


Unraveling Immunological Dynamics: HPV Infection in Women—Insights from Pregnancy


 , ,
, ,
Abstract
1. Introduction
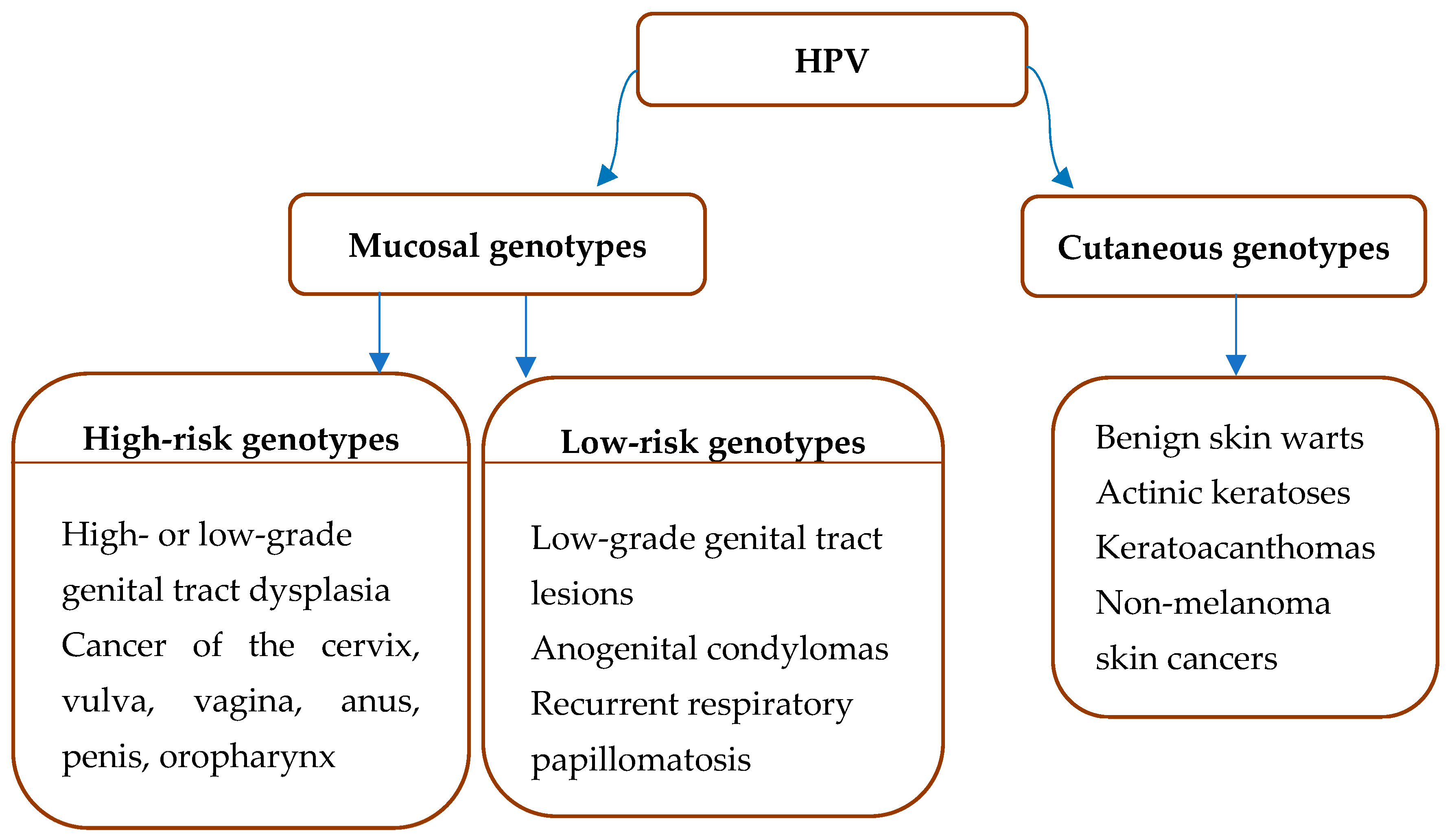
2. HPV Types and Associated Diseases
HPV Structure and Life Cycle
3. HPV Infection during Pregnancy
4. The Immune Response to HPV under Normal Conditions and during Pregnancy
4.1. The Innate Immune Response
4.2. The Adaptive Immune Response
5. HPV Vaccines
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hamborsky, J.; Kroger, A. Epidemiology and Prevention of Vaccine-Preventable Diseases; E-Book: The Pink Book; Public Health Foundation: Washington, DC, USA, 2015. [Google Scholar]
- Petca, A.; Borislavschi, A.; Zvanca, M.E.; Petca, R.C.; Sandru, F.; Dumitrascu, M.C. Non-sexual HPV transmission and role of vaccination for a better future (Review). Exp. Ther. Med. 2020, 20, 186. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Weaver, B. Human papillomavirus in older women: New infection or reactivation? J. Infect. Dis. 2013, 207, 211–212. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.K.; Aimagambetova, G.; Ukybassova, T.; Kongrtay, K.; Azizan, A. Human Papillomavirus Infection and Cervical Cancer: Epidemiology, Screening, and Vaccination-Review of Current Perspectives. J. Oncol. 2019, 2019, 3257939. [Google Scholar] [CrossRef] [PubMed]
- Szymonowicz, K.A.; Chen, J. Biological and clinical aspects of HPV-related cancers. Cancer Biol. Med. 2020, 17, 864–878. [Google Scholar] [CrossRef]
- Bzhalava, D.; Guan, P.; Franceschi, S.; Dillner, J.; Clifford, G. A systematic review of the prevalence of mucosal and cutaneous human papillomavirus types. Virology 2013, 445, 224–231. [Google Scholar] [CrossRef]
- Hufbauer, M.; Akgül, B. Molecular Mechanisms of Human Papillomavirus Induced Skin Carcinogenesis. Viruses 2017, 9, 187. [Google Scholar] [CrossRef]
- Forslund, O.; DeAngelis, P.M.; Beigi, M.; Schjølberg, A.R.; Clausen, O.P. Identification of human papillomavirus in keratoacanthomas. J. Cutan. Pathol. 2003, 30, 423–429. [Google Scholar] [CrossRef]
- Asgari, M.M.; Kiviat, N.B.; Critchlow, C.W.; Stern, J.E.; Argenyi, Z.B.; Raugi, G.J.; Berg, D.; Odland, P.B.; Hawes, S.E.; de Villiers, E.-M. Detection of human papillomavirus DNA in cutaneous squamous cell carcinoma among immunocompetent individuals. J. Investig. Dermatol. 2008, 128, 1409–1417. [Google Scholar] [CrossRef]
- Al-Eitan, L.N.; Tarkhan, A.H.; Alghamdi, M.A.; Al-Qarqaz, F.A.; Al-Kofahi, H.S. Transcriptome analysis of HPV-induced warts and healthy skin in humans. BMC Med. Genom. 2020, 13, 35. [Google Scholar] [CrossRef]
- Jaan, A.; Rajnik, M. TORCH Complex. In StatPearls; StatPearls Publishing LLC.: Tampa, FL, USA, 2023. [Google Scholar]
- Lynn, M.K.; Aquino, M.S.R.; Self, S.C.W.; Kanyangarara, M.; Campbell, B.A.; Nolan, M.S. TORCH Congenital Syndrome Infections in Central America’s Northern Triangle. Microorganisms 2023, 11, 257. [Google Scholar]
- Bzhalava, D.; Eklund, C.; Dillner, J. International standardization and classification of human papillomavirus types. Virology 2015, 476, 341–344. [Google Scholar] [CrossRef] [PubMed]
- International HPV Reference Center. Karolinska Universitetssjukhuset. Available online: https://www.hpvcenter.se/human_reference_clones/ (accessed on 24 January 2023).
- World Health Organization. Human Papillomaviruses. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Publications: Lyon, France, 2007; Volume 90. [Google Scholar]
- Arbyn, M.; Tommasino, M.; Depuydt, C.; Dillner, J. Are 20 human papillomavirus types causing cervical cancer? J. Pathol. 2014, 234, 431–435. [Google Scholar] [CrossRef] [PubMed]
- de Sanjosé, S.; Serrano, B.; Tous, S.; Alejo, M.; Lloveras, B.; Quirós, B.; Clavero, O.; Vidal, A.; Ferrándiz-Pulido, C.; Pavón, M.; et al. Burden of Human Papillomavirus (HPV)-Related Cancers Attributable to HPVs 6/11/16/18/31/33/45/52 and 58. JNCI Cancer Spectr. 2018, 2, pky045. [Google Scholar] [CrossRef] [PubMed]
- Capra, G.; Giovannelli, L.; Bellavia, C.; Migliore, M.C.; Caleca, M.P.; Perino, A.; Ammatuna, P. HPV genotype prevalence in cytologically abnormal cervical samples from women living in south Italy. Virus Res. 2008, 133, 195–200. [Google Scholar] [CrossRef]
- Ciavattini, A.; Delli Carpini, G.; Giannella, L.; del Fabro, A.; Banerji, V.; Hall, G.; Barbero, M.; Sopracordevole, F. An online survey on emotions, impact on everyday life, and educational needs of women with HPV positivity or abnormal Pap smear result. Medicine 2021, 100, e27177. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.C.; McMillan, N.A.J.; Antonsson, A. Human papillomavirus type spectrum in normal skin of individuals with or without a history of frequent sun exposure. J. Gen. Virol. 2008, 89, 2891–2897. [Google Scholar] [CrossRef]
- Accardi, R.; Gheit, T. Cutaneous HPV and skin cancer. Presse Med. 2014, 43, e435–e443. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Castle, P.E.; Maza, M. Prophylactic HPV vaccination: Past, present, and future. Epidemiol. Infect. 2016, 144, 449–468. [Google Scholar] [CrossRef]
- Silva Dalla Libera, L.; de Carvalho, K.P.A.; Enocencio Porto Ramos, J.; Oliveira Cabral, L.A.; de Cassia Goncalves de Alencar, R.; Villa, L.L.; Alves, R.R.F.; Rabelo Santos, S.H.; Aparecida Dos Santos Carneiro, M.; Saddi, V.A. Human Papillomavirus and Anal Cancer: Prevalence, Genotype Distribution, and Prognosis Aspects from Midwestern Region of Brazil. J. Oncol. 2019, 2019, 6018269. [Google Scholar] [CrossRef]
- Kreimer, A.R.; Chaturvedi, A.K.; Alemany, L.; Anantharaman, D.; Bray, F.; Carrington, M.; Doorbar, J.; D’Souza, G.; Fakhry, C.; Ferris, R.L.; et al. Summary from an international cancer seminar focused on human papillomavirus (HPV)-positive oropharynx cancer, convened by scientists at IARC and NCI. Oral Oncol. 2020, 108, 104736. [Google Scholar] [CrossRef] [PubMed]
- Baral, S.K.; Biswas, P.; Kaium, M.A.; Islam, M.A.; Dey, D.; Saber, M.A.; Rahaman, T.I.; M, A.; Emran, T.B.; Hasan, M.N.; et al. A Comprehensive Discussion in Vaginal Cancer Based on Mechanisms, Treatments, Risk Factors and Prevention. Front. Oncol. 2022, 12, 883805. [Google Scholar] [CrossRef] [PubMed]
- de Vuyst, H.; Clifford, G.M.; Nascimento, M.C.; Madeleine, M.M.; Franceschi, S. Prevalence and type distribution of human papillomavirus in carcinoma and intraepithelial neoplasia of the vulva, vagina and anus: A meta-analysis. Int. J. Cancer 2009, 124, 1626–1636. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Álvarez, M.I.; Gómez-Urquiza, J.L.; Husein-El Ahmed, H.; Albendín-García, L.; Gómez-Salgado, J.; la Fuente, G.A.C.-D. Prevalence and Risk Factors of Human Papillomavirus in Male Patients: A Systematic Review and Meta-Analysis. Int. J. Environ. Res. Public Health 2018, 15, 2210. [Google Scholar] [CrossRef]
- Moscicki, A.B.; Palefsky, J.M. Human papillomavirus in men: An update. J. Low. Genit. Tract Dis. 2011, 15, 231–234. [Google Scholar] [CrossRef]
- Capra, G.; Nyitray, A.G.; Lu, B.; Perino, A.; Marci, R.; Schillaci, R.; Matranga, D.; Firenze, A.; Caleca, M.; Bellavia, C.; et al. Analysis of persistence of human papillomavirus infection in men evaluated by sampling multiple genital sites. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4153–4163. [Google Scholar]
- Capra, G.; Notari, T.; Buttà, M.; Serra, N.; Rizzo, G.; Bosco, L. Human Papillomavirus (HPV) Infection and Its Impact on Male Infertility. Life 2022, 12, 1919. [Google Scholar] [CrossRef]
- Burley, M.; Roberts, S.; Parish, J.L. Epigenetic regulation of human papillomavirus transcription in the productive virus life cycle. Semin. Immunopathol. 2020, 42, 159–171. [Google Scholar] [CrossRef]
- Münger, K. Papillomaviruses. In Encyclopedia of Cancer, 2nd ed.; Bertino, J.R., Ed.; Academic Press: New York, NY, USA, 2002; pp. 393–401. [Google Scholar]
- Makowsky, R.; Lhaki, P.; Wiener, H.W.; Bhatta, M.P.; Cullen, M.; Johnson, D.C.; Perry, R.T.; Lama, M.; Boland, J.F.; Yeager, M.; et al. Genomic diversity and phylogenetic relationships of human papillomavirus 16 (HPV16) in Nepal. Infect. Genet. Evol. 2016, 46, 7–11. [Google Scholar] [CrossRef][Green Version]
- Yu, L.; Majerciak, V.; Zheng, Z.M. HPV16 and HPV18 Genome Structure, Expression, and Post-Transcriptional Regulation. Int. J. Mol. Sci. 2022, 23, 4943. [Google Scholar] [CrossRef]
- Zheng, Z.M.; Baker, C.C. Papillomavirus genome structure, expression, and post-transcriptional regulation. Front. Biosci. A J. Virtual Libr. 2006, 11, 2286–2302. [Google Scholar] [CrossRef]
- McBride, A.A.; Oliveira, J.G.; McPhillips, M.G. Partitioning viral genomes in mitosis: Same idea, different targets. Cell Cycle 2006, 5, 1499–1502. [Google Scholar] [CrossRef]
