


Use of Capsid Integrity-qPCR for Detecting Viral Capsid Integrity in Wastewater
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viral Stocks
2.2. PMAxx Optimisation
2.3. Wastewater Sample Collection
Wastewater Processing
2.4. Nucleic Acid Extraction
2.5. Real-Time Quantitative PCR (RT-qPCR) and Capsid Integrity-RT-qPCR (ci-RT-qPCR) Analysis
2.6. Data Analysis
3. Results
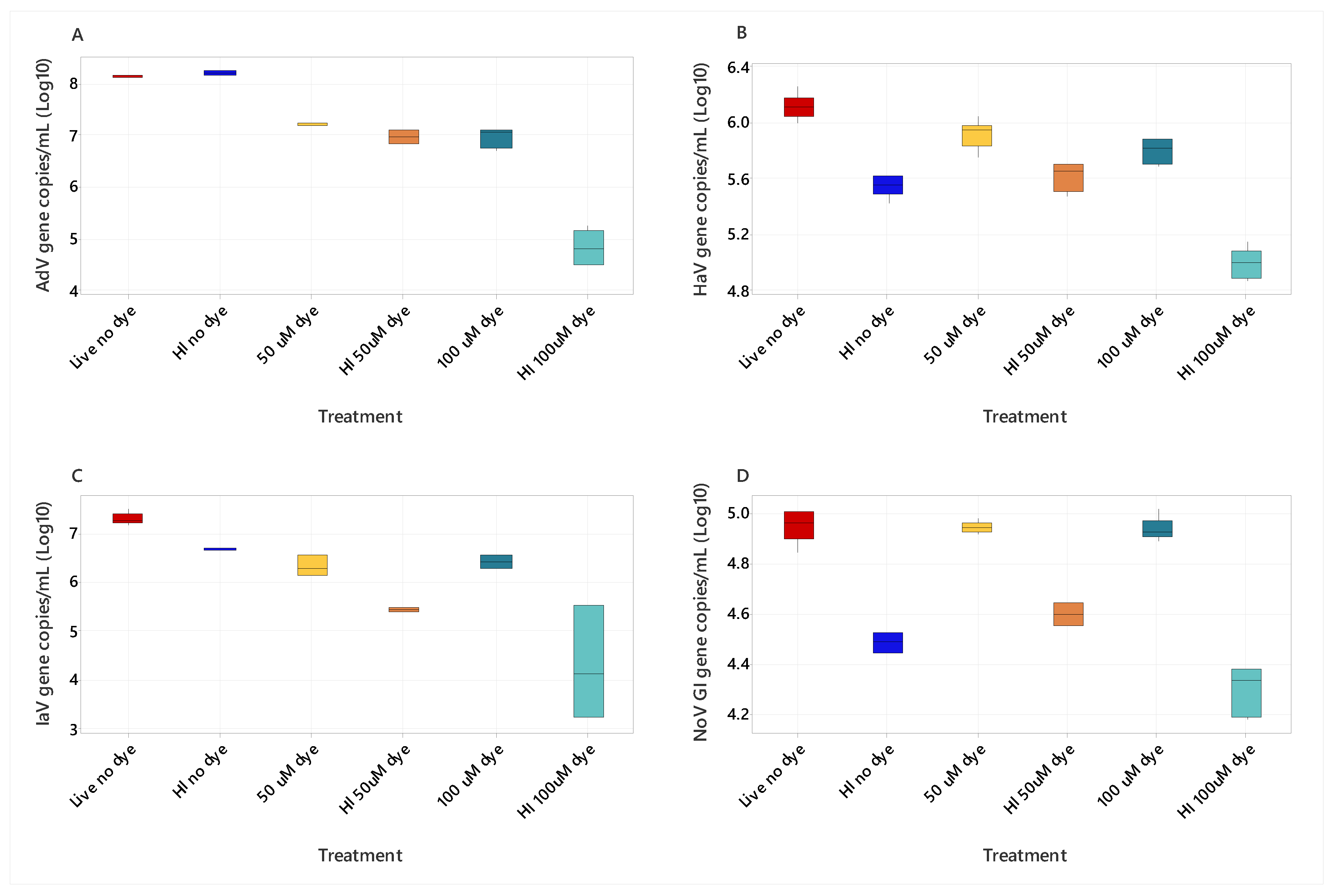
3.1. PMAXX Dye Concentration on Live and Heat-Inactivated Viruses (AdV, HAV, IAV, NoV GI)
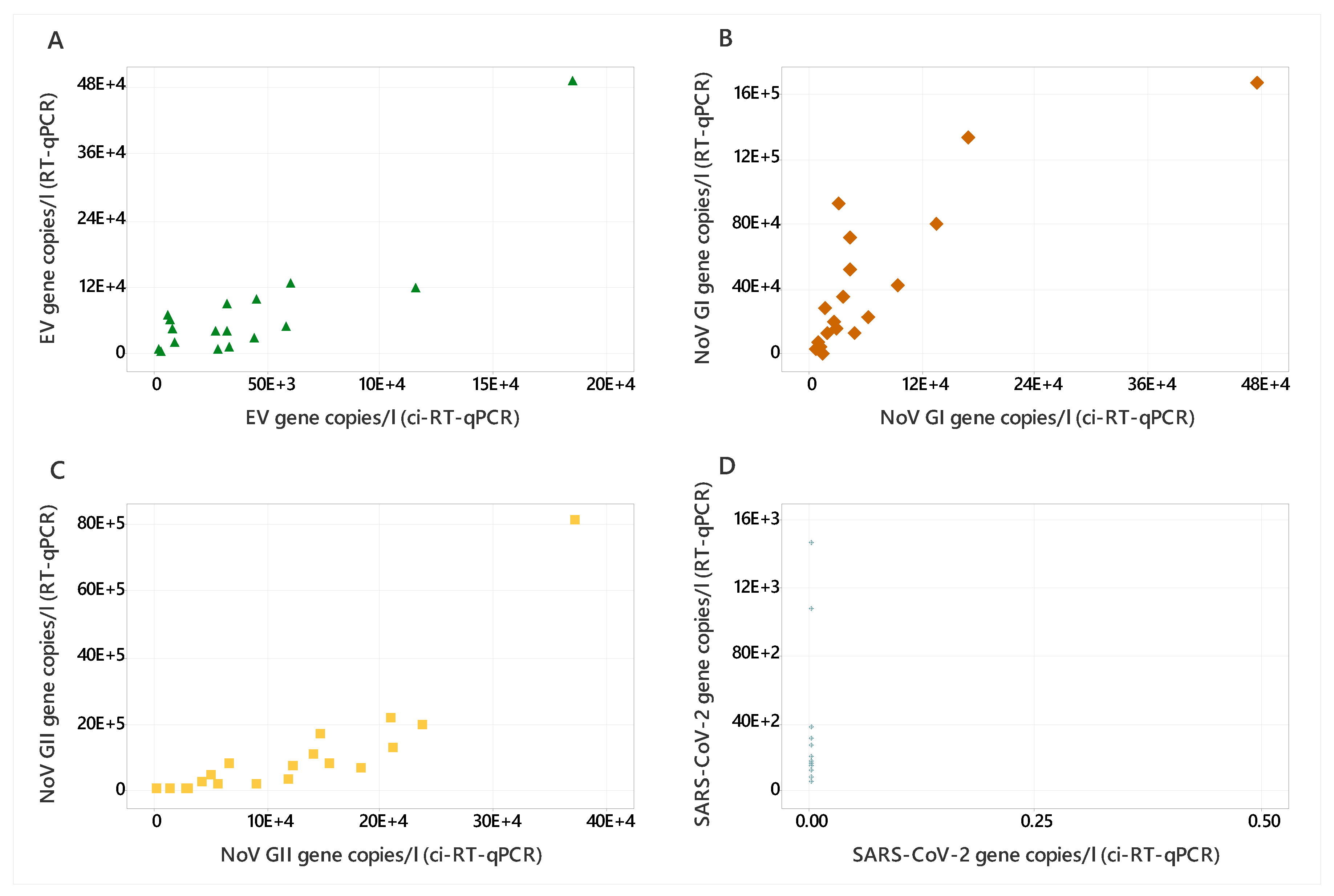
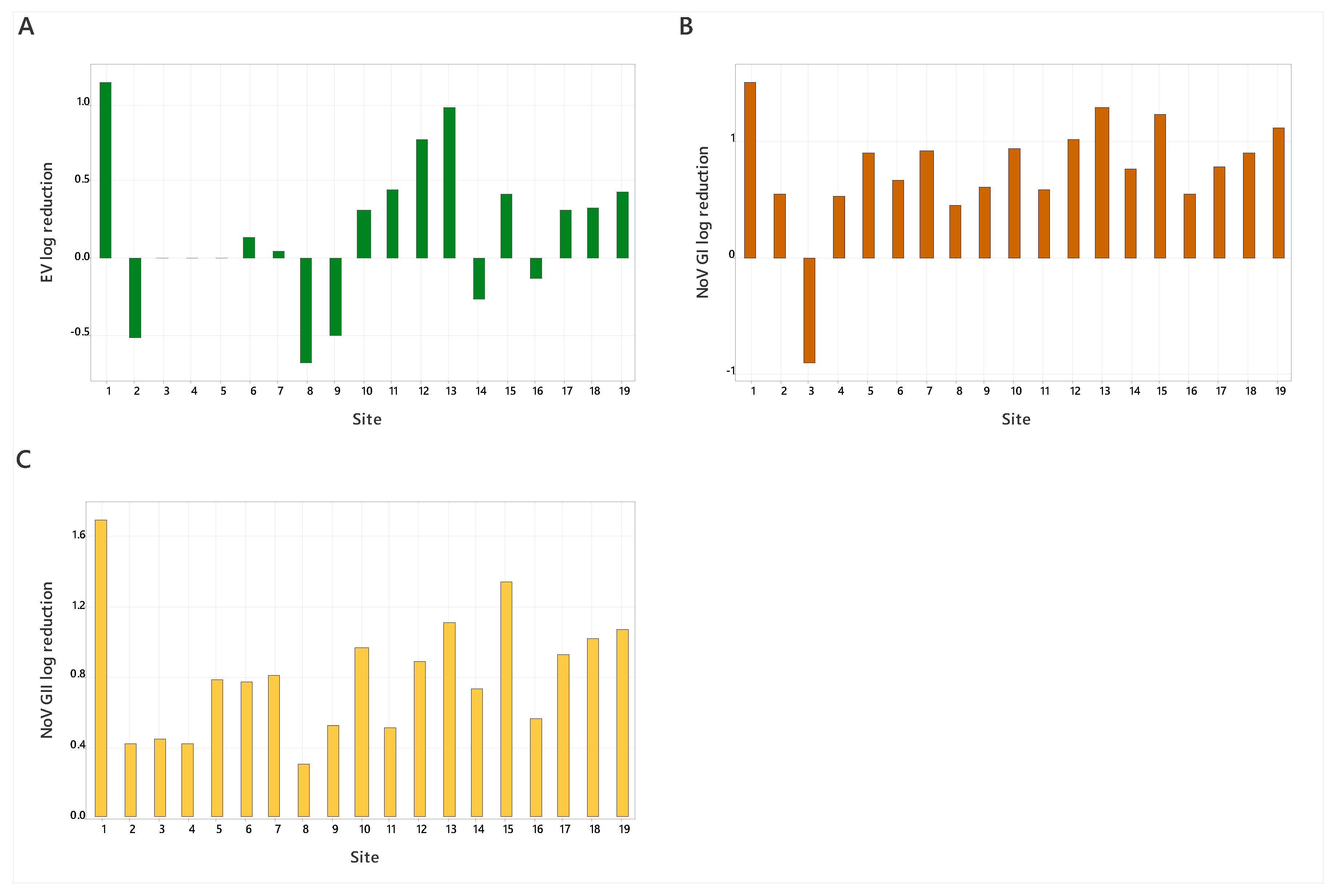
3.2. Wastewater Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barcellos, D.S.; Barquilha, C.E.R.; Oliveira, P.E.; Prokopiuk, M.; Etchepare, R.G. How Has the COVID-19 Pandemic Impacted Wastewater-Based Epidemiology? Sci. Total Environ. 2023, 892, 164561. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhou, N.; Zhang, S.; Yi, Y.; Han, Y.; Liu, M.; Han, Y.; Shi, N.; Yang, L.; Wang, Q.; et al. Norovirus Detection in Wastewater and Its Correlation with Human Gastroenteritis: A Systematic Review and Meta-Analysis. Environ. Sci. Pollut. Res. 2022, 29, 3. [Google Scholar] [CrossRef] [PubMed]
- Tedcastle, A.; Wilton, T.; Pegg, E.; Klapsa, D.; Bujaki, E.; Mate, R.; Fritzsche, M.; Majumdar, M.; Martin, J. Detection of Enterovirus D68 in Wastewater Samples from the UK between July and November 2021. Viruses 2022, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Kitakawa, K.; Kitamura, K.; Yoshida, H. Monitoring Enteroviruses and SARS-CoV-2 in Wastewater Using the Polio Environmental Surveillance System in Japan. Appl. Environ. Microbiol. 2023, 89, e01853-22. [Google Scholar] [CrossRef]
- Bisseux, M.; Debroas, D.; Mirand, A.; Archimbaud, C.; El Ene Peigue-Lafeuille, H.; Bailly, J.-L.; Henquell, E. Monitoring of Enterovirus Diversity in Wastewater by Ultra-Deep Sequencing: An Effective Complementary Tool for Clinical Enterovirus Surveillance. Water Res. 2019, 169, 115246. [Google Scholar] [CrossRef] [PubMed]
- Reyne, M.I.; Allen, D.M.; Levickas, A.; Allingham, P.; Lock, J.; Fitzgerald, A.; Mcsparron, C.; Nejad, B.F.; Mckinley, J.; Lee, A.; et al. Detection of Human Adenovirus F41 in Wastewater and Its Relationship to Clinical Cases of Acute Hepatitis of Unknown Aetiology. Sci. Total Environ. 2022, 857, 159579. [Google Scholar] [CrossRef] [PubMed]
