
Development and Application of Attenuated Plant Viruses as Biological Control Agents in Japan
 ,
,
Abstract
1. Introduction
2. History of the Development of Attenuated Viruses in Japan
2.1. Attenuated Virus Strains for Mosaic Disease of Cucumber
2.2. Attenuated Virus Strains Inhibit Mosaic Disease of Green Pepper
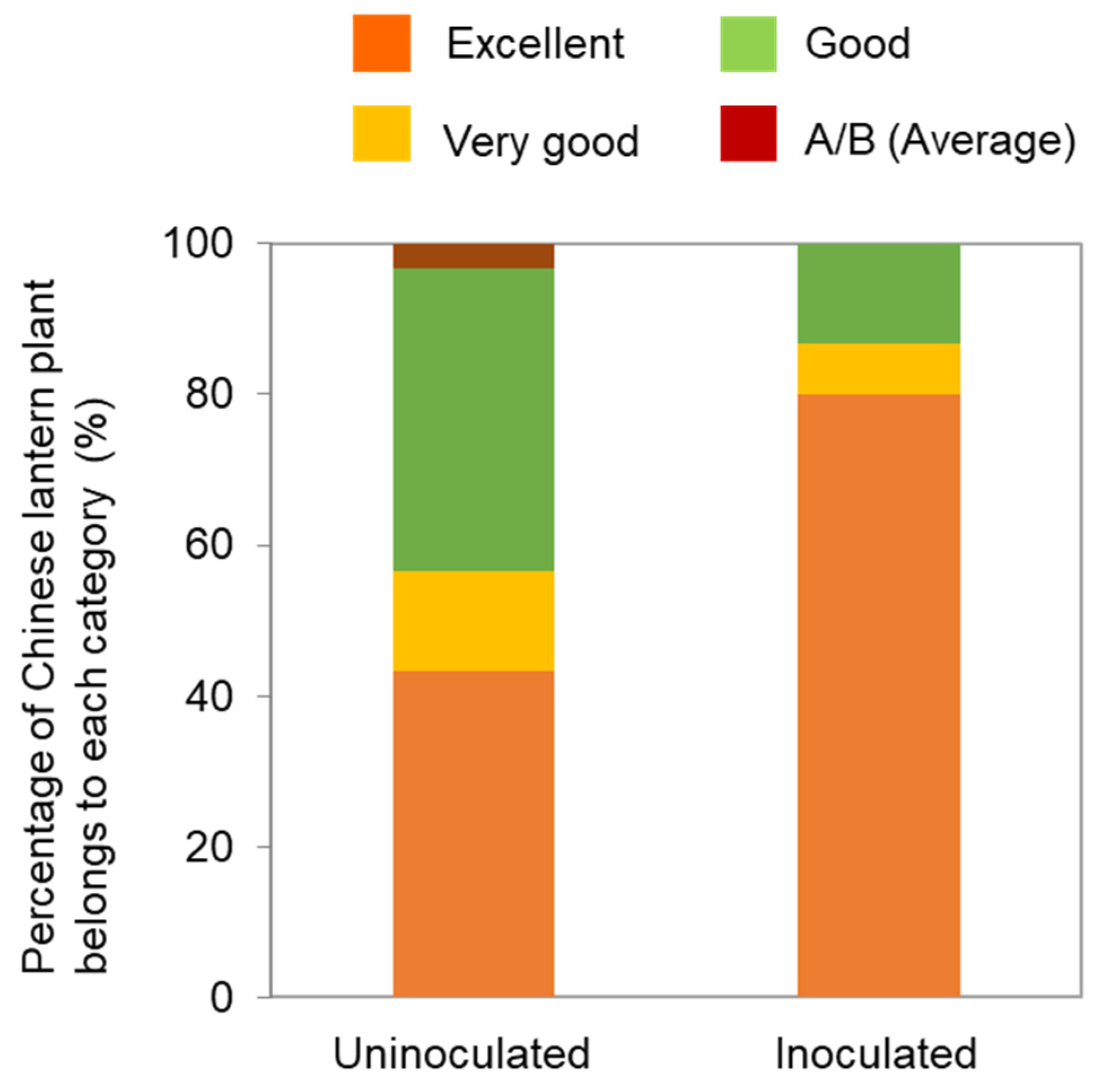
2.3. Attenuated Virus Strains for Mosaic and Necrotic Disease of Chinese Lantern Plants
2.4. Attenuated Viral Strains against Cucumber Spotted Wilt Disease
3. Inoculation of Attenuated Viruses
4. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mckinney, H.H. Mosaic diseases in the Canary Islands, West Africa, and Gibraltar. J. Agric. Res. 1929, 39, 557–578. [Google Scholar]
- Gal-On, A.; Shiboleth, Y.M. Cross-protection. In Natural Resistance Mechanisms of Plants to Viruses; Loebenstein, G., Carr, J.P., Eds.; Springer: Berlin, Germany, 2006; pp. 261–288. [Google Scholar]
- Rosner, A.; Lee, R.F.; Bar-Joseph, M. Differential hybridization with cloned cDNA sequences for detecting specific isolate of citrus tristeza virus. Phytopathology 1986, 76, 820–824. [Google Scholar] [CrossRef]
- Salaman, R.N. Protective inoculation against a plant virus. Nature 1933, 131, 468. [Google Scholar] [CrossRef]
- Webb, R.E.; Larson, R.H.; Walker, J.C. Relationships of potato leaf roll virus strains. Res. Bull. Wis. Agric. Exp. Stat. 1952, 178, 38. [Google Scholar]
- Holmes, F.O. A masked strain of tobacco-mosaic virus. Phytopathology 1934, 24, 845–873. [Google Scholar]
- Grant, T.J.; Costa, A.S. A mild strain of tristeza virus of citurs. Phytopathology 1951, 41, 114–122. [Google Scholar]
- Posnette, A.F.; Todd, J.M. Viruse diseases of Cacao in West Africa IX. Strain variation and interference in virus 1A. Ann. Appl. Biol. 1955, 43, 433–453. [Google Scholar] [CrossRef]
- Lecoq, H. Control of plant virus diseases by cross protection. In Plant Virus Disease Control; Hadidi, A., Khetarpal, R.K., Koganezawa, H., Eds.; APS Publishing: St. Paul, MN, USA, 1998; pp. 33–40. [Google Scholar]
- Fulton, R.W. Practices and precautions in the use of cross protection for plant virus disease control. Annu. Rev. Phytopathol. 1986, 24, 67–81. [Google Scholar] [CrossRef]
- Ziebell, H.; Carr, J.P. Cross-protection: A century of mystery. Adv. Virus Res. 2010, 76, 211–264. [Google Scholar] [PubMed]
