
Beyond the Syndemic of Opioid Use Disorders and HIV: The Impact of Opioids on Viral Reservoirs
 , , ,
, , ,
Abstract
1. Introduction
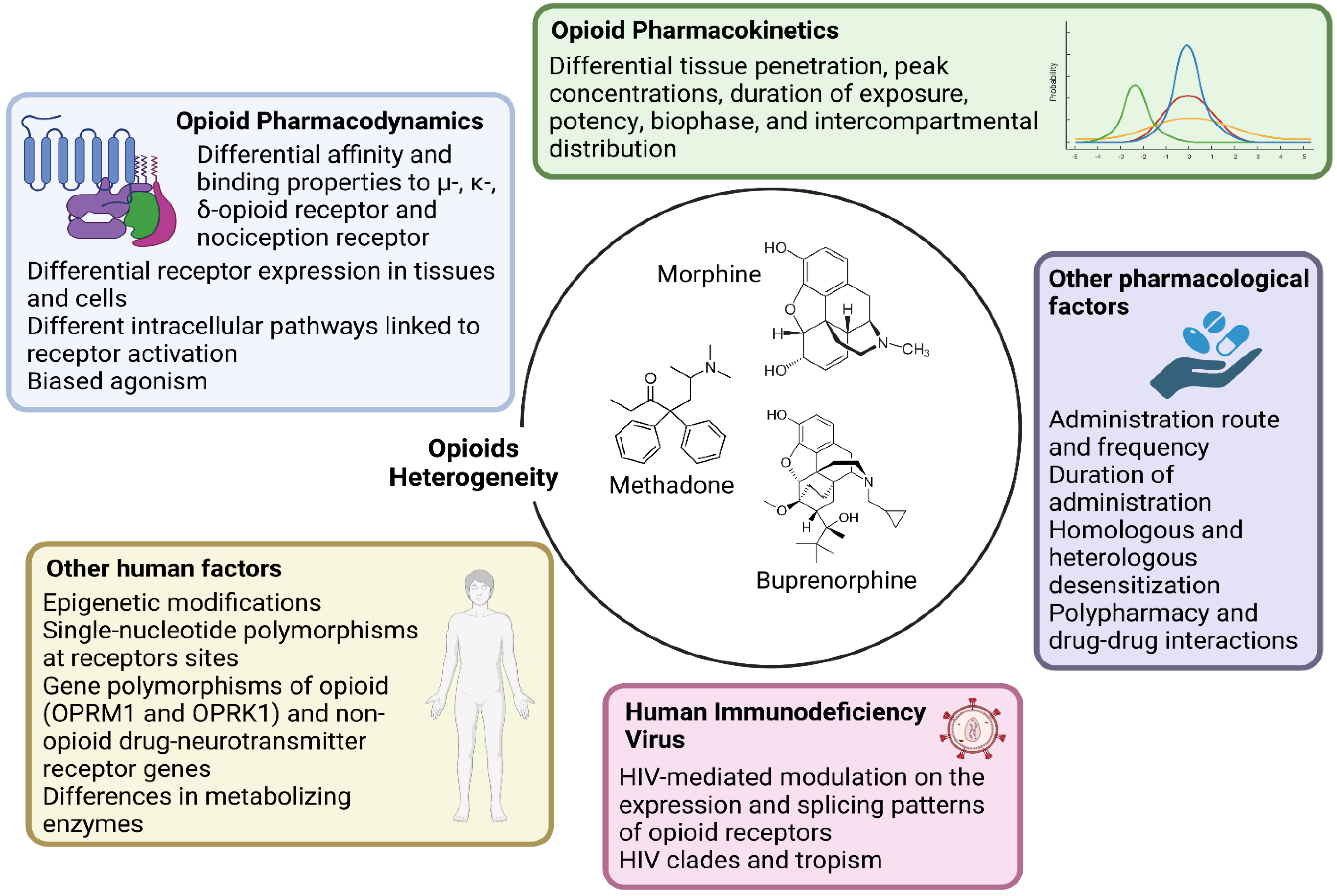
- Research should account for the heterogeneity within the large class of opioid drugs. Studies aiming to assess effects of opioids on viral and immune factors should consider the pharmacokinetic and pharmacodynamic features of each distinct opioid drug, and possibly other factors that can affect cell and tissue responses to opioids, as summarized in Figure 1. For instance, during chronic opioid exposure, compensatory mechanisms and tolerance could explain contradictory findings such as reduced B-cell responses following morphine administration (acute exposure) but effective humoral immune responses to vaccination in heroin and methadone users (chronic exposure) [11]. Differences between opioid drugs may also explain discrepancies with some, but not other opioids being associated with immunosuppression [11,17,18].
- Research should account for the heterogeneity within the cellular and tissue-based HIV reservoirs. HIV integrates its proviral genome into the host genome, establishing a pool of infected cells that can indefinitely harbor provirus during ART [13]. This cell reservoir is heterogeneous and mainly consists of memory CD4+ T-cells and T-cell subsets (e.g., follicular helper, regulatory, helper-1, and helper-17 cells) [13]. More recently, evidence has accumulated on the contribution of myeloid cells, which includes macrophages and microglia, in these HIV reservoirs [19,20]. These cell types display distinct proviral transcriptional activity and inducibility, as well as different expression and activity of opioid receptors and response patterns to opioid-induced stimuli. Further, the response to opioids on similar or even the same cell types may vary quantitatively and qualitatively across tissues, which may explain why the same dose of the same opioid could differentially impact the HIV reservoir of distinct tissues such as the gut-associated lymphoid tissue and central nervous system (CNS) [10,11,21,22].