- Takuissu, G.R.; Kenmoe, S.; Ebogo-Belobo, J.T.; Kengne-Ndé, C.; Mbaga, D.S.; Bowo-Ngandji, A.; Ondigui Ndzie, J.L.; Kenfack-Momo, R.; Tchatchouang, S.; Kenfack-Zanguim, J.; et al. Exploring Adenovirus in Water Environments: A Systematic Review and Meta-Analysis. Int. J. Environ. Health Res. 2023, 33. [Google Scholar] [CrossRef]
- Fong, T.T.; Phanikumar, M.S.; Xagoraraki, I.; Rose, J.B. Quantitative Detection of Human Adenoviruses in Wastewater and Combined Sewer Overflows Influencing a Michigan River. Appl. Environ. Microbiol. 2010, 76, 715–723. [Google Scholar] [CrossRef]
- Fantilli, A.; Cola, G.D.; Castro, G.; Sicilia, P.; Cachi, A.M.; de los Ángeles Marinzalda, M.; Ibarra, G.; López, L.; Valduvino, C.; Barbás, G.; et al. Hepatitis A Virus Monitoring in Wastewater: A Complementary Tool to Clinical Surveillance. Water Res. 2023, 241, 120102. [Google Scholar] [CrossRef]
- Rachida, S.; Taylor, M.B. Potentially Infectious Novel Hepatitis A Virus Strains Detected in Selected Treated Wastewater Discharge Sources, South Africa. Viruses 2020, 12, 1468. [Google Scholar] [CrossRef]
- Beyer, S.; Szewzyk, R.; Gnirss, R.; Johne, R.; Selinka, H.C. Detection and Characterization of Hepatitis E Virus Genotype 3 in Wastewater and Urban Surface Waters in Germany. Food Environ. Virol. 2020, 12, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Masclaux, F.G.; Hotz, P.; Friedli, D.; Savova-Bianchi, D.; Oppliger, A. High Occurrence of Hepatitis E Virus in Samples from Wastewater Treatment Plants in Switzerland and Comparison with Other Enteric Viruses. Water Res. 2013, 47, 5101–5109. [Google Scholar] [CrossRef] [PubMed]
- Boehm, A.B.; Hughes, B.; Duong, D.; Chan-Herur, V.; Buchman, A.; Wolfe, M.K.; White, B.J. Wastewater Concentrations of Human Influenza, Metapneumovirus, Parainfluenza, Respiratory Syncytial Virus, Rhinovirus, and Seasonal Coronavirus Nucleic-Acids during the COVID-19 Pandemic: A Surveillance Study. Lancet Microbe 2023, 4, e340–e348. [Google Scholar] [CrossRef]
- Wolfe, M.K.; Duong, D.; Bakker, K.M.; Ammerman, M.; Mortenson, L.; Hughes, B.; Arts, P.; Lauring, A.S.; Fitzsimmons, W.J.; Bendall, E.; et al. Wastewater-Based Detection of Two Influenza Outbreaks. Environ. Sci. Technol. Lett. 2022, 9, 687–692. [Google Scholar] [CrossRef]