- Van der Vlugt, R.A.A.; Stijger, C.C.M.M.; Verhoeven, J.T.J.; Lesemann, D.E. First report of Pepino mosaic virus on tomato. Plant Dis. 2000, 84, 103. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Conclusion on the peer review of the pesticide risk assessment of the active substance pepino mosaic virus strain CH2 isolate 1906. EFSA J. 2015, 13, 3977. [Google Scholar] [CrossRef][Green Version]
- European Food Safety Authority (EFSA). Conclusion on the peer review of the pesticide risk assessment of the active substance mild pepino mosaic virus isolate VX1. EFSA J. 2017, 15, e04650. [Google Scholar]
- European Food Safety Authority (EFSA). Conclusion on the peer review of the pesticide risk assessment of the active substance mild pepino mosaic virus isolate VC1. EFSA J. 2017, 15, 4651. [Google Scholar]
- Agüero, J.; Gόmez-Aix, C.; Sempere, R.N.; García-Villalba, J.; García-Núñez, J.; Hernando, Y.; Aranda, M.A. Stable and broad spectrum cross-protection against pepino mosaic virus attained by mixed infection. Front. Plant Sci. 2018, 9, 1810. [Google Scholar] [CrossRef]
- Hernando, Y.; Aranda, M.A. Cross-protection against pepino mosaic virus, more than a decade of efficient disease control. Ann. Appl. Biol. 2023, 184, 174–182. [Google Scholar] [CrossRef]
- Nishiguchi, M. Control of plant virus diseases by attenuated viruses: From past and present to future. Jpn. J. Pestic. Sci. 2017, 42, 326–333. [Google Scholar] [CrossRef]
- Kameya-Iwaki, M. Control of virus disease by attenuated viruses. Agric. Hortic. 1994, 69, 137–142. [Google Scholar]
- Hanada, K. Control of plant virus disease by attenuated virus and transgenic plant in Japan. In Proceedings of the International Workshop on the Pest Management Strategies in Asian Monsoon Agroecosystems, Kumamoto, Japan, 15–18 November 1995; pp. 293–299. [Google Scholar]
- Natsuaki, T. Studies on molecular mechanisms of viral attenuation and cross protection. J. Gen. Plant Pathol. 2011, 77, 354–357. [Google Scholar] [CrossRef]
- Nishiguchi, M.; Kobayashi, K. Attenuated plant viruses: Preventing virus diseases and understanding the molecular mechanism. J. Gen. Plant Pathol. 2011, 77, 221–229. [Google Scholar] [CrossRef][Green Version]
- Ieki, H.; Yamaguchi, A.; Kano, T.; Koizumi, M.; Iwanami, T. Control of stem pitting disease caused by citrus tristeza virus using protective mild strains in navel orange. Jpn. J. Phytopathol. 1997, 63, 170–175. [Google Scholar] [CrossRef]
- Sasaki, A. Studies on Hassaku dwarf. Spec. Bull. Fruit Tree Exp. Stn. Hiroshima 1974, 2, 1–106. [Google Scholar]
