
Novel Polymorphisms and Genetic Characteristics of the Shadow of Prion Protein Gene (SPRN) in Cats, Hosts of Feline Spongiform Encephalopathy



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statements
2.2. Samples
2.3. Genomic DNA Isolation
2.4. Genetic Analysis
2.5. Statistical Analyses
2.6. Multiple Sequence Alignments
2.7. Secondary Structural Analysis of mRNA
2.8. Prediction of the Signal Peptide of Feline Sho
2.9. Prediction of Omega-Site and Signal Sequence of GPI-Anchor of Feline Sho
3. Results
3.1. Identification of Novel Polymorphisms of the Feline SPRN Gene in 193 Cats
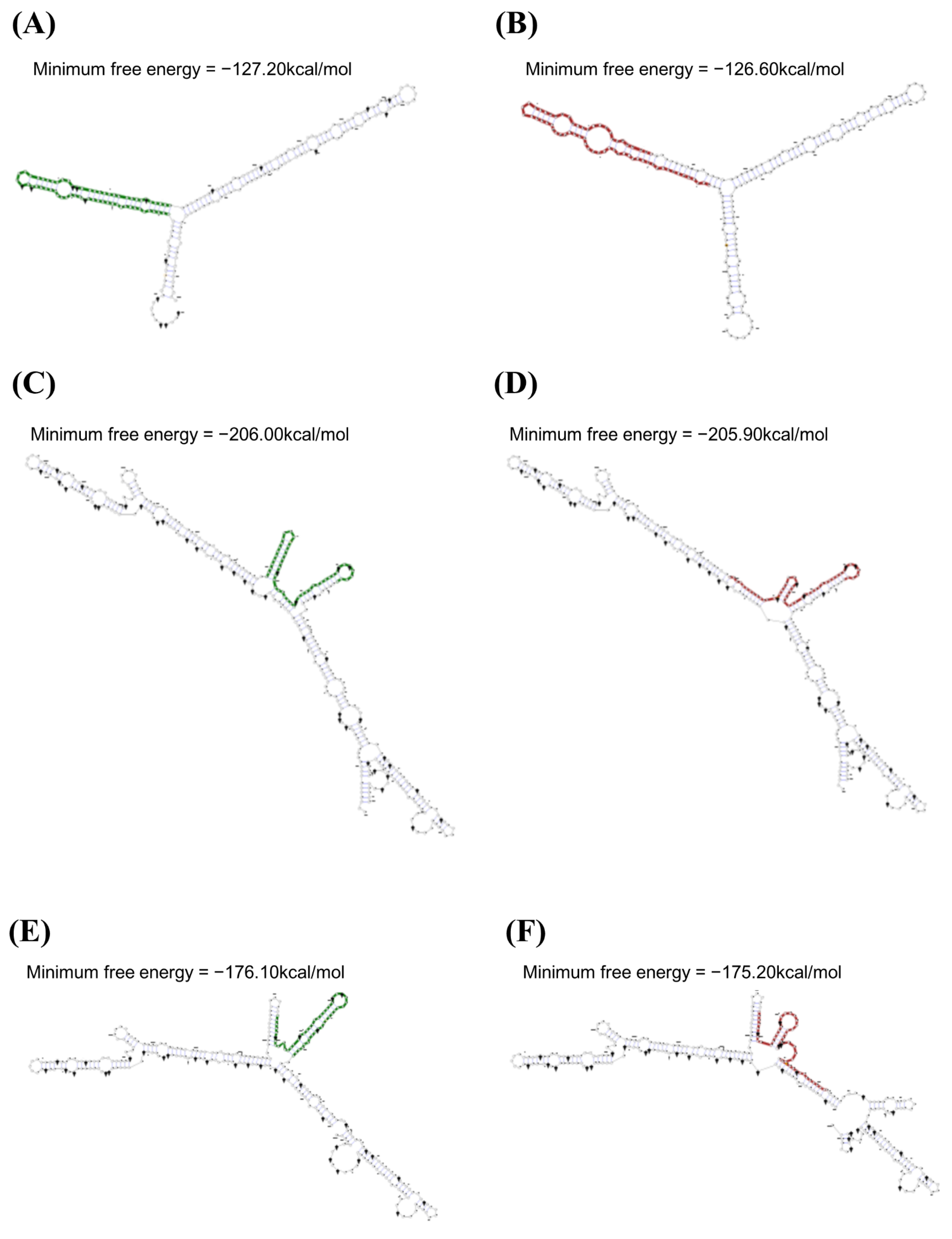
3.2. mRNA Secondary Structure Analysis of the Feline SPRN Gene According to Polymorphisms
3.3. Comparison of the Amino Acid Sequences of the Sho Protein among Species
3.4. Identification of Differences in the N-Terminal Signal Peptide of the Sho Protein among Species
3.5. Investigation of the Omega-Site and Signal Sequence of the GPl-Anchor of the Sho Protein
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prusiner, S.B. The prion diseases. Brain Pathol. 1998, 8, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Houston, F.; Andreoletti, O. Animal prion diseases: The risks to human health. Brain Pathol. 2019, 29, 248–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.C.; Park, K.J.; Hwang, J.Y.; Park, H.C.; Kang, H.E.; Sohn, H.J.; Jeong, B.H. In-depth examination of PrP(Sc) in Holstein cattle carrying the E211K somatic mutation of the bovine prion protein gene (PRNP). Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Won, S.Y.; Jeong, B.H. Altered expression of glymphatic system-related proteins in prion diseases: Implications for the role of the glymphatic system in prion diseases. Cell Mol. Immunol. 2021, 18, 2281–2283. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.J.; Samuel, M.D.; O’Rourke, K.I.; Johnson, C.J. The role of genetics in chronic wasting disease of North American cervids. Prion 2012, 6, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, I.S.; Kim, Y.C.; Won, S.Y.; Park, K.J.; Park, H.C.; Hwang, J.Y.; Kang, H.E.; Sohn, H.J.; Jeong, B.H. Association Study of the M132L Single Nucleotide Polymorphism with Susceptibility to Chronic Wasting Disease in Korean Elk: A Meta-Analysis. Front. Vet. Sci. 2021, 8, 804325. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Jeong, B.H. The First Meta-Analysis of the M129V Single-Nucleotide Polymorphism (SNP) of the Prion Protein Gene (PRNP) with Sporadic Creutzfeldt-Jakob Disease. Cells 2021, 10, 3132. [Google Scholar] [CrossRef]
- Kim, H.H.; Kim, Y.C.; Kim, K.; Kim, A.D.; Jeong, B.H. Novel Polymorphisms and Genetic Features of the Prion Protein Gene (PRNP) in Cats, Hosts of Feline Spongiform Encephalopathy. Genes 2020, 12, 13. [Google Scholar] [CrossRef]
- Baldwin, K.J.; Correll, C.M. Prion Disease. Semin. Neurol. 2019, 39, 428–439. [Google Scholar] [CrossRef]