- Research should account for the heterogeneity in the activity of HIV reservoirs. The reservoir can be distinguished based on its replication and transcriptional activity [13]. Defective provirus genomes do not encode replication-competent viruses due to the accumulation of several mutations and deletions [13,15]. However, defective proviruses can be transcribed and can contribute to HIV pathogenesis by producing viral proteins and noncoding RNA sequences [23]. Current HIV cure strategies primarily target intact provirus genomes, which account for less than 5% of all the infected cells in peripheral blood [24,25,26,27]. Since HIV transcripts and proteins can trigger immune activation, it is important to differentiate not only intact and defective provirus but also their transcriptional capacity to understand the respective contribution to HIV pathogenesis. Furthermore, transcriptionally active reservoirs could be better targeted for clearance by host immune responses [13,15,27,28]. As discussed in this review, opioids can shape the HIV reservoir primarily through immunological effects. This observation raises the possibility that the modulation of the HIV reservoir by opioids may be more significant for transcriptionally active proviruses compared to reservoirs that undergo deep latency and complete silencing. Similar hypotheses can be made with regard to the different degrees of inducibility from latency across reservoirs [13,29].
- Research should address concurrent direct and indirect biological mechanisms and confounding factors. In vitro and ex vivo studies can identify molecular pathways affected by opioids, but they usually focus on single cell types and relatively short-term exposure (hours–weeks). Translation of these findings to PWH may also vary since polysubstance use is common, and exposure to contaminants of opioid drugs may also have immunomodulatory effects. In this regard, animal and human studies allow concomitant evaluation of indirect effects mediated by immune modulation and cell-to-cell interactions while accounting for relevant moderating and confounding factors, including ART. In this regard, opioids may alter the metabolism, distribution, and elimination of ART drugs via their effects on drug metabolizing enzymes and efflux transporters [30,31], although few studies have addressed these interactions to date.
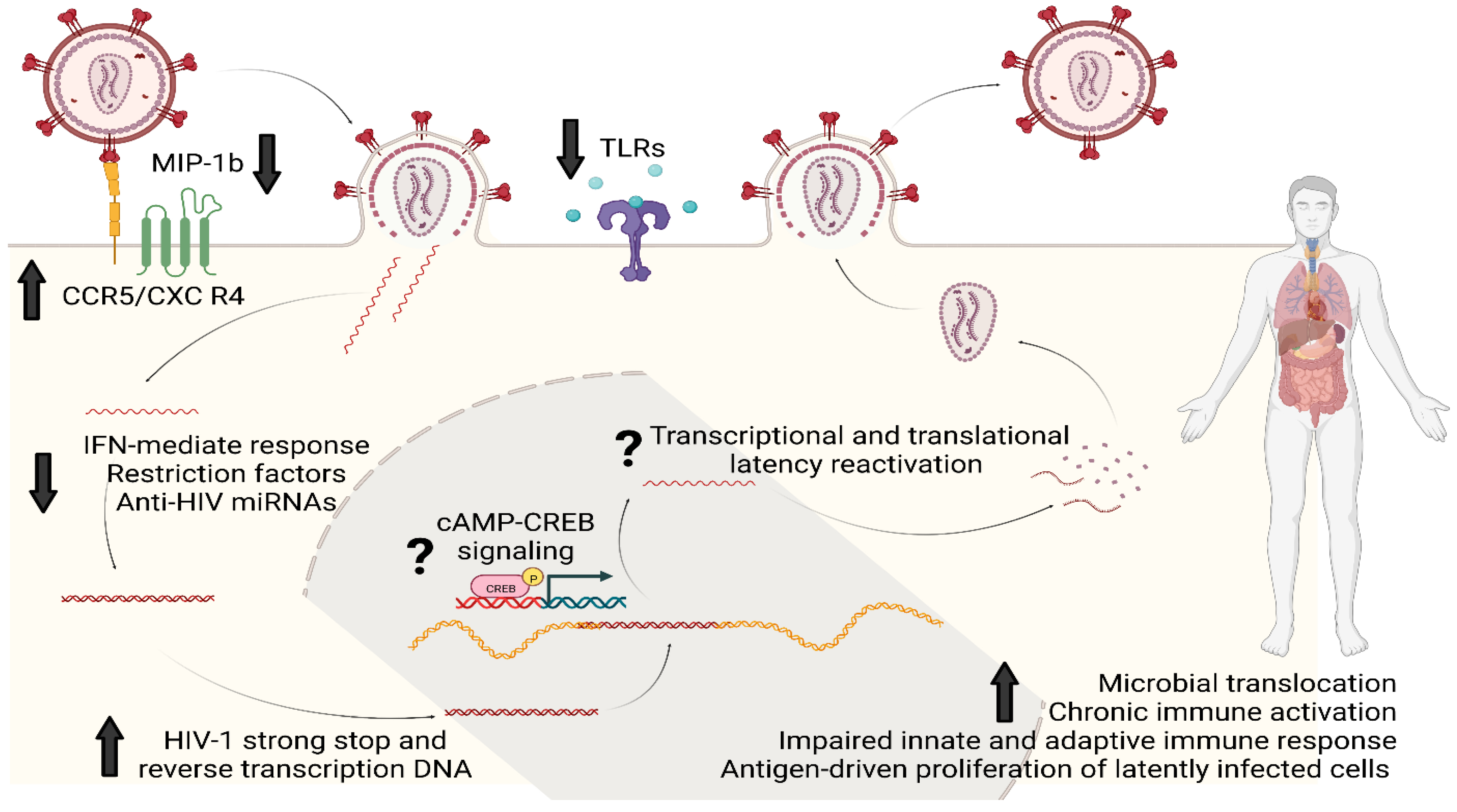
2. Opioids and HIV Infectivity and Replication
3. Opioids and HIV Latency
4. Opioids and the Immune System
5. Opioids, HIV, and the Central Nervous System
6. Future Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mattson, C.L.; Tanz, L.J.; Quinn, K.; Kariisa, M.; Patel, P.; Davis, N.L. Trends and Geographic Patterns in Drug and Synthetic Opioid Overdose Deaths—United States, 2013–2019. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 202–207. [Google Scholar] [CrossRef]
- World Drug Report. 2022. Available online: //www.unodc.org/unodc/en/data-and-analysis/world-drug-report-2022.html (accessed on 30 November 2022).