- Kajitani, N.; Satsuka, A.; Kawate, A.; Sakai, H. Productive Lifecycle of Human Papillomaviruses that Depends Upon Squamous Epithelial Differentiation. Front. Microbiol. 2012, 3, 152. [Google Scholar] [CrossRef] [PubMed]
- Sanclemente, G.; Gill, D.K. Human papillomavirus molecular biology and pathogenesis. J. Eur. Acad. Dermatol. Venereol. 2002, 16, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Peralta-Zaragoza, O.; Deas, J.; Gómez-Cerón, C.; García-Suastegui, W.A.; Fierros-Zárate, G.d.S.; Jacobo-Herrera, N.J. HPV-Based Screening, Triage, Treatment, and Followup Strategies in the Management of Cervical Intraepithelial Neoplasia. Obstet. Gynecol. Int. 2013, 2013, 912780. [Google Scholar] [CrossRef] [PubMed]
- Vande Pol, S.B.; Klingelhutz, A.J. Papillomavirus E6 oncoproteins. Virology 2013, 445, 115–137. [Google Scholar] [CrossRef] [PubMed]
- Roman, A.; Munger, K. The papillomavirus E7 proteins. Virology 2013, 445, 138–168. [Google Scholar] [CrossRef]
- Ishii, Y.; Ozaki, S.; Tanaka, K.; Kanda, T. Human Papillomavirus 16 Minor Capsid Protein L2 Helps Capsomeres Assemble Independently of Intercapsomeric Disulfide Bonding. Virus Genes 2005, 31, 321–328. [Google Scholar] [CrossRef]
- Yazdani, R.; Shams-Bakhsh, M.; Hassani-Mehraban, A.; Arab, S.S.; Thelen, N.; Thiry, M.; Crommen, J.; Fillet, M.; Jacobs, N.; Brans, A.; et al. Production and characterization of virus-like particles of grapevine fanleaf virus presenting L2 epitope of human papillomavirus minor capsid protein. BMC Biotechnol. 2019, 19, 81. [Google Scholar] [CrossRef]
- Burchell, A.N.; Winer, R.L.; de Sanjosé, S.; Franco, E.L. Chapter 6: Epidemiology and transmission dynamics of genital HPV infection. Vaccine 2006, 24 (Suppl. 3), S52–S61. [Google Scholar] [CrossRef]
- Chattopadhyay, K. A comprehensive review on host genetic susceptibility to human papillomavirus infection and progression to cervical cancer. Indian J. Hum. Genet. 2011, 17, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, P.K.; Sparén, P.; Gyllensten, U.B. Genetic link to cervical tumours. Nature 1999, 400, 29–30. [Google Scholar] [CrossRef] [PubMed]
- Pontillo, A.; Bricher, P.; Leal, V.N.; Lima, S.; Souza, P.R.; Crovella, S. Role of inflammasome genetics in susceptibility to HPV infection and cervical cancer development. J. Med. Virol. 2016, 88, 1646–1651. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.F.; Koutsky, L.A.; Castle, P.E.; Edelstein, Z.R.; Meyers, C.; Ho, J.; Schiffman, M. Relationship between cigarette smoking and human papilloma virus types 16 and 18 DNA load. Cancer Epidemiol. Biomark. Prev. 2009, 18, 3490–3496. [Google Scholar] [CrossRef]
- Smith, E.M.; Rubenstein, L.M.; Haugen, T.H.; Hamsikova, E.; Turek, L.P. Tobacco and alcohol use increases the risk of both HPV-associated and HPV-independent head and neck cancers. Cancer Causes Control 2010, 21, 1369–1378. [Google Scholar] [CrossRef]
- Schabath, M.B.; Thompson, Z.J.; Egan, K.M.; Torres, B.N.; Nguyen, A.; Papenfuss, M.R.; Abrahamsen, M.E.; Giuliano, A.R. Alcohol consumption and prevalence of human papillomavirus (HPV) infection among US men in the HPV in Men (HIM) study. Sex. Transm. Infect. 2015, 91, 61–67. [Google Scholar] [CrossRef]
- Itarat, Y.; Kietpeerakool, C.; Jampathong, N.; Chumworathayi, B.; Kleebkaow, P.; Aue-Aungkul, A.; Nhokaew, W. Sexual behavior and infection with cervical human papillomavirus types 16 and 18. Int. J. Womens Health 2019, 11, 489–494. [Google Scholar] [CrossRef]
- Farahmand, M.; Moghoofei, M.; Dorost, A.; Abbasi, S.; Monavari, S.H.; Kiani, S.J.; Tavakoli, A. Prevalence and genotype distribution of genital human papillomavirus infection in female sex workers in the world: A systematic review and meta-analysis. BMC Public Health 2020, 20, 1455. [Google Scholar] [CrossRef]
- González-Cano, M.; Garrido-Peña, F.; Gil-Garcia, E.; Lima-Serrano, M.; Cano-Caballero, M.D. Sexual behaviour, human papillomavirus and its vaccine: A qualitative study of adolescents and parents in Andalusia. BMC Public Health 2021, 21, 1476. [Google Scholar] [CrossRef]
- Ribeiro, A.A.; Costa, M.C.; Alves, R.R.F.; Villa, L.L.; Saddi, V.A.; Carneiro, M.A.d.S.; Zeferino, L.C.; Rabelo-Santos, S.H. HPV infection and cervical neoplasia: Associated risk factors. Infect. Agents Cancer 2015, 10, 16. [Google Scholar] [CrossRef]
- Wieland, U.; Kreuter, A.; Pfister, H. Human Papillomavirus and Immunosuppression. In Human Papillomavirus Bench to Bedside; Current Problems in Dermatology Series; Karger: Basel, Switzerland, 2014; Volume 45, pp. 154–165. [Google Scholar] [CrossRef]
- Basu, P.; Malvi, S.G.; Joshi, S.; Bhatla, N.; Muwonge, R.; Lucas, E.; Verma, Y.; Esmy, P.O.; Poli, U.R.R.; Shah, A.; et al. Vaccine efficacy against persistent human papillomavirus (HPV) 16/18 infection at 10 years after one, two, and three doses of quadrivalent HPV vaccine in girls in India: A multicentre, prospective, cohort study. Lancet Oncol. 2021, 22, 1518–1529. [Google Scholar] [CrossRef] [PubMed]
- Siddharthan, R. Anal intraepithelial neoplasia: Diagnosis, screening, and treatment. Ann. Gastroenterol. 2019, 32, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Adebamowo, S.N.; Adeyemo, A.A.; Rotimi, C.N.; Olaniyan, O.; Offiong, R.; Adebamowo, C.A.; Odutola, M.; Dareng, E.O.; Famooto, A.O.; Adebiyi, R.; et al. Genome-wide association study of prevalent and persistent cervical high-risk human papillomavirus (HPV) infection. BMC Med. Genet. 2020, 21, 231. [Google Scholar] [CrossRef] [PubMed]
- Bernal-Silva, S.; Granados, J.; Gorodezky, C.; Aláez, C.; Flores-Aguilar, H.; Cerda-Flores, R.M.; Guerrero-González, G.; Valdez-Chapa, L.D.; Morales-Casas, J.; González-Guerrero, J.F.; et al. HLA-DRB1 Class II antigen level alleles are associated with persistent HPV infection in Mexican women; a pilot study. Infect. Agents Cancer 2013, 8, 31. [Google Scholar] [CrossRef]
- Hubert, W.G.; Laimins, L.A. Human papillomavirus type 31 replication modes during the early phases of the viral life cycle depend on transcriptional and posttranscriptional regulation of E1 and E2 expression. J. Virol. 2002, 76, 2263–2273. [Google Scholar] [CrossRef][Green Version]
- Sapp, M.; Bienkowska-Haba, M. Viral entry mechanisms: Human papillomavirus and a long journey from extracellular matrix to the nucleus. FEBS J. 2009, 276, 7206–7216. [Google Scholar] [CrossRef]
- Richards, K.F.; Bienkowska-Haba, M.; Dasgupta, J.; Chen, X.S.; Sapp, M. Multiple heparan sulfate binding site engagements are required for the infectious entry of human papillomavirus type 16. J. Virol. 2013, 87, 11426–11437. [Google Scholar] [CrossRef]
- Richards, K.F.; Mukherjee, S.; Bienkowska-Haba, M.; Pang, J.; Sapp, M. Human Papillomavirus Species-Specific Interaction with the Basement Membrane-Resident Non-Heparan Sulfate Receptor. Viruses 2014, 6, 4856–4879. [Google Scholar] [CrossRef]
- DiGiuseppe, S.; Bienkowska-Haba, M.; Hilbig, L.; Sapp, M. The nuclear retention signal of HPV16 L2 protein is essential for incoming viral genome to transverse the trans-Golgi network. Virology 2014, 458–459, 93–105. [Google Scholar] [CrossRef]
- Culp, T.D.; Christensen, N.D. Kinetics of in vitro adsorption and entry of papillomavirus virions. Virology 2004, 319, 152–161. [Google Scholar] [CrossRef]
- Selinka, H.-C.; Giroglou, T.; Sapp, M. Analysis of the infectious entry pathway of human papillomavirus type 33 pseudovirions. Virology 2002, 299, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Spoden, G.; Freitag, K.; Husmann, M.; Boller, K.; Sapp, M.; Lambert, C.; Florin, L. Clathrin-and caveolin-independent entry of human papillomavirus type 16—Involvement of tetraspanin-enriched microdomains (TEMs). PLoS ONE 2008, 3, e3313. [Google Scholar] [CrossRef] [PubMed]
- Abbate, E.A.; Berger, J.M.; Botchan, M.R. The X-ray structure of the papillomavirus helicase in complex with its molecular matchmaker E2. Genes Dev. 2004, 18, 1981–1996. [Google Scholar] [CrossRef] [PubMed]
- Bergvall, M.; Melendy, T.; Archambault, J. The E1 proteins. Virology 2013, 445, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Dakic, A.; Zhang, Y.; Dai, Y.; Chen, R.; Schlegel, R. HPV E6 protein interacts physically and functionally with the cellular telomerase complex. Proc. Natl. Acad. Sci. USA 2009, 106, 18780–18785. [Google Scholar] [CrossRef]
- McMurray, H.R.; McCance, D.J. Human papillomavirus type 16 E6 activates TERT gene transcription through induction of c-Myc and release of USF-mediated repression. J. Virol. 2003, 77, 9852–9861. [Google Scholar] [CrossRef]
- Zouboulis, C.C.; Adjaye, J.; Akamatsu, H.; Moe-Behrens, G.; Niemann, C. Human skin stem cells and the ageing process. Exp. Gerontol. 2008, 43, 986–997. [Google Scholar] [CrossRef]
- White, E.A. Manipulation of Epithelial Differentiation by HPV Oncoproteins. Viruses 2019, 11, 369. [Google Scholar] [CrossRef]