- Zenbayashi, R.; Hanada, K.; Iwaki, M.; Shibukawa, S. A strain of cucumber mosaic virus (CMV-SR) causing rugose symptom on spinach. Jpn. J. Phytopathol. 1983, 49, 716–719. [Google Scholar] [CrossRef]
- Hanada, K.; Tochihara, H. Agriculture useful microorganismes. Its use and prospects. Sponsor Minist. Agric. For. Fish. Natl. Agric. Res. Cent. 1990, 18, 70–78. [Google Scholar]
- Sayama, H. Field testing of a satellite-containing attenuated strain of cucumber mosaic virus for tomato protection in Japan. Phytopathology 1993, 83, 405–410. [Google Scholar] [CrossRef]
- Yoshida, K.; Goto, T.; Iizuka, N. Attenuated isolates of cucumber mosaic virus produced by satellite RNA and cross protection between attenuated isolates and virulent ones. Jpn. J. Phytopathol. 1985, 51, 238–242. [Google Scholar] [CrossRef]
- Nakazono-Nagaoka, E.; Takahashi, T.; Shimizu, T.; Kosaka, Y.; Natsuaki, T.; Omura, T.; Sasaya, T. Cross-protection against Bean yellow mosaic virus (BYMV) and clover yellow vein virus by attenuated BYMV isolate M11. Phytopathology 2009, 99, 251–257. [Google Scholar] [CrossRef]
- Sato, H.; Seo, N.; Nakamura, S. Attenuated mutants of Lily mottle virus obtained by Tissu culture of Lily bulb. Hort. Res. 2012, 11, 113–120, (In Japanese with English Abstract). [Google Scholar] [CrossRef][Green Version]
- Ogawa, T.; Nakamura, S.; Sayama, M.; Ohshima, K. Attenuated mutants of potato virus Y necrotic strain produced by nitrous acid treatment and mutagenesis-in-tissue culture methods. Eur. J. Plant Pathol. 2013, 135, 745–760. [Google Scholar] [CrossRef]
- Kosaka, Y. Attenuated isolates of soybean mosaic virus derived at a low temperature. Plant Dis. 1993, 77, 882–886. [Google Scholar] [CrossRef]
- Kosaka, Y.; Ryang, B.S.; Kobori, T.; Shiomi, H.; Yasuhara, H.; Kataoka, M. Effectiveness of an attenuated Zucchini yellow mosaic virus Isolate for Cross-Protecting Cucumber. Plant Dis. 2006, 90, 67–72. [Google Scholar] [CrossRef]
- Yamasaki, S.; Sakai, J.; Kamisoyama, S.; Goto, H.; Okuda, M.; Hanada, K. Control of russet crack disease in sweetpotato plants using a protective mild strain of Sweet potato feathery mottle virus. Plant Dis. 2009, 93, 190–194. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Motoyoshi, F.; Nishiguchi, M. Control of virus diseases by attenuated virus strains: Comparison between attenuated strains of cucumber green mottle mosaic virus and tobacco mosaic virus. Gamma Field Symp. 1988, 27, 91–109. [Google Scholar]