- Full Report—In Danger: UNAIDS Global AIDS Update. 2022. Available online: https://www.unaids.org/en/resources/documents/2022/in-danger-global-aids-update (accessed on 30 November 2022).
- Degenhardt, L.; Peacock, A.; Colledge, S.; Leung, J.; Grebely, J.; Vickerman, P.; Stone, J.; Cunningham, E.B.; Trickey, A.; Dumchev, K.; et al. Global prevalence of injecting drug use and sociodemographic characteristics and prevalence of HIV, HBV, and HCV in people who inject drugs: A multistage systematic review. Lancet Glob. Health 2017, 5, e1192–e1207. [Google Scholar] [CrossRef]
- Cunningham, C.O. Opioids and HIV Infection: From Pain Management to Addiction Treatment. Top. Antivir. Med. 2018, 25, 143–146. [Google Scholar]
- Flores, J.; Liang, Y.; Ketchum, N.S.; Turner, B.J.; Bullock, D.; Villarreal, R.; Potter, J.S.; Taylor, B.S. Prescription Opioid Use is Associated with Virologic Failure in People Living with HIV. AIDS Behav. 2018, 22, 1323–1328. [Google Scholar] [CrossRef]
- Canan, C.E.; Chander, G.; Monroe, A.K.; Gebo, K.A.; Moore, R.D.; Agwu, A.L.; Alexander, G.C.; Lau, B.; HIV Research Network. High-Risk Prescription Opioid Use Among People Living with HIV. J. Acquir. Immune Defic. Syndr. 2018, 78, 283–290. [Google Scholar] [CrossRef]
- McNamara, K.F.; Biondi, B.E.; Hernández-Ramírez, R.U.; Taweh, N.; Grimshaw, A.A.; Springer, S.A. A Systematic Review and Meta-Analysis of Studies Evaluating the Effect of Medication Treatment for Opioid Use Disorder on Infectious Disease Outcomes. Open Forum Infect. Dis. 2021, 8, ofab289. [Google Scholar] [CrossRef] [PubMed]
- Trickey, A.; May, M.T.; Vehreschild, J.; Obel, N.; Gill, M.J.; Crane, H.; Boesecke, C.; Samji, H.; Grabar, S.; Cazanave, C.; et al. Cause-Specific Mortality in HIV-Positive Patients Who Survived Ten Years after Starting Antiretroviral Therapy. PLoS ONE 2016, 11, e0160460. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.; Barbaro, J.; Martínez-Aguado, P.; Chilunda, V.; Jaureguiberry-Bravo, M.; Berman, J.W. The Effects of Opioids on HIV Neuropathogenesis. Front. Immunol. 2019, 10, 2445. [Google Scholar] [CrossRef]
- Azzoni, L.; Metzger, D.; Montaner, L.J. Effect of Opioid Use on Immune Activation and HIV Persistence on ART. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2020, 15, 643–657. [Google Scholar] [CrossRef]
- Jaureguiberry-Bravo, M.; Lopez, L.; Berman, J.W. Frontline Science: Buprenorphine decreases CCL2-mediated migration of CD14+ CD16+ monocytes. J. Leukoc. Biol. 2018, 104, 1049–1059. [Google Scholar] [CrossRef] [PubMed]
- Cohn, L.B.; Chomont, N.; Deeks, S.G. The Biology of the HIV-1 Latent Reservoir and Implications for Cure Strategies. Cell Host Microbe 2020, 27, 519–530. [Google Scholar] [CrossRef]
- Henderson, L.J.; Reoma, L.B.; Kovacs, J.A.; Nath, A. Advances toward Curing HIV-1 Infection in Tissue Reservoirs. J. Virol. 2020, 94, e00375-19. [Google Scholar] [CrossRef]
- Kreider, E.F.; Bar, K.J. HIV-1 Reservoir Persistence and Decay: Implications for Cure Strategies. Curr. HIV/AIDS Rep. 2022, 19, 194–206. [Google Scholar] [CrossRef]
- Dufour, C.; Gantner, P.; Fromentin, R.; Chomont, N. The multifaceted nature of HIV latency. J. Clin. Investig. 2020, 130, 3381–3390. [Google Scholar] [CrossRef] [PubMed]
- Gornalusse, G.G.; Vojtech, L.N.; Levy, C.N.; Hughes, S.M.; Kim, Y.; Valdez, R.; Pandey, U.; Ochsenbauer, C.; Astronomo, R.; McElrath, J.; et al. Buprenorphine Increases HIV-1 Infection In Vitro but Does Not Reactivate HIV-1 from Latency. Viruses 2021, 13, 1472. [Google Scholar] [CrossRef]
- Sacerdote, P.; Manfredi, B.; Mantegazza, P.; Panerai, A.E. Antinociceptive and immunosuppressive effects of opiate drugs: A structure-related activity study. Br. J. Pharmacol. 1997, 121, 834–840. [Google Scholar] [CrossRef]