- Taylor, D.R.; Whitehouse, I.J.; Hooper, N.M. Glypican-1 mediates both prion protein lipid raft association and disease isoform formation. PLoS Pathog. 2009, 5, e1000666. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Lee, J.; Lee, D.W.; Jeong, B.H. Large-scale lipidomic profiling identifies novel potential biomarkers for prion diseases and highlights lipid raft-related pathways. Vet. Res. 2021, 52, 105. [Google Scholar] [CrossRef]
- Deleault, N.R.; Piro, J.R.; Walsh, D.J.; Wang, F.; Ma, J.; Geoghegan, J.C.; Supattapone, S. Isolation of phosphatidylethanolamine as a solitary cofactor for prion formation in the absence of nucleic acids. Proc. Natl. Acad. Sci. USA 2012, 109, 8546–8551. [Google Scholar] [CrossRef] [Green Version]
- Lewis, V.; Hooper, N.M. The role of lipid rafts in prion protein biology. Front. Biosci. 2011, 16, 151–168. [Google Scholar] [CrossRef] [Green Version]
- Ciric, D.; Richard, C.A.; Moudjou, M.; Chapuis, J.; Sibille, P.; Daude, N.; Westaway, D.; Adrover, M.; Beringue, V.; Martin, D.; et al. Interaction between Shadoo and PrP Affects the PrP-Folding Pathway. J. Virol. 2015, 89, 6287–6293. [Google Scholar] [CrossRef] [Green Version]
- Daude, N.; Westaway, D. Biological properties of the PrP-like Shadoo protein. Front. Biosci. 2011, 16, 1505–1516. [Google Scholar] [CrossRef]
- Beck, J.A.; Campbell, T.A.; Adamson, G.; Poulter, M.; Uphill, J.B.; Molou, E.; Collinge, J.; Mead, S. Association of a null allele of SPRN with variant Creutzfeldt-Jakob disease. J. Med. Genet. 2008, 45, 813–817. [Google Scholar] [CrossRef]