- Toribio-Avedillo, D.; Gómez-Gómez, C.; Sala-Comorera, L.; Rodríguez-Rubio, L.; Carcereny, A.; García-Pedemonte, D.; Pintó, R.M.; Guix, S.; Galofré, B.; Bosch, A.; et al. Monitoring Influenza and Respiratory Syncytial Virus in Wastewater. Beyond COVID-19. Sci. Total Environ. 2023, 892, 164495. [Google Scholar] [CrossRef]
- Dzinamarira, T.; Murewanhema, G.; Iradukunda, P.G.; Madziva, R.; Herrera, H.; Cuadros, D.F.; Tungwarara, N.; Chitungo, I.; Musuka, G. Utilization of SARS-CoV-2 Wastewater Surveillance in Africa—A Rapid Review. Int. J. Environ. Res. Public. Health 2022, 19, 969. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar Patel, A.; Shah, A.V.; Raval, J.; Rajpara, N.; Joshi, M.; Joshi, C.G. First Proof of the Capability of Wastewater Surveillance for COVID-19 in India through Detection of Genetic Material of SARS-CoV-2. Sci. Total Environ. 2020, 746, 141326. [Google Scholar] [CrossRef]
- Yu, A.T.; Hughes, B.; Wolfe, M.K.; Leon, T.; Duong, D.; Rabe, A.; Kennedy, L.C.; Ravuri, S.; White, B.J.; Wigginton, K.R.; et al. Estimating Relative Abundance of 2 SARS-CoV-2 Variants through Wastewater Surveillance at 2 Large Metropolitan Sites, United States. Emerg. Infect. Dis. 2022, 28, 940. [Google Scholar] [CrossRef]
- Agrawal, S.; Orschler, L.; Schubert, S.; Zachmann, K.; Heijnen, L.; Tavazzi, S.; Gawlik, B.M.; de Graaf, M.; Medema, G.; Lackner, S. Prevalence and Circulation Patterns of SARS-CoV-2 Variants in European Sewage Mirror Clinical Data of 54 European Cities. Water Res. 2022, 214, 118162. [Google Scholar] [CrossRef]
- Brunner, F.S.; Brown, M.R.; Bassano, I.; Denise, H.; Khalifa, M.S.; Wade, M.J.; van Aerle, R.; Kevill, J.L.; Jones, D.L.; Farkas, K.; et al. City-Wide Wastewater Genomic Surveillance through the Successive Emergence of SARS-CoV-2 Alpha and Delta Variants. Water Res. 2022, 226, 119306. [Google Scholar] [CrossRef]