- Wahl, A.; Al-Harthi, L. HIV infection of non-classical cells in the brain. Retrovirology 2023, 20, 1. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chaillon, A.; Gianella, S.; Wong, L.M.; Li, D.; Simermeyer, T.L.; Porrachia, M.; Ignacio, C.; Woodworth, B.; Zhong, D.; et al. Brain microglia serve as a persistent HIV reservoir despite durable antiretroviral therapy. J. Clin. Investig. 2023, 133, e167417. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Jiang, Y.; Liang, S.; Cheng, Z.; Zhu, X.; Guo, Q. Opioids Regulate the Immune System: Focusing on Macrophages and Their Organelles. Front. Pharmacol. 2021, 12, 814241. [Google Scholar] [CrossRef]
- Acharya, A.; Olwenyi, O.A.; Thurman, M.; Pandey, K.; Morsey, B.M.; Lamberty, B.; Ferguson, N.; Callen, S.; Fang, Q.; Buch, S.J.; et al. Chronic morphine administration differentially modulates viral reservoirs in SIVmac251 infected rhesus macaque model. J. Virol. 2020, 95, e01657-20. [Google Scholar] [CrossRef]
- Imamichi, H.; Smith, M.; Adelsberger, J.W.; Izumi, T.; Scrimieri, F.; Sherman, B.T.; Rehm, C.A.; Imamichi, T.; Pau, A.; Catalfamo, M.; et al. Defective HIV-1 proviruses produce viral proteins. Proc. Natl. Acad. Sci. USA 2020, 117, 3704–3710. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, F.R.; White, J.A.; Tumiotto, C.; Ritter, K.D.; Cai, M.; Gandhi, R.T.; Deeks, S.G.; Howell, B.J.; Montaner, L.J.; Blankson, J.N.; et al. Intact proviral DNA assay analysis of large cohorts of people with HIV provides a benchmark for the frequency and composition of persistent proviral DNA. Proc. Natl. Acad. Sci. USA 2020, 117, 18692–18700. [Google Scholar] [CrossRef] [PubMed]
- White, J.A.; Simonetti, F.R.; Beg, S.; McMyn, N.F.; Dai, W.; Bachmann, N.; Lai, J.; Ford, W.C.; Bunch, C.; Jones, J.L.; et al. Complex decay dynamics of HIV virions, intact and defective proviruses, and 2LTR circles following initiation of antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2022, 119, e2120326119. [Google Scholar] [CrossRef]
- Bruner, K.M.; Wang, Z.; Simonetti, F.R.; Bender, A.M.; Kwon, K.J.; Sengupta, S.; Fray, E.J.; Beg, S.A.; Antar, A.A.R.; Jenike, K.M.; et al. A quantitative approach for measuring the reservoir of latent HIV-1 proviruses. Nature 2019, 566, 120–125. [Google Scholar] [CrossRef]
- Gandhi, R.T.; Cyktor, J.C.; Bosch, R.J.; Mar, H.; Laird, G.M.; Martin, A.; Collier, A.C.; Riddler, S.A.; Macatangay, B.J.; Rinaldo, C.R.; et al. Selective Decay of Intact HIV-1 Proviral DNA on Antiretroviral Therapy. J. Infect. Dis. 2020, 223, 225–233. [Google Scholar] [CrossRef]
- Peluso, M.J.; Bacchetti, P.; Ritter, K.D.; Beg, S.; Lai, J.; Martin, J.N.; Hunt, P.W.; Henrich, T.J.; Siliciano, J.D.; Siliciano, R.F.; et al. Differential Decay of Intact and Defective Proviral DNA in HIV-1–Infected Individuals on Suppressive Antiretroviral Therapy. JCI Insight 2020, 5, e132997. [Google Scholar] [CrossRef]
- Dhummakupt, A.; Rubens, J.H.; Anderson, T.; Powell, L.; Nonyane, B.A.S.; Siems, L.V.; Collinson-Streng, A.; Nilles, T.; Jones, R.B.; Tepper, V.; et al. Differences in inducibility of the latent HIV reservoir in perinatal and adult infection. JCI Insight 2020, 5, e134105. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Kwatra, D.; Minocha, M.; Paturi, D.K.; Budda, B.; Mitra, A.K. Efflux transporters- and cytochrome P-450-mediated interactions between drugs of abuse and antiretrovirals. Life Sci. 2011, 88, 959–971. [Google Scholar] [CrossRef]
- Eap, C.B.; Buclin, T.; Baumann, P. Interindividual variability of the clinical pharmacokinetics of methadone: Implications for the treatment of opioid dependence. Clin. Pharmacokinet. 2002, 41, 1153–1193. [Google Scholar] [CrossRef]
- Madhuravasal Krishnan, J.; Kong, L.; Karns, R.; Medvedovic, M.; Sherman, K.E.; Blackard, J.T. The Synthetic Opioid Fentanyl Increases HIV Replication and Chemokine Co-Receptor Expression in Lymphocyte Cell Lines. Viruses 2023, 15, 1027. [Google Scholar] [CrossRef]
- Kong, L.; Shata, M.T.M.; Brown, J.L.; Lyons, M.S.; Sherman, K.E.; Blackard, J.T. The synthetic opioid fentanyl increases HIV replication and chemokine co-receptor expression in vitro. J. Neurovirol. 2022, 28, 583–594. [Google Scholar] [CrossRef]
