

Low Pathogenic Avian Influenza H9N2 Viruses in Morocco: Antigenic and Molecular Evolution from 2021 to 2023
 , , , ,
, , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Virus Extraction and Real-Time RT-PCR for H9 Detection
2.3. Virus Isolation and cDNA Purification
2.4. Sequencing and Phylogenetic Analysis
3. Results
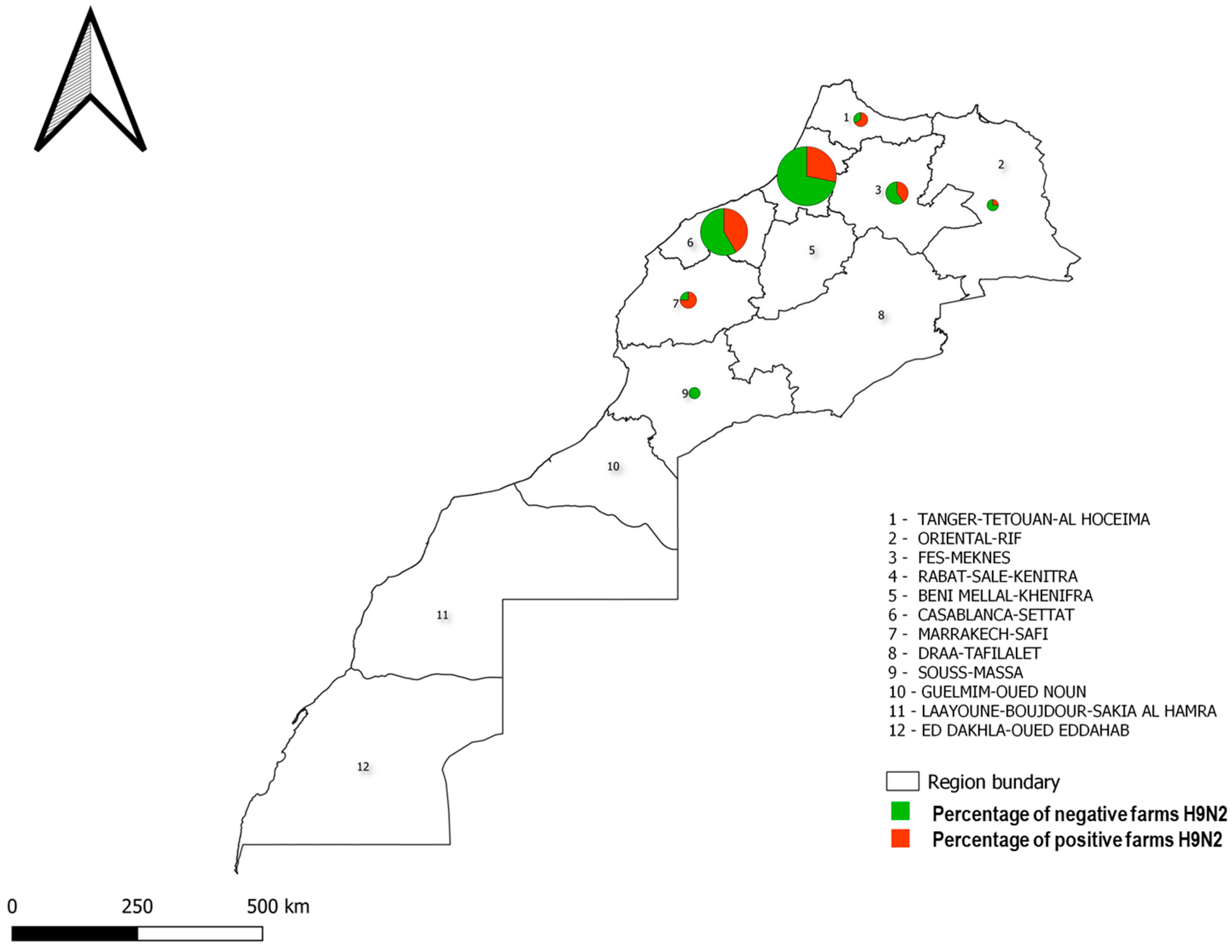
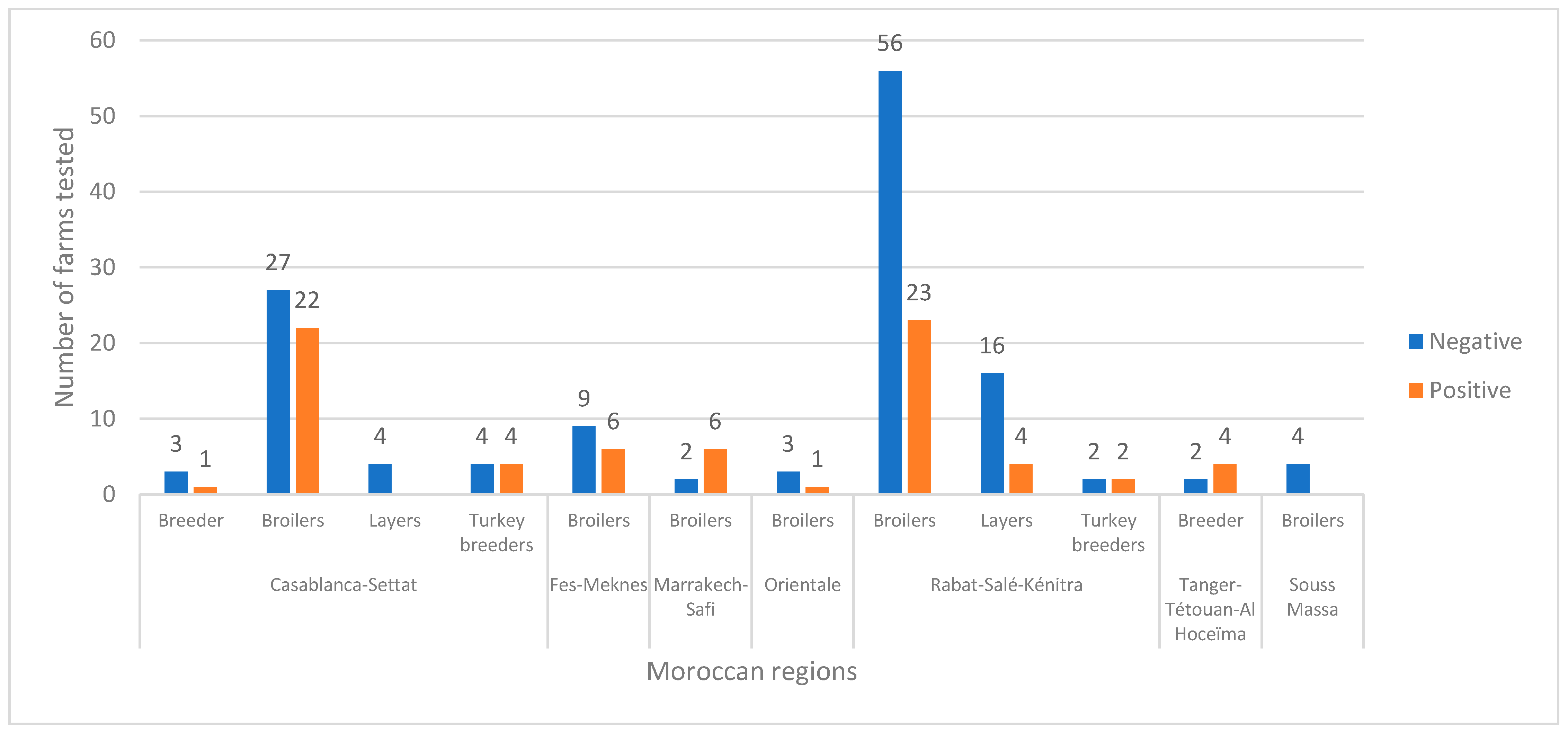
3.1. Epidemiological Survey and Real-Time RT-PCR H9N2 Results
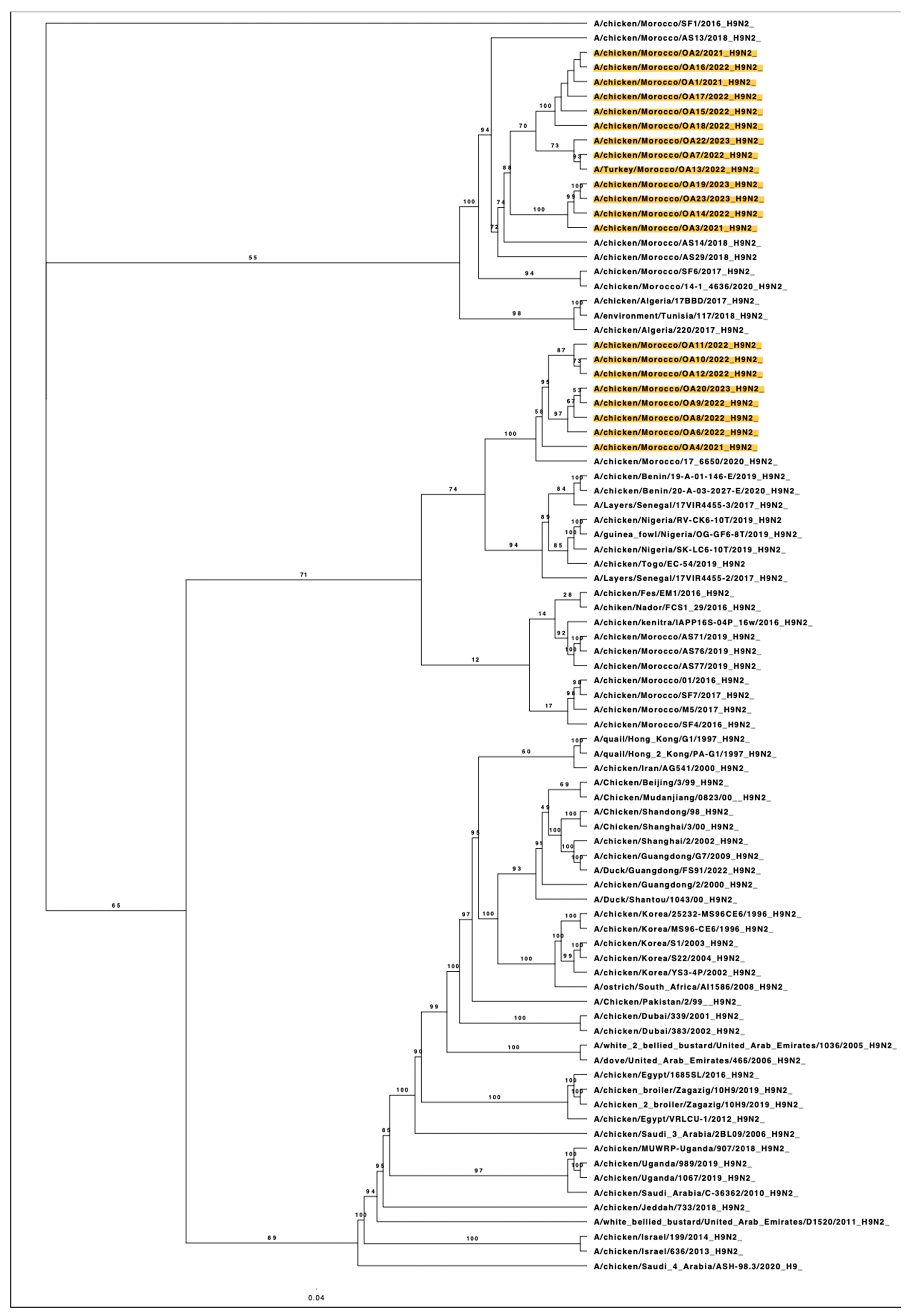
3.2. Phylogenetic Analysis of HA Gene Segment
3.3. Genetic Diversity
3.4. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Current Moroccan Isolates | Represented H9N2 Group | ||||||
|---|---|---|---|---|---|---|---|
| a Morocco/2016 | b Morocco/2017 | c Morocco/2018 | d Nigeria/2019 | e Benin/2019 | f Algeria/2017 | g Tunisia/2018 | |
| A/chicken/Morocco/OA1/2021 | 97.18 | 97.02 | 97.03 | 95.81 | 96.24 | 97.06 | 97.12 |
| A/chicken/Morocco/OA2/2021 | 97.18 | 97.02 | 97.03 | 95.81 | 96.24 | 97.06 | 97.12 |
| A/chicken/Morocco/OA3/2021 | 96.26 | 96.11 | 97.36 | 95.17 | 95.33 | 96.10 | 96.18 |
| A/chicken/Morocco/OA4/2021 | 97.61 | 97.44 | 96.08 | 96.76 | 96.59 | 97.50 | 97.56 |
| A/chicken/Morocco/OA6/2022 | 96.33 | 96.16 | 94.31 | 95.83 | 95.33 | 96.33 | 96.25 |
| A/chicken/Morocco/OA7/2022 | 96.57 | 96.51 | 97.35 | 95.10 | 95.63 | 96.59 | 96.50 |
| A/chicken/Morocco/OA8/2022 | 97.18 | 97.05 | 95.45 | 96.16 | 96.39 | 97.06 | 97.12 |
| A/chicken/Morocco/OA9/2022 | 97.04 | 96.88 | 95.32 | 96.57 | 96.10 | 96.91 | 96.97 |
| A/chicken/Morocco/OA10/2022 | 97.33 | 97.15 | 95.73 | 97.15 | 96.44 | 97.33 | 97.33 |
| A/chicken/Morocco/OA11/2022 | 97.27 | 97.27 | 97.09 | 97.09 | 96.90 | 97.27 | 97.27 |
| A/chicken/Morocco/OA12/2022 | 97.33 | 97.15 | 96.09 | 96.80 | 96.98 | 97.33 | 97.33 |
| A/Turkey/Morocco/OA13/2022 | 97.00 | 97.17 | 97.5 | 95.71 | 96.17 | 97.0 | 97.0 |
| A/chicken/Morocco/OA14/2022 | 96.72 | 96.56 | 97.82 | 95.62 | 95.78 | 96.58 | 96.65 |
| A/chicken/Morocco/OA15/2022 | 97.18 | 97.02 | 97.03 | 95.81 | 96.39 | 97.06 | 97.12 |
| A/chicken/Morocco/OA16/2022 | 97.18 | 97.02 | 97.03 | 95.81 | 96.24 | 97.06 | 97.12 |
| A/chicken/Morocco/OA17/2022 | 97.18 | 97.02 | 96.55 | 95.81 | 96.24 | 97.06 | 97.12 |
| A/chicken/Morocco/OA18/2022 | 97.18 | 97.02 | 96.55 | 95.81 | 96.08 | 97.06 | 97.12 |
| A/chicken/Morocco/OA19/2023 | 96.55 | 96.39 | 97.87 | 95.4 | 95.37 | 96.40 | 96.48 |
| A/chicken/Morocco/OA20/2023 | 95.37 | 95.12 | 94.81 | 95.37 | 95.12 | 95.37 | 95.37 |
| A/chicken/Morocco/OA22/2023 | 97.87 | 97.7 | 98.20 | 96.72 | 96.89 | 97.95 | 97.82 |
| A/chicken/Morocco/OA23/2023 | 96.39 | 96.22 | 97.70 | 95.24 | 95.20 | 96.23 | 96.31 |

References