- Goto, T.; Nemoto, M. Studies on control of plant virus diseases by interference of attenuated virus-selection of TMV attenuated strain and influence on various plants inoculated with the attenuated strain. Res. Bull. Hokkaido Natl. Agric. Exp. Stn. 1971, 99, 67–76. (In Japanese) [Google Scholar]
- Ichiki, T.U.; Nagaoka, E.N.; Hagiwara, K.; Uchikawa, K.; Tsuda, S.; Omura, T. Integration of mutations responsible for the attenuated phenotype of Pepper mild mottle virus strains results in a symptomless cross-protecting strain. Arch. Virol. 2005, 150, 2009–2020. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Iizuka, N.; Komochi, S. Selection and utilization of an attenuated isolate of pepper strain of tobacco mosaic virus. Jpn. J. Phytopathol. 1984, 50, 221–228, (In Japanese with English Summary). [Google Scholar] [CrossRef]
- Nagai, Y. Production of C-1421, an attenuated mutant of pepper strain of tobacco mosaic virus. Jpn. J. Phytopathol. 1987, 53, 168–174. [Google Scholar] [CrossRef][Green Version]
- Ogai, R.; Kanda-Hojo, A.; Tsuda, S. An attenuated isolate of Pepper mild mottle virus for cross protection of cultivated green pepper (Capsicum annuum L.) carrying the L3 resistance gene. Crop Prot. 2013, 54, 29–34. [Google Scholar] [CrossRef]
- Kubo, S.; Tomaru, K.; Kimura, T.; Murano, H. Control of tobacco mosaic disease by inoculation with attenuated virus. Bull. Hatano Tob. Exp. Stn. 1973, 71, 303–320. (In Japanese) [Google Scholar]
- Yoneda, E.; Ishimatsu, H.; Tomitaka, Y. Cross protection effect of attenuated strains of tomato mosaic virus and tobacco mild green mosaic virus against mosaic and necrosis diseases of Chinese lantern plants (Physalis alkekengi L. var. franchetii M). Kyushu Plant Prot. Res. 2019, 65, 11–17. (In Japanese) [Google Scholar] [CrossRef]
- Oshima, N.; Komochi, S.; Goto, T. Study on control of plant virus diseases by vaccination of attenuated virus (1). Control of tomato mosaic disease. Hokkaido. Natl. Agric. Exp. Stn. Bull. 1965, 85, 23–33, (In Japanese with English Summary). [Google Scholar]
- Oshima, N.; Osawa, T.; Morita, H.; Mori, K. A new attenuated virus L11A237 (in Japanese with English abstract). Jpn. J. Phytopathol. 1978, 44, 504–508. [Google Scholar] [CrossRef]
- Shimomoto, Y.; Ishikawa, K.; Okada, T.; Oki, T.; Hayashi, K.; Yano, K.; Morita, Y.; Takeuchi, S. Isolation and characterization of an attenuated strain of an orthotospovirus, melon yellow spot virus. J. Gen. Plant Pathol. 2021, 87, 300–306. [Google Scholar] [CrossRef]
