
Targeting Host PIM Protein Kinases Reduces Mayaro Virus Replication

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells Lines, Virus Strains and Reagents
2.2. Cell Viability Analysis
2.3. Evaluation of PIM Kinase Inhibitors’ Antiviral Effects
2.4. Viral Plaque-Forming Assay
2.5. Time of Addition Experiment
2.6. Immunofluorescence Assay
2.7. Protein Analysis
2.8. Data Analysis
3. Results
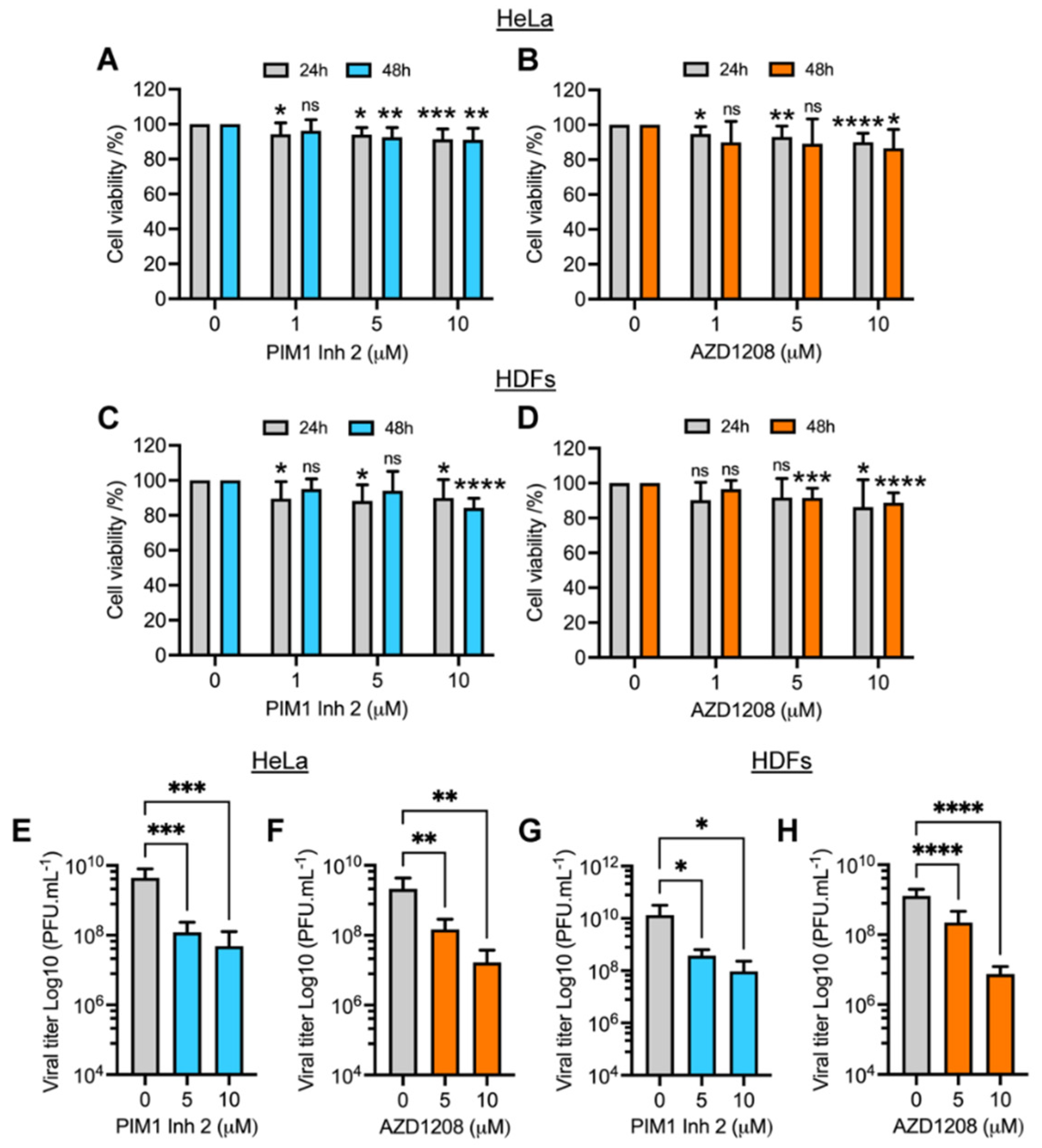