- Peiris, M.; Yuen, K.Y.; Leung, C.W.; Chan, K.H.; Ip, P.L.S.; Lai, R.W.M.; Orr, W.K.; Shortridge, K.F. Human Infection with Influenza H9N2. Lancet 1999, 354, 916–917. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.P.; Sealy, J.E.; Harvey, W.T.; Benton, D.J.; Reeve, R.; Iqbal, M. Genetic Determinants of Receptor-Binding Preference and Zoonotic Potential of H9N2 Avian Influenza Viruses. J. Virol. 2020, 95, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Calvo, P.A.; Malide, D.; Gibbs, J.; Schubert, U.; Bacik, I.; Basta, S.; O’Neill, R.; Schickli, J.; Palese, P.; et al. A Novel Influenza A Virus Mitochondrial Protein That Induces Cell Death. Nat. Med. 2001, 7, 1306–1312. [Google Scholar] [CrossRef]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Webster, R.G. Molecular Characterization of H9N2 Influenza Viruses: Were They the Donors of the “Internal” Genes of H5N1 Viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 1999, 96, 9363–9367. [Google Scholar] [CrossRef] [PubMed]
- Aamir, U.B.; Wernery, U.; Ilyushina, N.; Webster, R.G. Characterization of Avian H9N2 Influenza Viruses from United Arab Emirates 2000 to 2003. Virology 2007, 361, 45–55. [Google Scholar] [CrossRef]
- Homme, P.J.; Easterday, B.C. Avian Influenza Virus Infections. I. Characteristics of Influenza A-Turkey-Wisconsin-1966 Virus. Avian Dis. 1970, 14, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E.; Slemons, R.D. Proceedings of the Fourth International Symposium on Avian Influenza; American Association of Avian Pathologists: Jacksonville, FL, USA, 1998. [Google Scholar]
- Alexander, D.J. An Overview of the Epidemiology of Avian Influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef]
- Alexander, D.J. Summary of Avian Influenza Activity in Europe, Asia, Africa, and Australasia, 2002–2006. Avian Dis. 2007, 51, 161–166. [Google Scholar] [CrossRef]
- Nagy, A.; Mettenleiter, T.C.; Abdelwhab, E.M. A Brief Summary of the Epidemiology and Genetic Relatedness of Avian Influenza H9N2 Virus in Birds and Mammals in the Middle East and North Africa. Epidemiol. Infect. 2017, 145, 3320–3333. [Google Scholar] [CrossRef]
- Peacock, T.P.; James, J.; Sealy, J.E.; Iqbal, M. A Global Perspective on H9N2 Avian Influenza Virus. Viruses 2019, 11, 620. [Google Scholar] [CrossRef]
- Adlhoch, C.; Authority, E.F.S.; European Centre for Disease Prevention and Control; European Union Reference Laboratory for Avian Influenza; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Mirinaviciute, G.; Niqueux, É.; et al. Avian Influenza Overview December 2022–March 2023. EFSA J. 2023, 21, e07917. [Google Scholar] [CrossRef] [PubMed]
- Kammon, A.; Heidari, A.; Dayhum, A.; Eldaghayes, I.; Sharif, M.; Monne, I.; Cattoli, G.; Asheg, A.; Farhat, M.; Kraim, E. Characterization of Avian Influenza and Newcastle Disease Viruses from Poultry in Libya. Avian Dis. 2015, 59, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Abdelwhab, E.M.; Grund, C.; Aly, M.M.; Beer, M.; Harder, T.C.; Hafez, H.M. Influence of Maternal Immunity on Vaccine Efficacy and Susceptibility of One Day Old Chicks against Egyptian Highly Pathogenic Avian Influenza H5N1. Vet. Microbiol. 2012, 155, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Tombari, W.; Paul, M.; Bettaieb, J.; Larbi, I.; Nsiri, J.; Elbehi, I.; Gribaa, L.; Ghram, A. Risk Factors and Characteristics of Low Pathogenic Avian Influenza Virus Isolated from Commercial Poultry in Tunisia. PLoS ONE 2013, 8, e53524. [Google Scholar] [CrossRef] [PubMed]
