

Evaluation of Histone Demethylase Inhibitor ML324 and Acyclovir against Cyprinid herpesvirus 3 Infection
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses, Cells, and Histone Demethylase Inhibitors
2.2. CyHV-3 Plaque Reduction Assay Following Drug Treatment
2.3. Histone Demethylase Inhibitor Cytotoxicity Assay In Vitro
2.4. Viral Gene Transcription in the Presence and Absence of Histone Demethylase Inhibitor
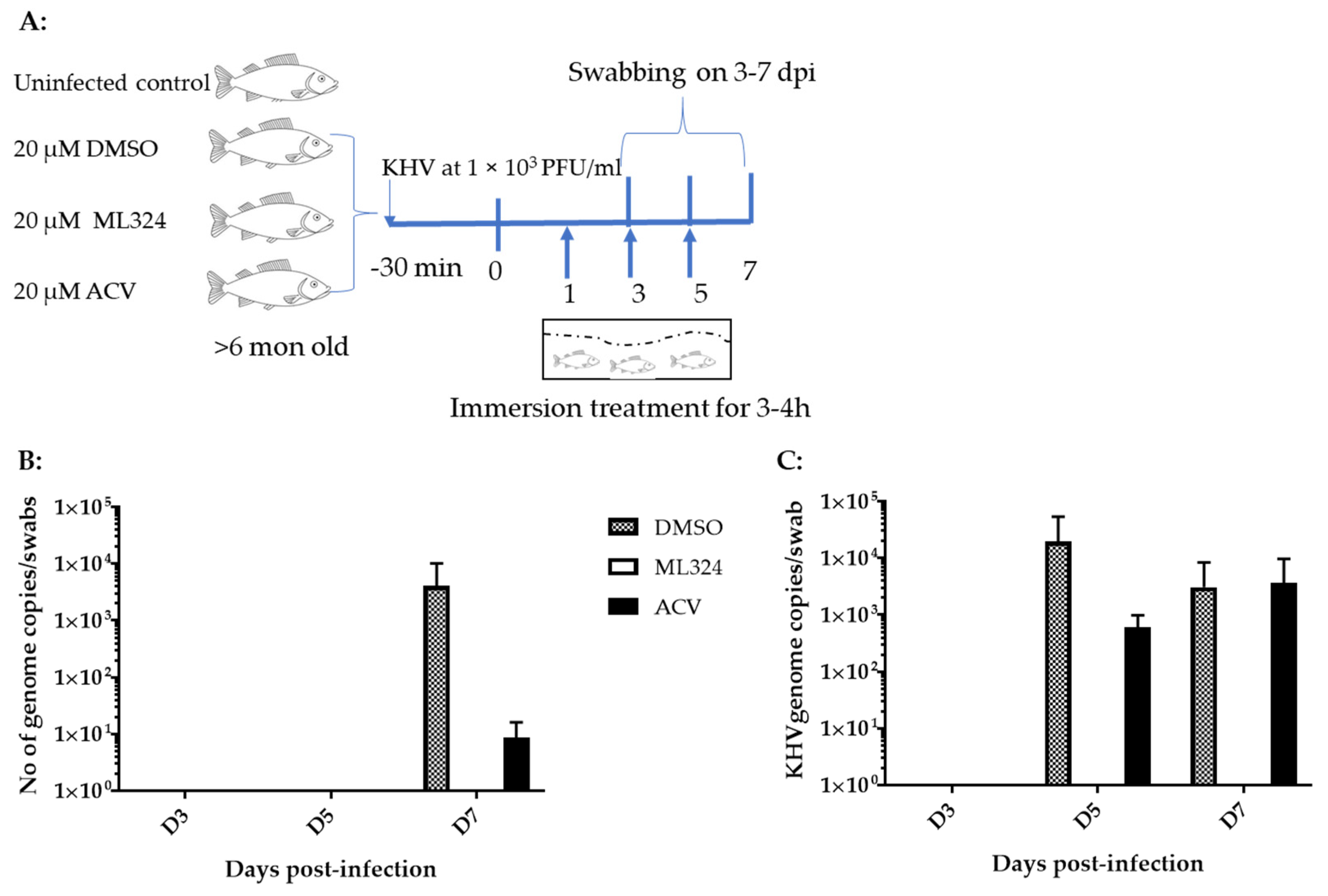
2.5. Koi Infection and Drug Treatment
2.6. Examination of CyHV-3 Infection in Body Secretions
2.7. Tissue Collection for Histopathology and Total DNA Isolation
2.8. CyHV-3 DNA Real-Time PCR
2.9. Statistical Analysis
3. Results
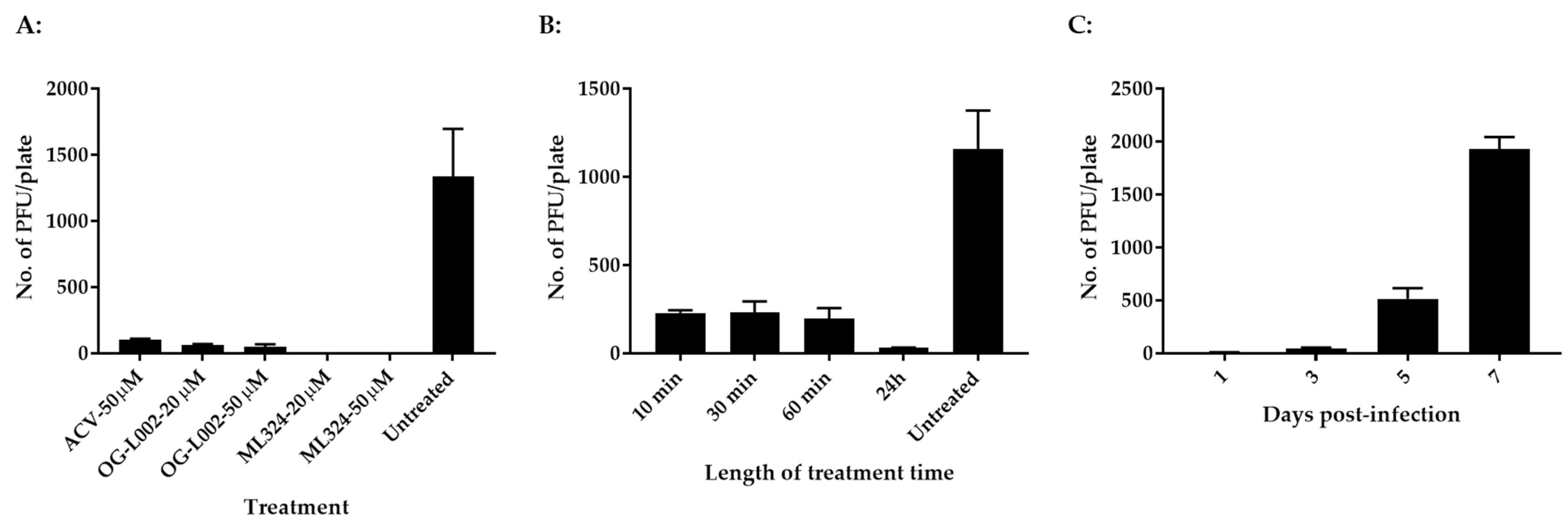
3.1. Histone Demethylase Inhibitors Reduce CyHV-3 Replication In Vitro
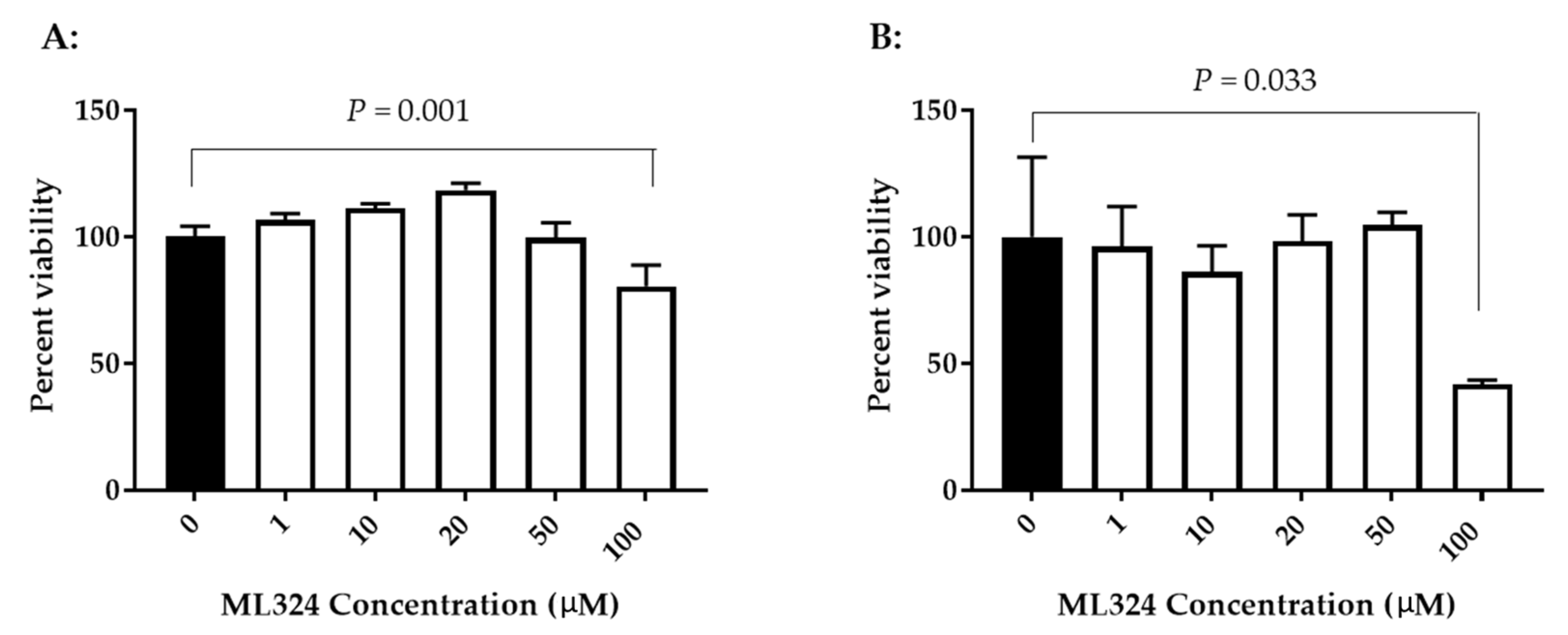
3.2. Cytotoxicity of Histone Demethylase Inhibitor ML324
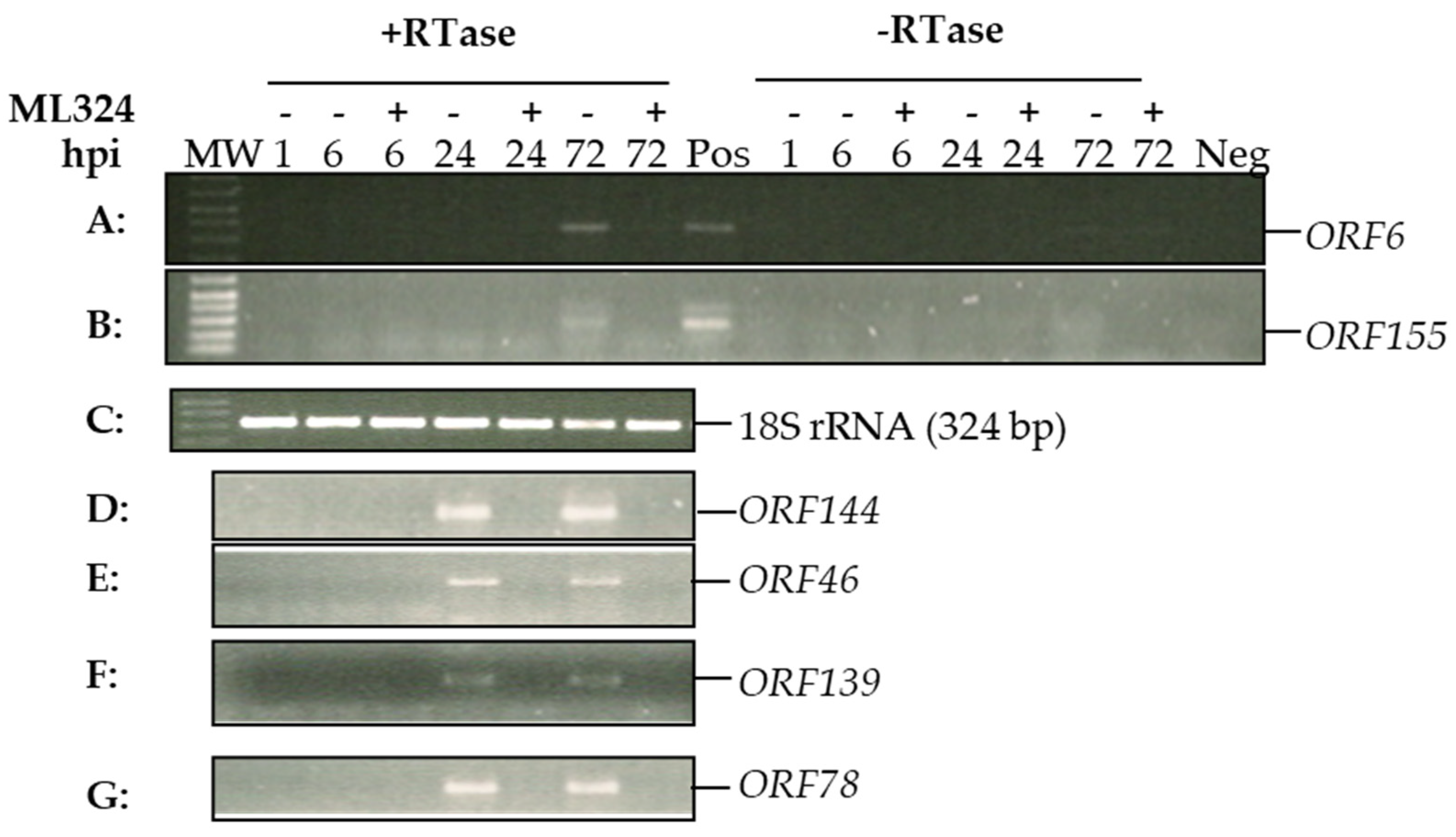
3.3. Histone Demethylase Inhibitor ML324 Blocks Transcription of CyHV-3 Immediate Early Genes
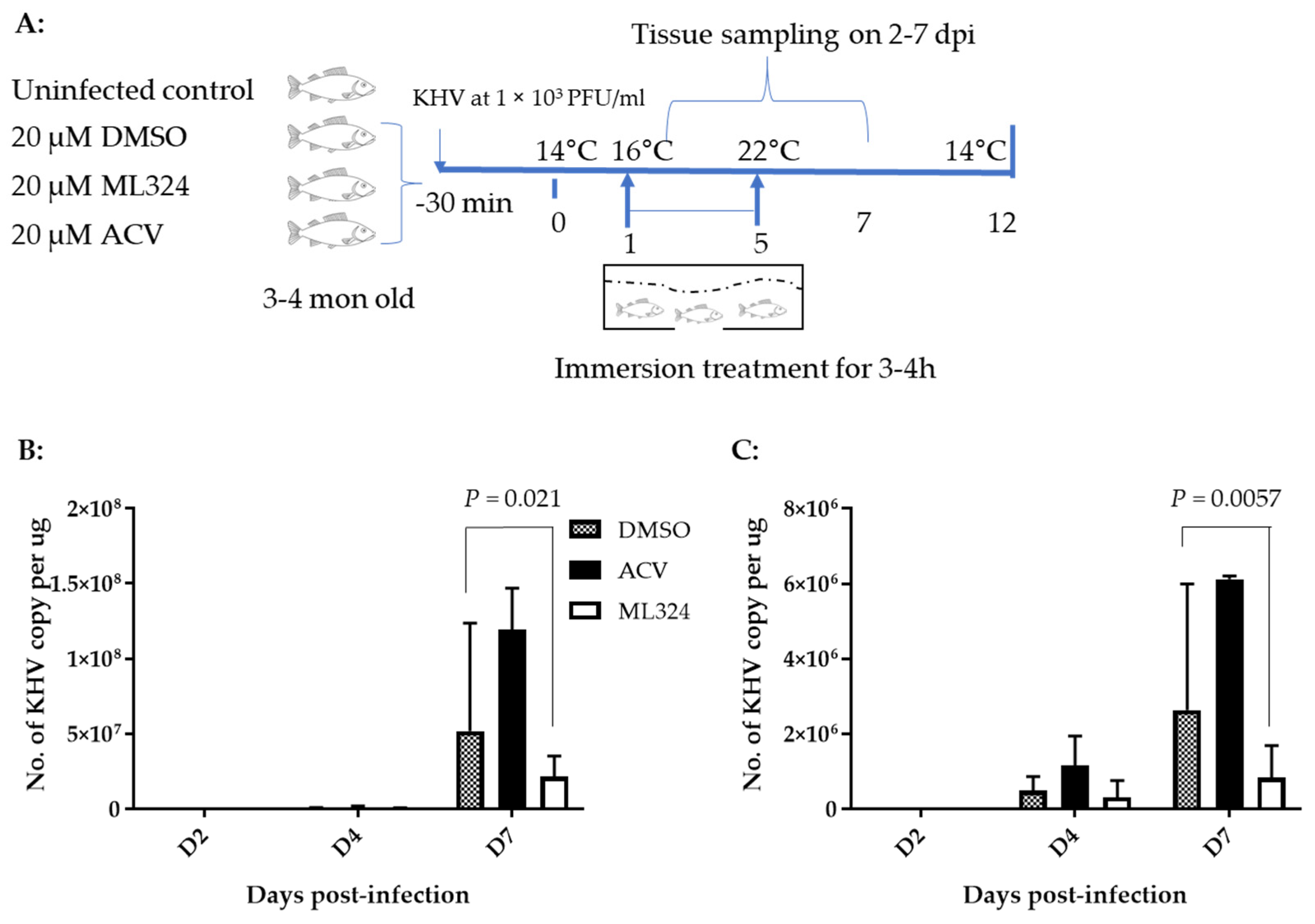
3.4. ML324 Reduced CyHV-3 Shedding in over 6-Month-Old Koi at Low Temperature
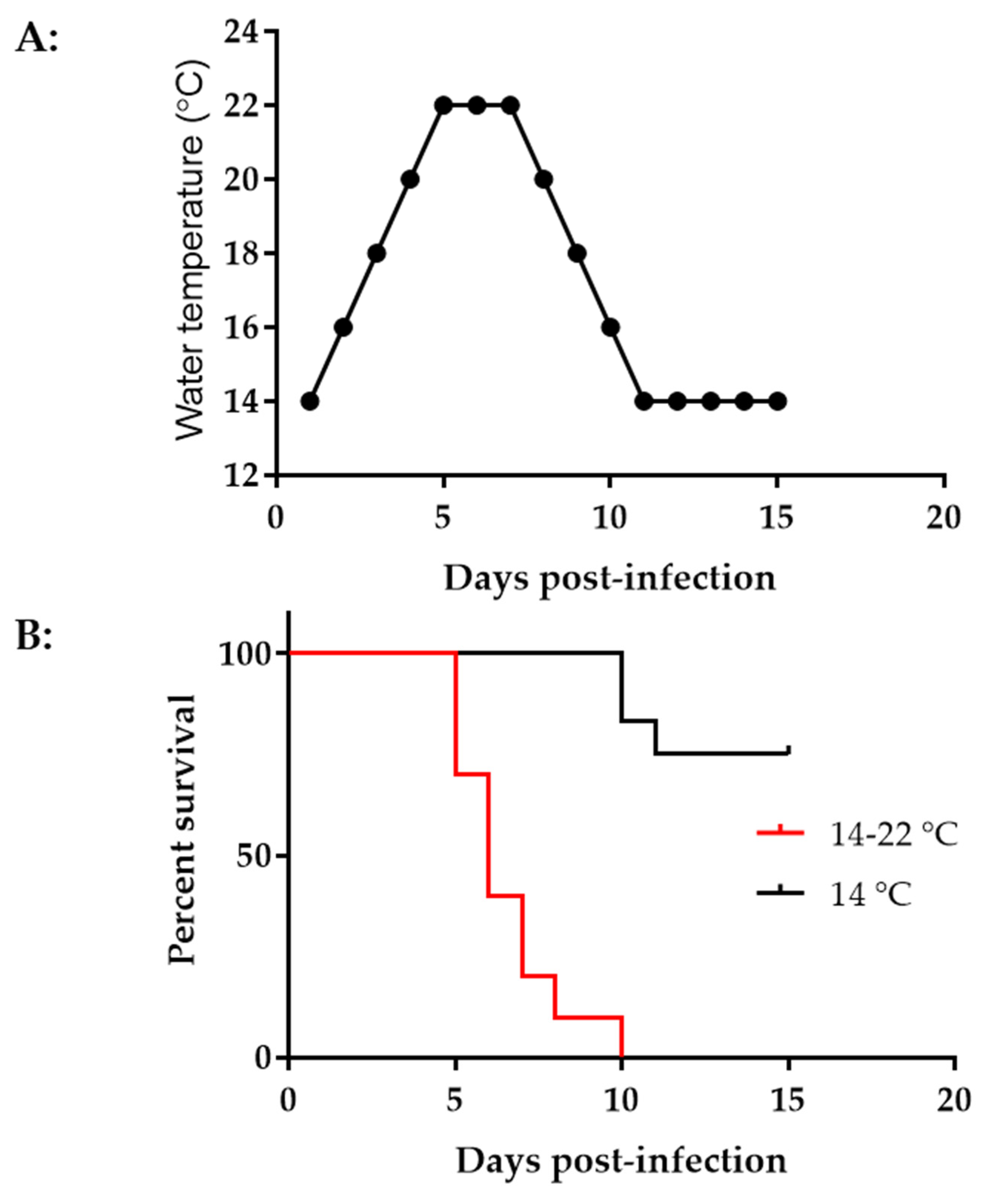
3.5. CyHV-3 Induced Mortality Is Different at Different Temperatures in Younger Koi
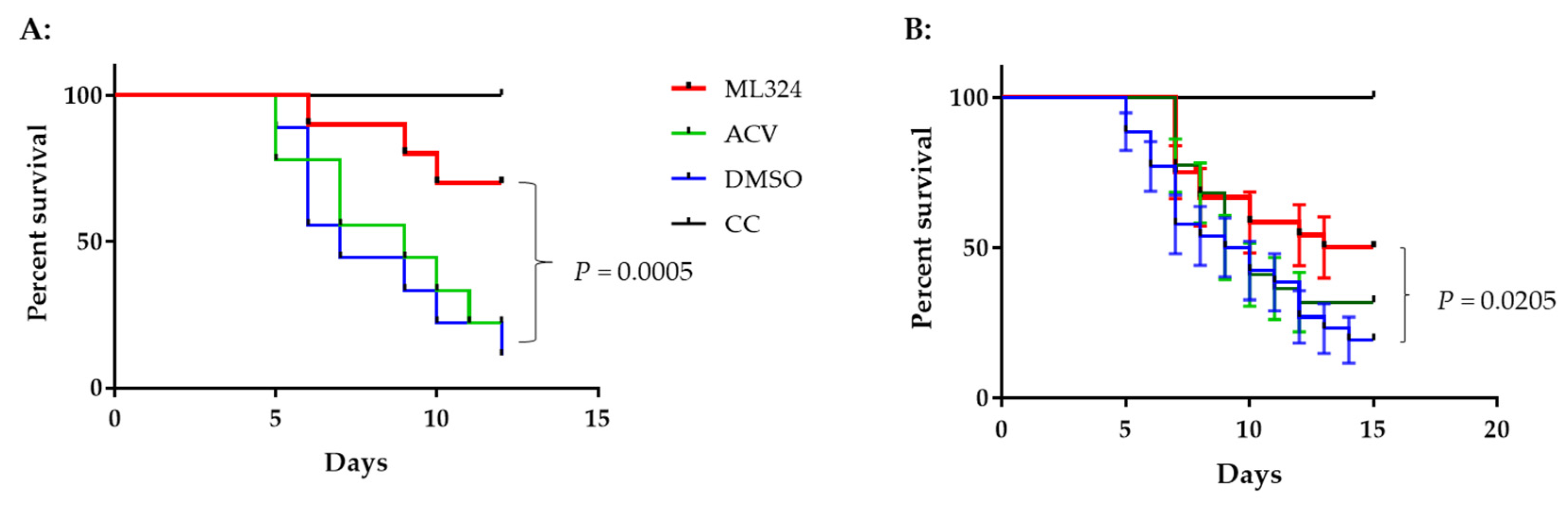
3.6. ML324 Reduced Mortality in CyHV-3 Infected Younger Koi
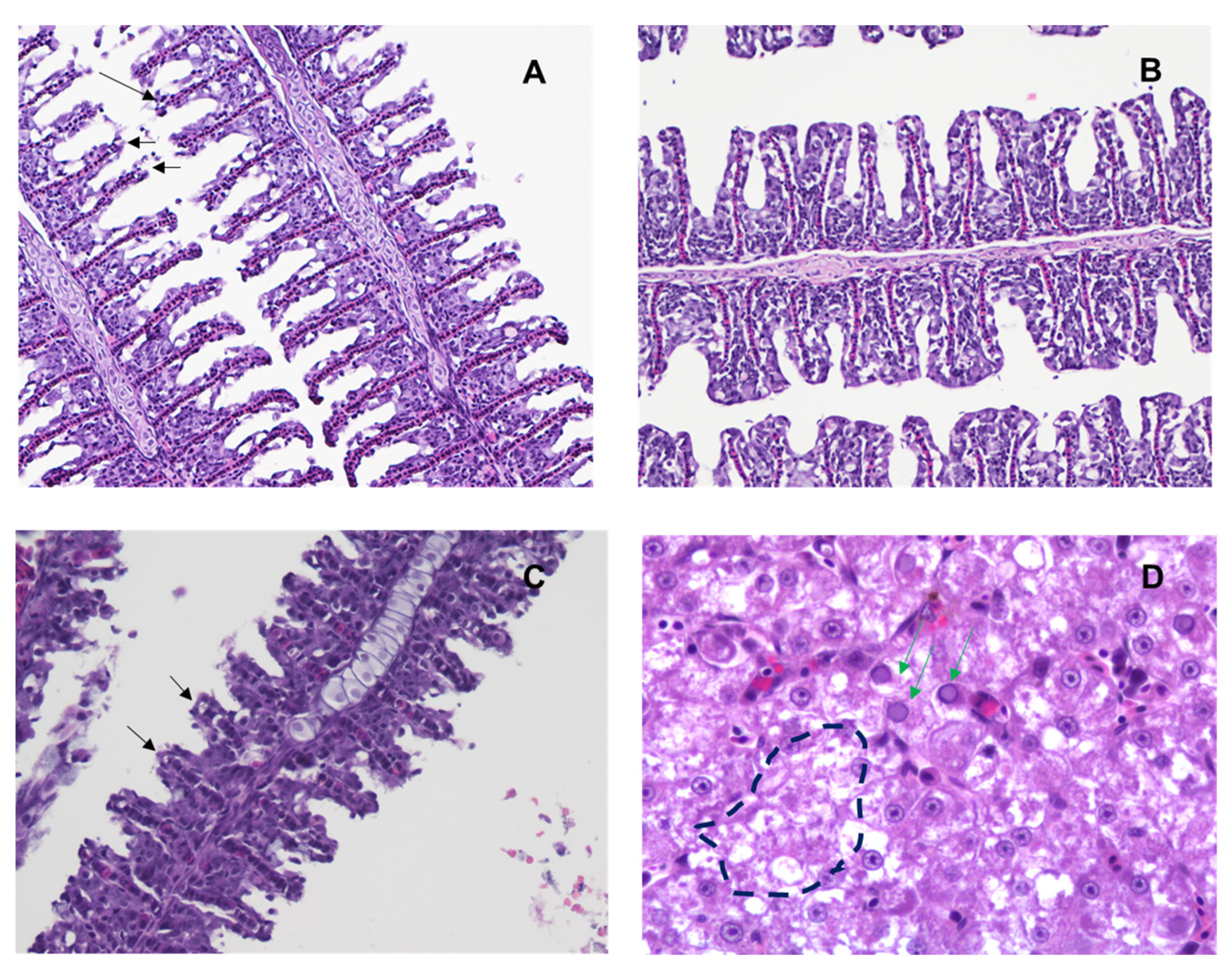
3.7. ML324 Reduced Histopathology in CyHV-3 Infected Younger Koi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilad, O.; Yun, S.; Andree, K.B.; Adkison, M.A.; Zlotkin, A.; Bercovier, H.; Eldar, A.; Hedrick, R.P. Initial characteristics of koi herpesvirus and development of a polymerase chain reaction assay to detect the virus in koi, Cyprinus carpio koi. Dis. Aquat. Organ. 2002, 48, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, R.P.; Gilad, O.; Yun, S.; Spangenberg, J.V.; Marty, G.D.; Nordhausen, R.W.; Kebus, M.J.; Bercovier, H.; Eldar, A. A Herpesvirus Associated with Mass Mortality of Juvenile and Adult Koi, a Strain of Common Carp. J. Aquat. Anim. Health 2000, 12, 44–57. [Google Scholar] [CrossRef]