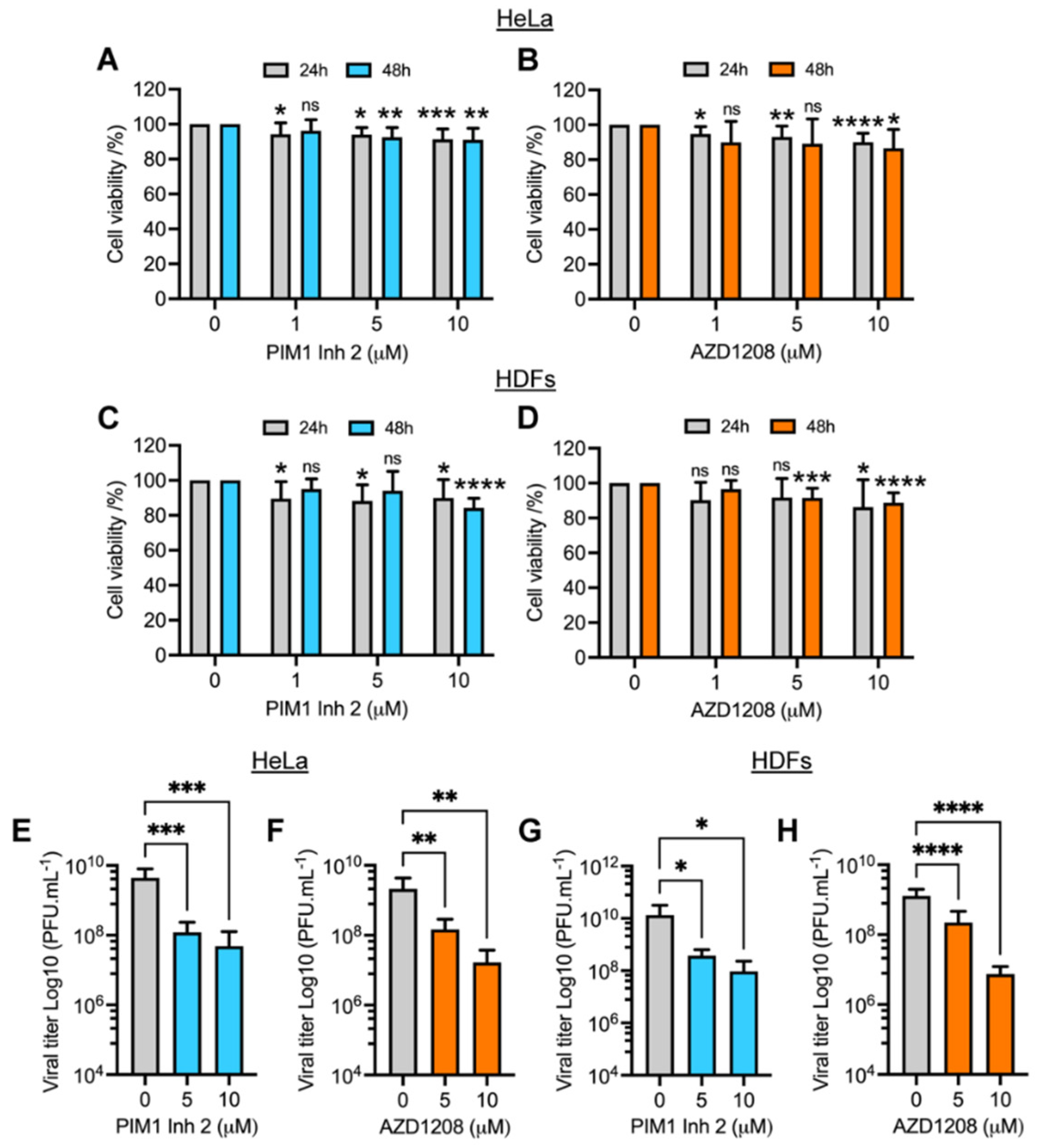
3.1. PIM1 Inhibitor 2 and AZD1208 Partially Prevent MAYV-Induced Cytopathic Effects and Reduce Viral