- EL Houadfi, M.; Fellahi, S.; Nassik, S.; Guérin, J.-L.; Ducatez, M.F. First Outbreaks and Phylogenetic Analyses of Avian Influenza H9N2 Viruses Isolated from Poultry Flocks in Morocco. Virol. J. 2016, 13, 140. [Google Scholar] [CrossRef]
- Jeevan, T.; Darnell, D.; Gradi, E.A.; Benali, Y.; Kara, R.; Guetarni, D.; Rubrum, A.; Seiler, P.J.; Crumpton, J.C.; Webby, R.J.; et al. A(H9N2) Influenza Viruses Associated with Chicken Mortality in Outbreaks in Algeria 2017. Influenza Other Respir. Viruses 2019, 13, 622–626. [Google Scholar] [CrossRef] [PubMed]
- Zecchin, B.; Minoungou, G.; Fusaro, A.; Moctar, S.; Ouedraogo-Kaboré, A.; Schivo, A.; Salviato, A.; Marciano, S.; Monne, I. Influenza A(H9N2) Virus, Burkina Faso. Emerg. Infect. Dis. 2017, 23, 2118–2119. [Google Scholar] [CrossRef]
- Awuni, J.A.; Bianco, A.; Dogbey, O.J.; Fusaro, A.; Yingar, D.T.; Salviato, A.; Ababio, P.T.; Milani, A.; Bonfante, F.; Monne, I. Avian Influenza H9N2 Subtype in Ghana: Virus Characterization and Evidence of Co-Infection. Avian Pathol. 2019, 48, 470–476. [Google Scholar] [CrossRef]
- Kariithi, H.M.; Welch, C.N.; Ferreira, H.L.; Pusch, E.A.; Ateya, L.O.; Binepal, Y.S.; Apopo, A.A.; Dulu, T.D.; Afonso, C.L.; Suarez, D.L. Genetic Characterization and Pathogenesis of the First H9N2 Low Pathogenic Avian Influenza Viruses Isolated from Chickens in Kenyan Live Bird Markets. Infect. Genet. Evol. 2020, 78, 104074. [Google Scholar] [CrossRef]
- Fusade-Boyer, M.; Djegui, F.; Batawui, K.; Byuragaba, D.K.; Jones, J.C.; Wabwire-Mangeni, F.; Erima, B.; Atim, G.; Ukuli, Q.A.; Tugume, T.; et al. Antigenic and Molecular Characterization of Low Pathogenic Avian Influenza A(H9N2) Viruses in Sub-Saharan Africa from 2017 through 2019. Emerg. Microbes Infect. 2021, 10, 753–761. [Google Scholar] [CrossRef]
- Boumart, Z.; Bamouh, Z.; Jazouli, M.; Zecchin, B.; Fusaro, A.; Salviato, A.; Monne, I.; Tadlaoui, K.O.; Harrak, M.E. Pathogenicity and Full Genome Sequencing of the Avian Influenza H9N2 Moroccan Isolate 2016. Avian Dis 2019, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- El Khantour, A.; Soulaymani, A.; Salek, M.; Maltouf, A.F.; El Mellouli, F.; Ducatez, M.F.; Fellahi, S. Molecular Characterization of the Hemagglutinin Gene of H9N2 Avian Influenza Viruses Isolated from Broiler Flocks in Morocco from 2016 to 2018. Vet. Arh. 2020, 90, 477–484. [Google Scholar] [CrossRef]
- Guo, Y.; Li, J.; Cheng, X. Discovery of men infected by avian influenza A (H9N2) virus. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 1999, 13, 105–108. [Google Scholar] [PubMed]
- Jin, Y.; Cui, H.; Jiang, L.; Zhang, C.; Li, J.; Cheng, H.; Chen, Z.; Zheng, J.; Zhang, Y.; Fu, Y.; et al. Evidence for Human Infection with Avian Influenza A(H9N2) Virus via Environmental Transmission inside Live Poultry Market in Xiamen, China. J. Med. Virol. 2023, 95, e28242. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, J.; Zhang, C.; Li, S.; Yuan, M.; Zhang, C.; Shen, C.; Yang, Y.; Fu, L.; Xu, G.; et al. Genetic, Biological and Epidemiological Study on a Cluster of H9N2 Avian Influenza Virus Infections among Chickens, a Pet Cat, and Humans at a Backyard Farm in Guangxi, China. Emerg. Microbes Infect. 2023, 12, 2143282. [Google Scholar] [CrossRef]