- Cheng, L.; Chen, C.Y.; Tsai, M.A.; Wang, P.C.; Hsu, J.P.; Chern, R.S.; Chen, S.C. Koi herpesvirus epizootic in cultured carp and koi, Cyprinus carpio L., in Taiwan. J. Fish Dis. 2011, 34, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Gilad, O.; Yun, S.; Zagmutt-Vergara, F.J.; Leutenegger, C.M.; Bercovier, H.; Hedrick, R.P. Concentrations of a Koi herpesvirus (KHV) in tissues of experimentally infected Cyprinus carpio koi as assessed by real-time TaqMan PCR. Dis. Aquat. Organ. 2004, 60, 179–187. [Google Scholar] [CrossRef]
- Eide, K.; Miller-Morgan, T.; Heidel, J.; Bildfell, R.; Jin, L. Results of total DNA measurement in koi tissue by Koi Herpes Virus real-time PCR. J. Virol. Methods 2011, 172, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Waltzek, T.B.; Kelley, G.O.; Stone, D.M.; Way, K.; Hanson, L.; Fukuda, H.; Hirono, I.; Aoki, T.; Davison, A.J.; Hedrick, R.P. Koi herpesvirus represents a third cyprinid herpesvirus (CyHV-3) in the family Herpesviridae. J. Gen. Virol. 2005, 86, 1659–1667. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Beevers, N.; Joiner, C.; Hedrick, R.P.; Way, K. Antibody response of two populations of common carp, Cyprinus carpio L., exposed to koi herpesvirus. J. Fish Dis. 2009, 32, 311–320. [Google Scholar] [CrossRef]
- Eide, K.E.; Miller-Morgan, T.; Heidel, J.R.; Kent, M.L.; Bildfell, R.J.; Lapatra, S.; Watson, G.; Jin, L. Investigation of koi herpesvirus latency in koi. J. Virol. 2011, 85, 4954–4962. [Google Scholar] [CrossRef]
- Xu, J.R.; Bently, J.; Beck, L.; Reed, A.; Miller-Morgan, T.; Heidel, J.R.; Kent, M.L.; Rockey, D.D.; Jin, L. Analysis of koi herpesvirus latency in wild common carp and ornamental koi in Oregon, USA. J. Virol. Methods 2013, 187, 372–379. [Google Scholar] [CrossRef]
- Amin, M.; Adrianti, D.N.; Lasmika, N.L.A.; Ali, M. Detection of koi herpesvirus in healthy common carps, Cyprinus carpio L. Virusdisease 2018, 29, 445–452. [Google Scholar] [CrossRef]