- Gurgul, A.; Polak, M.P.; Larska, M.; Slota, E. PRNP and SPRN genes polymorphism in atypical bovine spongiform encephalopathy cases diagnosed in Polish cattle. J. Appl. Genet. 2012, 53, 337–342. [Google Scholar] [CrossRef]
- Sabarinathan, R.; Tafer, H.; Seemann, S.E.; Hofacker, I.L.; Stadler, P.F.; Gorodkin, J. The RNAsnp web server: Predicting SNP effects on local RNA secondary structure. Nucleic Acids Res. 2013, 41, W475–W479. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal omega. Curr. Protoc. Bioinf. 2014, 48, 3–13. [Google Scholar] [CrossRef]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gislason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022. [Google Scholar] [CrossRef]
- Pierleoni, A.; Martelli, P.L.; Casadio, R. PredGPI: A GPI-anchor predictor. BMC Bioinf. 2008, 9, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, J.E.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, J.C.; Drisaldi, B.; Ng, V.; Yang, J.; Strome, B.; Horne, P.; Sy, M.S.; Yoong, L.; Young, R.; Mastrangelo, P.; et al. The CNS glycoprotein Shadoo has PrP(C)-like protective properties and displays reduced levels in prion infections. EMBO J. 2007, 26, 4038–4050. [Google Scholar] [CrossRef] [PubMed]
- Ciric, D.; Rezaei, H. Biochemical insight into the prion protein family. Front. Cell Dev. Biol. 2015, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Dondapati, D.T.; Cingaram, P.R.; Ayaydin, F.; Nyeste, A.; Kanyó, A.; Welker, E.; Fodor, E. Membrane Domain Localization and Interaction of the Prion-Family Proteins, Prion and Shadoo with Calnexin. Membranes 2021, 11, 978. [Google Scholar] [CrossRef]