- Wang, M.-R.; Wu, D.-D.; Luo, F.; Zhong, C.-J.; Wang, X.; Zhu, N.; Wu, Y.-J.; Hu, H.-T.; Feng, Y.; Wang, X.; et al. Methadone Inhibits Viral Restriction Factors and Facilitates HIV Infection in Macrophages. Front. Immunol. 2020, 11, 1253. [Google Scholar] [CrossRef]
- Karagiannis, T.T.; Cleary, J.P.; Gok, B.; Henderson, A.J.; Martin, N.G.; Yajima, M.; Nelson, E.C.; Cheng, C.S. Single cell transcriptomics reveals opioid usage evokes widespread suppression of antiviral gene program. Nat. Commun. 2020, 11, 2611. [Google Scholar] [CrossRef]
- Ma, H.; Wang, H.; Li, M.; Barreto-de-Souza, V.; Reinecke, B.A.; Gunta, R.; Zheng, Y.; Kang, G.; Nassehi, N.; Zhang, H.; et al. Bivalent Ligand Aiming Putative Mu Opioid Receptor and Chemokine Receptor CXCR4 Dimers in Opioid Enhanced HIV-1 Entry. ACS Med. Chem. Lett. 2020, 11, 2318–2324. [Google Scholar] [CrossRef]
- Chao, C.C.; Gekker, G.; Sheng, W.S.; Hu, S.; Peterson, P.K. U50488 inhibits HIV-1 expression in acutely infected monocyte-derived macrophages. Drug Alcohol Depend. 2001, 62, 149–154. [Google Scholar] [CrossRef]
- Swaminathan, G.; Rossi, F.; Sierra, L.-J.; Gupta, A.; Navas-Martín, S.; Martín-García, J. A role for microRNA-155 modulation in the anti-HIV-1 effects of Toll-like receptor 3 stimulation in macrophages. PLoS Pathog. 2012, 8, e1002937. [Google Scholar] [CrossRef] [PubMed]
- Pilakka-Kanthikeel, S.; Nair, M.P.N. Interaction of drugs of abuse and microRNA with HIV: A brief review. Front. Microbiol. 2015, 6, 967. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Jiang, J.; Liang, B.; Wei, F.; Huang, J.; Pan, P.; Su, J.; Zhou, B.; Zang, N.; Ye, L.; et al. Opiate use inhibits TLR9 signaling pathway in vivo: Possible role in pathogenesis of HIV-1 infection. Sci. Rep. 2017, 7, 13071. [Google Scholar] [CrossRef]
- Vibholm, L.K.; Konrad, C.V.; Schleimann, M.H.; Frattari, G.; Winckelmann, A.; Klastrup, V.; Jensen, N.M.; Jensen, S.S.; Schmidt, M.; Wittig, B.; et al. Effects of 24-week Toll-like receptor 9 agonist treatment in HIV type 1+ individuals. AIDS 2019, 33, 1315–1325. [Google Scholar] [CrossRef]
- Arnatt, C.K.; Falls, B.A.; Yuan, Y.; Raborg, T.J.; Masvekar, R.R.; El-Hage, N.; Selley, D.E.; Nicola, A.V.; Knapp, P.E.; Hauser, K.F.; et al. Exploration of bivalent ligands targeting putative mu opioid receptor and chemokine receptor CCR5 dimerization. Bioorg. Med. Chem. 2016, 24, 5969–5987. [Google Scholar] [CrossRef] [PubMed]
- Szabo, I.; Wetzel, M.A.; Zhang, N.; Steele, A.D.; Kaminsky, D.E.; Chen, C.; Liu-Chen, L.-Y.; Bednar, F.; Henderson, E.E.; Howard, O.M.Z.; et al. Selective inactivation of CCR5 and decreased infectivity of R5 HIV-1 strains mediated by opioid-induced heterologous desensitization. J. Leukoc. Biol. 2003, 74, 1074–1082. [Google Scholar] [CrossRef]
- Huang, J.; Wang, F.; Argyris, E.; Chen, K.; Liang, Z.; Tian, H.; Huang, W.; Squires, K.; Verlinghieri, G.; Zhang, H. Cellular microRNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat. Med. 2007, 13, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, N.; von Siebenthal, C.; Vongrad, V.; Turk, T.; Neumann, K.; Beerenwinkel, N.; Bogojeska, J.; Fellay, J.; Roth, V.; Kok, Y.L.; et al. Determinants of HIV-1 reservoir size and long-term dynamics during suppressive ART. Nat. Commun. 2019, 10, 3193. [Google Scholar] [CrossRef]
- Gianella, S.; Rawlings, S.A.; Dobrowolski, C.; Nakazawa, M.; Chaillon, A.; Strain, M.; Layman, L.; Caballero, G.; Scully, E.; Scott, B.; et al. Sex Differences in Human Immunodeficiency Virus Persistence and Reservoir Size During Aging. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2022, 75, 73–80. [Google Scholar] [CrossRef]
- Kirk, G.D.; Astemborski, J.; Mehta, S.H.; Ritter, K.D.; Laird, G.M.; Bordi, R.; Sekaly, R.; Siliciano, J.D.; Siliciano, R.F. Nonstructured Treatment Interruptions Are Associated with Higher Human Immunodeficiency Virus Reservoir Size Measured by Intact Proviral DNA Assay in People Who Inject Drugs. J. Infect. Dis. 2020, 223, 1905–1913. [Google Scholar] [CrossRef]