- Lin, L.; Chen, S.; Russell, D.S.; Lohr, C.V.; Milston-Clements, R.; Song, T.; Miller-Morgan, T.; Jin, L. Analysis of stress factors associated with KHV reactivation and pathological effects from KHV reactivation. Virus Res. 2017, 240, 200–206. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E. Antivirals for the treatment of herpesvirus infections. J. Antimicrob. Chemother. 1993, 32 (Suppl. A), 121–132. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, E.C. Current and potential therapies for the treatment of herpes-virus infections. Prog. Drug Res. 2003, 60, 263–307. [Google Scholar] [CrossRef] [PubMed]
- Troszok, A.; Kolek, L.; Szczygiel, J.; Ostrowski, T.; Adamek, M.; Irnazarow, I. Anti-CyHV-3 Effect of Fluorescent, Tricyclic Derivative of Acyclovir 6-(4-MeOPh)-TACV in vitro. J. Vet. Res. 2019, 63, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Troszok, A.; Kolek, L.; Szczygiel, J.; Wawrzeczko, J.; Borzym, E.; Reichert, M.; Kaminska, T.; Ostrowski, T.; Jurecka, P.; Adamek, M.; et al. Acyclovir inhibits Cyprinid herpesvirus 3 multiplication in vitro. J. Fish Dis. 2018, 41, 1709–1718. [Google Scholar] [CrossRef]
- Quijano Carde, E.M.; Yazdi, Z.; Yun, S.; Hu, R.; Knych, H.; Imai, D.M.; Soto, E. Pharmacokinetic and Efficacy Study of Acyclovir Against Cyprinid Herpesvirus 3 in Cyprinus carpio. Front. Vet. Sci. 2020, 7, 587952. [Google Scholar] [CrossRef]
- Roizman, B.; Pellette, P.E. The Family Herpesviridae: A Brief Introduction, 4th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott William & Wilkins: Philadelphia, PA, USA, 2001; pp. 2381–2397. [Google Scholar]
- Dulbecco, R.; Ginsberg, H. Herpesviruses. In Virology; J.B. Lippincott Company: Philadelphia, PA, USA, 1988; Volume 2, pp. 161–177. [Google Scholar]
- Rice, S.A.; Knipe, D.M. Gene-specific transactivation by herpes simplex virus type 1 alpha protein ICP27. J. Virol. 1988, 62, 3814–3823. [Google Scholar] [CrossRef]
- Culhane, J.C.; Cole, P.A. LSD1 and the chemistry of histone demethylation. Curr. Opin. Chem. Biol. 2007, 11, 561–568. [Google Scholar] [CrossRef]
- Shi, Y.; Lan, F.; Matson, C.; Mulligan, P.; Whetstine, J.R.; Cole, P.A.; Casero, R.A.; Shi, Y. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell 2004, 119, 941–953. [Google Scholar] [CrossRef]