- Masterson, P.J.; Stanley, M.A.; Lewis, A.P.; Romanos, M.A. A C-terminal helicase domain of the human papillomavirus E1 protein binds E2 and the DNA polymerase alpha-primase p68 subunit. J. Virol. 1998, 72, 7407–7419. [Google Scholar] [CrossRef]
- Yajid, A.I.; Zakariah, M.A.; Mat Zin, A.A.; Othman, N.H. Potential Role of E4 Protein in Human Papillomavirus Screening: A Review. Asian Pac. J. Cancer Prev. 2017, 18, 315–319. [Google Scholar] [CrossRef]
- Galloway, D.A. Human papillomaviruses: A growing field. Genes Dev. 2009, 23, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Roden, R.B. L2, the minor capsid protein of papillomavirus. Virology 2013, 445, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Siddiqa, A.; Massimi, P.; Pim, D.; Banks, L. Diverse Papillomavirus Types Induce Endosomal Tubulation. Front. Cell. Infect. Microbiol. 2019, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Stanley, M. Pathology and epidemiology of HPV infection in females. Gynecol. Oncol. 2010, 117, S5–S10. [Google Scholar] [CrossRef] [PubMed]
- Anaya-Saavedra, G.; Ramírez-Amador, V.; Irigoyen-Camacho, M.E.; García-Cuellar, C.M.; Guido-Jiménez, M.; Méndez-Martínez, R.; García-Carrancá, A. High association of human papillomavirus infection with oral cancer: A case-control study. Arch. Med. Res. 2008, 39, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Gillison, M.L.; D’Souza, G.; Westra, W.; Sugar, E.; Xiao, W.; Begum, S.; Viscidi, R. Distinct risk factor profiles for human papillomavirus type 16-positive and human papillomavirus type 16-negative head and neck cancers. J. Natl. Cancer Inst. 2008, 100, 407–420. [Google Scholar] [CrossRef]
- Smith, E.M.; Ritchie, J.M.; Summersgill, K.F.; Klussmann, J.P.; Lee, J.H.; Wang, D.; Haugen, T.H.; Turek, L.P. Age, sexual behavior and human papillomavirus infection in oral cavity and oropharyngeal cancers. Int. J. Cancer 2004, 108, 766–772. [Google Scholar] [CrossRef]
- Frega, A.; Cenci, M.; Stentella, P.; Cipriano, L.; de Ioris, A.; Alderisio, M.; Vecchione, A. Human papillomavirus in virgins and behaviour at risk. Cancer Lett. 2003, 194, 21–24. [Google Scholar] [CrossRef]
- Syrjänen, S.; Puranen, M. Human papillomavirus infections in children: The potential role of maternal transmission. Crit. Rev. Oral Biol. Med. 2000, 11, 259–274. [Google Scholar] [CrossRef]
- Rombaldi, R.L.; Serafini, E.P.; Mandelli, J.; Zimmermann, E.; Losquiavo, K.P. Perinatal transmission of human papilomavirus DNA. Virol. J. 2009, 6, 83. [Google Scholar] [CrossRef]
- Fredericks, B.D.; Balkin, A.; Daniel, H.W.; Schonrock, J.; Ward, B.; Frazer, I.H. Transmission of human papillomaviruses from mother to child. Aust. N. Z. J. Obstet. Gynaecol. 1993, 33, 30–32. [Google Scholar] [CrossRef]
- Cason, J.; Kaye, J.N.; Jewers, R.J.; Kambo, P.K.; Bible, J.M.; Kell, B.; Shergill, B.; Pakarian, F.; Raju, K.S.; Best, J.M. Perinatal infection and persistence of human papillomavirus types 16 and 18 in infants. J. Med. Virol. 1995, 47, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, T.Q.; Derkay, C.S. Recurrent respiratory papillomatosis: Update 2008. Curr. Opin. Otolaryngol. Head Neck Surg. 2008, 16, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Derkay, C.S.; Wiatrak, B. Recurrent respiratory papillomatosis: A review. Laryngoscope 2008, 118, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Puranen, M.H.; Yliskoski, M.H.; Saarikoski, S.V.; Syrjänen, K.J.; Syrjänen, S.M. Exposure of an infant to cervical human papillomavirus infection of the mother is common. Am. J. Obstet. Gynecol. 1997, 176, 1039–1045. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, Q.; Rao, H. Maternal-fetal transmission of human papillomavirus. Chin. Med. J. 1998, 111, 726–727. [Google Scholar]
- Tseng, C.J.; Liang, C.C.; Soong, Y.K.; Pao, C.C. Perinatal transmission of human papillomavirus in infants: Relationship between infection rate and mode of delivery. Obstet. Gynecol. 1998, 91, 92–96. [Google Scholar] [CrossRef]
- Çağlar, G.S.; Garrido, N. The implications of male human papilloma virus infection in couples seeking assisted reproduction technologies. J. Turk. Ger. Gynecol. Assoc. 2018, 19, 48–52. [Google Scholar] [CrossRef]
- Armbruster-Moraes, E.; Ioshimoto, L.M.; Leão, E.; Zugaib, M. Presence of human papillomavirus DNA in amniotic fluids of pregnant women with cervical lesions. Gynecol. Oncol. 1994, 54, 152–158. [Google Scholar] [CrossRef]
- Favre, M.; Majewski, S.; De Jesus, N.; Malejczyk, M.; Orth, G.; Jablonska, S. A possible vertical transmission of human papillomavirus genotypes associated with epidermodysplasia verruciformis. J. Investig. Dermatol. 1998, 111, 333–336. [Google Scholar] [CrossRef]
- Xu, S.; Liu, L.; Lu, S.; Ren, S. Clinical observation on vertical transmission of human papillomavirus. Chin. Med. Sci. J. 1998, 13, 29–31. [Google Scholar] [PubMed]
- Yoshida, K.; Furumoto, H.; Abe, A.; Kato, T.; Nishimura, M.; Kuwahara, A.; Maeda, K.; Matsuzaki, T.; Irahara, M. The possibility of vertical transmission of human papillomavirus through maternal milk. J. Obstet. Gynaecol. 2011, 31, 503–506. [Google Scholar] [CrossRef] [PubMed]
- Sarkola, M.; Rintala, M.; Grénman, S.; Syrjänen, S. Human Papillomavirus DNA Detected in Breast Milk. Pediatr. Infect. Dis. J. 2008, 27, 557–558. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, M.; Gheit, T.; Casadio, C.; Khan, N.; Macis, D.; Valenti, F.; Miller, M.J.; Sylla, B.S.; Akiba, S.; Bonanni, B.; et al. Analysis of the presence of cutaneous and mucosal papillomavirus types in ductal lavage fluid, milk and colostrum to evaluate its role in breast carcinogenesis. Breast Cancer Res. Treat. 2009, 114, 599–605. [Google Scholar] [CrossRef]
- You, H.; Liu, Y.; Agrawal, N.; Prasad, C.K.; Chiriva-Internati, M.; Lowery, C.L.; Kay, H.H.; Hermonat, P.L. Infection, replication, and cytopathology of human papillomavirus type 31 in trophoblasts. Virology 2003, 316, 281–289. [Google Scholar] [CrossRef]
- Boulenouar, S.; Weyn, C.; van Noppen, M.; Moussa Ali, M.; Favre, M.; Delvenne, P.O.; Bex, F.; Noël, A.; Englert, Y.; Fontaine, V. Effects of HPV-16 E5, E6 and E7 proteins on survival, adhesion, migration and invasion of trophoblastic cells. Carcinogenesis 2010, 31, 473–480. [Google Scholar] [CrossRef]
- Srinivas, S.K.; Ma, Y.; Sammel, M.D.; Chou, D.; McGrath, C.; Parry, S.; Elovitz, M.A. Placental inflammation and viral infection are implicated in second trimester pregnancy loss. Am. J. Obstet. Gynecol. 2006, 195, 797–802. [Google Scholar] [CrossRef]
- Zuo, Z.; Goel, S.; Carter, J.E. Association of cervical cytology and HPV DNA status during pregnancy with placental abnormalities and preterm birth. Am. J. Clin. Pathol. 2011, 136, 260–265. [Google Scholar] [CrossRef]
- Gomez, L.; Ma, Y.; Ho, C.; McGrath, C.; Nelson, D.; Parry, S. Placental infection with human papillomavirus is associated with spontaneous preterm delivery. Hum. Reprod. 2008, 23, 709–715. [Google Scholar] [CrossRef]
- He, Y.; Wu, Y.-M.; Wang, T.; Song, F.; Wang, Y.; Zhu, L. Perinatal outcomes of pregnant women with cervical intraepithelial neoplasia. Arch. Gynecol. Obstet. 2013, 288, 1237–1242. [Google Scholar] [CrossRef]
- Niyibizi, J.; Zanré, N.; Mayrand, M.-H.; Trottier, H. The association between adverse pregnancy outcomes and maternal human papillomavirus infection: A systematic review protocol. Syst. Rev. 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, L.R.; Ethur, A.B.d.M.; Hilgert, J.B.; Zanini, R.R.; Berwanger, O.; Bozzetti, M.C.; Mylius, L.C. Vertical transmission of the human papillomavirus: A systematic quantitative review. Cad. Saude Publica 2005, 21, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Sarkola, M.E.; Grénman, S.E.; Rintala, M.A.; Syrjänen, K.J.; Syrjänen, S.M. Human papillomavirus in the placenta and umbilical cord blood. Acta Obstet. Gynecol. Scand. 2008, 87, 1181–1188. [Google Scholar] [CrossRef] [PubMed]
- Tenti, P.; Zappatore, R.; Migliora, P.; Spinillo, A.; Belloni, C.; Carnevali, L. Perinatal transmission of human papillomavirus from gravidas with latent infections. Obstet. Gynecol. 1999, 93, 475–479. [Google Scholar] [PubMed]
- Tenti, P.; Zappatore, R.; Migliora, P.; Spinillo, A.; Maccarini, U.; De Benedittis, M.; Vesentini, N.; Marchitelli, G.; Silini, E.; Carnevali, L. Latent human papillomavirus infection in pregnant women at term: A case-control study. J. Infect. Dis. 1997, 176, 277–280. [Google Scholar] [CrossRef][Green Version]
- Værnesbranden, M.R.; Wiik, J.; Sjøborg, K.; Staff, A.C.; Carlsen, K.C.L.; Haugen, G.; Hedlin, G.; Hilde, K.; Nordlund, B.; Nystrand, C.F.; et al. Maternal human papillomavirus infections at mid-pregnancy and delivery in a Scandinavian mother–child cohort study. Int. J. Infect. Dis. 2021, 108, 574–581. [Google Scholar] [CrossRef]