Progeny Production in HeLa Cells and Human Dermal Fibroblasts (HDFs)
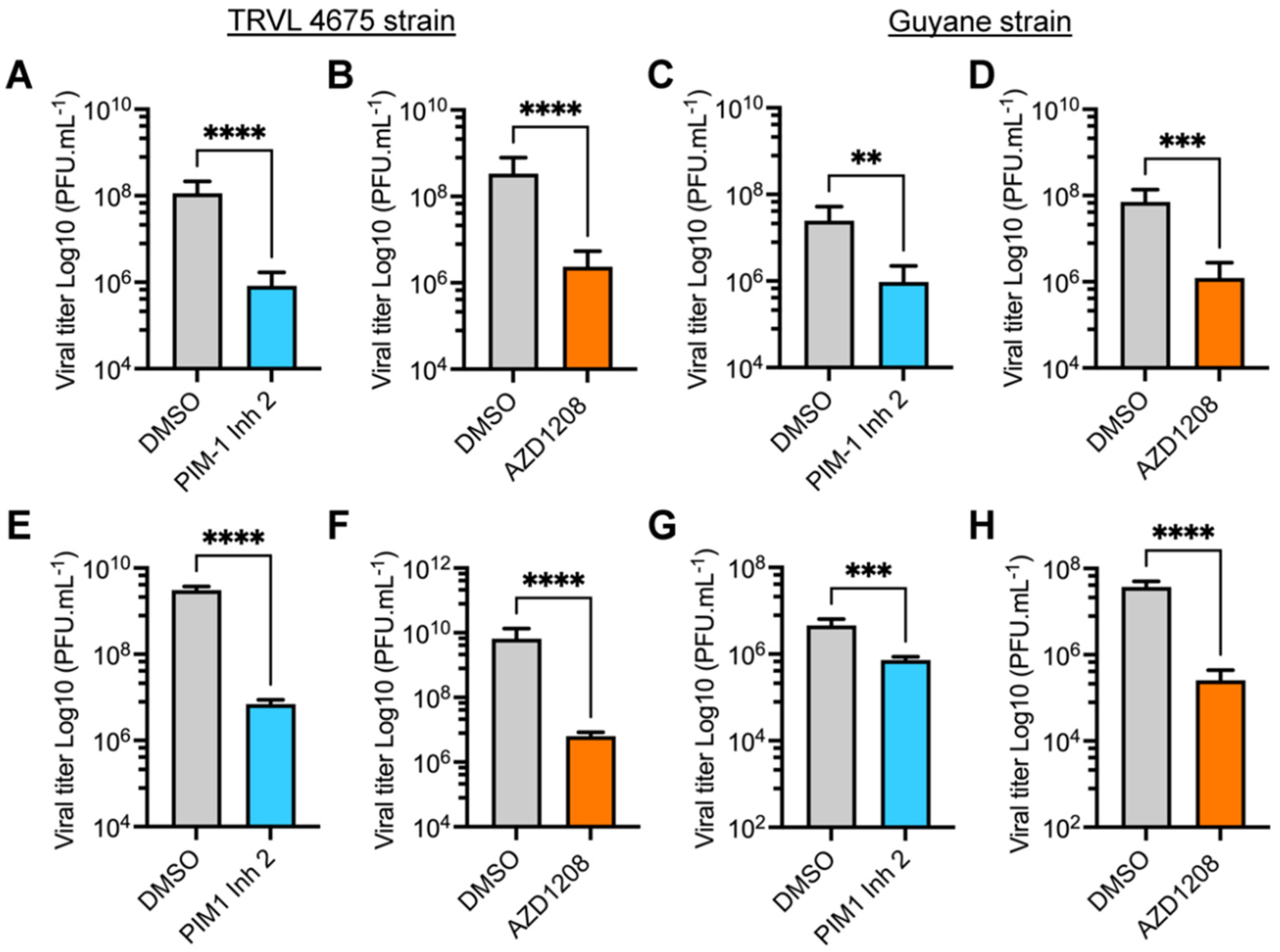
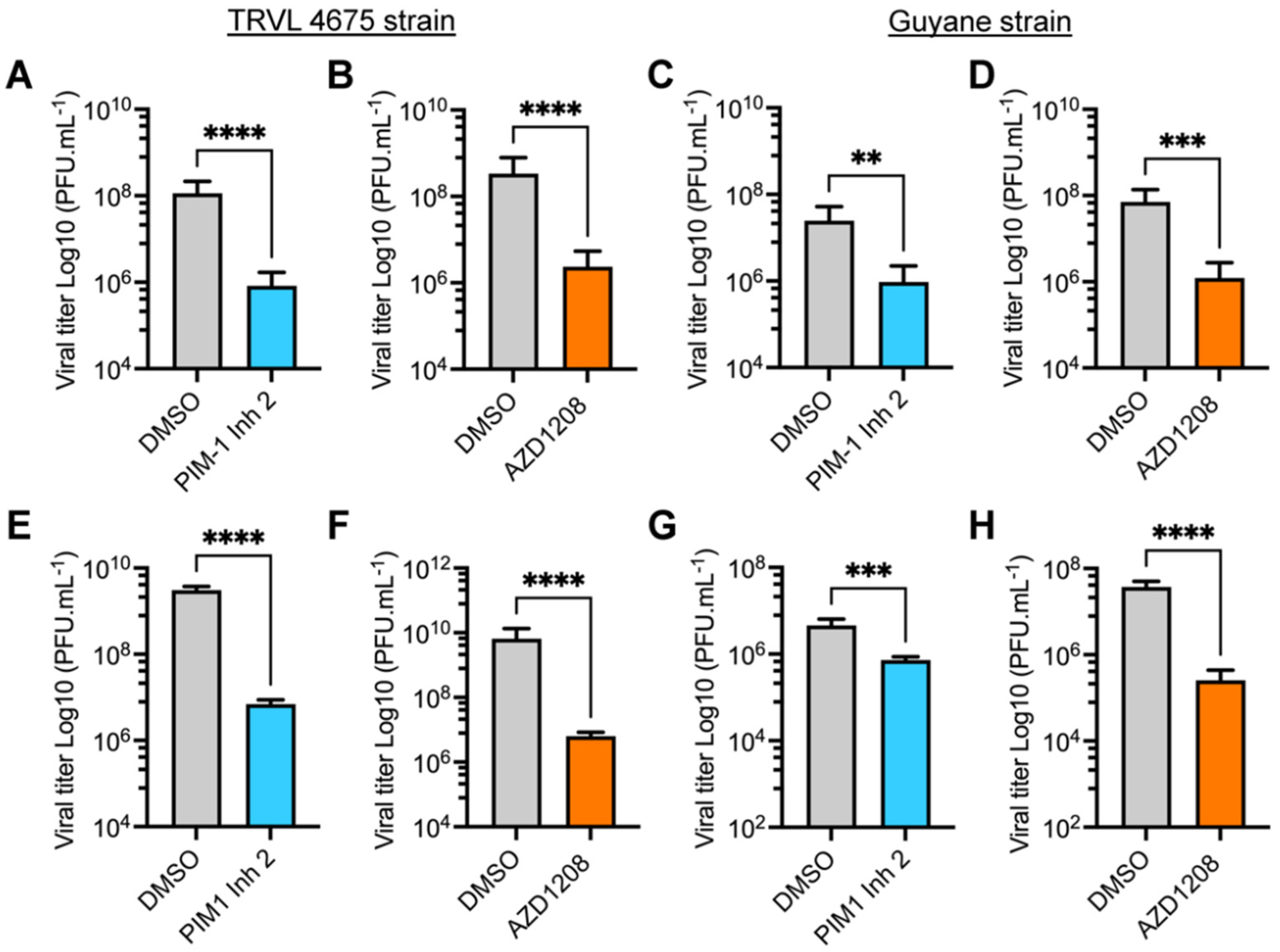