- Hillary, L.S.; Farkas, K.; Maher, K.H.; Lucaci, A.; Thorpe, J.; Distaso, M.A.; Gaze, W.H.; Paterson, S.; Burke, T.; Connor, T.R.; et al. Monitoring SARS-CoV-2 in Municipal Wastewater to Evaluate the Success of Lockdown Measures for Controlling COVID-19 in the UK. Water Res. 2021, 200, 117214. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Angel, N.; Edson, J.; Bibby, K.; Bivins, A.; O’brien, J.W.; Choi, P.M.; Kitajima, M.; Simpson, S.L.; Li, J.; et al. First Confirmed Detection of SARS-CoV-2 in Untreated Wastewater in Australia: A Proof of Concept for the Wastewater Surveillance of COVID-19 in the Community. Sci. Total Environ. 2020, 728, 138764. [Google Scholar] [CrossRef] [PubMed]
- Hassard, F.; Sharp, J.H.; Taft, H.; Levay, L.; Harris, J.P.; Mcdonald, J.E.; Tuson, K.; Wilson, J.; Jones, L.D.; Malham, S.K. Critical Review on the Public Health Impact of Norovirus Contamination in Shellfish and the Environment: A UK Perspective. Food Environ. Virol. 2017, 9, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Botturi, A.; Ozbayram, E.G.; Tondera, K.; Gilbert, N.I.; Rouault, P.; Caradot, N.; Gutierrez, O.; Daneshgar, S.; Frison, N.; Akyol, Ç.; et al. Combined Sewer Overflows: A Critical Review on Best Practice and Innovative Solutions to Mitigate Impacts on Environment and Human Health. Crit. Rev. Environ. Sci. Technol. 2021, 51, 1585–1618. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, G.-C.; Kim, J.I.; Yi, H.A.; Lee, C.H. Development of a New Cell Culture-Based Method and Optimized Protocol for the Detection of Enteric Viruses. J. Virol. Methods 2013, 191, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Iaconelli, M.; Muscillo, M.; Libera, S.D.; Fratini, M.; Meucci, L.; De Ceglia, M.; Giacosa, D.; Rosa, G. La One-Year Surveillance of Human Enteric Viruses in Raw and Treated Wastewaters, Downstream River Waters, and Drinking Waters. Food Environ. Virol. 2017, 9, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.J.; Ho, H.H.; Riggs, J.L.; Lennette, E.H. Comparative Sensitivity of Various Cell Culture Systems for Isolation of Viruses from Wastewater and Fecal Samples. Appl. Environ. Microbiol. 1978, 36, 480–486. [Google Scholar] [CrossRef]
- Fittipaldi, M.; Nocker, A.; Codony, F. Progress in Understanding Preferential Detection of Live Cells Using Viability Dyes in Combination with DNA Amplification. J. Microbiol. Methods 2012, 91, 276–289. [Google Scholar] [CrossRef]