- Xu, J.; Li, S.; Yang, Y.; Liu, B.; Yang, H.; Li, T.; Zhang, L.; Li, W.; Luo, X.; Zhang, L.; et al. Human Infection with a Further Evolved Avian H9N2 Influenza A Virus in Sichuan, China. Sci. China Life Sci. 2018, 61, 604–606. [Google Scholar] [CrossRef]
- Butt, K.M.; Smith, G.J.D.; Chen, H.; Zhang, L.J.; Leung, Y.H.C.; Xu, K.M.; Lim, W.; Webster, R.G.; Yuen, K.Y.; Peiris, J.S.M.; et al. Human Infection with an Avian H9N2 Influenza A Virus in Hong Kong in 2003. J. Clin. Microbiol. 2005, 43, 5760–5767. [Google Scholar] [CrossRef]
- Jallow, M.M.; Fall, A.; Barry, M.A.; Diop, B.; Sy, S.; Goudiaby, D.; Fall, M.; Enouf, V.; Niang, M.N.; Dia, N. Genetic Characterization of the First Detected Human Case of Low Pathogenic Avian Influenza A/H9N2 in Sub-Saharan Africa, Senegal. Emerg. Microbes Infect. 2020, 9, 1092–1095. [Google Scholar] [CrossRef]
- Lin, Y.P.; Shaw, M.; Gregory, V.; Cameron, K.; Lim, W.; Klimov, A.; Subbarao, K.; Guan, Y.; Krauss, S.; Shortridge, K.; et al. Avian-to-Human Transmission of H9N2 Subtype Influenza A Viruses: Relationship between H9N2 and H5N1 Human Isolates. Proc. Natl. Acad. Sci. USA 2000, 97, 9654–9658. [Google Scholar] [CrossRef]
- Belser, J.A.; Sun, X.; Brock, N.; Pappas, C.; Pulit-Penaloza, J.A.; Zeng, H.; Jang, Y.; Jones, J.; Carney, P.J.; Chang, J.; et al. Genetically and Antigenically Divergent Influenza A(H9N2) Viruses Exhibit Differential Replication and Transmission Phenotypes in Mammalian Models. J. Virol. 2020, 94, e00451-20. [Google Scholar] [CrossRef]
- Pusch, L.; Suarez, D. The Multifaceted Zoonotic Risk of H9N2 Avian Influenza. Vet. Sci. 2018, 5, 82. [Google Scholar] [CrossRef] [PubMed]
- Monne, I.; Ormelli, S.; Salviato, A.; De Battisti, C.; Bettini, F.; Salomoni, A.; Drago, A.; Zecchin, B.; Capua, I.; Cattoli, G. Development and Validation of a One-Step Real-Time PCR Assay for Simultaneous Detection of Subtype H5, H7, and H9 Avian Influenza Viruses. J. Clin. Microbiol. 2008, 46, 1769–1773. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.; Stech, J.; Guan, Y.; Webster, R.G.; Perez, D.R. Universal Primer Set for the Full-Length Amplification of All Influenza A Viruses. Arch. Virol. 2001, 146, 2275–2289. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Sikht, F.-Z.; Ducatez, M.; Touzani, C.D.; Rubrum, A.; Webby, R.; El Houadfi, M.; Tligui, N.-S.; Camus, C.; Fellahi, S. Avian Influenza a H9N2 Viruses in Morocco, 2018–2019. Viruses 2022, 14, 529. [Google Scholar] [CrossRef]
- Belkasmi, S.F.Z.; Fellahi, S.; Touzani, C.D.; Faraji, F.Z.; Maaroufi, I.; Delverdier, M.; Guérin, J.-L.; Fihri, O.F.; El Houadfi, M.; Ducatez, M.F. Co-Infections of Chickens with Avian Influenza Virus H9N2 and Moroccan Italy 02 Infectious Bronchitis Virus: Effect on Pathogenesis and Protection Conferred by Different Vaccination Programmes. Avian Pathol. 2020, 49, 21–28. [Google Scholar] [CrossRef]