- Fang, Z.; Rajewsky, N. The impact of miRNA target sites in coding sequences and in 3′UTRs. PLoS ONE 2011, 6, e18067. [Google Scholar] [CrossRef]
- Didiano, D.; Hobert, O. Molecular architecture of a miRNA-regulated 3′ UTR. RNA 2008, 14, 1297–1317. [Google Scholar] [CrossRef] [Green Version]
- Peletto, S.; Bertolini, S.; Maniaci, M.G.; Colussi, S.; Modesto, P.; Biolatti, C.; Bertuzzi, S.; Caramelli, M.; Maurella, C.; Acutis, P.L. Association of an indel polymorphism in the 3′UTR of the caprine SPRN gene with scrapie positivity in the central nervous system. J. Gen. Virol. 2012, 93, 1620–1623. [Google Scholar] [CrossRef]
- Passet, B.; Castille, J.; Makhzami, S.; Truchet, S.; Vaiman, A.; Floriot, S.; Moazami-Goudarzi, K.; Vilotte, M.; Gaillard, A.L.; Helary, L.; et al. The Prion-like protein Shadoo is involved in mouse embryonic and mammary development and differentiation. Sci. Rep. 2020, 10, 6765. [Google Scholar] [CrossRef] [Green Version]
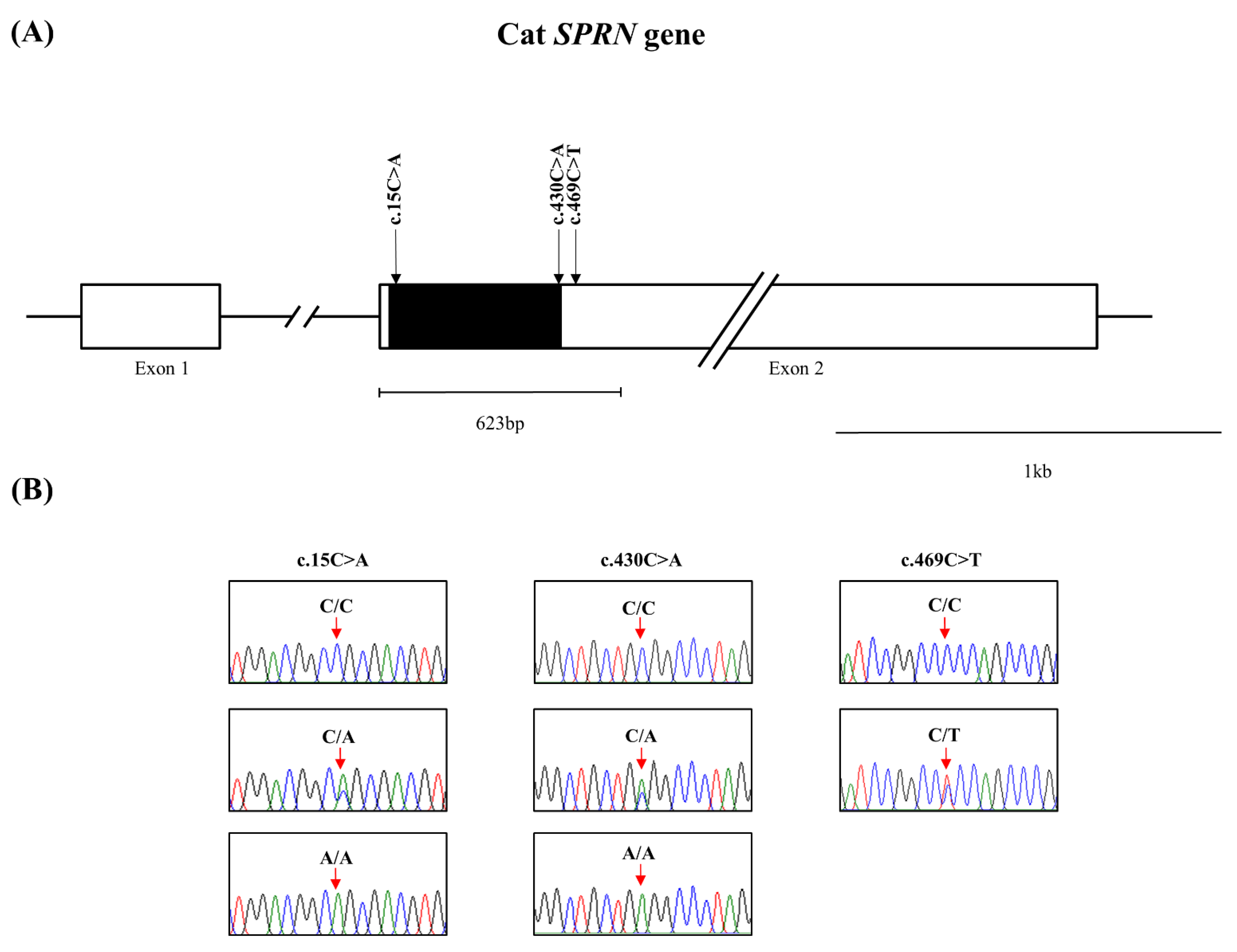



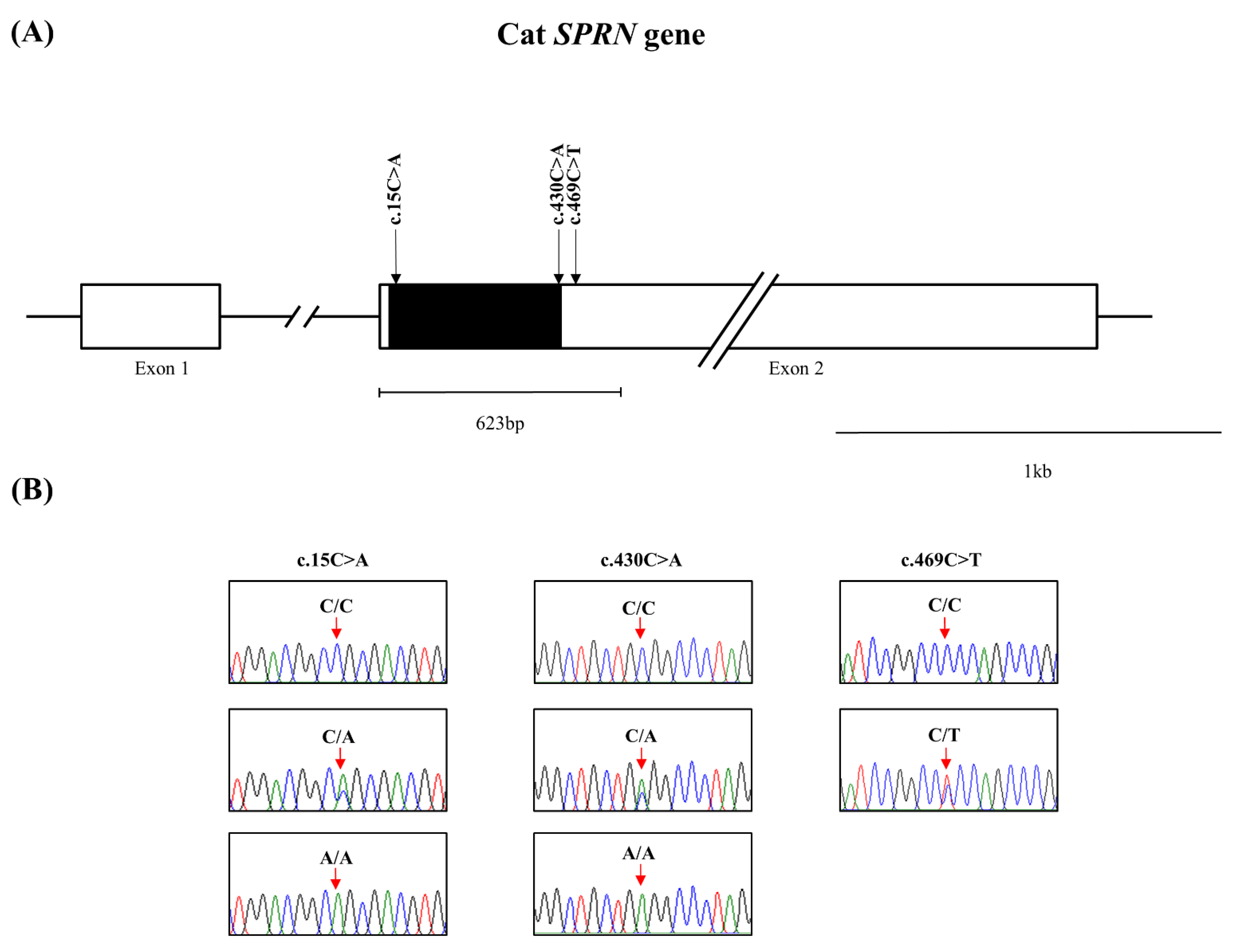{kind=link}
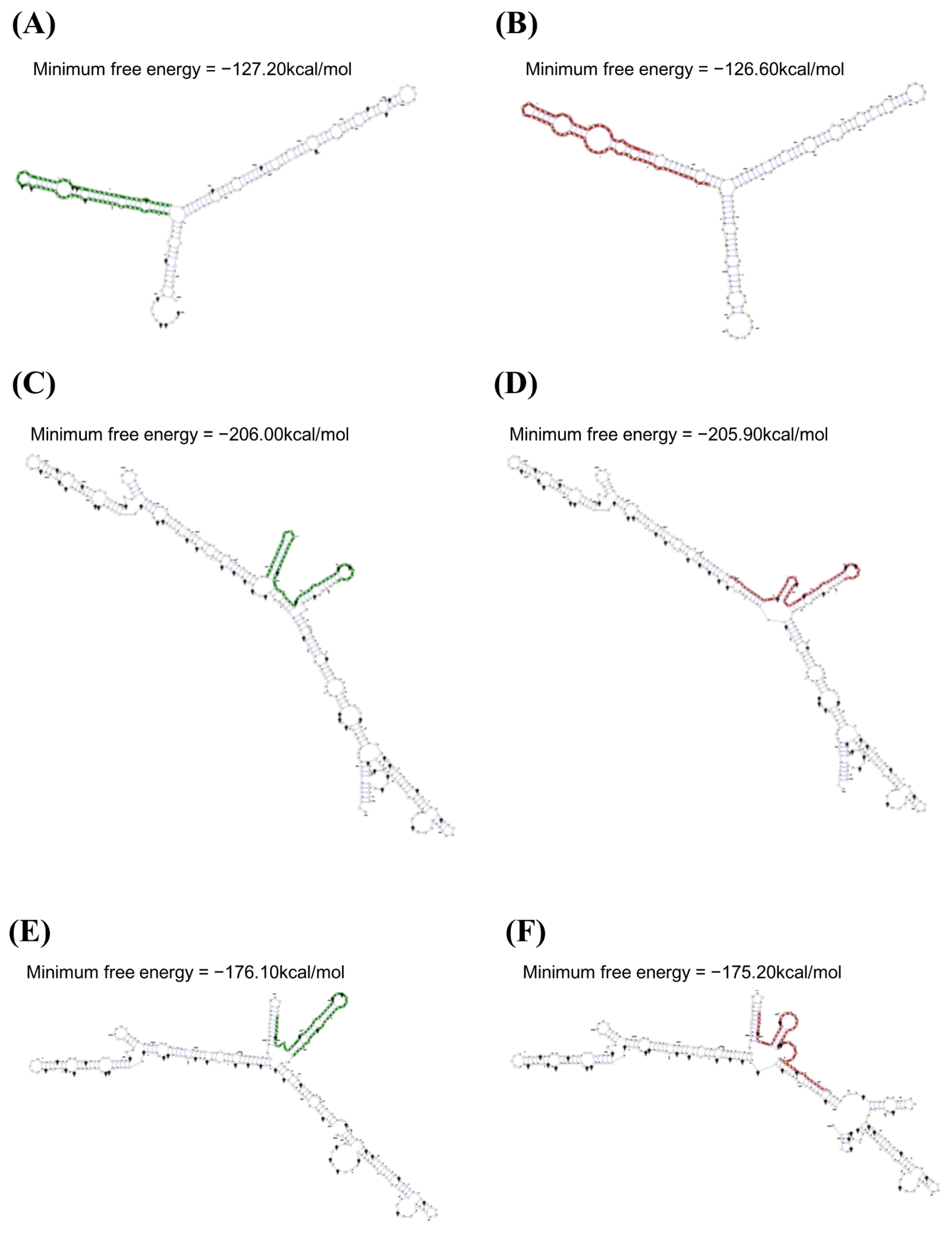{kind=link}
{kind=link}
{kind=link}
| Polymorphisms | Genotype Frequency, n (%) | Allele Frequency, n (%) | HWE | |||
|---|---|---|---|---|---|---|
| c.15C > A | CC 109 (56.48) | CA 67 (34.72) | AA 17 (8.81) | C 285 (73.83) | A 101 (26.17) | 0.158 |
| c.430C > A | CC 157 (81.35) | CA 33 (17.10) | AA 3 (1.55) | C 346 (89.90) | A 39 (10.10) | 0.414 |