- Prevost, B.; Goulet, M.; Lucas, F.S.; Joyeux, M.; Moulin, L.; Wurtzer, S. Viral Persistence in Surface and Drinking Water: Suitability of PCR Pre-Treatment with Intercalating Dyes. Water Res. 2016, 91, 68–76. [Google Scholar] [CrossRef]
- Randazzo, W.; Khezri, M.; Ollivier, J.; Le Guyader, F.S.; Rodríguez-Díaz, J.; Aznar, R.; Sánchez, G. Optimization of PMAxx Pretreatment to Distinguish between Human Norovirus with Intact and Altered Capsids in Shellfish and Sewage Samples. Int. J. Food Microbiol. 2018, 266, 1–7. [Google Scholar] [CrossRef]
- Wurtzer, S.; Waldman, P.; Ferrier-Rembert, A.; Frenois-Veyrat, G.; Mouchel, J.M.; Boni, M.; Maday, Y.; Consortium, O.; Marechal, V.; Moulin, L. Several Forms of SARS-CoV-2 RNA Can Be Detected in Wastewaters: Implication for Wastewater-Based Epidemiology and Risk Assessment. Water Res. 2021, 198, 117183. [Google Scholar] [CrossRef] [PubMed]
- Stobnicka-Kupiec, A.; Gołofit-Szymczak, M.; Cyprowski, M.; Górny, R.L. Detection and Identification of Potentially Infectious Gastrointestinal and Respiratory Viruses at Workplaces of Wastewater Treatment Plants with Viability QPCR/RT-QPCR. Sci. Rep. 2022, 12, 4517. [Google Scholar] [CrossRef] [PubMed]
- Palowski, A.; Balestreri, C.; Urriola, P.E.; van de Ligt, J.L.G.; Sampedro, F.; Dee, S.; Shah, A.; Yancy, H.F.; Shurson, G.C.; Schroeder, D.C. Survival of a Surrogate African Swine Fever Virus-like Algal Virus in Feed Matrices Using a 23-Day Commercial United States Truck Transport Model. Front. Microbiol. 2022, 13, 1059118. [Google Scholar] [CrossRef] [PubMed]
- Anfruns-Estrada, E.; Bottaro, M.; Pintó, R.M.; Guix, S.; Bosch, A. Effectiveness of Consumers Washing with Sanitizers to Reduce Human Norovirus on Mixed Salad. Foods 2019, 8, 637. [Google Scholar] [CrossRef] [PubMed]
- Shirasaki, N.; Matsushita, T.; Matsui, Y.; Koriki, S. Suitability of Pepper Mild Mottle Virus as a Human Enteric Virus Surrogate for Assessing the Efficacy of Thermal or Free-Chlorine Disinfection Processes by Using Infectivity Assays and Enhanced Viability PCR. Water Res. 2020, 186, 116409. [Google Scholar] [CrossRef] [PubMed]
- Leifels, M.; Jurzik, L.; Wilhelm, M.; Hamza, I.A. Use of Ethidium Monoazide and Propidium Monoazide to Determine Viral Infectivity upon Inactivation by Heat, UV- Exposure and Chlorine. Int. J. Hyg. Environ. Health 2015, 218, 686–693. [Google Scholar] [CrossRef]