- El Mellouli, F.; Mouahid, M.; Fusaro, A.; Zecchin, B.; Zekhnini, H.; El Khantour, A.; Giussani, E.; Palumbo, E.; Rguibi Idrissi, H.; Monne, I.; et al. Spatiotemporal Dynamics, Evolutionary History and Zoonotic Potential of Moroccan H9N2 Avian Influenza Viruses from 2016 to 2021. Viruses 2022, 14, 509. [Google Scholar] [CrossRef]
- Capua, I.; Terregino, C.; Cattoli, G.; Toffan, A. Increased Resistance of Vaccinated Turkeys to Experimental Infection with an H7N3 Low-Pathogenicity Avian Influenza Virus. Avian Pathol. 2004, 33, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Park, K.J.; Kwon, H.-I.; Song, M.-S.; Pascua, P.N.Q.; Baek, Y.H.; Lee, J.H.; Jang, H.-L.; Lim, J.-Y.; Mo, I.-P.; Moon, H.-J.; et al. Rapid Evolution of Low-Pathogenic H9N2 Avian Influenza Viruses Following Poultry Vaccination Programmes. J. Gen. Virol. 2011, 92, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-W.; Senne, D.A.; Suarez, D.L. Effect of Vaccine Use in the Evolution of Mexican Lineage H5N2 Avian Influenza Virus. J. Virol. 2004, 78, 8372–8381. [Google Scholar] [CrossRef] [PubMed]
- Bi, J.; Deng, G.; Dong, J.; Kong, F.; Li, X.; Xu, Q.; Zhang, M.; Zhao, L.; Qiao, J. Phylogenetic and Molecular Characterization of H9N2 Influenza Isolates from Chickens in Northern China from 2007–2009. PLoS ONE 2010, 5, e13063. [Google Scholar] [CrossRef]
- Larbi, I.; Ghedira, K.; Arbi, M.; Butcher, G.D.; Rego, N.; Naya, H.; Tougorti, H.; Lachhab, J.; Behi, I.E.; Nsiri, J.; et al. Phylogenetic Analysis and Assessment of the Pathogenic Potential of the First H9N2 Avian Influenza Viruses Isolated from Wild Birds and Lagoon Water in Tunisia. Virus Res. 2022, 322, 198929. [Google Scholar] [CrossRef]
- Khantour, A.E.; Houadfi, M.E.; Nassik, S.; Tligui, N.S.; Mellouli, F.E.; Sikht, F.-Z.; Ducatez, M.F.; Soulaymani, A.; Fellahi, S. Protective Efficacy Evaluation of Four Inactivated Commercial Vaccines Against Low Pathogenic Avian Influenza H9N2 Virus Under Experimental Conditions in Broiler Chickens. Avian Dis. 2021, 65, 15. [Google Scholar] [CrossRef]
- Amal, E.-B.; Saâdi, N.; Asma, F.; Moncef, B.; Ouafae, F.F. Characterization and Phylogenetic Analysis of the Hemagglutinin Gene in H9 Influenza Viruses from Chickens in Morocco from 2017 to 2019. Avian Dis. 2020, 64, 310–314. [Google Scholar] [CrossRef]
- Bush, R.M.; Fitch, W.M.; Bender, C.A.; Cox, N.J. Positive Selection on the H3 Hemagglutinin Gene of Human Influenza Virus A. Mol. Biol. Evol. 1999, 16, 1457–1465. [Google Scholar] [CrossRef]
- García, M.; Suarez, D.L.; Crawford, J.M.; Latimer, J.W.; Slemons, R.D.; Swayne, D.E.; Perdue, M.L. Evolution of H5 Subtype Avian Influenza A Viruses in North America. Virus Res. 1997, 51, 115–124. [Google Scholar] [CrossRef]
- Bahl, J.; Vijaykrishna, D.; Holmes, E.C.; Smith, G.J.D.; Guan, Y. Gene Flow and Competitive Exclusion of Avian Influenza A Virus in Natural Reservoir Hosts. Virology 2009, 390, 289–297. [Google Scholar] [CrossRef]