- Smith, E.M.; Johnson, S.R.; Jiang, D.; Zaleski, S.; Lynch, C.F.; Brundage, S.; Anderson, R.D.; Turek, L.P. The association between pregnancy and human papilloma virus prevalence. Cancer Detect. Prev. 1991, 15, 397–402. [Google Scholar]
- Pandey, D.; Solleti, V.; Jain, G.; Das, A.; Shama Prasada, K.; Acharya, S.; Satyamoorthy, K. Human Papillomavirus (HPV) Infection in Early Pregnancy: Prevalence and Implications. Infect. Dis. Obstet. Gynecol. 2019, 2019, 4376902. [Google Scholar] [CrossRef]
- Luo, D.; Peng, M.; Wei, X.; Pan, D.; Xue, H.; Xu, Y.; Dong, B. Prevalence of human papillomavirus and genotype distribution in pregnant and non-pregnant Women in China. Risk Manag. Health Policy 2021, 14, 3147–3157. [Google Scholar] [CrossRef]
- Schneider, A.; Hotz, M.; Gissmann, L. Increased prevalence of human papillomaviruses in the lower genital tract of pregnant women. Int. J. Cancer 1987, 40, 198–201. [Google Scholar] [CrossRef]
- Garry, R.; Jones, R. Relationship between cervical condylomata, pregnancy and subclinical papillomavirus infection. J. Reprod. Med. 1985, 30, 393–399. [Google Scholar] [PubMed]
- Muñoz, N.; Franceschi, S.; Bosetti, C.; Moreno, V.; Herrero, R.; Smith, J.S.; Shah, K.V.; Meijer, C.J.; Bosch, F.X. Role of parity and human papillomavirus in cervical cancer: The IARC multicentric case-control study. Lancet 2002, 359, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Tekalegn, Y.; Sahiledengle, B.; Woldeyohannes, D.; Atlaw, D.; Degno, S.; Desta, F.; Bekele, K.; Aseffa, T.; Gezahegn, H.; Kene, C. High parity is associated with increased risk of cervical cancer: Systematic review and meta-analysis of case-control studies. Women’s Health 2022, 18, 17455065221075904. [Google Scholar] [CrossRef]
- Jensen, K.E.; Schmiedel, S.; Norrild, B.; Frederiksen, K.; Iftner, T.; Kjaer, S.K. Parity as a cofactor for high-grade cervical disease among women with persistent human papillomavirus infection: A 13-year follow-up. Br. J. Cancer 2013, 108, 234–239. [Google Scholar] [CrossRef] [PubMed]
- James, C.D.; Morgan, I.M.; Bristol, M.L. The Relationship between Estrogen-Related Signaling and Human Papillomavirus Positive Cancers. Pathogens 2020, 9, 403. [Google Scholar] [CrossRef] [PubMed]
- Läsche, M.; Gallwas, J.; Gründker, C. Like Brothers in Arms: How Hormonal Stimuli and Changes in the Metabolism Signaling Cooperate, Leading HPV Infection to Drive the Onset of Cervical Cancer. Int. J. Mol. Sci. 2022, 23, 5050. [Google Scholar] [CrossRef]
- Delvenne, P.; Herman, L.; Kholod, N.; Caberg, J.-H.; Herfs, M.; Boniver, J.; Jacobs, N.; Hubert, P. Role of hormone cofactors in the human papillomavirus-induced carcinogenesis of the uterine cervix. Mol. Cell. Endocrinol. 2007, 264, 1–5. [Google Scholar] [CrossRef]
- Gariglio, P.; Gutiérrez, J.; Cortés, E.; Vázquez, J. The role of retinoid deficiency and estrogens as cofactors in cervical cancer. Arch. Med. Res. 2009, 40, 449–465. [Google Scholar] [CrossRef]
- Eibye, S.; Kjær, S.K.; Nielsen, T.S.; Mellemkjær, L. Mortality Among Women With Cervical Cancer During or Shortly After a Pregnancy in Denmark 1968 to 2006. Int. J. Gynecol. Cancer 2016, 26, 951–958. [Google Scholar] [CrossRef]
- Nobbenhuis, M.A.E.; Helmerhorst, T.J.M.; van den Brule, A.J.C.; Rozendaal, L.; Bezemer, P.D.; Voorhorst, F.J.; Meijer, C.J.L.M. High-risk human papillomavirus clearance in pregnant women: Trends for lower clearance during pregnancy with a catch-up postpartum. Br. J. Cancer 2002, 87, 75–80. [Google Scholar] [CrossRef]
- Sugai, S.; Nishijima, K.; Enomoto, T. Management of Condyloma Acuminata in Pregnancy: A Review. Sex. Transm. Dis. 2021, 48, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Devi, L.T.; Pathania, K. Pregnancy with HPV associated Viral Warts. Med. J. Armed Forces India 2009, 65, 272–273. [Google Scholar] [CrossRef] [PubMed]
- Qaderi, K.; Mirmolaei, S.T.; Geranmayeh, M.; Farnam, F.; Sheikh Hasani, S. ‘Does HPV affect my fertility?’ Reproductive concerns of HPV-positive women: A qualitative study. Reprod. Health 2021, 18, 72. [Google Scholar] [CrossRef] [PubMed]
- Singhal, P.; Naswa, S.; Marfatia, Y.S. Pregnancy and sexually transmitted viral infections. Indian J. Sex. Transm. Dis. AIDS 2009, 30, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Condrat, C.E.; Filip, L.; Gherghe, M.; Cretoiu, D.; Suciu, N. Maternal HPV Infection: Effects on Pregnancy Outcome. Viruses 2021, 13, 2455. [Google Scholar] [CrossRef] [PubMed]
- Isaguliants, M.; Krasnyak, S.; Smirnova, O.; Colonna, V.; Apolikhin, O.; Buonaguro, F.M. Genetic instability and anti-HPV immune response as drivers of infertility associated with HPV infection. Infect. Agents Cancer 2021, 16, 29. [Google Scholar] [CrossRef]
- Muscianisi, F.; de Toni, L.; Giorato, G.; Carosso, A.; Foresta, C.; Garolla, A. Is HPV the Novel Target in Male Idiopathic Infertility? A Systematic Review of the Literature. Front. Endocrinol. 2021, 12, 643539. [Google Scholar] [CrossRef]
- Wiik, J.; Nilsson, S.; Kärrberg, C.; Strander, B.; Jacobsson, B.; Sengpiel, V. Associations of treated and untreated human papillomavirus infection with preterm delivery and neonatal mortality: A Swedish population-based study. PLoS Med. 2021, 18, e1003641. [Google Scholar] [CrossRef]
- Hermonat, P.L.; Han, L.; Wendel, P.J.; Quirk, J.G.; Stern, S.; Lowery, C.L.; Rechtin, T.M. Human papillomavirus is more prevalent in first trimester spontaneously aborted products of conception compared to elective specimens. Virus Genes 1997, 14, 13–17. [Google Scholar] [CrossRef]
- Bober, L.; Guzowski, G.; Moczulska, H.; Sieroszewski, P. Influence of human Papilloma Virus (hPV) infection on early pregnancy. Ginekol. Pol. 2019, 90, 72–75. [Google Scholar] [CrossRef]
- Slatter, T.L.; Hung, N.G.; Clow, W.M.; Royds, J.A.; Devenish, C.J.; Hung, N.A. A clinicopathological study of episomal papillomavirus infection of the human placenta and pregnancy complications. Mod. Pathol. 2015, 28, 1369–1382. [Google Scholar] [CrossRef] [PubMed]
- McDonnold, M.; Dunn, H.; Hester, A.; Pacheco, L.D.; Hankins, G.D.; Saade, G.R.; Costantine, M.M. High risk human papillomavirus at entry to prenatal care and risk of preeclampsia. Am. J. Obstet. Gynecol. 2014, 210, 138.e1–138.e5. [Google Scholar] [CrossRef] [PubMed]
- Joo, E.-J.; Chang, Y.; Kwon, M.-J.; Cho, A.; Cheong, H.S.; Ryu, S. High-Risk Human Papillomavirus Infection and the Risk of Cardiovascular Disease in Korean Women. Circ. Res. 2019, 124, 747–756. [Google Scholar] [CrossRef]
- Kanodia, S.; Fahey, L.M.; Kast, W.M. Mechanisms used by human papillomaviruses to escape the host immune response. Curr. Cancer Drug Targets 2007, 7, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Stanley, M. HPV—Immune response to infection and vaccination. Infect. Agents Cancer 2010, 5, 19. [Google Scholar] [CrossRef] [PubMed]
- Stanley, M. Immunobiology of HPV and HPV vaccines. Gynecol. Oncol. 2008, 109, S15–S21. [Google Scholar] [CrossRef]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef]
- Kourtis, A.P.; Read, J.S.; Jamieson, D.J. Pregnancy and infection. N. Engl. J. Med. 2014, 370, 2211–2218. [Google Scholar] [CrossRef]
- Alberca, R.W.; Pereira, N.Z.; Oliveira, L.M.D.S.; Gozzi-Silva, S.C.; Sato, M.N. Pregnancy, Viral Infection, and COVID-19. Front. Immunol. 2020, 11, 1672. [Google Scholar] [CrossRef]
- Yu, W.; Hu, X.; Cao, B. Viral Infections During Pregnancy: The Big Challenge Threatening Maternal and Fetal Health. Matern. Med. 2022, 4, 72–86. [Google Scholar] [CrossRef]
- Sappenfield, E.; Jamieson, D.J.; Kourtis, A.P. Pregnancy and susceptibility to infectious diseases. Infect. Dis. Obstet. Gynecol. 2013, 2013, 752852. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xu, L.; Sun, Y.; Wang, Z. The prevalence and risk of human papillomavirus infection in pregnant women. Epidemiol. Infect. 2014, 142, 1567–1578. [Google Scholar] [CrossRef] [PubMed]
- Cotton-Caballero, A.; Dudley, D.; Ferguson, J.; Pettit, K.; Boyle, A.J.O. Maternal human papillomavirus infection increases the risk of premature rupture of membranes [19M]. Obstet. Gynecol. 2017, 129, S137. [Google Scholar] [CrossRef]
- Faas, M.M.; de Vos, P. Uterine NK cells and macrophages in pregnancy. Placenta 2017, 56, 44–52. [Google Scholar] [CrossRef]
- Sharkey, A.M.; Xiong, S.; Kennedy, P.R.; Gardner, L.; Farrell, L.E.; Chazara, O.; Ivarsson, M.A.; Hiby, S.E.; Colucci, F.; Moffett, A. Tissue-Specific Education of Decidual NK Cells. J. Immunol. 2015, 195, 3026–3032. [Google Scholar] [CrossRef]
- Tartaglia, E.; Bordoni, V.; Oliva, A.; Vergori, A.; Girardi, E.; Antinori, A.; Agrati, C.; Liuzzi, G. T helper profile in pregnant women recovered from COVID-19. J. Reprod. Immunol. 2022, 153, 103661. [Google Scholar] [CrossRef]