- Prottengeier, J.; Koutsilieri, E.; Scheller, C. The effects of opioids on HIV reactivation in latently-infected T-lymphoblasts. AIDS Res. Ther. 2014, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ye, L.; Zhou, Y.; Liu, M.-Q.; Zhou, D.-J.; Ho, W.-Z. Inhibition of anti-HIV microRNA expression: A mechanism for opioid-mediated enhancement of HIV infection of monocytes. Am. J. Pathol. 2011, 178, 41–47. [Google Scholar] [CrossRef]
- Peterson, P.K.; Gekker, G.; Hu, S.; Anderson, W.R.; Kravitz, F.; Portoghese, P.S.; Balfour, H.H.; Chao, C.C. Morphine amplifies HIV-1 expression in chronically infected promonocytes cocultured with human brain cells. J. Neuroimmunol. 1994, 50, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Zou, Z.; Li, X.; Cheng, N.; Guo, N.; Sun, G.; Liu, D. A new side-effect of sufentanil: Increased monocyte-endothelial adhesion. BMC Anesthesiol. 2021, 21, 267. [Google Scholar] [CrossRef]
- Basukala, B.; Rossi, S.; Bendiks, S.; Gnatienko, N.; Patts, G.; Krupitsky, E.; Lioznov, D.; So-Armah, K.; Sagar, M.; Cheng, C.; et al. Virally Suppressed People Living with HIV Who Use Opioids Have Diminished Latency Reversal. Viruses 2023, 15, 415. [Google Scholar] [CrossRef]
- Börner, C.; Lanciotti, S.; Koch, T.; Höllt, V.; Kraus, J. μ opioid receptor agonist-selective regulation of interleukin-4 in T lymphocytes. J. Neuroimmunol. 2013, 263, 35–42. [Google Scholar] [CrossRef]
- Kafami, L.; Etesami, I.; Felfeli, M.; Enayati, N.; Ghiaghi, R.; Aminian, A.; Dehpour, A. Methadone diminishes neuroinflammation and disease severity in EAE through modulating T cell function. J. Neuroimmunol. 2013, 255, 39–44. [Google Scholar] [CrossRef]
- Peterson, P.K.; Sharp, B.; Gekker, G.; Brummitt, C.; Keane, W.F. Opioid-mediated suppression of interferon-gamma production by cultured peripheral blood mononuclear cells. J. Clin. Investig. 1987, 80, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Kholodnaia, A.; So-Armah, K.; Cheng, D.; Gnatienko, N.; Patts, G.; Samet, J.H.; Freiberg, M.; Lioznov, D. Impact of illicit opioid use on markers of monocyte activation and systemic inflammation in people living with HIV. PLoS ONE 2022, 17, e0265504. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.L.; Nguyen, T.; Uebelhoer, L.S.; Kunkel, L.E.; Korthuis, P.T.; Lancioni, C.L. Altered monocyte phenotype and dysregulated innate cytokine responses among people living with HIV and opioid-use disorder. AIDS 2020, 34, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Sindberg, G.M.; Roy, S. Disruption of gut homeostasis by opioids accelerates HIV disease progression. Front. Microbiol. 2015, 6, 643. [Google Scholar] [CrossRef] [PubMed]
- Hileman, C.O.; Bowman, E.R.; Gabriel, J.; Kettelhut, A.; Labbato, D.; Smith, C.; Avery, A.; Parran, T.; Funderburg, N.; McComsey, G.A. Impact of Heroin and HIV on Gut Integrity and Immune Activation. J. Acquir. Immune Defic. Syndr. 2022, 89, 519–526. [Google Scholar] [CrossRef]
- Meng, J.; Yu, H.; Ma, J.; Wang, J.; Banerjee, S.; Charboneau, R.; Barke, R.A.; Roy, S. Morphine induces bacterial translocation in mice by compromising intestinal barrier function in a TLR-dependent manner. PLoS ONE 2013, 8, e54040. [Google Scholar] [CrossRef]
- Müller-Lissner, S.; Bassotti, G.; Coffin, B.; Drewes, A.M.; Breivik, H.; Eisenberg, E.; Emmanuel, A.; Laroche, F.; Meissner, W.; Morlion, B. Opioid-Induced Constipation and Bowel Dysfunction: A Clinical Guideline. Pain Med. Malden Mass 2017, 18, 1837–1863. [Google Scholar] [CrossRef]
- Herlihy, B.; Roy, S. Gut-Microbiome Implications in Opioid Use Disorder and Related Behaviors. Adv. Drug Alcohol Res. 2022, 2, 10311. [Google Scholar] [CrossRef]
- Le Bastard, Q.; Al-Ghalith, G.A.; Grégoire, M.; Chapelet, G.; Javaudin, F.; Dailly, E.; Batard, E.; Knights, D.; Montassier, E. Systematic review: Human gut dysbiosis induced by non-antibiotic prescription medications. Aliment. Pharmacol. Ther. 2018, 47, 332–345. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Sindberg, G.; Wang, F.; Meng, J.; Sharma, U.; Zhang, L.; Dauer, P.; Chen, C.; Dalluge, J.; Johnson, T.; et al. Opioid-induced gut microbial disruption and bile dysregulation leads to gut barrier compromise and sustained systemic inflammation. Mucosal Immunol. 2016, 9, 1418–1428. [Google Scholar] [CrossRef]