- Gambaryan, A.; Webster, R.; Matrosovich, M. Differences between Influenza Virus Receptors on Target Cells of Duck and Chicken. Arch. Virol. 2002, 147, 1197–1208. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Belser, J.A.; Maines, T.R. Adaptation of H9N2 Influenza Viruses to Mammalian Hosts: A Review of Molecular Markers. Viruses 2020, 12, 541. [Google Scholar] [CrossRef] [PubMed]
- Vines, A.; Wells, K.; Matrosovich, M.; Castrucci, M.R.; Ito, T.; Kawaoka, Y. The Role of Influenza A Virus Hemagglutinin Residues 226 and 228 in Receptor Specificity and Host Range Restriction. J. Virol. 1998, 72, 7626–7631. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Zhao, Z.; Sang, J.; Jiang, W.; Chen, J.; Tang, T.; Li, Y.; Kan, Q.; Shao, H.; Zhang, J.; et al. Amino Acid Variation at Hemagglutinin Position 193 Impacts the Properties of H9N2 Avian Influenza Virus. J. Virol. 2023, 97, e0137922. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Perez, D.R. Amino Acid 226 in the Hemagglutinin of H9N2 Influenza Viruses Determines Cell Tropism and Replication in Human Airway Epithelial Cells. J. Virol. 2007, 81, 5181–5191. [Google Scholar] [CrossRef]
- Baigent, S.J.; McCauley, J.W. Influenza Type A in Humans, Mammals and Birds: Determinants of Virus Virulence, Host-Range and Interspecies Transmission. Bioessays 2003, 25, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.; Wolff, T.; Herwig, A.; Pleschka, S.; Klenk, H.-D. Interdependence of Hemagglutinin Glycosylation and Neuraminidase as Regulators of Influenza Virus Growth: A Study by Reverse Genetics. J. Virol. 2000, 74, 6316–6323. [Google Scholar] [CrossRef]
- Xu, K.M.; Li, K.S.; Smith, G.J.D.; Li, J.W.; Tai, H.; Zhang, J.X.; Webster, R.G.; Peiris, J.S.M.; Chen, H.; Guan, Y. Evolution and Molecular Epidemiology of H9N2 Influenza A Viruses from Quail in Southern China, 2000 to 2005. J. Virol. 2007, 81, 2635–2645. [Google Scholar] [CrossRef]



| Virus Identification | Date | Location | Type of Production | Age of Birds * | Tissue | Ct Value of qRT-PCR | Accession Number |
|---|---|---|---|---|---|---|---|
| A/chicken/Morocco/OA1/2021 | 07/04/2021 | Rabat | Layers | 34w | Trachea and lung | 29.09 | OR592450 |
| A/chicken/Morocco/OA2/2021 | 02/08/2021 | Casablanca | Broilers | 32d | Trachea and lung | 23.69 | OR592441 |
| A/chicken/Morocco/OA3/2021 | 01/12/2021 | Rabat | Broilers | 34d | Oropharyngeal swabs | 10.29 | OR592474 |
| A/chicken/Morocco/OA4/2021 | 17/12/2021 | Temara | Broilers | 19d | Oropharyngeal swabs | 14.71 | OR592443 |
| A/chicken/Morocco/OA6/2022 | 27/01/2022 | Kénitra | Broilers | 22d | Oropharyngeal swabs | 10.84 | OR592455 |
| A/chicken/Morocco/OA7/2022 | 31/01/2022 | Kénitra | Broilers | 18d | Trachea and lung | 12.02 | OR592451 |
| A/chicken/Morocco/OA8/2022 | 14/03/2022 | Salé | Broilers | 31d | Oropharyngeal swabs | 13.05 | OR592475 |
| A/chicken/Morocco/OA9/2022 | 14/03/2022 | Salé | Broilers | 31d | Trachea and lung | 11.56 | OR592452 |
| A/chicken/Morocco/OA10/2022 | 14/11/2022 | Rabat | Layers | 34w | Trachea and lung | 26.07 | OR592453 |