- Aghaeepour, N.; Ganio, E.A.; Mcilwain, D.; Tsai, A.S.; Tingle, M.; Van Gassen, S.; Gaudilliere, D.K.; Baca, Q.; McNeil, L.; Okada, R.; et al. An immune clock of human pregnancy. Sci. Immunol. 2017, 2, eaan2946. [Google Scholar] [CrossRef]
- Derzsy, Z.; Prohászka, Z.; Rigó Jr, J.; Füst, G.; Molvarec, A. Activation of the complement system in normal pregnancy and preeclampsia. Mol. Immunol. 2010, 47, 1500–1506. [Google Scholar] [CrossRef]
- Chen, K.; Liu, J.; Liu, S.; Xia, M.; Zhang, X.; Han, D.; Jiang, Y.; Wang, C.; Cao, X. Methyltransferase SETD2-mediated methylation of STAT1 is critical for interferon antiviral activity. Cell 2017, 170, 492–506.e414. [Google Scholar] [CrossRef]
- Le Gars, M.; Seiler, C.; Kay, A.W.; Bayless, N.L.; Starosvetsky, E.; Moore, L.; Shen-Orr, S.S.; Aziz, N.; Khatri, P.; Dekker, C.L.; et al. Pregnancy-induced alterations in NK cell phenotype and function. Front. Immunol. 2019, 10, 2469. [Google Scholar] [CrossRef]
- Moffett, A.; Colucci, F. Uterine NK cells: Active regulators at the maternal-fetal interface. J. Clin. Investig. 2014, 124, 1872–1879. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Innate immunity. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef] [PubMed]
- Giannini, S.L.; Hubert, P.; Doyen, J.; Boniver, J.; Delvenne, P. Influence of the mucosal epithelium microenvironment on Langerhans cells: Implications for the development of squamous intraepithelial lesions of the cervix. Int. J. Cancer 2002, 97, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Prata, T.T.M.; Bonin, C.M.; Ferreira, A.M.T.; Padovani, C.T.J.; dos Santos Fernandes, C.E.; Machado, A.P.; Tozetti, I.A. Local immunosuppression induced by high viral load of human papillomavirus: Characterization of cellular phenotypes producing interleukin-10 in cervical neoplastic lesions. Immunology 2015, 146, 113–121. [Google Scholar] [CrossRef]
- Woodham, A.W.; Raff, A.B.; Raff, L.M.; da Silva, D.M.; Yan, L.; Skeate, J.G.; Wong, M.K.; Lin, Y.G.; Kast, W.M. Inhibition of Langerhans cell maturation by human papillomavirus type 16: A novel role for the annexin A2 heterotetramer in immune suppression. J. Immunol. 2014, 192, 4748–4757. [Google Scholar] [CrossRef] [PubMed]
- Fausch, S.C.; da Silva, D.M.; Kast, W.M. Differential uptake and cross-presentation of human papillomavirus virus-like particles by dendritic cells and Langerhans cells. Cancer Res. 2003, 63, 3478–3482. [Google Scholar]
- Lucs, A.V.; DeVoti, J.A.; Hatam, L.; Afzal, A.; Abramson, A.L.; Steinberg, B.M.; Bonagura, V.R. Immune Dysregulation in Patients Persistently Infected with Human Papillomaviruses 6 and 11. J. Clin. Med. 2015, 4, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Bonagura, V.R.; Du, Z.; Ashouri, E.; Luo, L.; Hatam, L.J.; DeVoti, J.A.; Rosenthal, D.W.; Steinberg, B.M.; Abramson, A.L.; Gjertson, D.W.; et al. Activating killer cell immunoglobulin-like receptors 3DS1 and 2DS1 protect against developing the severe form of recurrent respiratory papillomatosis. Hum. Immunol. 2010, 71, 212–219. [Google Scholar] [CrossRef]
- Para, R.; Romero, R.; Miller, D.; Panaitescu, B.; Varrey, A.; Chaiworapongsa, T.; Hassan, S.S.; Hsu, C.D.; Gomez-Lopez, N. Human β-defensin-3 participates in intra-amniotic host defense in women with labor at term, spontaneous preterm labor and intact membranes, and preterm prelabor rupture of membranes. J. Matern. Fetal Neonatal Med. 2020, 33, 4117–4132. [Google Scholar] [CrossRef]
- Varrey, A.; Romero, R.; Panaitescu, B.; Miller, D.; Chaiworapongsa, T.; Patwardhan, M.; Faro, J.; Pacora, P.; Hassan, S.S.; Hsu, C.; et al. Human β-defensin-1: A natural antimicrobial peptide present in amniotic fluid that is increased in spontaneous preterm labor with intra-amniotic infection. Am. J. Reprod. Immunol. 2018, 80, e13031. [Google Scholar] [CrossRef]
- Buck, C.B.; Day, P.M.; Thompson, C.D.; Lubkowski, J.; Lu, W.; Lowy, D.R.; Schiller, J.T. Human alpha-defensins block papillomavirus infection. Proc. Natl. Acad. Sci. USA 2006, 103, 1516–1521. [Google Scholar] [CrossRef]
- Wiens, M.E.; Smith, J.G. Alpha-defensin HD5 inhibits furin cleavage of human papillomavirus 16 L2 to block infection. J. Virol. 2015, 89, 2866–2874. [Google Scholar] [CrossRef] [PubMed]
- Wiens, M.E.; Smith, J.G. α-Defensin HD5 Inhibits Human Papillomavirus 16 Infection via Capsid Stabilization and Redirection to the Lysosome. mBio 2017, 8, e02304-16. [Google Scholar] [CrossRef] [PubMed]
- Quayle, A.J.; Porter, E.M.; Nussbaum, A.A.; Wang, Y.M.; Brabec, C.; Yip, K.P.; Mok, S.C. Gene expression, immunolocalization, and secretion of human defensin-5 in human female reproductive tract. Am. J. Pathol. 1998, 152, 1247–1258. [Google Scholar] [PubMed]
- Svinarich, D.M.; Gomez, R.; Romero, R. Detection of human defensins in the placenta. Am. J. Reprod. Immunol. 1997, 38, 252–255. [Google Scholar] [CrossRef]
- Svinarich, D.M.; Wolf, N.A.; Gomez, R.; Gonik, B.; Romero, R. Detection of human defensin 5 in reproductive tissues. Am. J. Obstet. Gynecol. 1997, 176, 470–475. [Google Scholar] [CrossRef]
- Escribese, M.M.; Rodríguez-García, M.; Sperling, R.; Engel, S.M.; Gallart, T.; Moran, T.M. Alpha-defensins 1-3 release by dendritic cells is reduced by estrogen. Reprod. Biol. Endocrinol. 2011, 9, 118. [Google Scholar] [CrossRef]
- Dorfer, S.; Strasser, K.; Reipert, S.; Fischer, M.B.; Shafti-Keramat, S.; Bonelli, M.; Schröckenfuchs, G.; Bauer, W.; Kancz, S.; Müller, L.; et al. Deficiency of Cathelicidin-related Antimicrobial Peptide Promotes Skin Papillomatosis in Mus musculus Papillomavirus 1-infected Mice. Acta Derm. Venereol. 2021, 101, adv00367. [Google Scholar] [CrossRef]
- Lee, S.; Yoo, I.; Han, J.; Ka, H. Antimicrobial peptides cathelicidin, PMAP23, and PMAP37: Expression in the endometrium throughout the estrous cycle and at the maternal-conceptus interface during pregnancy and regulation by steroid hormones and calcitriol in pigs. Theriogenology 2021, 160, 1–9. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Coyne, C.B.; Zeh, H.J.; Lotze, M.T. PAMPs and DAMPs: Signal 0s that spur autophagy and immunity. Immunol. Rev. 2012, 249, 158–175. [Google Scholar] [CrossRef]
- Chan, J.K.; Roth, J.; Oppenheim, J.J.; Tracey, K.J.; Vogl, T.; Feldmann, M.; Horwood, N.; Nanchahal, J. Alarmins: Awaiting a clinical response. J. Clin. Investig. 2012, 122, 2711–2719. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.; Huebener, P.; Schwabe, R.F. Damage-associated molecular patterns in cancer: A double-edged sword. Oncogene 2016, 35, 5931–5941. [Google Scholar] [CrossRef] [PubMed]
- Urban-Wojciuk, Z.; Khan, M.M.; Oyler, B.L.; Fåhraeus, R.; Marek-Trzonkowska, N.; Nita-Lazar, A.; Hupp, T.R.; Goodlett, D.R. The Role of TLRs in Anti-cancer Immunity and Tumor Rejection. Front. Immunol. 2019, 10, 2388. [Google Scholar] [CrossRef] [PubMed]
- Douzandeh-Mobarrez, B.; Kariminik, A.; Kazemi Arababadi, M.; Kheirkhah, B. TLR9 in the Human Papilloma Virus Infections: Friend or Foe? Viral Immunol. 2022, 35, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.T.; Cheng, A.C.; Liu, Y.T.; Yan, C.; Cheng, Y.C.; Chang, C.F.; Tseng, P.H. Persistent TLR4 Activation Promotes Hepatocellular Carcinoma Growth through Positive Feedback Regulation by LIN28A/Let-7g miRNA. Int. J. Mol. Sci. 2022, 23, 8419. [Google Scholar] [CrossRef]
- Yuan, X.; Zhou, Y.; Wang, W.; Li, J.; Xie, G.; Zhao, Y.; Xu, D.; Shen, L. Activation of TLR4 signaling promotes gastric cancer progression by inducing mitochondrial ROS production. Cell Death Dis. 2013, 4, e794. [Google Scholar] [CrossRef]
- Ou, T.; Lilly, M.; Jiang, W. The Pathologic Role of Toll-Like Receptor 4 in Prostate Cancer. Front. Immunol. 2018, 9, 1188. [Google Scholar] [CrossRef]
- Kelly, M.G.; Alvero, A.B.; Chen, R.; Silasi, D.-A.; Abrahams, V.M.; Chan, S.; Visintin, I.; Rutherford, T.; Mor, G. TLR-4 Signaling Promotes Tumor Growth and Paclitaxel Chemoresistance in Ovarian Cancer. Cancer Res. 2006, 66, 3859–3868. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhu, K.; Cheng, H. Toll-like receptors in human papillomavirus infection. Arch. Immunol. Ther. Exp. 2013, 61, 203–215. [Google Scholar] [CrossRef]
- Joyce, J.G.; Tung, J.-S.; Przysiecki, C.T.; Cook, J.C.; Lehman, E.D.; Sands, J.A.; Jansen, K.U.; Keller, P.M. The L1 major capsid protein of human papillomavirus type 11 recombinant virus-like particles interacts with heparin and cell-surface glycosaminoglycans on human keratinocytes. J. Biol. Chem. 1999, 274, 5810–5822. [Google Scholar] [CrossRef]
- Hasan, U.A.; Bates, E.; Takeshita, F.; Biliato, A.; Accardi, R.; Bouvard, V.; Mansour, M.; Vincent, I.; Gissmann, L.; Iftner, T.; et al. TLR9 expression and function is abolished by the cervical cancer-associated human papillomavirus type 16. J. Immunol. 2007, 178, 3186–3197. [Google Scholar] [CrossRef] [PubMed]