| c.469C > T | CC 192 (99.48) | CT 1 (0.52) | TT 0 (0) | C 385 (99.74) | T 1 (0.26) | 0.971 |
| r2 | c.15C > A | c.430C > A | c.469C > T |
|---|---|---|---|
| c.15C > A | - | ||
| c.430C > A | 0.04 | - | |
| c.469C > T | 0.001 | 0 | - |
| Haplotype | c.15C > A | c.430C > A | c.469C > T | Frequency (n = 386) |
|---|---|---|---|---|
| ht1 | C | C | C | 245 (0.635) |
| ht2 | A | C | C | 101 (0.262) |
| ht3 | C | A | C | 39 (0.101) |
| ht4 | C | C | T | 1 (0.003) |
| r2 | SPRN c.15C > A | SPRN c.430C > A | SPRN c.469C > T |
|---|---|---|---|
| PRNP c.-3G > A | 0.002 | 0.007 | 0.0 |
| PRNP c.128G > A | 0.007 | 0.0 | 0.0 |
| PRNP c.171C > T | 0.001 | 0.007 | 0.002 |
| PRNP c.201C > T | 0.003 | 0.004 | 0.005 |
| PRNP c.214_240del CCCCACGCC GGCGGAGGC TGGGGTCAG | 0.007 | 0.03 | 0.0 |
| PRNP c.255T > C, G | 0.0 | 0.004 | 0.032 |
| PRNP c.264T > C | 0.0 | 0.008 | 0.015 |
| PRNP c.279C > T | 0.0 | 0.0 | 0.0 |
| PRNP c.457G > A | 0.003 | 0.001 | 0.023 |
| PRNP c.734C > T | 0.003 | 0.007 | 0.0 |
| PRNP c.774C > T | 0.0 | 0.002 | 0.0 |
| PRNP c.787C > T | 0.002 | 0.046 | 0.0 |
| PRNP c.789G > A | 0.004 | 0.009 | 0.0 |
| PRNP c.790C > T | 0.0 | 0.011 | 0.0 |
| PRNP c.979G > A | 0.002 | 0.007 | 0.0 |
| Species | Omega-Site | Signal Sequence | |||
|---|---|---|---|---|---|
| Position | Amino Acid | Position | Length | Protein Sequence | |
| Humans | 125 | S | 125–151 | 27 | SGAGPTRGPRLCLVLGGALGALGLLRP |
| Mice | 116 | Y | 116–147 | 32 | YWAWTSGSGSVHSPRICLLLGGTLGALELLRP |
| Cattle | 112 | Y | 112–143 | 32 | YWAWTSGTGPTGHRHLCPLLGGALGALRLLRP |