- Canh, V.D.; Liu, M.; Sangsanont, J.; Katayama, H. Capsid Integrity Detection of Pathogenic Viruses in Waters: Recent Progress and Potential Future Applications. Sci. Total Environ. 2022, 827, 154258. [Google Scholar] [CrossRef]
- Leifels, M.; Cheng, D.; Sozzi, E.; Shoults, D.C.; Wuertz, S.; Mongkolsuk, S.; Sirikanchana, K. Capsid Integrity Quantitative PCR to Determine Virus Infectivity in Environmental and Food Applications—A Systematic Review. Water Res. X 2021, 11, 100080. [Google Scholar] [CrossRef]
- Leifels, M.; Hamza, I.A.; Krieger, M.; Wilhelm, M.; Mackowiak, M.; Jurzik, L. From Lab to Lake—Evaluation of Current Molecular Methods for the Detection of Infectious Enteric Viruses in Complex Water Matrices in an Urban Area. PLoS ONE 2016, 11, e0167105. [Google Scholar] [CrossRef]
- Parshionikar, S.; Laseke, I.; Fout, G.S. Use of propidium monoazide in reverse transcriptase PCR to distinguish between infectious and noninfectious enteric viruses in water samples. Appl. Environ. Microbiol. 2010, 76, 4318–4326. [Google Scholar] [CrossRef]
- Gyawali, P.; Hewitt, J. Detection of Infectious Noroviruses from Wastewater and Seawater Using PEMAXTM Treatment Combined with RT-QPCR. Water 2018, 10, 841. [Google Scholar] [CrossRef]
- Randazzo, W.; Piqueras, J.; Rodriguez, D.; Iaz, J.; Aznar, R.; Anchez, G.S. Improving Efficiency of Viability-QPCR for Selective Detection of Infectious HAV in Food and Water Samples. J. Appl. Microbiol. 2017, 124, 958–964. [Google Scholar] [CrossRef] [PubMed]
- Yee, R.A.; Leifels, M.; Scott, C.; Ashbolt, N.J.; Liu, Y. Evaluating Microbial and Chemical Hazards in Commercial Struvite Recovered from Wastewater. Environ. Sci. Technol. 2019, 53, 5378–5386. [Google Scholar] [CrossRef] [PubMed]
- Ogorzaly, L.; Cauchie, H.M.; Penny, C.; Perrin, A.; Gantzer, C.; Bertrand, I. Two-Day Detection of Infectious Enteric and Non-Enteric Adenoviruses by Improved ICC-QPCR. Appl. Microbiol. Biotechnol. 2013, 97, 4159–4166. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Bertsch, P.M.; Bibby, K.; Haramoto, E.; Hewitt, J.; Huygens, F.; Gyawali, P.; Korajkic, A.; Riddell, S.; Sherchan, S.P.; et al. Decay of SARS-CoV-2 and Surrogate Murine Hepatitis Virus RNA in Untreated Wastewater to Inform Application in Wastewater-Based Epidemiology. Environ. Res. 2020, 191, 110092. [Google Scholar] [CrossRef]