- Daud, I.I.; Scott, M.E.; Ma, Y.; Shiboski, S.; Farhat, S.; Moscicki, A.B. Association between toll-like receptor expression and human papillomavirus type 16 persistence. Int. J. Cancer 2011, 128, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.R.; Ramalho, A.C.; Marques, M.; Ribeiro, D. The Interplay between Antiviral Signalling and Carcinogenesis in Human Papillomavirus Infections. Cancers 2020, 12, 646. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Cheng, Y.; Li, C. The role of TLRs in cervical cancer with HPV infection: A review. Signal Transduct. Target. Ther. 2017, 2, 17055. [Google Scholar] [CrossRef]
- Cheng, Y.; Chen, G.; Wang, X.; Huang, Y.; Ding, J.; Huang, J.; Hong, L. TLR4 may accelerate hypoxia reaction to promote the occurrence and progress of cervical lesions by infected pathogenic microorganisms other than HPV. J. Cancer Ther. 2013, 4, 549–553. [Google Scholar] [CrossRef][Green Version]
- Castro-Muñoz, L.J.; Rocha-Zavaleta, L.; Lizano, M.; Ramírez-Alcántara, K.M.; Madrid-Marina, V.; Manzo-Merino, J. Alteration of the IFN-Pathway by Human Papillomavirus Proteins: Antiviral Immune Response Evasion Mechanism. Biomedicines 2022, 10, 2965. [Google Scholar] [CrossRef] [PubMed]
- Perea, S.; Lopez-Ocejo, O.; Von Gabain, A.; Araña, M. Human papillomavirus type-16 (HPV-16) major transforming proteins functionally interact with interferon signaling mechanisms. Int. J. Oncol. 1997, 11, 169–173. [Google Scholar] [CrossRef]
- Rice, S.; Kim, S.M.; Rodriguez, C.; Songock, W.; Raikhy, G.; Lopez, R.; Henderson, L.; Yusufji, A.; Bodily, J. Suppression of a Subset of Interferon-Induced Genes by Human Papillomavirus Type 16 E7 via a Cyclin Dependent Kinase 8-Dependent Mechanism. Viruses 2020, 12, 311. [Google Scholar] [CrossRef]
- Arany, I.; Goel, A.; Tyring, S.K. Interferon response depends on viral transcription in human papillomavirus-containing lesions. Anticancer Res. 1995, 15, 2865–2869. [Google Scholar]
- Um, S.J.; Rhyu, J.W.; Kim, E.J.; Jeon, K.C.; Hwang, E.S.; Park, J.S. Abrogation of IRF-1 response by high-risk HPV E7 protein in vivo. Cancer Lett. 2002, 179, 205–212. [Google Scholar] [CrossRef]
- Park, J.S.; Kim, E.J.; Kwon, H.J.; Hwang, E.S.; Namkoong, S.E.; Um, S.J. Inactivation of interferon regulatory factor-1 tumor suppressor protein by HPV E7 oncoprotein. Implication for the E7-mediated immune evasion mechanism in cervical carcinogenesis. J. Biol. Chem. 2000, 275, 6764–6769. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz-Fathi, M.; Rezaei, N. Chapter 3—Vaccines, Adjuvants, and Delivery Systems. In Vaccines for Cancer Immunotherapy; Rezaei, N., Keshavarz-Fathi, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 45–59. [Google Scholar]
- Pezda, A.C.; Penn, A.; Barton, G.M.; Coscoy, L. Suppression of TLR9 immunostimulatory motifs in the genome of a gammaherpesvirus. J. Immunol. 2011, 187, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Hasan, U.A.; Zannetti, C.; Parroche, P.; Goutagny, N.; Malfroy, M.; Roblot, G.; Carreira, C.; Hussain, I.; Müller, M.; Taylor-Papadimitriou, J.; et al. The human papillomavirus type 16 E7 oncoprotein induces a transcriptional repressor complex on the Toll-like receptor 9 promoter. J. Exp. Med. 2013, 210, 1369–1387. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Luquez, K.; Schadock, I.C.; Gonçalves, C.V.; Tornatore, M.; Finger-Jardim, F.; Avila, E.C.; Soares, M.A.; de Martínez, A.M.B.; Ellwanger, J.H.; Chies, J.A.B.; et al. Impact of TLR7 and TLR9 polymorphisms on susceptibility to placental infections and pregnancy complications. J. Reprod. Immunol. 2021, 146, 103342. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, V.M.; Schaefer, T.M.; Fahey, J.V.; Visintin, I.; Wright, J.A.; Aldo, P.B.; Romero, R.; Wira, C.R.; Mor, G. Expression and secretion of antiviral factors by trophoblast cells following stimulation by the TLR-3 agonist, Poly(I:C). Hum. Reprod. 2006, 21, 2432–2439. [Google Scholar] [CrossRef] [PubMed]
- Koga, K.; Mor, G. Toll-like receptors at the maternal-fetal interface in normal pregnancy and pregnancy disorders. Am. J. Reprod. Immunol. 2010, 63, 587–600. [Google Scholar] [CrossRef]
- Koga, K.; Cardenas, I.; Aldo, P.; Abrahams, V.M.; Peng, B.; Fill, S.; Romero, R.; Mor, G. Activation of TLR3 in the trophoblast is associated with preterm delivery. Am. J. Reprod. Immunol. 2009, 61, 196–212. [Google Scholar] [CrossRef]
- Rehwinkel, J.; Gack, M.U. RIG-I-like receptors: Their regulation and roles in RNA sensing. Nat. Rev. Immunol. 2020, 20, 537–551. [Google Scholar] [CrossRef]
- Li, P.; Zhang, X.; Cao, W.; Yang, F.; Du, X.; Shi, Z.; Zhang, M.; Liu, X.; Zhu, Z.; Zheng, H. RIG-I is responsible for activation of type I interferon pathway in Seneca Valley virus-infected porcine cells to suppress viral replication. Virol. J. 2018, 15, 162. [Google Scholar] [CrossRef]
- Chiang, C.; Pauli, E.K.; Biryukov, J.; Feister, K.F.; Meng, M.; White, E.A.; Münger, K.; Howley, P.M.; Meyers, C.; Gack, M.U. The Human Papillomavirus E6 Oncoprotein Targets USP15 and TRIM25 To Suppress RIG-I-Mediated Innate Immune Signaling. J. Virol. 2018, 92, e01737-17. [Google Scholar] [CrossRef]
- Ishikawa, H.; Barber, G.N. STING is an endoplasmic reticulum adaptor that facilitates innate immune signalling. Nature 2008, 455, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Ma, Z.; Barber, G.N. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 2009, 461, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Liu, P. Cytosolic DNA sensing by cGAS: Regulation, function, and human diseases. Signal Transduct. Target. Ther. 2021, 6, 170. [Google Scholar] [CrossRef] [PubMed]
- Gui, X.; Yang, H.; Li, T.; Tan, X.; Shi, P.; Li, M.; Du, F.; Chen, Z.J. Autophagy induction via STING trafficking is a primordial function of the cGAS pathway. Nature 2019, 567, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Sun, H.; You, F.; Sun, W.; Zhou, X.; Chen, L.; Yang, J.; Wang, Y.; Tang, H.; Guan, Y.; et al. Activation of STAT6 by STING is critical for antiviral innate immunity. Cell 2011, 147, 436–446. [Google Scholar] [CrossRef]
- Lau, L.; Gray, E.E.; Brunette, R.L.; Stetson, D.B. DNA tumor virus oncogenes antagonize the cGAS-STING DNA-sensing pathway. Science 2015, 350, 568–571. [Google Scholar] [CrossRef]
- Delorme-Axford, E.; Klionsky, D.J. Inflammatory-dependent Sting activation induces antiviral autophagy to limit zika virus in the Drosophila brain. Autophagy 2019, 15, 1–3. [Google Scholar] [CrossRef]
- Guess, J.C.; McCance, D.J. Decreased migration of Langerhans precursor-like cells in response to human keratinocytes expressing human papillomavirus type 16 E6/E7 is related to reduced macrophage inflammatory protein-3α production. J. Virol. 2005, 79, 14852–14862. [Google Scholar] [CrossRef]
- Kindt, N.; Descamps, G.; Lechien, J.R.; Remmelink, M.; Colet, J.M.; Wattiez, R.; Berchem, G.; Journe, F.; Saussez, S. Involvement of HPV Infection in the Release of Macrophage Migration Inhibitory Factor in Head and Neck Squamous Cell Carcinoma. J. Clin. Med. 2019, 8, 75. [Google Scholar] [CrossRef]
- Mazibrada, J.; Rittà, M.; Mondini, M.; De Andrea, M.; Azzimonti, B.; Borgogna, C.; Ciotti, M.; Orlando, A.; Surico, N.; Chiusa, L.; et al. Interaction between inflammation and angiogenesis during different stages of cervical carcinogenesis. Gynecol. Oncol. 2008, 108, 112–120. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, Z.; Gao, S.; Li, C.; Feng, Y.; Zhou, X. Tumor-Associated Macrophages: Recent Insights and Therapies. Front. Oncol. 2020, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xie, Y.; Wang, X.; Jiang, C.; Yuan, X.; Zhang, A.; Liu, C.; Pang, L.; Li, F.; Hu, J. Overexpression of VEGF-C and MMP-9 predicts poor prognosis in Kazakh patients with esophageal squamous cell carcinoma. PeerJ 2019, 7, e8182. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, L.; Li, Y.; Zhao, X. Research Progress on Tumor-Associated Macrophages and Inflammation in Cervical Cancer. BioMed Res. Int. 2020, 2020, 6842963. [Google Scholar] [CrossRef] [PubMed]
- Lurie, S.; Rahamim, E.; Piper, I.; Golan, A.; Sadan, O. Total and differential leukocyte counts percentiles in normal pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 136, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Xu, X.-H.; Jin, L. Macrophage polarization in physiological and pathological pregnancy. Front. Immunol. 2019, 10, 792. [Google Scholar] [CrossRef] [PubMed]
- Ambühl, L.M.M.; Leonhard, A.K.; Widen Zakhary, C.; Jørgensen, A.; Blaakaer, J.; Dybkaer, K.; Baandrup, U.; Uldbjerg, N.; Sørensen, S. Human papillomavirus infects placental trophoblast and Hofbauer cells, but appears not to play a causal role in miscarriage and preterm labor. Acta Obstet. Gynecol. Scand. 2017, 96, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Cho, Y.S.; Cho, M.C.; Shim, J.H.; Lee, K.A.; Ko, K.K.; Choe, Y.K.; Park, S.N.; Hoshino, T.; Kim, S.; et al. Both E6 and E7 oncoproteins of human papillomavirus 16 inhibit IL-18-induced IFN-gamma production in human peripheral blood mononuclear and NK cells. J. Immunol. 2001, 167, 497–504. [Google Scholar] [CrossRef]