| Sheep | 119 | S | 119–145 | 27 | SGAGPTGHRHLCPLLGGALGALRLLRP |
| Goats | 117 | S | 117–146 | 30 | SGAGPTGHRHLCPLLGGALGALRLLRPLRP |
| Red deer | 117 | S | 117–143 | 27 | SGAGPTGHRHLCPLLGGTLGALRLLRP |
| Dogs | 121 | S | 121–147 | 27 | SGAGPTGSPRLCLLLGGALGARGLLPP |
| Horses | 117 | S | 117–147 | 31 | SAWTSGTEPTRGLCLCLLLGGALGALGLLRP |
| Cats | 120 | D | 120–145 | 26 | DAGPTGGPRLCLLLGGALVALGLLRP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-C.; Kim, H.-H.; Kim, K.; Kim, A.-D.; Jeong, B.-H. Novel Polymorphisms and Genetic Characteristics of the Shadow of Prion Protein Gene (SPRN) in Cats, Hosts of Feline Spongiform Encephalopathy. Viruses 2022, 14, 981. https://doi.org/10.3390/v14050981
Kim Y-C, Kim H-H, Kim K, Kim A-D, Jeong B-H. Novel Polymorphisms and Genetic Characteristics of the Shadow of Prion Protein Gene (SPRN) in Cats, Hosts of Feline Spongiform Encephalopathy. Viruses. 2022; 14(5):981. https://doi.org/10.3390/v14050981
Chicago/Turabian StyleKim, Yong-Chan, Hyeon-Ho Kim, Kiwon Kim, An-Dang Kim, and Byung-Hoon Jeong. 2022. "Novel Polymorphisms and Genetic Characteristics of the Shadow of Prion Protein Gene (SPRN) in Cats, Hosts of Feline Spongiform Encephalopathy" Viruses 14, no. 5: 981. https://doi.org/10.3390/v14050981
APA StyleKim, Y.-C., Kim, H.-H., Kim, K., Kim, A.-D., & Jeong, B.-H. (2022). Novel Polymorphisms and Genetic Characteristics of the Shadow of Prion Protein Gene (SPRN) in Cats, Hosts of Feline Spongiform Encephalopathy. Viruses, 14(5), 981. https://doi.org/10.3390/v14050981