- Beattie, R.E.; Denene Blackwood, A.; Clerkin, T.; Dinga, C.; Noble, R.T. Evaluating the Impact of Sample Storage, Handling, and Technical Ability on the Decay and Recovery of SARS-CoV-2 in Wastewater. PLoS ONE 2022, 17, e0270659. [Google Scholar] [CrossRef]
- Farkas, K.; Hillary, L.S.; Thorpe, J.; Walker, D.I.; Lowther, J.A.; McDonald, J.E.; Malham, S.K.; Jones, D.L. Concentration and Quantification of SARS-CoV-2 RNA in Wastewater Using Polyethylene Glycol-Based Concentration and QRT-PCR. Methods Protoc. 2021, 4, 17. [Google Scholar] [CrossRef]
- Lewis, G.D.; Metcalf, T.G. Polyethylene Glycol Precipitation for Recovery of Pathogenic Viruses, Including Hepatitis A Virus and Human Rotavirus, from Oyster, Water, and Sediment Samples. Appl. Environ. Microbiol. 1988, 54, 1983–1988. [Google Scholar] [CrossRef]
- Kevill, J.L.; Pellett, C.; Farkas, K.; Brown, M.R.; Bassano, I.; Denise, H.; Mcdonald, J.E.; Malham, S.K.; Porter, J.; Warren, J.; et al. A Comparison of Precipitation and Filtration-Based SARS-CoV-2 Recovery Methods and the Influence of Temperature, Turbidity, and Surfactant Load in Urban Wastewater. Sci. Total Environ. 2021, 808, 151916. [Google Scholar] [CrossRef]
- WHO. Laboratory Biosafety Manual, 3rd ed.; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Farkas, K.; Pellett, C.; Alex-Sanders, N.; Bridgman, M.T.P.; Corbishley, A.; Grimsley, J.M.S.; Kasprzyk-Hordern, B.; Kevill, J.L.; Pântea, I.; Richardson-O’Neill, I.S.; et al. Comparative Assessment of Filtration- and Precipitation-Based Methods for the Concentration of SARS-CoV-2 and Other Viruses from Wastewater. Microbiol. Spectr. 2022, 10, e01102-22. [Google Scholar] [CrossRef] [PubMed]
- Dunn, O.J. Multiple Comparisons Using Rank Sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Kolassa, J.E.; Jankowski, S. R: Multivariate Nonparametric Methods. Available online: https://search.r-project.org/CRAN/refmans/MultNonParam/html/00Index.html (accessed on 1 November 2023).
- Dinno, A. Dunn’s Test of Multiple Comparisons Using Rank Sums. Available online: https://cran.r-project.org/web/packages/dunn.test/dunn.test.pdf (accessed on 1 November 2023).