| A/chicken/Morocco/OA11/2022 | 22/11/2022 | Rabat | Layers | 25w | Trachea and lung | 28.08 | OR592442 |
| A/chicken/Morocco/OA12/2022 | 29/01/2022 | Casablanca | Broilers | 26d | Trachea and lung | 25.22 | OR592454 |
| A/Turkey/Morocco/OA13/2022 | 19/11/2022 | Bouznika | Turkey breeders | 42d | Trachea and lung | 25.06 | OR592514 |
| A/chicken/Morocco/OA14/2022 | 15/02/2022 | Marrakech | Broilers | 24d | Trachea and lung | 25.75 | OR592449 |
| A/chicken/Morocco/OA15/2022 | 10/12/2022 | East region | Broilers | 35d | Trachea and lung | 25.25 | OR592456 |
| A/chicken/Morocco/OA16/2022 | 10/12/2022 | East region | Broilers | 41d | Trachea and lung | 24.85 | OR592448 |
| A/chicken/Morocco/OA17/2022 | 07/11/2022 | Rabat | Broilers | 19d | Trachea and lung | 23.72 | OR592457 |
| A/chicken/Morocco/OA18/2022 | 09/11/2022 | Fes | Broilers | 27d | Trachea and lung | 21.92 | OR592447 |
| A/chicken/Morocco/OA19/2023 | 03/01/2023 | Meknes | Broilers | 21d | Trachea and lung | 19.44 | OR592444 |
| A/chicken/Morocco/OA20/2023 | 13/01/2023 | Casablanca | Layers | 30w | Trachea and lung | 28.6 | OR592513 |
| A/chicken/Morocco/OA22/2023 | 12/01/2023 | Fes | Broilers | 47d | Trachea and lung | 23.89 | OR592446 |
| A/chicken/Morocco/OA23/2023 | 01/03/2023 | Temara | Broilers | 28d | Trachea and lung | 28.68 | OR592445 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amino-Acid Genetic Distance | ||||||||||||||||
| SF1/2016 | 0.004 | 0.09 | 0.004 | 0.009 | 0 | 0.002 | 0.004 | 0.025 | 0.02 | 0.016 | 0.03 | 0.011 | 0.032 | 0.032 | 0.011 | |
| AS13/2018 | 0.01 | 0.092 | 0.007 | 0.013 | 0.005 | 0.005 | 0.007 | 0.03 | 0.015 | 0.011 | 0.025 | 0.017 | 0.04 | 0.027 | 0.011 | |
| Hong_Kong/G1/1997 | 0.117 | 0.128 | 0.09 | 0.094 | 0.033 | 0.092 | 0.095 | 0.05 | 0.051 | 0.043 | 0.06 | 0.039 | 0.048 | 0.06 | 0.039 | |
| Algeria/220/2017 | 0.007 | 0.02 | 0.123 | 0.009 | 0 | 0.005 | 0.007 | 0.025 | 0.02 | 0.016 | 0.03 | 0.011 | 0.032 | 0.032 | 0.011 | |
| Tunisia/117/2018 | 0.016 | 0.03 | 0.124 | 0.019 | 0 | 0.011 | 0.013 | 0.025 | 0.02 | 0.016 | 0.03 | 0.011 | 0.032 | 0.032 | 0.011 | |
| Fes/EM1/2016 | 0 | 0.018 | 0.071 | 0.004 | 0 | 0 | 0 | 0.011 | 0.011 | 0.016 | 0.016 | 0.011 | 0.033 | 0.033 | 0.011 | |
| SF7/2017 | 0.002 | 0.015 | 0.119 | 0.008 | 0.018 | 0.002 | 0.005 | 0.025 | 0.02 | 0.016 | 0.03 | 0.011 | 0.032 | 0.032 | 0.011 | |
| Morocco/14-1_4636/2020 | 0.019 | 0.022 | 0.132 | 0.026 | 0.034 | 0.016 | 0.021 | 0.026 | 0.021 | 0.016 | 0.031 | 0.011 | 0.033 | 0.033 | 0.011 | |
| OA2/2021 | 0.04 | 0.045 | 0.101 | 0.043 | 0.04 | 0.027 | 0.041 | 0.047 | 0.025 | 0.027 | 0.035 | 0.022 | 0.032 | 0.043 | 0.022 | |
| OA13/2022 | 0.035 | 0.027 | 0.099 | 0.039 | 0.035 | 0.024 | 0.034 | 0.031 | 0.044 | 0.016 | 0.02 | 0.022 | 0.04 | 0.033 | 0.022 | |
| OA23/2023 | 0.036 | 0.025 | 0.087 | 0.039 | 0.036 | 0.037 | 0.038 | 0.03 | 0.041 | 0.038 | 0.021 | 0.028 | 0.04 | 0.038 | 0.022 | |
| OA3/2021 | 0.052 | 0.039 | 0.102 | 0.055 | 0.051 | 0.035 | 0.053 | 0.049 | 0.052 | 0.044 | 0.016 | 0.028 | 0.056 | 0.038 | 0.022 | |