- Klose, R.J.; Yamane, K.; Bae, Y.; Zhang, D.; Erdjument-Bromage, H.; Tempst, P.; Wong, J.; Zhang, Y. The transcriptional repressor JHDM3A demethylates trimethyl histone H3 lysine 9 and lysine 36. Nature 2006, 442, 312–316. [Google Scholar] [CrossRef]
- Whetstine, J.R.; Nottke, A.; Lan, F.; Huarte, M.; Smolikov, S.; Chen, Z.; Spooner, E.; Li, E.; Zhang, G.; Colaiacovo, M.; et al. Reversal of histone lysine trimethylation by the JMJD2 family of histone demethylases. Cell 2006, 125, 467–481. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Quenelle, D.; Vogel, J.L.; Mascaro, C.; Ortega, A.; Kristie, T.M. A novel selective LSD1/KDM1A inhibitor epigenetically blocks herpes simplex virus lytic replication and reactivation from latency. mBio 2013, 4, e00558-00512. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Vogel, J.L.; Arbuckle, J.H.; Rai, G.; Jadhav, A.; Simeonov, A.; Maloney, D.J.; Kristie, T.M. Targeting the JMJD2 histone demethylases to epigenetically control herpesvirus infection and reactivation from latency. Sci. Transl. Med. 2013, 5, 167ra165. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Vogel, J.L.; Narayanan, A.; Peng, H.; Kristie, T.M. Inhibition of the histone demethylase LSD1 blocks alpha-herpesvirus lytic replication and reactivation from latency. Nat. Med. 2009, 15, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.K.; Smyth, G.K.; Greenfield, P.F. Accuracy of the endpoint assay for virus titration. Cytotechnology 1992, 8, 231–236. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Beevers, N.; Way, K.; Le Deuff, R.M.; Martin, P.; Joiner, C. Reactivation of koi herpesvirus infections in common carp Cyprinus carpio. Dis. Aquat. Organ. 2005, 67, 15–23. [Google Scholar] [CrossRef]
- Zhou, X.; Ma, H. Evolutionary history of histone demethylase families: Distinct evolutionary patterns suggest functional divergence. BMC Evol. Biol. 2008, 8, 294. [Google Scholar] [CrossRef]
- Ilouze, M.; Dishon, A.; Kotler, M. Coordinated and sequential transcription of the cyprinid herpesvirus-3 annotated genes. Virus Res. 2012, 169, 98–106. [Google Scholar] [CrossRef]
- Reed, A.; Lin, L.; Ostertag-Hill, C.; Wang, Q.; Wu, Z.; Miller-Morgan, T.; Jin, L. Detection of ORF6 protein associated with latent KHV infection. Virology 2017, 500, 82–90. [Google Scholar] [CrossRef]