- Cuevas-Ferrando, E.; Pérez-Cataluña, A.; Falcó, I.; Randazzo, W.; Sánchez, G. Monitoring Human Viral Pathogens Reveals Potential Hazard for Treated Wastewater Discharge or Reuse. Front. Microbiol. 2022, 13, 836193. [Google Scholar] [CrossRef] [PubMed]
- Morales, E.; Badilla, A.; Rivera, P.; Alfaro, E.; Symonds, E.M.; Barrantes, K.; Ruepert, C.; Crowe, J.; Chacón, L. A Novel Approach to Biological Risk Surveillance in Occupational Hygiene: A Case Study on the Use of Quantitative Microbial Risk Assessment to Determine Occupational Risks for Wastewater Treatment Plants Workers in Costa Rica. Microb. Risk Anal. 2023, 24, 100262. [Google Scholar] [CrossRef]
- Randazzo, W.; López-Gálvez, F.; Allende, A.; Aznar, R.; Sánchez, G. Evaluation of Viability PCR Performance for Assessing Norovirus Infectivity in Fresh-Cut Vegetables and Irrigation Water. Int. J. Food Microbiol. 2016, 229, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.; D’souza, D.H.; Davidson, P.M. Thermal inactivation of foodborne enteric viruses and their viral surrogates in foods. J. Food Prot. 2015, 78, 1597–1617. [Google Scholar] [CrossRef] [PubMed]
- Da Graça Pedrosa De Macena, L.; Simone De Oliveira Pereira, J.; Couto Da Silva, J.; Ferreira, F.C.; Gonçalves Maranhão, A.; Lanzarini, N.M.; Pereira Miagostovich, M. Environmental Microbiology-Quantification of Infectious Human Mastadenovirus in Environmental Matrices Using PMAxx-QPCR. Braz. J. Microbiol. 2022, 53, 1456–1471. [Google Scholar] [CrossRef]
- Roos, Y.H. Water and Pathogenic Viruses Inactivation-Food Engineering Perspectives. Food Eng. Rev. 2020, 12, 251–267. [Google Scholar] [CrossRef]
- Gregory, J.B.; Litaker, R.W.; Noble, R.T. Rapid One-Step Quantitative Reverse Transcriptase PCR Assay with Competitive Internal Positive Control for Detection of Enteroviruses in Environmental Samples. Appl. Environ. Microbiol. 2006, 72, 3960–3967. [Google Scholar] [CrossRef]
- Blanco, A.; Guix, S.; Fuster, N.; Fuentes, C.; Bartolomé, R.; Cornejo, T.; Pintó, R.M.; Bosch, A. Norovirus in Bottled Water Associated with Gastroenteritis Outbreak, Spain, 2016. Emerg. Infect. Dis. 2016, 23, 1531–1534. [Google Scholar] [CrossRef]
- Fuster, N.; Pint, R.M.; Fuentes, C.; Beguiristain, N.; Bosch, A.; Guix, S. Propidium Monoazide RTqPCR Assays for the Assessment of Hepatitis A Inactivation and for a Better Estimation of the Health Risk of Contaminated Waters. Water Res. 2016, 101, 226–232. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Comparison | AdV | HAV | IAV | Nov GI |
|---|---|---|---|---|
| Live 100 µM—Live 50 µM | 0.2700 | 0.4300 | 0.6800 | 0.8813 |
| Live 100 µM—HI 100 µM | 0.2500 | 0.0037 | 0.0260 | 0.0003 |
| Live 100 µM—HI 50 µM | 0.8800 | 0.3200 | 0.0820 | 0.1419 |
| Live 100 µM—HI no dye | 0.0024 | 0.0870 | 0.1700 | 0.0112 |
| Live 100 µM—Live no dye | 0.0250 | 0.0520 | 0.0160 | 0.5462 |
| Live 50 µM—HI 100 µM | 0.0140 | 0.0002 | 0.0680 | 0.0002 |
| Live 50 µM—HI 50 µM | 0.2000 | 0.0890 | 0.1700 | 0.1091 |
| Live 50 µM—HI no dye | 0.0310 | 0.0130 | 0.0770 | 0.0072 |
| Live 50 µM—Live no dye | 0.2000 | 0.2500 | 0.0049 | 0.6447 |
| HI 100 µM—HI 50 µM | 0.3300 | 0.1100 | 0.8000 | 0.0780 |
| HI 100 µM—HI no dye | <0.0001 | 0.2300 | 0.0003 | 0.2825 |
| HI 100 µM—Live no dye | 0.0003 | 0.0000 | <0.0001 | 0.0001 |
| HI 50 µM—HI no dye | 0.0013 | 0.6000 | 0.0031 | 0.4231 |
| HI 50 µM—Live no dye | 0.0160 | 0.0062 | 0.0001 | 0.0502 |
| HI no dye—Live no dye | 0.3700 | 0.0003 | 0.3000 | 0.0025 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kevill, J.L.; Farkas, K.; Ridding, N.; Woodhall, N.; Malham, S.K.; Jones, D.L. Use of Capsid Integrity-qPCR for Detecting Viral Capsid Integrity in Wastewater. Viruses 2024, 16, 40. https://doi.org/10.3390/v16010040
Kevill JL, Farkas K, Ridding N, Woodhall N, Malham SK, Jones DL. Use of Capsid Integrity-qPCR for Detecting Viral Capsid Integrity in Wastewater. Viruses. 2024; 16(1):40. https://doi.org/10.3390/v16010040
Chicago/Turabian StyleKevill, Jessica L., Kata Farkas, Nicola Ridding, Nicholas Woodhall, Shelagh K. Malham, and Davey L. Jones. 2024. "Use of Capsid Integrity-qPCR for Detecting Viral Capsid Integrity in Wastewater" Viruses 16, no. 1: 40. https://doi.org/10.3390/v16010040
APA StyleKevill, J. L., Farkas, K., Ridding, N., Woodhall, N., Malham, S. K., & Jones, D. L. (2024). Use of Capsid Integrity-qPCR for Detecting Viral Capsid Integrity in Wastewater. Viruses, 16(1), 40. https://doi.org/10.3390/v16010040