- Sindberg, G.M.; Sharma, U.; Banerjee, S.; Anand, V.; Dutta, R.; Gu, C.-J.; Volsky, D.J.; Roy, S. An infectious murine model for studying the systemic effects of opioids on early HIV pathogenesis in the gut. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2015, 10, 74–87. [Google Scholar] [CrossRef]
- Abdel-Mohsen, M.; Kuri-Cervantes, L.; Grau-Exposito, J.; Spivak, A.M.; Nell, R.A.; Tomescu, C.; Vadrevu, S.K.; Giron, L.B.; Serra-Peinado, C.; Genescà, M.; et al. CD32 is expressed on cells with transcriptionally active HIV but does not enrich for HIV DNA in resting T cells. Sci. Transl. Med. 2018, 10, eaar6759. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Y.; Meng, J.; Girotra, M.; Ramakrishnan, S.; Roy, S. Immune modulation mediated by extracellular vesicles of intestinal organoids is disrupted by opioids. Mucosal Immunol. 2021, 14, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.; Olson, R.K.; Erhart, F.N.; Zhang, L.; Meng, J.; Segura, B.; Banerjee, S.; Sharma, M.; Saluja, A.K.; Ramakrishnan, S.; et al. Prescription Opioids induce Gut Dysbiosis and Exacerbate Colitis in a Murine Model of Inflammatory Bowel Disease. J. Crohns Colitis 2020, 14, 801–817. [Google Scholar] [CrossRef] [PubMed]
- Abdel Shaheed, C.; Beardsley, J.; Day, R.O.; McLachlan, A.J. Immunomodulatory effects of pharmaceutical opioids and antipyretic analgesics: Mechanisms and relevance to infection. Br. J. Clin. Pharmacol. 2022, 88, 3114–3131. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, M.; Chen, C.; Liu, L.; Wei, X.; Zeng, S. Toll-Like Receptor 4 (TLR4)/Opioid Receptor Pathway Crosstalk and Impact on Opioid Analgesia, Immune Function, and Gastrointestinal Motility. Front. Immunol. 2020, 11, 1455. [Google Scholar] [CrossRef]
- Chaillon, A.; Gianella, S.; Dellicour, S.; Rawlings, S.A.; Schlub, T.E.; Oliveira, M.F.D.; Ignacio, C.; Porrachia, M.; Vrancken, B.; Smith, D.M. HIV persists throughout deep tissues with repopulation from multiple anatomical sources. J. Clin. Investig. 2020, 130, 1699–1712. [Google Scholar] [CrossRef]
- Fitting, S.; McRae, M.; Hauser, K.F. Opioid and neuroHIV Comorbidity—Current and Future Perspectives. J. Neuroimmune Pharmacol. Off. J. Soc. NeuroImmune Pharmacol. 2020, 15, 584–627. [Google Scholar] [CrossRef]
- Jaureguiberry-Bravo, M.; Kelschenbach, J.; Murphy, A.; Carvallo, L.; Hadas, E.; Tesfa, L.; Scott, T.M.; Rivera-Mindt, M.; Cunningham, C.O.; Arnsten, J.H.; et al. Treatment with buprenorphine prior to EcoHIV infection of mice prevents the development of neurocognitive impairment. J. Leukoc. Biol. 2021, 109, 675–681. [Google Scholar] [CrossRef]
- Patel, S.H.; Ismaiel, O.A.; Mylott, W.R.; Yuan, M.; McClay, J.L.; Paris, J.J.; Hauser, K.F.; McRae, M. Cell-type specific differences in antiretroviral penetration and the effects of HIV-1 Tat and morphine among primary human brain endothelial cells, astrocytes, pericytes, and microglia. Neurosci. Lett. 2019, 712, 134475. [Google Scholar] [CrossRef]
- Fox, H.S.; Niu, M.; Morsey, B.M.; Lamberty, B.G.; Emanuel, K.; Periyasamy, P.; Callen, S.; Acharya, A.; Kubik, G.; Eudy, J.; et al. Morphine suppresses peripheral responses and transforms brain myeloid gene expression to favor neuropathogenesis in SIV infection. Front. Immunol. 2022, 13, 1012884. [Google Scholar] [CrossRef]
- Leibrand, C.R.; Paris, J.J.; Jones, A.M.; Ohene-Nyako, M.; Rademeyer, K.M.; Nass, S.R.; Kim, W.-K.; Knapp, P.E.; Hauser, K.F.; McRae, M. Independent actions by HIV-1 Tat and morphine to increase recruitment of monocyte-derived macrophages into the brain in a region-specific manner. Neurosci. Lett. 2022, 788, 136852. [Google Scholar] [CrossRef] [PubMed]