- Omori, R.; Adams, B. Disrupting seasonality to control disease outbreaks: The case of koi herpes virus. J. Theor. Biol. 2011, 271, 159–165. [Google Scholar] [CrossRef]
- Choi, D.L.; Sohn, S.G.; Bang, J.D.; Do, J.W.; Park, M.S. Ultrastructural identification of a herpes-like virus infection in common carp Cyprinus carpio in Korea. Dis. Aquat. Organ. 2004, 61, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Minamoto, T.; Honjo, M.N.; Uchii, K.; Yamanaka, H.; Suzuki, A.A.; Kohmatsu, Y.; Iida, T.; Kawabata, Z. Detection of cyprinid herpesvirus 3 DNA in river water during and after an outbreak. Vet. Microbiol. 2009, 135, 261–266. [Google Scholar] [CrossRef]
- Boutier, M.; Gao, Y.; Donohoe, O.; Vanderplasschen, A. Current knowledge and future prospects of vaccines against cyprinid herpesvirus 3 (CyHV-3). Fish Shellfish Immunol. 2019, 93, 531–541. [Google Scholar] [CrossRef]
- Schroder, L.; Klafack, S.; Bergmann, S.M.; Fichtner, D.; Jin, Y.; Lee, P.Y.; Hoper, D.; Mettenleiter, T.C.; Fuchs, W. Generation of a potential koi herpesvirus live vaccine by simultaneous deletion of the viral thymidine kinase and dUTPase genes. J. Gen. Virol. 2019, 100, 642–655. [Google Scholar] [CrossRef]
- Ma, Y.; Liu, Z.; Hao, L.; Wu, J.; Qin, B.; Liang, Z.; Ma, J.; Ke, H.; Yang, H.; Li, Y.; et al. Oral vaccination using Artemia coated with recombinant Saccharomyces cerevisiae expressing cyprinid herpesvirus-3 envelope antigen induces protective immunity in common carp (Cyprinus carpio var. Jian) larvae. Res. Vet. Sci. 2020, 130, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Li, Y.; Wang, Q.; Wang, G.; Zhu, B.; Wang, Y.; Zeng, W.; Yin, J.; Liu, C.; Bergmann, S.M.; et al. Carbon nanotube-based DNA vaccine against koi herpesvirus given by intramuscular injection. Fish Shellfish Immunol. 2020, 98, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Embregts, C.W.E.; Tadmor-Levi, R.; Vesely, T.; Pokorova, D.; David, L.; Wiegertjes, G.F.; Forlenza, M. Intra-muscular and oral vaccination using a Koi Herpesvirus ORF25 DNA vaccine does not confer protection in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2019, 85, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Perelberg, A.; Ilouze, M.; Kotler, M.; Steinitz, M. Antibody response and resistance of Cyprinus carpio immunized with cyprinid herpes virus 3 (CyHV-3). Vaccine 2008, 26, 3750–3756. [Google Scholar] [CrossRef]
