremblay-Sher, D.; Fortin, C.; Martel-Laferrière, V.; Trottier, B.; Richard, J.; Durand, M.; Kaufmann, D.; et al. Covid-19 Vaccine Immunogenicity in People Living with HIV-1. Vaccine 2022, 40, 3633–3637. [Google Scholar] [CrossRef] [PubMed]
- Hassold, N.; Brichler, S.; Ouedraogo, E.; Leclerc, D.; Carroue, S.; Gater, Y.; Alloui, C.; Carbonnelle, E.; Bouchaud, O.; Mechai, F.; et al. Impaired Antibody Response to COVID-19 Vaccination in Advanced HIV Infection. AIDS 2022, 36, F1–F5. [Google Scholar] [CrossRef] [PubMed]
- Antinori, A.; Cicalini, S.; Meschi, S.; Bordoni, V.; Lorenzini, P.; Vergori, A.; Lanini, S.; De Pascale, L.; Matusali, G.; Mariotti, D.; et al. Humoral and Cellular Immune Response Elicited by mRNA Vaccination Against Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in People Living With Human Immunodeficiency Virus Receiving Antiretroviral Therapy Based on Current CD4 T-Lymphocyte Count. Clin. Infect. Dis. 2022, 75, e552–e563. [Google Scholar] [CrossRef] [PubMed]
- Touizer, E.; Alrubbayi, A.; Ford, R.; Hussain, N.; Gerber, P.P.; Shum, H.-L.; Rees-Spear, C.; Muir, L.; Gea-Mallorquí, E.; Kopycinski, J.; et al. Attenuated Humoral Responses in HIV after SARS-CoV-2 Vaccination Linked to B Cell Defects and Altered Immune Profiles. iScience 2022, 26, 105862. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, M.A.; Jones, B.L.H.; Gandhi, M. COVID-19 Outcomes and Risk Factors Among People Living with HIV. Curr. HIV/AIDS Rep. 2022, 19, 425–432. [Google Scholar] [CrossRef]
- Oliver, S. Updates to COVID-19 Vaccine Policy: Considerations for Future Planning. cdc.gov.2023. Available online: https://www.cdc.gov/vaccines/acip/meetings/downloads/slides-2023-04-19/06-COVID-Oliver-508.pdf (accessed on 27 March 2024).
- Zhang, W.; Ruan, L. Recent Advances in Poor HIV Immune Reconstitution: What Will the Future Look Like? Front. Microbiol. 2023, 14, 1236460. [Google Scholar] [CrossRef]
- Handoko, R.; Colby, D.J.; Kroon, E.; Sacdalan, C.; de Souza, M.; Pinyakorn, S.; Prueksakaew, P.; Munkong, C.; Ubolyam, S.; Akapirat, S.; et al. Determinants of Suboptimal CD4+ T Cell Recovery after Antiretroviral Therapy Initiation in a Prospective Cohort of Acute HIV-1 Infection. J. Int. AIDS Soc. 2020, 23, e25585. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Pallikkuth, S.; Pahwa, S. Metabolic Phenotype of B Cells from Young and Elderly HIV Individuals. Immun. Ageing 2021, 18, 35. [Google Scholar] [CrossRef] [PubMed]
- Jedicke, N.; Stankov, M.V.; Cossmann, A.; Dopfer-Jablonka, A.; Knuth, C.; Ahrenstorf, G.; Ramos, G.M.; Behrens, G.M.N. Humoral Immune Response Following Prime and Boost BNT162b2 Vaccination in People Living with HIV on Antiretroviral Therapy. HIV Med. 2022, 23, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Tau, L.; Turner, D.; Adler, A.; Marom, R.; Ahsanov, S.; Matus, N.; Levi, I.; Gerber, G.; Lev, S.; Ziv-Baran, T.; et al. SARS-CoV-2 Humoral and Cellular Immune Responses of Patients with HIV After Vaccination With BNT162b2 mRNA COVID-19 Vaccine in the Tel-Aviv Medical Center. Open Forum Infect. Dis. 2022, 9, ofac089. [Google Scholar] [CrossRef] [PubMed]