- Javanbakht, M.; Khan, L.; Mustanski, B.; Shoptaw, S.; Baum, M.K.; Mehta, S.H.; Kirk, G.D.; Lai, S.; Moore, R.; Milloy, M.-J.; et al. Substance use and other factors associated with COVID-19 vaccine uptake among people at risk for or living with HIV: Findings from the C3PNO consortium. Prev. Med. Rep. 2023, 35, 102300. [Google Scholar] [CrossRef]
- Sun, J.; Mehta, S.H.; Astemborski, J.; Piggott, D.A.; Genberg, B.L.; Woodson-Adu, T.; Benson, E.-M.; Thomas, D.L.; Celentano, D.D.; Vlahov, D.; et al. Mortality among people who inject drugs: A prospective cohort followed over three decades in Baltimore, MD, USA. Addict. Abingdon Engl. 2022, 117, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Mustanski, B.; Swann, G.; Newcomb, M.E.; Prachand, N. Effects of parental monitoring and knowledge on substance use and HIV risk behaviors among young men who have sex with men: Results from three studies. AIDS Behav. 2017, 21, 2046–2058. [Google Scholar] [CrossRef] [PubMed]
- Schultheis, A.; Sanchez, M.; Pedersen, S.; Kyriakides, T.; Ho, Y.-C.; Kluger, Y.; Springer, S.A. Design and implementation of a cohort study of persons living with HIV infection who are initiating medication treatment for opioid use disorder to evaluate HIV-1 persistence. Contemp. Clin. Trials Commun. 2021, 24, 100866. [Google Scholar] [CrossRef]
- Riggs, P.K.; Chaillon, A.; Jiang, G.; Letendre, S.L.; Tang, Y.; Taylor, J.; Kaytes, A.; Smith, D.M.; Dubé, K.; Gianella, S. Lessons for Understanding Central Nervous System HIV Reservoirs from the Last Gift Program. Curr. HIV/AIDS Rep. 2022, 19, 566–579. [Google Scholar] [CrossRef]
- Gianella, S.; Taylor, J.; Brown, T.R.; Kaytes, A.; Achim, C.L.; Moore, D.J.; Little, S.J.; Ellis, R.J.; Smith, D.M. Can research at the end of life be a useful tool to advance HIV cure? AIDS 2017, 31, 1–4. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Research Question | Clinical and Experimental Implications |
|---|---|
| Which are the exact regulatory mechanisms in promoting reservoir establishment, maintenance, residual transcription, activity, and reactivation and how does each opioid affect these mechanisms according to dose and in vivo concentrations, route of administration, frequency, and duration of exposure? |
|
| What are the differences among the available options of medications for opioid use disorder and among the analgesic opioid alternatives in terms of modulation of pro-viral landscape and the immune system? | |
| Is there any interaction between opioids, antiretroviral regimens and other concurrent medications/substances in modulating HIV reservoir and the immune system? | |
| Does HIV itself modulate host tissues and cell sensitivity and responses to opioids? | |
| What are the differences in reservoir establishment, maintenance, and reactivation among cell- and tissue-based HIV reservoirs and how does each opioid affect these mechanisms in each cell and body compartment? |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trunfio, M.; Chaillon, A.; Beliakova-Bethell, N.; Deiss, R.; Letendre, S.L.; Riggs, P.K.; Higgins, N.; Gianella, S. Beyond the Syndemic of Opioid Use Disorders and HIV: The Impact of Opioids on Viral Reservoirs. Viruses 2023, 15, 1712. https://doi.org/10.3390/v15081712
Trunfio M, Chaillon A, Beliakova-Bethell N, Deiss R, Letendre SL, Riggs PK, Higgins N, Gianella S. Beyond the Syndemic of Opioid Use Disorders and HIV: The Impact of Opioids on Viral Reservoirs. Viruses. 2023; 15(8):1712. https://doi.org/10.3390/v15081712
Chicago/Turabian StyleTrunfio, Mattia, Antoine Chaillon, Nadejda Beliakova-Bethell, Robert Deiss, Scott L. Letendre, Patricia K. Riggs, Niamh Higgins, and Sara Gianella. 2023. "Beyond the Syndemic of Opioid Use Disorders and HIV: The Impact of Opioids on Viral Reservoirs" Viruses 15, no. 8: 1712. https://doi.org/10.3390/v15081712
APA StyleTrunfio, M., Chaillon, A., Beliakova-Bethell, N., Deiss, R., Letendre, S. L., Riggs, P. K., Higgins, N., & Gianella, S. (2023). Beyond the Syndemic of Opioid Use Disorders and HIV: The Impact of Opioids on Viral Reservoirs. Viruses, 15(8), 1712. https://doi.org/10.3390/v15081712