- Buchmann, K. Evolution of Innate Immunity: Clues from Invertebrates via Fish to Mammals. Front. Immunol. 2014, 5, 459. [Google Scholar] [CrossRef]
- Boehm, T.; Iwanami, N.; Hess, I. Evolution of the immune system in the lower vertebrates. Annu. Rev. Genomics Hum. Genet. 2012, 13, 127–149. [Google Scholar] [CrossRef]
- Cowen, P.; Li, S.; Guy, J.S.; Erickson, G.A.; Blanchard, D. Reactivation of latent pseudorabies virus infection in vaccinated commercial sows. Am. J. Vet. Res. 1990, 51, 354–358. [Google Scholar] [PubMed]
- Mengeling, W.L. Virus reactivation in pigs latently infected with a thymidine kinase negative vaccine strain of pseudorabies virus. Arch. Virol. 1991, 120, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Yoon, S.S.; Kim, S.E.; Ahn Jo, S. KDM4B histone demethylase and G9a regulate expression of vascular adhesion proteins in cerebral microvessels. Sci. Rep. 2017, 7, 45005. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Gao, Y.; Wang, S.; Xu, F.F.; Dai, L.S.; Jiang, H.; Yu, X.F.; Chen, C.Z.; Yuan, B.; Zhang, J.B. Expression pattern of JMJD1C in oocytes and its impact on early embryonic development. Genet. Mol. Res. 2015, 14, 18249–18258. [Google Scholar] [CrossRef]
- Carnesecchi, J.; Forcet, C.; Zhang, L.; Tribollet, V.; Barenton, B.; Boudra, R.; Cerutti, C.; Billas, I.M.; Serandour, A.A.; Carroll, J.S.; et al. ERRalpha induces H3K9 demethylation by LSD1 to promote cell invasion. Proc. Natl. Acad. Sci. USA 2017, 114, 3909–3914. [Google Scholar] [CrossRef]
- Shi, Y.G.; Tsukada, Y. The discovery of histone demethylases. Cold Spring Harb. Perspect. Biol. 2013, 5, a017947. [Google Scholar] [CrossRef]
- Harancher, M.R.; Packard, J.E.; Cowan, S.P.; DeLuca, N.A.; Dembowski, J.A. Antiviral Properties of the LSD1 Inhibitor SP-2509. J. Virol. 2020, 94, e00974-20. [Google Scholar] [CrossRef]