- Malin, J.J.; Suárez, I.; Biehl, L.M.; Schommers, P.; Knops, E.; Di Cristanziano, V.; Heger, E.; Pflieger, E.; Wyen, C.; Bettin, D.; et al. Immune Response to mRNA–Based COVID-19 Booster Vaccination in People Living with HIV. HIV Med. 2023, 24, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.S.; Toyoda, M.; Ode, H.; Barabona, G.; Hamana, H.; Kitamatsu, M.; Kishi, H.; Motozono, C.; Iwatani, Y.; Ueno, T. Dissecting Naturally Arising Amino Acid Substitutions at Position L452 of SARS-CoV-2 Spike. J. Virol. 2022, 96, e0116222. [Google Scholar] [CrossRef] [PubMed]
- Kaku, Y.; Kuwata, T.; Zahid, H.M.; Hashiguchi, T.; Noda, T.; Kuramoto, N.; Biswas, S.; Matsumoto, K.; Shimizu, M.; Kawanami, Y.; et al. Resistance of SARS-CoV-2 Variants to Neutralization by Antibodies Induced in Convalescent Patients with COVID-19. Cell Rep. 2021, 36, 109385. [Google Scholar] [CrossRef] [PubMed]
- Judicate, G.P.; Barabona, G.; Kamori, D.; Mahiti, M.; Tan, T.S.; Ozono, S.; Mgunya, A.S.; Kuwata, T.; Matsushita, S.; Sunguya, B.; et al. Phenotypic and Genotypic Co-Receptor Tropism Testing in HIV-1 Epidemic Region of Tanzania Where Multiple Non-B Subtypes Co-Circulate. Front. Microbiol. 2021, 12, 1788. [Google Scholar] [CrossRef] [PubMed]
- Ozono, S.; Zhang, Y.; Tobiume, M.; Kishigami, S.; Tokunaga, K. Super-Rapid Quantitation of the Production of HIV-1 Harboring a Luminescent Peptide Tag. J. Biol. Chem. 2020, 295, 13023–13030. [Google Scholar] [CrossRef]
- Ozono, S.; Zhang, Y.; Ode, H.; Sano, K.; Tan, T.S.; Imai, K.; Miyoshi, K.; Kishigami, S.; Ueno, T.; Iwatani, Y.; et al. SARS-CoV-2 D614G Spike Mutation Increases Entry Efficiency with Enhanced ACE2-Binding Affinity. Nat. Commun. 2021, 12, 1–9. [Google Scholar] [CrossRef]
- Montefiori, D.C. Measuring HIV Neutralization in a Luciferase Reporter Gene Assay. Methods Mol. Biol. 2009, 485, 395–405. [Google Scholar] [CrossRef]
- Schmidt, F.; Weisblum, Y.; Muecksch, F.; Hoffmann, H.H.; Michailidis, E.; Lorenzi, J.C.C.; Mendoza, P.; Rutkowska, M.; Bednarski, E.; Gaebler, C.; et al. Measuring SARS-CoV-2 Neutralizing Antibody Activity Using Pseudotyped and Chimeric Viruses. J. Exp. Med. 2020, 217, e20201181. [Google Scholar] [CrossRef]
- Cytoplasmic Tail Truncation of SARS-CoV-2 Spike Protein Enhances Titer of Pseudotyped Vectors But Masks the Effect of the D614G Mutation. Available online: https://journals.asm.org/doi/epdf/10.1128/jvi.00966-21?src=getftr (accessed on 30 December 2023).
- Burrer, R.; Spiridon, G.; Einius-Haessig, S.; Richert, S.; Salmon-Ceron, D.; Pancino, G.; Aubertin, A.M.; Moog, C. Short Communication: Efavirenz in Plasma from HIV-Infected Patients Does Not Directly Block Reverse Transcriptase Activity in Cell-Free Assays But Inhibits HIV Replication in Cellular Assays. AIDS Res. Hum. Retroviruses 2006, 22, 865–869. [Google Scholar] [CrossRef]
- Burrer, R.; Salmon-Ceron, D.; Richert, S.; Pancino, G.; Spiridon, G.; Haessig, S.; Roques, V.; Barre-Sinoussi, F.; Aubertin, A.-M.; Moog, C. Immunoglobulin G (IgG) and IgA, but Also Nonantibody Factors, Account for In Vitro Neutralization of Human Immunodeficiency Virus (HIV) Type 1 Primary Isolates by Serum and Plasma of HIV-Infected Patients. J. Virol. 2004, 78, 11450. [Google Scholar] [CrossRef]
- Klingler, J.; Weiss, S.; Itri, V.; Liu, X.; Oguntuyo, K.Y.; Stevens, C.; Ikegame, S.; Hung, C.-T.; Enyindah-Asonye, G.; Amanat, F.; et al. Role of Immunoglobulin M and A Antibodies in the Neutralization of Severe Acute Respiratory Syndrome Coronavirus 2. J. Infect. Dis. 2021, 223, 957–970. [Google Scholar] [CrossRef]