- Brégeon, D.; Sarasin, A. Hypothetical role of RNA damage avoidance in preventing human disease. Mutat. Res. 2005, 577, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y. Utilization of attenuated strain of tobacco mosaic virus for control of tomato mosaic disease. Shokubutsu Boeki 1984, 38, 345–348. (In Japanese) [Google Scholar]
- Aoki, H.; Ogiwara, S. Studies on control of tobacco mosaic virus by interference of attenuated virus on greenhouse tomatoes. Bull. Chiba Ken Agric. Exp. Stn. 1974, 14, 135–143. (In Japanese) [Google Scholar]
- Oshima, N. Control of tomato mosaic disease by attenuated virus. Jpn. Agric. Res. Q. 1981, 14, 222–228. [Google Scholar]
- Nishiguchi, M.; Kikuchi, S.; Kiho, Y.; Ohno, T.; Meshi, T.; Okada, Y. Molecular basis of plant viral virulence; the complete nucleotide sequence of an attenuated strain of tobacco mosaic virus. Nucleic Acids Res. 1985, 13, 5585–5590. [Google Scholar] [CrossRef][Green Version]
- Chen, H.; Ino, M.; Shimono, M.; Wagh, S.G.; Kobayashi, K.; Yaeno, T.; Yamaoka, N.; Bai, G.; Nishiguchi, M. A single amino acid substitution in the intervening region of 129K protein of cucumber green mottle mosaic virus resulted in attenuated symptoms. Phytopathology 2020, 110, 146–152. [Google Scholar] [CrossRef]
- Sayama, H.; Kominato, M.; Atarashi, H.; Takayanagi, N.; Yamada, M.; Hikage, T.; Yoshiike, T. Control of Cucumber mosaic virus (CMV) in gentian by satellite RNA containing attenuated CMV strains. Acta Hortic. 2006, 722, 147–153. [Google Scholar] [CrossRef]
- Iwasaki, M.; Inaba, T. Viral wilt of cucumber plants grafted on Squash rootstocks. Jpn. J. Phytopathol. 1988, 54, 584–592. [Google Scholar] [CrossRef]
- Kosaka, Y.; Fukunishi, T. Multiple inoculation with three attenuated viruses for the control of cucumber virus disease. Plant Dis. 1997, 81, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Wetter, C.; Conti, M.; Pepper mild mottle virus. No. 330. In: AAB Descriptions of Plant Viruses. Assoc. J. Appl. Biol. Warwick UK 1988. Available online: https://www.dpvweb.net/dpv/showdpv/?dpvno=330 (accessed on 11 February 2024).
- Ikegashira, Y.; Ohki, T.; Ichiki, U.T.; Higashi, T.; Hagiwara, K.; Omura, T.; Honda, Y.; Tsuda, S. An immunological system for the detection of pepper mild mottle virus in soil from green pepper fields. Plant Dis. 2004, 88, 650–656. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Report of the 28th Meeting of the Open-Ended Working Group of the Parties to the Montreal Protocol 2008. Available online: http://ozone.unep.org/Meeting_Documents/oewg/28oewg/OEWG-28-5E.pdf (accessed on 13 February 2024).