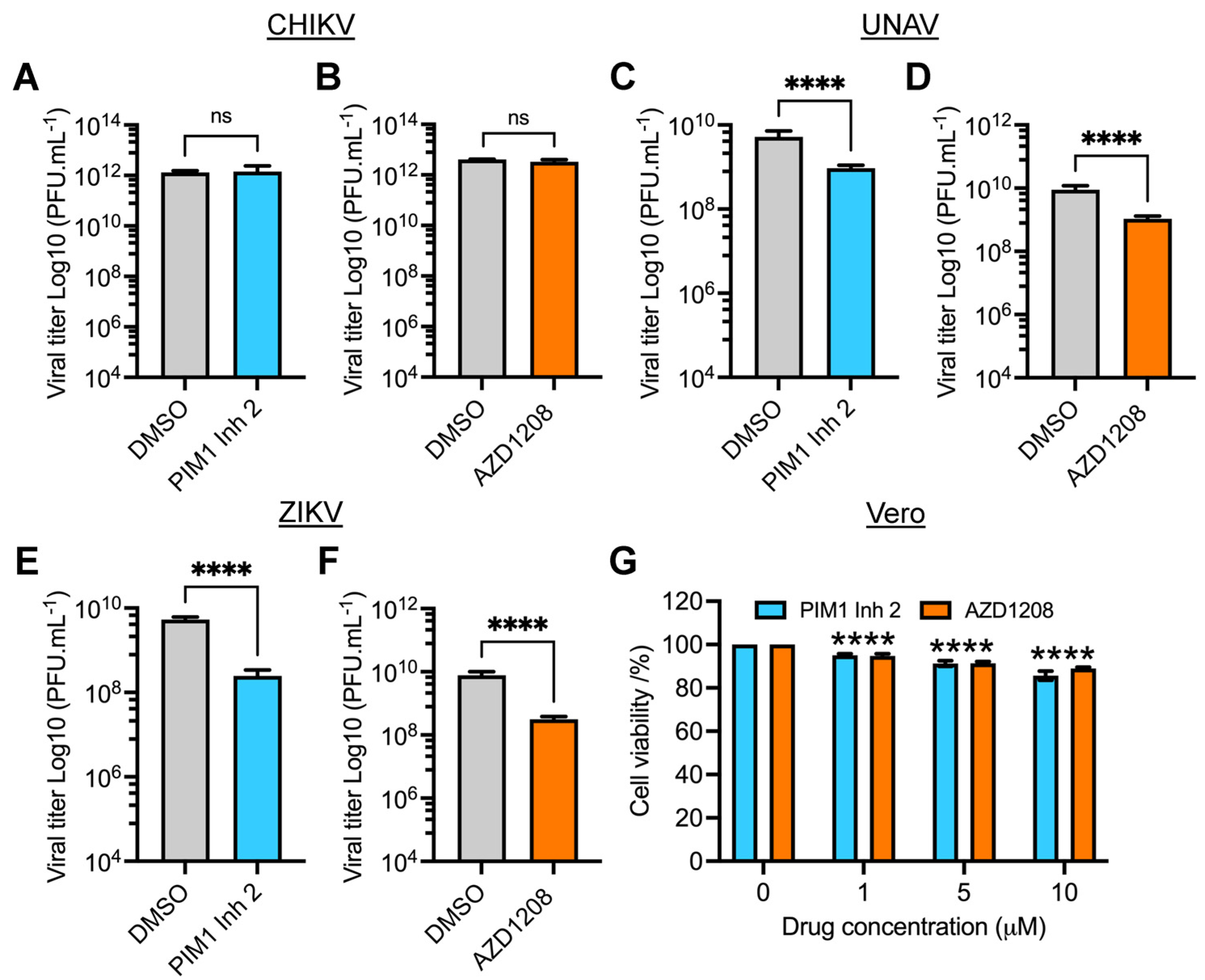
3.2. PIM Kinase Inhibitors Reduce Viral Replication in Multiple MAYV Strains
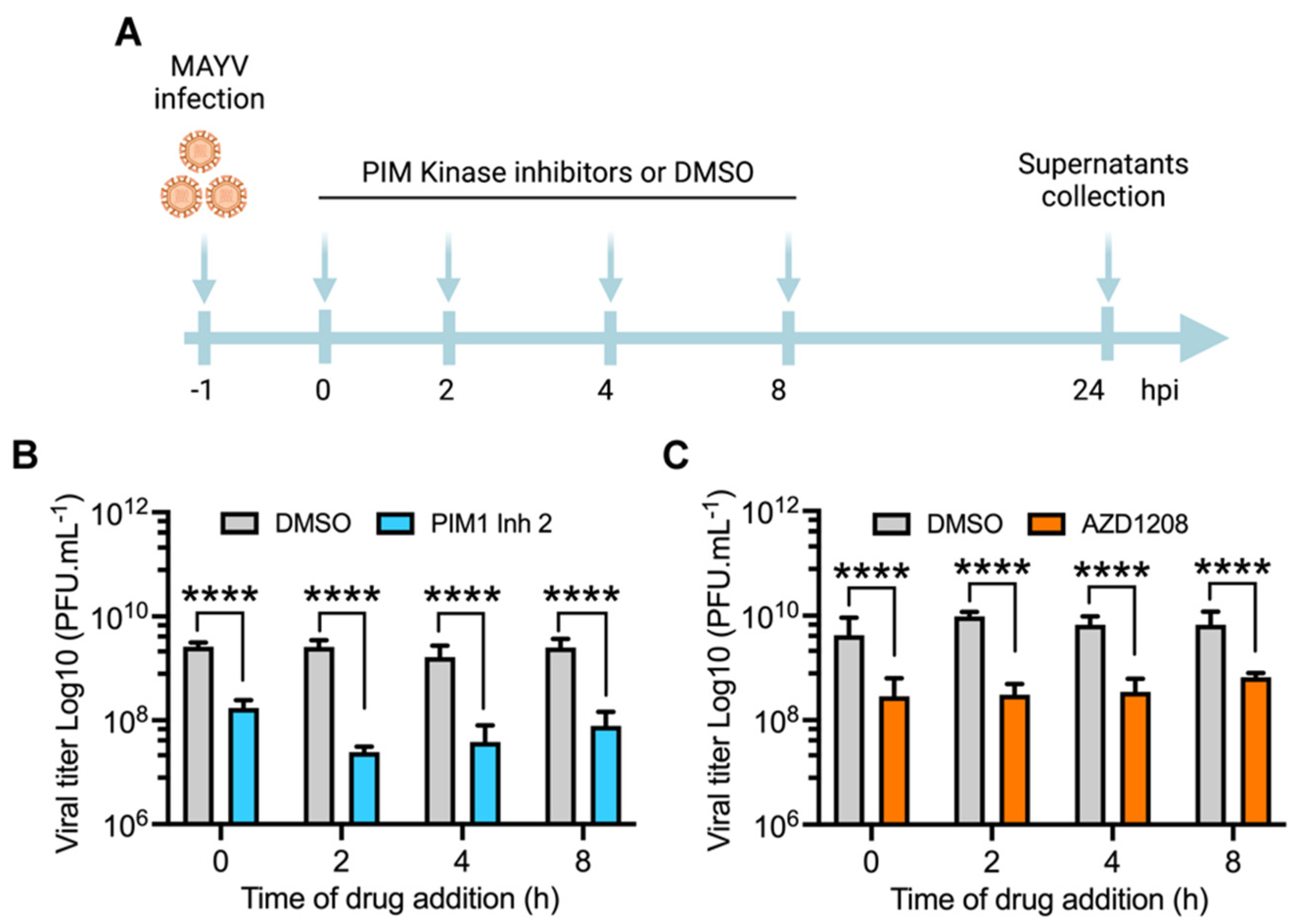