- Senju, H.; Kumagai, A.; Nakamura, Y.; Yamaguchi, H.; Nakatomi, K.; Fukami, S.; Shiraishi, K.; Harada, Y.; Nakamura, M.; Okamura, H.; et al. Effect of IL-18 on the Expansion and Phenotype of Human Natural Killer Cells: Application to Cancer Immunotherapy. Int. J. Biol. Sci. 2018, 14, 331–340. [Google Scholar] [CrossRef]
- Song, Y.; Wu, X.; Xu, Y.; Zhu, J.; Li, J.; Zou, Z.; Chen, L.; Zhang, B.; Hua, C.; Rui, H.; et al. HPV E7 inhibits cell pyroptosis by promoting TRIM21-mediated degradation and ubiquitination of the IFI16 inflammasome. Int. J. Biol. Sci. 2020, 16, 2924–2937. [Google Scholar] [CrossRef]
- Gutiérrez-Hoya, A.; Soto-Cruz, I. NK Cell Regulation in Cervical Cancer and Strategies for Immunotherapy. Cells 2021, 10, 3104. [Google Scholar] [CrossRef]
- Koopman, L.A.; Kopcow, H.D.; Rybalov, B.; Boyson, J.E.; Orange, J.S.; Schatz, F.; Masch, R.; Lockwood, C.J.; Schachter, A.D.; Park, P.J.; et al. Human decidual natural killer cells are a unique NK cell subset with immunomodulatory potential. J. Exp. Med. 2003, 198, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Kopcow, H.D.; Allan, D.S.; Chen, X.; Rybalov, B.; Andzelm, M.M.; Ge, B.; Strominger, J.L. Human decidual NK cells form immature activating synapses and are not cytotoxic. Proc. Natl. Acad. Sci. USA 2005, 102, 15563–15568. [Google Scholar] [CrossRef] [PubMed]
- Siewiera, J.; El Costa, H.; Tabiasco, J.; Berrebi, A.; Cartron, G.; le Bouteiller, P.; Jabrane-Ferrat, N. Human cytomegalovirus infection elicits new decidual natural killer cell effector functions. PLoS Pathog. 2013, 9, e1003257. [Google Scholar] [CrossRef]
- Crespo, Â.C.; Strominger, J.L.; Tilburgs, T. Expression of KIR2DS1 by decidual natural killer cells increases their ability to control placental HCMV infection. Proc. Natl. Acad. Sci. USA 2016, 113, 15072–15077. [Google Scholar] [CrossRef] [PubMed]
- Sanchita Roy, P.; Sutapa, M.; Dipanwita, G.; Pranab Kumar, S.; Sinjini, S.; Ranita, P.; Vilas, D.N. Human Papillomavirus Infections in Pregnant Women and Its Impact on Pregnancy Outcomes: Possible Mechanism of Self-Clearance. In Human Papillomavirus; Rajamanickam, R., Ed.; IntechOpen: Rijeka, Croatia, 2020; p. Ch.4. [Google Scholar]
- Deligeoroglou, E.; Giannouli, A.; Athanasopoulos, N.; Karountzos, V.; Vatopoulou, A.; Dimopoulos, K.; Creatsas, G. HPV infection: Immunological aspects and their utility in future therapy. Infect. Dis. Obstet. Gynecol. 2013, 2013, 540850. [Google Scholar] [CrossRef] [PubMed]
- Viscidi, R.P.; Schiffman, M.; Hildesheim, A.; Herrero, R.; Castle, P.E.; Bratti, M.C.; Rodriguez, A.C.; Sherman, M.E.; Wang, S.; Clayman, B.; et al. Seroreactivity to human papillomavirus (HPV) types 16, 18, or 31 and risk of subsequent HPV infection: Results from a population-based study in Costa Rica. Cancer Epidemiol. Biomark. Prev. 2004, 13, 324–327. [Google Scholar] [CrossRef]
- van der Burg, S.H.; de Jong, A.; Welters, M.J.; Offringa, R.; Melief, C.J. The status of HPV16-specific T-cell reactivity in health and disease as a guide to HPV vaccine development. Virus Res. 2002, 89, 275–284. [Google Scholar] [CrossRef]
- Porras, C.; Bennett, C.; Safaeian, M.; Coseo, S.; Rodríguez, A.C.; González, P.; Hutchinson, M.; Jiménez, S.; Sherman, M.E.; Wacholder, S.; et al. Determinants of seropositivity among HPV-16/18 DNA positive young women. BMC Infect. Dis. 2010, 10, 238. [Google Scholar] [CrossRef]
- Faust, H.; Jelen, M.M.; Poljak, M.; Klavs, I.; Učakar, V.; Dillner, J. Serum antibodies to human papillomavirus (HPV) pseudovirions correlate with natural infection for 13 genital HPV types. J. Clin. Virol. 2013, 56, 336–341. [Google Scholar] [CrossRef]
- Coseo, S.; Porras, C.; Hildesheim, A.; Rodriguez, A.C.; Schiffman, M.; Herrero, R.; Wacholder, S.; Gonzalez, P.; Wang, S.S.; Sherman, M.E.; et al. Seroprevalence and correlates of human papillomavirus 16/18 seropositivity among young women in Costa Rica. Sex. Transm. Dis. 2010, 37, 706–714. [Google Scholar] [CrossRef]
- Cho, K.J.; Roche, P.A. Regulation of MHC Class II-Peptide Complex Expression by Ubiquitination. Front. Immunol. 2013, 4, 369. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; Salnikov, M.; Tessier, T.M.; Mymryk, J.S. Reduced MHC Class I and II Expression in HPV—Negative vs. HPV—Positive Cervical Cancers. Cells 2022, 11, 3911. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; Salnikov, M.; Gameiro, S.F.; Maleki Vareki, S.; Mymryk, J.S. HPV-Positive and-Negative Cervical Cancers Are Immunologically Distinct. J. Clin. Med. 2022, 11, 4825. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, S.F.; Zhang, A.; Ghasemi, F.; Barrett, J.W.; Nichols, A.C.; Mymryk, J.S. Analysis of Class I Major Histocompatibility Complex Gene Transcription in Human Tumors Caused by Human Papillomavirus Infection. Viruses 2017, 9, 252. [Google Scholar] [CrossRef]
- Heller, C.; Weisser, T.; Mueller-Schickert, A.; Rufer, E.; Hoh, A.; Leonhardt, R.M.; Knittler, M.R. Identification of key amino acid residues that determine the ability of high risk HPV16-E7 to dysregulate major histocompatibility complex class I expression. J. Biol. Chem. 2011, 286, 10983–10997. [Google Scholar] [CrossRef]
- Li, W.; Deng, X.M.; Wang, C.X.; Zhang, X.; Zheng, G.X.; Zhang, J.; Feng, J.B. Down-regulation of HLA class I antigen in human papillomavirus type 16 E7 expressing HaCaT cells: Correlate with TAP-1 expression. Int. J. Gynecol. Cancer 2010, 20, 227–232. [Google Scholar] [CrossRef]
- Liu, D.W.; Tsao, Y.P.; Hsieh, C.H.; Hsieh, J.T.; Kung, J.T.; Chiang, C.L.; Huang, S.J.; Chen, S.L. Induction of CD8 T cells by vaccination with recombinant adenovirus expressing human papillomavirus type 16 E5 gene reduces tumor growth. J. Virol. 2000, 74, 9083–9089. [Google Scholar] [CrossRef]
- Campo, M.; Graham, S.; Cortese, M.; Ashrafi, G.; Araibi, E.; Dornan, E.; Miners, K.; Nunes, C.; Man, S. HPV-16 E5 down-regulates expression of surface HLA class I and reduces recognition by CD8 T cells. Virology 2010, 407, 137–142. [Google Scholar] [CrossRef]
- Einstein, M.H.; Leanza, S.; Chiu, L.G.; Schlecht, N.F.; Goldberg, G.L.; Steinberg, B.M.; Burk, R.D. Genetic variants in TAP are associated with high-grade cervical neoplasia. Clin. Cancer Res. 2009, 15, 1019–1023. [Google Scholar] [CrossRef]
- Stelzle, D.; Tanaka, L.F.; Lee, K.K.; Ibrahim Khalil, A.; Baussano, I.; Shah, A.S.V.; McAllister, D.A.; Gottlieb, S.L.; Klug, S.J.; Winkler, A.S.; et al. Estimates of the global burden of cervical cancer associated with HIV. Lancet Glob. Health 2021, 9, e161–e169. [Google Scholar] [CrossRef]
- Lekoane, K.M.B.; Kuupiel, D.; Mashamba-Thompson, T.P.; Ginindza, T.G. The interplay of HIV and human papillomavirus-related cancers in sub-Saharan Africa: Scoping review. Syst. Rev. 2020, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Mbuya, W.; McHaro, R.; Mhizde, J.; Mnkai, J.; Mahenge, A.; Mwakatima, M.; Mwalongo, W.; Chiwerengo, N.; Hölscher, M.; Lennemann, T.; et al. Depletion and activation of mucosal CD4 T cells in HIV infected women with HPV-associated lesions of the cervix uteri. PLoS ONE 2020, 15, e0240154. [Google Scholar] [CrossRef] [PubMed]
- Mbulawa, Z.Z.; Marais, D.J.; Johnson, L.F.; Boulle, A.; Coetzee, D.; Williamson, A.-L. Influence of human immunodeficiency virus and CD4 count on the prevalence of human papillomavirus in heterosexual couples. J. Gen. Virol. 2010, 91, 3023–3031. [Google Scholar] [CrossRef] [PubMed]
- Morrow, M.P.; Kraynyak, K.A.; Sylvester, A.J.; Dallas, M.; Knoblock, D.; Boyer, J.D.; Yan, J.; Vang, R.; Khan, A.S.; Humeau, L.; et al. Clinical and Immunologic Biomarkers for Histologic Regression of High-Grade Cervical Dysplasia and Clearance of HPV16 and HPV18 after Immunotherapy. Clin. Cancer Res. 2018, 24, 276–294. [Google Scholar] [CrossRef]
- Pennock, N.D.; White, J.T.; Cross, E.W.; Cheney, E.E.; Tamburini, B.A.; Kedl, R.M. T cell responses: Naive to memory and everything in between. Adv. Physiol. Educ. 2013, 37, 273–283. [Google Scholar] [CrossRef]
- Zhu, J.; Yamane, H.; Paul, W.E. Differentiation of effector CD4 T cell populations. Annu. Rev. Immunol. 2010, 28, 445–489. [Google Scholar] [CrossRef]
- Kobayashi, A.; Weinberg, V.; Darragh, T.; Smith-McCune, K. Evolving immunosuppressive microenvironment during human cervical carcinogenesis. Mucosal Immunol. 2008, 1, 412–420. [Google Scholar] [CrossRef]
- Peghini, B.C.; Abdalla, D.R.; Barcelos, A.C.; Teodoro, L.; Murta, E.F.; Michelin, M.A. Local cytokine profiles of patients with cervical intraepithelial and invasive neoplasia. Hum. Immunol. 2012, 73, 920–926. [Google Scholar] [CrossRef]