- Boukema, I.W. Resistance to TMV in Capsicum chacoense Hunz. is governed by an allele of the L-locus. Capsicum Newsl. 1984, 3, 47–48. [Google Scholar]
- Rast, A.T.B. Pepper tobamoviruses and pathotypes used in resistance breeding. Capsicum Newsl. 1989, 7, 20–23. [Google Scholar]
- Tsuda, S.; Kirita, M.; Watanabe, Y. Characterization of a Pepper mild mottle Tobamovirus strain capable of overcoming the L3 gene-mediated resistance, distinct from the resistance-breaking Italian isolate. Mol. Plant Microbe Interact. 1998, 11, 327–331. [Google Scholar] [CrossRef]
- Hamada, H.; Tomita, R.; Iwadate, Y.; Kobayashi, K.; Munemura, I.; Takeuchi, S.; Hikichi, Y.; Suzuki, K. Cooperative effect of two amino acid mutations in the coat protein of Pepper mild mottle virus overcomes L3-mediated resistance in Capsicum plants. Virus Gen. 2007, 34, 205–214. [Google Scholar] [CrossRef]
- Ogai, R.; Kanda, A.; Kubota, K.; Tsuda, S. Characterization and field assessment of L3-163, an attenuated strain of Pepper mild mottle virus. In XV International Congress of Virology; The Unlimited World of Microbes: Sapporo, Japan, 2011; Final Program VI-PO23-7. [Google Scholar]
- Yoneda, E.; Kodama, Y.; Ishimatsu, T.; Tomitaka, Y. A survey of occurrence of viral diseases in Chinese lantern plants (Physalis alkekengi L. var. franchetii Makino)in Oita Prefecture, Japan. Kyushu Plant Prot. Res. 2017, 63, 23–29. (In Japanese) [Google Scholar] [CrossRef]
- Kato, K.; Hanada, K.; Kameya-Iwaki, M. Transmission mode, host range and electron microscopy of a pathogen causing a new disease of melon (Cucumis melo) in Japan. Ann. Phytopathol. Soc. Jpn. 1999, 65, 624–627. [Google Scholar] [CrossRef]
- Kato, K.; Handa, K.; Kameya-Iwaki, M. Melon yellow spot virus: A distinct species of the genus Tospovirus isolated from melon. Phytopathology 2000, 90, 422–426. [Google Scholar] [CrossRef]
- Takeuchi, S.; Okuda, M.; Hanada, K.; Kawada, Y.; Kameya, M. Spotted wilt disease of cucumber (Cucumis sativus) caused by Melon yellow spot virus. Jpn. J. Phytopathol. 2001, 67, 46–51, (In Japanese with English Summary). [Google Scholar] [CrossRef][Green Version]
- Peng, J.C.; Yeh, S.D.; Huang, L.H.; Li, J.T.; Cheng, Y.F.; Chen, T.C. Emerging threat of thrips-borne Melon yellow spot virus on melon and watermelon in Taiwan. Eur. J. Plant Pathol. 2011, 130, 205–214. [Google Scholar] [CrossRef]
- Hull, R. (Ed.) Matthew’s Plant Virology, 4th ed.; Academic Press: San Diego, CA, USA, 2002; p. 1029. [Google Scholar]
- Kobayashi, K.; Nishiguchi, M. (Eds.) Antiviral Resistance in Plants: Methods and Protocols; Springer: Berlin/Heidelberg, Germany, 2019; pp. 261–272. [Google Scholar]
- Folimonova, S.Y.; Achor, D.; Bar-Joseph, M. Walking together: Cross-protection, genome conservation, and the replication machinery of Citurs tristeza virus. Viruses 2020, 12, 1353. [Google Scholar] [CrossRef] [PubMed]
- Pechinger, K.; Chooi, K.M.; MacDiarmid, R.M.; Harper, S.J.; Ziebell, H. A new era for mild strain cross-protection. Viruses 2019, 11, 670. [Google Scholar] [CrossRef] [PubMed]
- Gonsalves, D. Control of papaya ringspot virus in Papaya: A case study. Annu. Rev. Phytopathol. 1998, 36, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S. A method to mechanically inoculate tomato plants with tomato chlorosis virus. Jpn. J. Phytopathol. 2022, 88, 98–104, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Kimura, S. A method to mechanically inoculate tomato plants with tomato yellow leaf curl virus. Jpn. J. Phytopathol. 2022, 88, 240–249, (In Japanese with English Abstract). [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Virus Name (Abbreviation) | Isolate | Method of Attenuated Virus Strain Development | Reference |