- Tallmadge, R.L.; Zygelyte, E.; Van de Walle, G.R.; Kristie, T.M.; Felippe, M.J.B. Effect of a Histone Demethylase Inhibitor on Equine Herpesvirus-1 Activity In Vitro. Front. Vet. Sci. 2018, 5, 34. [Google Scholar] [CrossRef]
- Hill, J.M.; Quenelle, D.C.; Cardin, R.D.; Vogel, J.L.; Clement, C.; Bravo, F.J.; Foster, T.P.; Bosch-Marce, M.; Raja, P.; Lee, J.S.; et al. Inhibition of LSD1 reduces herpesvirus infection, shedding, and recurrence by promoting epigenetic suppression of viral genomes. Sci. Transl. Med. 2014, 6, 265ra169. [Google Scholar] [CrossRef]
- Moerdyk-Schauwecker, M.; Stein, D.A.; Eide, K.; Blouch, R.E.; Bildfell, R.; Iversen, P.; Jin, L. Inhibition of HSV-1 ocular infection with morpholino oligomers targeting ICP0 and ICP27. Antivir. Res. 2009, 84, 131–141. [Google Scholar] [CrossRef]
- Rai, G.; Kawamura, A.; Tumber, A.; Liang, Y.; Vogel, J.L.; Arbuckle, J.H.; Rose, N.R.; Dexheimer, T.S.; Foley, T.L.; King, O.N.; et al. Discovery of ML324, a JMJD2 demethylase inhibitor with demonstrated antiviral activity. In Probe Reports from the NIH Molecular Libraries Program; Bethesda: Rockville, MD, USA, 2010. [Google Scholar]
- Hamada, S.; Suzuki, T.; Mino, K.; Koseki, K.; Oehme, F.; Flamme, I.; Ozasa, H.; Itoh, Y.; Ogasawara, D.; Komaarashi, H.; et al. Design, synthesis, enzyme-inhibitory activity, and effect on human cancer cells of a novel series of jumonji domain-containing protein 2 histone demethylase inhibitors. J. Med. Chem. 2010, 53, 5629–5638. [Google Scholar] [CrossRef]
- Mishra, S.; Van Rechem, C.; Pal, S.; Clarke, T.L.; Chakraborty, D.; Mahan, S.D.; Black, J.C.; Murphy, S.E.; Lawrence, M.S.; Daniels, D.L.; et al. Cross-talk between Lysine-Modifying Enzymes Controls Site-Specific DNA Amplifications. Cell 2018, 174, 803–817 e816. [Google Scholar] [CrossRef] [PubMed]
- Fellous, A.; Earley, R.L.; Silvestre, F. The Kdm/Kmt gene families in the self-fertilizing mangrove rivulus fish, Kryptolebias marmoratus, suggest involvement of histone methylation machinery in development and reproduction. Gene 2019, 687, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Eide, K.; Moerdyk-Schauwecker, M.; Stein, D.A.; Bildfell, R.; Koelle, D.M.; Jin, L. Reduction of herpes simplex virus type-2 replication in cell cultures and in rodent models with peptide-conjugated morpholino oligomers. Antivir. Ther. 2010, 15, 1141–1149. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene | Primer Sequences (5′-3′) |