| OA11/2022 | 0.026 | 0.044 | 0.088 | 0.03 | 0.026 | 0.026 | 0.028 | 0.038 | 0.043 | 0.05 | 0.048 | 0.05 | 0.024 | 0.022 | 0 | |
| OA20/2023 | 0.048 | 0.069 | 0.092 | 0.051 | 0.048 | 0.046 | 0.051 | 0.063 | 0.06 | 0.074 | 0.063 | 0.071 | 0.013 | 0.04 | 0.025 | |
| OA6/2022 | 0.034 | 0.047 | 0.093 | 0.038 | 0.034 | 0.031 | 0.036 | 0.046 | 0.052 | 0.054 | 0.051 | 0.052 | 0.009 | 0.018 | 0.016 | |
| OA4/2021 | 0.026 | 0.041 | 0.086 | 0.029 | 0.026 | 0.026 | 0.027 | 0.037 | 0.048 | 0.048 | 0.048 | 0.05 | 0.011 | 0.025 | 0.014 | |
| Nucleotide Genetic Distance | ||||||||||||||||
| Strains | Position NT (H3 Numbering) | Receptor Binding Sites (H3 Numbering) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 196 | 197 | 198 | 230 | 191 | 192 | 193 | 194 | 195 | 221 | 225 | 226 | 227 | 228 | |
| Beijing/3/99 a | N | R | R | I | N | N | K | C | D | * | V | A | R | S |
| Hong_2_Kong/1997 b | I | R | N | I | Q | T | N | L | Y | P | G | L | Q | G |
| SF1/2016 c | T | R | T | I | Q | T | N | L | Y | P | G | L | I | G |
| AS14/2018 d | T | R | T | I | Q | T | N | L | Y | P | G | L | I | G |
| AS77/2019 e | T | R | T | I | Q | T | N | L | Y | P | G | L | I | G |
| 17-6650/2020 f | T | R | T | I | Q | T | N | L | Y | P | G | L | I | G |
| OA1/2021 g | R | Q | L | R | G | F | Q | L | K | M | N | S | V | R |
| OA2/2021 h | R | Q | L | R | G | F | Q | L | K | M | N | S | V | R |
| OA15/2022 i | R | Q | L | R | G | F | Q | L | K | M | N | S | V | R |
| OA16/2022 j | R | Q | L | R | G | F | Q | L | K | M | N | S | V | R |
| OA17/2022 k | R | Q | L | R | G | F | Q | L | K | M | N | S | V | R |
| OA18/2022 l | R | Q | L | R | G | F | Q | L | K | M | N | S | V | R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arbani, O.; Ducatez, M.F.; Mahmoudi, S.; Salamat, F.; Khayi, S.; Mouahid, M.; Selim, K.M.; Kichou, F.; Ouchhour, I.; El Houadfi, M.; et al. Low Pathogenic Avian Influenza H9N2 Viruses in Morocco: Antigenic and Molecular Evolution from 2021 to 2023. Viruses 2023, 15, 2355. https://doi.org/10.3390/v15122355
Arbani O, Ducatez MF, Mahmoudi S, Salamat F, Khayi S, Mouahid M, Selim KM, Kichou F, Ouchhour I, El Houadfi M, et al. Low Pathogenic Avian Influenza H9N2 Viruses in Morocco: Antigenic and Molecular Evolution from 2021 to 2023. Viruses. 2023; 15(12):2355. https://doi.org/10.3390/v15122355
Chicago/Turabian StyleArbani, Oumayma, Mariette F. Ducatez, Salma Mahmoudi, Faiçal Salamat, Slimane Khayi, Mohamed Mouahid, Karim M. Selim, Faouzi Kichou, Ikram Ouchhour, Mohammed El Houadfi, and et al. 2023. "Low Pathogenic Avian Influenza H9N2 Viruses in Morocco: Antigenic and Molecular Evolution from 2021 to 2023" Viruses 15, no. 12: 2355. https://doi.org/10.3390/v15122355
APA StyleArbani, O., Ducatez, M. F., Mahmoudi, S., Salamat, F., Khayi, S., Mouahid, M., Selim, K. M., Kichou, F., Ouchhour, I., El Houadfi, M., & Fellahi, S. (2023). Low Pathogenic Avian Influenza H9N2 Viruses in Morocco: Antigenic and Molecular Evolution from 2021 to 2023. Viruses, 15(12), 2355. https://doi.org/10.3390/v15122355