|---|---|---|---|---|
| Closterovirus | Citrus tristeza virus (CTV) | M-16A | Isolation by using host plants Heat treatment | [23] |
| HM-55 | Isolation by using host plants | [24] | ||
| Cucumovirus | Cucumber mosaic virus (CMV) | SR | Isolation by using host plants | [25] |
| SRO | Exchange of RNA genome | [26] | ||
| SRK | Exchange of RNA genome | [26] | ||
| K02 | Addition of satellite RNA | [27] | ||
| P+fl | Addition of satellite RNA | [28] | ||
| Potyvirus | Bean yellow mosaic virus (BYMV) | M11 | Heat treatment | [29] |
| Lily mottle virus (LMoV) | LMm76-2 | Tissue culture | [30] | |
| LMm93 | Tissue culture | [30] | ||
| Potato virus Y (PVY) | N-NA10 | Sodium nitrite treatment | [31] | |
| N-MY10 | Mutagenesis-in-tissue culture | [31] | ||
| Soybean mosaic virus (SMV) | – | Heat treatment | [32] | |
| Sweet potato feathery mottle virus (SPFMV) | 10-O | Isolation by using host plants | [34] | |
| Zucchini yellow mosaic virus (ZYMV) | ZYMV-2002 | Heat treatment | [33] | |
| Tobamovirus | Cucumber mild green mosaic virus (CGMMV) | SH33b | Sodium nitrite treatment | [35] |
| Pepper mild mottle virus (PMMoV) | Pa18 | Heat treatment | [38] | |
| C-1421 | Heat treatment | [39] | ||
| L3-163 | Heat treatment | [40] | ||
| Tobacco mosaic virus (TMV) | M3 | Sodium nitrite treatment | [41] | |
| Tobacco mild green mosaic virus (TMGMV) | No. 4 | Unknown | [42] | |
| Tomato mosaic virus (ToMV) | L11 | Heat treatment | [43] | |
| L11A | Isolation by using host plants | [36] | ||
| L11A237 | Isolation by using host plants | [44] | ||
| Tospovirus | Melon yellow spot virus (MYSV) | SA08-8 | Heat treatment | [45] |
| Treatment | Percentage of Plants Showing Symptoms | |
|---|---|---|
| Mosaic | Necrosis | |
| Greenhouse | ||
| Uninoculated | 100% | 0% |
| TMGMV-No. 4 | 30% | 0% |
| ToMV-L11A | 66.7% | 0% |
| TMGMV-No. 4 and ToMV-L11A | 23.3% | 0% |
| Field | ||
| Uninoculated | 0% | 16.7% |
| TMGMV-No. 4 | 0% | 47.4% |
| ToMV-L11A | 0% | 80.8% |
| TMGMV-No. 4 and ToMV-L11A | 0% | 0% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomitaka, Y.; Shimomoto, Y.; Ryang, B.-S.; Hayashi, K.; Oki, T.; Matsuyama, M.; Sekine, K.-T. Development and Application of Attenuated Plant Viruses as Biological Control Agents in Japan. Viruses 2024, 16, 517. https://doi.org/10.3390/v16040517
Tomitaka Y, Shimomoto Y, Ryang B-S, Hayashi K, Oki T, Matsuyama M, Sekine K-T. Development and Application of Attenuated Plant Viruses as Biological Control Agents in Japan. Viruses. 2024; 16(4):517. https://doi.org/10.3390/v16040517
Chicago/Turabian StyleTomitaka, Yasuhiro, Yoshifumi Shimomoto, Bo-Song Ryang, Kazusa Hayashi, Tomoka Oki, Momoko Matsuyama, and Ken-Taro Sekine. 2024. "Development and Application of Attenuated Plant Viruses as Biological Control Agents in Japan" Viruses 16, no. 4: 517. https://doi.org/10.3390/v16040517
APA StyleTomitaka, Y., Shimomoto, Y., Ryang, B.-S., Hayashi, K., Oki, T., Matsuyama, M., & Sekine, K.-T. (2024). Development and Application of Attenuated Plant Viruses as Biological Control Agents in Japan. Viruses, 16(4), 517. https://doi.org/10.3390/v16040517