- Chammartin, F.; Kusejko, K.; Pasin, C.; Trkola, A.; Briel, M.; Amico, P.; Stoekle, M.P.; Eichenberger, A.L.; Hasse, B.; Braun, D.L.; et al. Determinants of Antibody Response to Severe Acute Respiratory Syndrome Coronavirus 2 mRNA Vaccines in People with HIV. AIDS 2022, 36, 1465. [Google Scholar] [CrossRef]
- Steensels, D.; Pierlet, N.; Penders, J.; Mesotten, D.; Heylen, L. Comparison of SARS-CoV-2 Antibody Response Following Vaccination With BNT162b2 and mRNA-1273. JAMA 2021, 326, 1533–1535. [Google Scholar] [CrossRef]
- Lau, B.; Sharrett, R.; Kingsley, L.A.; Post, W.; Palella, F.J.; Visscher, B.; Gange, S.J. C-Reactive Protein Is a Marker for Human Immunodeficiency Virus Disease Progression. Arch. Intern. Med. 2006, 166, 64–70. [Google Scholar] [CrossRef]
- Vergori, A.; Lepri, A.C.; Cicalini, S.; Matusali, G.; Bordoni, V.; Lanini, S.; Meschi, S.; Iannazzo, R.; Mazzotta, V.; Colavita, F.; et al. Immunogenicity to COVID-19 mRNA Vaccine Third Dose in People Living with HIV. Nat. Commun. 2022, 13, 4922. [Google Scholar] [CrossRef]
- Gianserra, L.; Donà, M.G.; Giuliani, E.; Stingone, C.; Pontone, M.; Buonomini, A.R.; Giuliani, M.; Pimpinelli, F.; Morrone, A.; Latini, A. Immunogenicity and Safety of BNT162b2 Homologous Booster Vaccination in People Living with HIV under Effective cART. Vaccines 2022, 10, 1243. [Google Scholar] [CrossRef]
- Miyazaki, R.; Miyagi, K.; Yoshida, M.; Suzuki, Y.; Hibino, S. Robust Antibody Response after the Third mRNA Coronavirus Vaccination in Japanese Hemodialysis Patients. Ren. Replace. Ther. 2023, 9, 38. [Google Scholar] [CrossRef]
- Fujigaki, H.; Yamamoto, Y.; Koseki, T.; Banno, S.; Ando, T.; Ito, H.; Fujita, T.; Naruse, H.; Hata, T.; Moriyama, S.; et al. Antibody Responses to BNT162b2 Vaccination in Japan: Monitoring Vaccine Efficacy by Measuring IgG Antibodies against the Receptor-Binding Domain of SARS-CoV-2. Microbiol. Spectr. 2022, 10, e0118121. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, T.; Ikeda, K.; Tanaka, S.; Taniguchi, T.; Igari, H.; Onouchi, Y.; Kaneda, A.; Matsushita, K.; Hanaoka, H.; Nakada, T.-A.; et al. Antibody Responses to BNT162b2 mRNA COVID-19 Vaccine and Their Predictors among Healthcare Workers in a Tertiary Referral Hospital in Japan. Clin. Microbiol. Infect. 2021, 27, 1861.e1–1861.e5. [Google Scholar] [CrossRef] [PubMed]
- Pallikkuth, S.; De Armas, L.R.; Pahwa, R.; Rinaldi, S.; George, V.K.; Sanchez, C.M.; Pan, L.; Dickinson, G.; Rodriguez, A.; Fischl, M.; et al. Impact of Aging and HIV Infection on Serologic Response to Seasonal Influenza Vaccination. AIDS 2018, 32, 1085. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.L.; Linterman, M.A. Mechanisms Underpinning Poor Antibody Responses to Vaccines in Ageing. Immunol. Lett. 2022, 241, 1–14. [Google Scholar] [CrossRef]
- Wing, E.J. HIV and Aging. Int. J. Infect. Dis. 2016, 53, 61–68. [Google Scholar] [CrossRef]
- Lombardi, A.; Butta, G.M.; Donnici, L.; Bozzi, G.; Oggioni, M.; Bono, P.; Matera, M.; Consonni, D.; Ludovisi, S.; Muscatello, A.; et al. Anti-Spike Antibodies and Neutralising Antibody Activity in People Living with HIV Vaccinated with COVID-19 mRNA-1273 Vaccine: A Prospective Single-Centre Cohort Study. Lancet Reg. Health-Eur. 2022, 13, 100287. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| 2-Dose Vaccinated Participants (n = 82) | |