|---|---|---|
| IE * | ||
| ORF139-116-F | GCCTACTGGGAGGATATGTA | |
| ORF139-116-R | CCCTGGTCTTGACAGAAATAG | |
| ORF155-134-F | GGAGAGGAGAAGGGAGAAAT | |
| ORF155-134-R | GAGTAGTTGTGGGTGATGAAG | |
| E ** | ||
| ORF46-103-F | GTCGATAGCGTCCTACTTTG | |
| ORF46-103R | GACGCTCTGGTTGATGTT | |
| ORF144-122-F | CGGTGCGACAGATACATAGA | |
| ORF144-122-R | GATAGAGGAGAGGGTGAAGAG | |
| L *** | ||
| ORF78-103-F | CCTCTGTACAACAACCCAATAA | |
| ORF78-103-R | GTGTATTGCTGGATGGAAGG | |
| Latency ^ | ||
| ORF6s-F291 | GACCCAGGGGACAGCTCTAT | |
| ORF6s-R291 | AGTGGTACAAGTGGCGCTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuoka, S.; Petri, G.; Larson, K.; Behnke, A.; Wang, X.; Peng, M.; Spagnoli, S.; Lohr, C.; Milston-Clements, R.; Divilov, K.; et al. Evaluation of Histone Demethylase Inhibitor ML324 and Acyclovir against Cyprinid herpesvirus 3 Infection. Viruses 2023, 15, 163. https://doi.org/10.3390/v15010163
Matsuoka S, Petri G, Larson K, Behnke A, Wang X, Peng M, Spagnoli S, Lohr C, Milston-Clements R, Divilov K, et al. Evaluation of Histone Demethylase Inhibitor ML324 and Acyclovir against Cyprinid herpesvirus 3 Infection. Viruses. 2023; 15(1):163. https://doi.org/10.3390/v15010163
Chicago/Turabian StyleMatsuoka, Shelby, Gloria Petri, Kristen Larson, Alexandra Behnke, Xisheng Wang, Muhui Peng, Sean Spagnoli, Christiane Lohr, Ruth Milston-Clements, Konstantin Divilov, and et al. 2023. "Evaluation of Histone Demethylase Inhibitor ML324 and Acyclovir against Cyprinid herpesvirus 3 Infection" Viruses 15, no. 1: 163. https://doi.org/10.3390/v15010163
APA StyleMatsuoka, S., Petri, G., Larson, K., Behnke, A., Wang, X., Peng, M., Spagnoli, S., Lohr, C., Milston-Clements, R., Divilov, K., & Jin, L. (2023). Evaluation of Histone Demethylase Inhibitor ML324 and Acyclovir against Cyprinid herpesvirus 3 Infection. Viruses, 15(1), 163. https://doi.org/10.3390/v15010163