- Suominen, H.; Paaso, A.; Koskimaa, H.-M.; Grénman, S.; Syrjänen, K.; Syrjänen, S.; Louvanto, K. Peripheral Blood T-lymphocyte Phenotypes in Mother-Child Pairs Stratified by the Maternal HPV Status: Persistent HPV16 vs. HPV-Negative: A Case-Control Study. Viruses 2022, 14, 2633. [Google Scholar] [CrossRef]
- Rodríguez, A.C.; García-Piñeres, A.J.; Hildesheim, A.; Herrero, R.; Trivett, M.; Williams, M.; Atmella, I.; Ramírez, M.; Villegas, M.; Schiffman, M.; et al. Alterations of T-cell surface markers in older women with persistent human papillomavirus infection. Int. J. Cancer 2011, 128, 597–607. [Google Scholar] [CrossRef]
- Rincon-Orozco, B.; Halec, G.; Rosenberger, S.; Muschik, D.; Nindl, I.; Bachmann, A.; Ritter, T.M.; Dondog, B.; Ly, R.; Bosch, F.X.; et al. Epigenetic silencing of interferon-kappa in human papillomavirus type 16-positive cells. Cancer Res. 2009, 69, 8718–8725. [Google Scholar] [CrossRef] [PubMed]
- Mougiakakos, D.; Choudhury, A.; Lladser, A.; Kiessling, R.; Johansson, C.C. Regulatory T cells in cancer. Adv. Cancer Res. 2010, 107, 57–117. [Google Scholar] [CrossRef]
- Cedeno-Laurent, F.; Dimitroff, C.J. Galectin-1 research in T cell immunity: Past, present and future. Clin. Immunol. 2012, 142, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Ao, C.; Zeng, K. The role of regulatory T cells in pathogenesis and therapy of human papillomavirus-related diseases, especially in cancer. Infect. Genet. Evol. 2018, 65, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Zenclussen, A.C. Regulatory T cells in pregnancy. Springer Semin. Immunopathol. 2006, 28, 31–39. [Google Scholar] [CrossRef]
- Jørgensen, N.; Persson, G.; Hviid, T.V.F. The Tolerogenic Function of Regulatory T Cells in Pregnancy and Cancer. Front. Immunol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Jash, S.; Sharma, S. In utero immune programming of autism spectrum disorder (ASD). Hum. Immunol. 2021, 82, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Kamolratanakul, S.; Pitisuttithum, P. Human Papillomavirus Vaccine Efficacy and Effectiveness against Cancer. Vaccines 2021, 9, 1413. [Google Scholar] [CrossRef]
- Yuan, H.; Estes, P.A.; Chen, Y.; Newsome, J.; Olcese, V.A.; Garcea, R.L.; Schlegel, R. Immunization with a pentameric L1 fusion protein protects against papillomavirus infection. J. Virol. 2001, 75, 7848–7853. [Google Scholar] [CrossRef]
- Balasundaram, P.; Sakr, M. Understanding and Application of CDC Immunization Guidelines. In StatPearls; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Kreimer, A.R.; Sampson, J.N.; Porras, C.; Schiller, J.T.; Kemp, T.; Herrero, R.; Wagner, S.; Boland, J.; Schussler, J.; Lowy, D.R.; et al. Evaluation of Durability of a Single Dose of the Bivalent HPV Vaccine: The CVT Trial. J. Natl. Cancer Inst. 2020, 112, 1038–1046. [Google Scholar] [CrossRef]
- Baandrup, L.; Dehlendorff, C.; Kjaer, S.K. One-dose human papillomavirus vaccination and the risk of genital warts: A Danish nationwide population-based study. Clin. Infect. Dis. 2021, 73, e3220–e3226. [Google Scholar] [CrossRef] [PubMed]
- Barnabas, R.V.; Brown, E.R.; Onono, M.A.; Bukusi, E.A.; Njoroge, B.; Winer, R.L.; Galloway, D.A.; Pinder, L.F.; Donnell, D.; Wakhungu, I.; et al. Efficacy of single-dose HPV vaccination among young African women. NEJM Evid. 2022, 1, EVIDoa2100056. [Google Scholar] [CrossRef] [PubMed]
- Batmunkh, T.; Dalmau, M.T.; Munkhsaikhan, M.E.; Khorolsuren, T.; Namjil, N.; Surenjav, U.; Toh, Z.Q.; Licciardi, P.V.; Russell, F.M.; Garland, S.M.; et al. A single dose of quadrivalent human papillomavirus (HPV) vaccine is immunogenic and reduces HPV detection rates in young women in Mongolia, six years after vaccination. Vaccine 2020, 38, 4316–4324. [Google Scholar] [CrossRef] [PubMed]
- Narducci, A.; Einarson, A.; Bozzo, P. Human papillomavirus vaccine and pregnancy. Can. Fam. Physician 2012, 58, 268–269. [Google Scholar] [PubMed]
- Garland, S.M.; Ault, K.A.; Gall, S.A.; Paavonen, J.; Sings, H.L.; Ciprero, K.L.; Saah, A.; Marino, D.; Ryan, D.; Radley, D.; et al. Pregnancy and infant outcomes in the clinical trials of a human papillomavirus type 6/11/16/18 vaccine: A combined analysis of five randomized controlled trials. Obstet. Gynecol. 2009, 114, 1179–1188. [Google Scholar] [CrossRef]
- Capra, G.; Giovannelli, L.; Matranga, D.; Bellavia, C.; Guarneri, M.F.; Fasciana, T.; Scaduto, G.; Firenze, A.; Vassiliadis, A.; Perino, A. Potential impact of a nonavalent HPV vaccine on HPV related low-and high-grade cervical intraepithelial lesions: A referral hospital-based study in Sicily. Hum. Vaccines Immunother. 2017, 13, 1839–1843. [Google Scholar] [CrossRef]
- Illah, O.; Olaitan, A. Updates on HPV Vaccination. Diagnostics 2023, 13, 243. [Google Scholar] [CrossRef]
- Bosco, L.; Serra, N.; Fasciana, T.; Pistoia, D.; Vella, M.; Di Gregorio, L.; Schillaci, R.; Perino, A.; Calagna, G.; Firenze, A.; et al. Potential impact of a nonavalent anti HPV vaccine in Italian men with and without clinical manifestations. Sci. Rep. 2021, 11, 4096. [Google Scholar] [CrossRef]
- Foresta, C.; Garolla, A.; Parisi, S.; Ghezzi, M.; Bertoldo, A.; di Nisio, A.; de Toni, L. HPV prophylactic vaccination in males improves the clearance of semen infection. EBioMedicine 2015, 2, 1487–1493. [Google Scholar] [CrossRef]
- Alli, B.Y.; Madathil, S.; Tran, S.D.; Nicolau, B. Protocol: Carrageenan for the prevention of oral HPV infection—A feasibility randomised clinical trial. BMJ Open 2023, 13, e074498. [Google Scholar] [CrossRef]
- Laurie, C.; El-Zein, M.; Tota, J.; Khosrow-Khavar, F.; Tellier, P.P.; Coutlée, F.; de Pokomandy, A.; Franco, E.L. Efficacy of a Carrageenan Gel in Increasing Clearance of Anal Human Papillomavirus Infections in Men: Interim Analysis of a Double-Blind, Randomized Controlled Trial. J. Infect. Dis. 2023, 227, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Calagna, G.; Maranto, M.; Paola, C.; Capra, G.; Perino, A.; Chiantera, V.; Cucinella, G. ‘Secondary prevention’ against female HPV infection: Literature review of the role of carrageenan. Expert Rev. Anti Infect. Ther. 2020, 18, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Magnan, S.; Tota, J.E.; El-Zein, M.; Burchell, A.N.; Schiller, J.T.; Ferenczy, A.; Tellier, P.P.; Coutlée, F.; Franco, E.L. Efficacy of a Carrageenan gel Against Transmission of Cervical HPV (CATCH): Interim analysis of a randomized, double-blind, placebo-controlled, phase 2B trial. Clin. Microbiol. Infect. 2019, 25, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Kreimer, A.R.; Cernuschi, T.; Rees, H.; Brotherton, J.M.L.; Porras, C.; Schiller, J. Public health opportunities resulting from sufficient HPV vaccine supply and a single-dose vaccination schedule. J. Natl. Cancer Inst. 2023, 115, 246–249. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Source | ||
|---|---|---|
| External Factors | ||
| Lifestyle habits | Cigarette smoking Alcohol consumption Early sexual activity Multiple sexual partners or having a partner with multiple sexual partners Engaging in sex trading Not using a barrier contraceptive method | [49] [50] [51] [52,53,54] |
| Medication | Long-term use of oral contraceptives Immunosuppressants Not having been vaccinated against HPV | [55] [56] [57] |
| Internal Factors | ||
| Co-infections | HIV and/or other sexually transmitted infections Other HR-HPV genotypes | [58] |
| Host genetic risk factors | KLF12 gene CTNND2 gene DAP gene | [59] |
| Defective immune response | Inflammasome genetics Allelic variations of the HLA locus | [48] [60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Condrat, C.E.; Cretoiu, D.; Radoi, V.E.; Mihele, D.M.; Tovaru, M.; Bordea, C.I.; Voinea, S.C.; Suciu, N. Unraveling Immunological Dynamics: HPV Infection in Women—Insights from Pregnancy. Viruses 2023, 15, 2011. https://doi.org/10.3390/v15102011
Condrat CE, Cretoiu D, Radoi VE, Mihele DM, Tovaru M, Bordea CI, Voinea SC, Suciu N. Unraveling Immunological Dynamics: HPV Infection in Women—Insights from Pregnancy. Viruses. 2023; 15(10):2011. https://doi.org/10.3390/v15102011
Chicago/Turabian StyleCondrat, Carmen Elena, Dragos Cretoiu, Viorica Elena Radoi, Dana Mihaela Mihele, Mihaela Tovaru, Cristian Ioan Bordea, Silviu Cristian Voinea, and Nicolae Suciu. 2023. "Unraveling Immunological Dynamics: HPV Infection in Women—Insights from Pregnancy" Viruses 15, no. 10: 2011. https://doi.org/10.3390/v15102011
APA StyleCondrat, C. E., Cretoiu, D., Radoi, V. E., Mihele, D. M., Tovaru, M., Bordea, C. I., Voinea, S. C., & Suciu, N. (2023). Unraveling Immunological Dynamics: HPV Infection in Women—Insights from Pregnancy. Viruses, 15(10), 2011. https://doi.org/10.3390/v15102011