|---|---|
| Age in years, median (IQR) | 48 (40–56) |
| Male, n (%) | 80 (98) |
| mRNA vaccine received c | |
| BNT162b2, n (%) | 48 (71.6) |
| mRNA-1273, n (%) | 19 (28.4) |
| Days since 2-dose vaccination | 53 (27–83) |
| Receiving ART, n (%) | 82 (100) |
| Pre-ART HIV viral load in log copies/mL, median (IQR) | 4.8 (3.1–5.2) a |
| Undetectable HIV viral load (at sample collection) n, (%) | 74 (94%) b |
| Pre-ART CD4+ count in cells/µL, median (IQR) | 211 (28–376) a |
| CD4+ count (at sample collection) in cells/µL, median (IQR) | 470 (314–643) a |
| CD4+/CD8+ ratio (at sample collection), median (IQR) | 0.9 (0.4–1.1) a |
| hsCRP (at sample collection) in mg/dL, median (IQR) | 0.12 (0.05–0.3) c |
| 3rd Booster Vaccinated Participants (n = 18) d | |
| Age in years, median (IQR) | 49 (36–66) |
| Male, n (%) | 18 (100) |
| mRNA booster received b | |
| BNT162b2, n (%) | 5 (33.3) |
| mRNA-1273, n (%) | 10 (66.7) |
| Days since 3rd booster vaccination, median (IQR) | 51 (37–70) |
| Days from 2-dose to 3rd booster vaccination, median (IQR) | 207 (196–215) |
| Variables | Bivariate | Multivariate a |
|---|---|---|
| Age | −0.42 (p = 0.001) | −0.33 (p = 0.007) |
| Days since 2-dose vaccination | −0.45 (p = 0.001) | −0.34 (p = 0.005) |
| Pre-ART viral load (copies/mL) | 0.21 (p = 0.087) | 0.14 (p = 0.266) |
| Pre-ART CD4+ count (cells/µL) | 0.18 (p = 0.141) | −0.05 (p = 0.691) |
| CD4+ count (at sample collection) (cells/µL) | 0.28 (p = 0.017) | 0.17 (p = 0.158) |
| CD4+/CD8+ ratio (at sample collection) | 0.15 (p = 0.204) | 0.04 (p = 0.767) |
| hsCRP (at sample collection) (mg/dL) | −0.16 (p = 0.193) | −0.18 (p = 0.158) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngare, I.; Tan, T.S.; Toyoda, M.; Kuwata, T.; Takahama, S.; Nakashima, E.; Yamasaki, N.; Motozono, C.; Fujii, T.; Minami, R.; et al. Factors Associated with Neutralizing Antibody Responses following 2-Dose and 3rd Booster Monovalent COVID-19 Vaccination in Japanese People Living with HIV. Viruses 2024, 16, 555. https://doi.org/10.3390/v16040555
Ngare I, Tan TS, Toyoda M, Kuwata T, Takahama S, Nakashima E, Yamasaki N, Motozono C, Fujii T, Minami R, et al. Factors Associated with Neutralizing Antibody Responses following 2-Dose and 3rd Booster Monovalent COVID-19 Vaccination in Japanese People Living with HIV. Viruses. 2024; 16(4):555. https://doi.org/10.3390/v16040555
Chicago/Turabian StyleNgare, Isaac, Toong Seng Tan, Mako Toyoda, Takeo Kuwata, Soichiro Takahama, Eriko Nakashima, Naoya Yamasaki, Chihiro Motozono, Teruhisa Fujii, Rumi Minami, and et al. 2024. "Factors Associated with Neutralizing Antibody Responses following 2-Dose and 3rd Booster Monovalent COVID-19 Vaccination in Japanese People Living with HIV" Viruses 16, no. 4: 555. https://doi.org/10.3390/v16040555
APA StyleNgare, I., Tan, T. S., Toyoda, M., Kuwata, T., Takahama, S., Nakashima, E., Yamasaki, N., Motozono, C., Fujii, T., Minami, R., Barabona, G., & Ueno, T. (2024). Factors Associated with Neutralizing Antibody Responses following 2-Dose and 3rd Booster Monovalent COVID-19 Vaccination in Japanese People Living with HIV. Viruses, 16(4), 555. https://doi.org/10.3390/v16040555