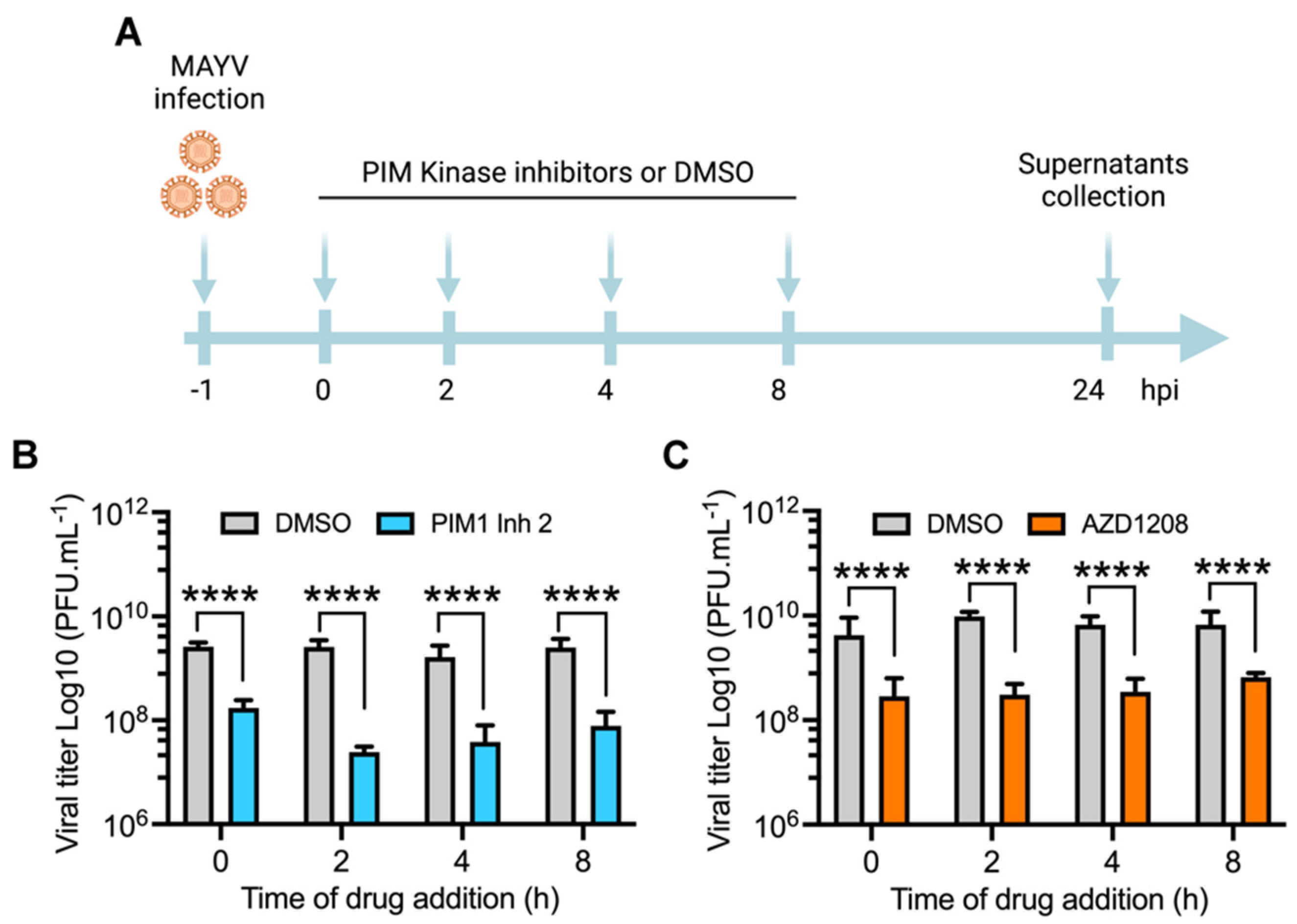
3.3. PIM Kinase Inhibitors Reduce MAYV Replication at Late Stages after Viral Adsorption
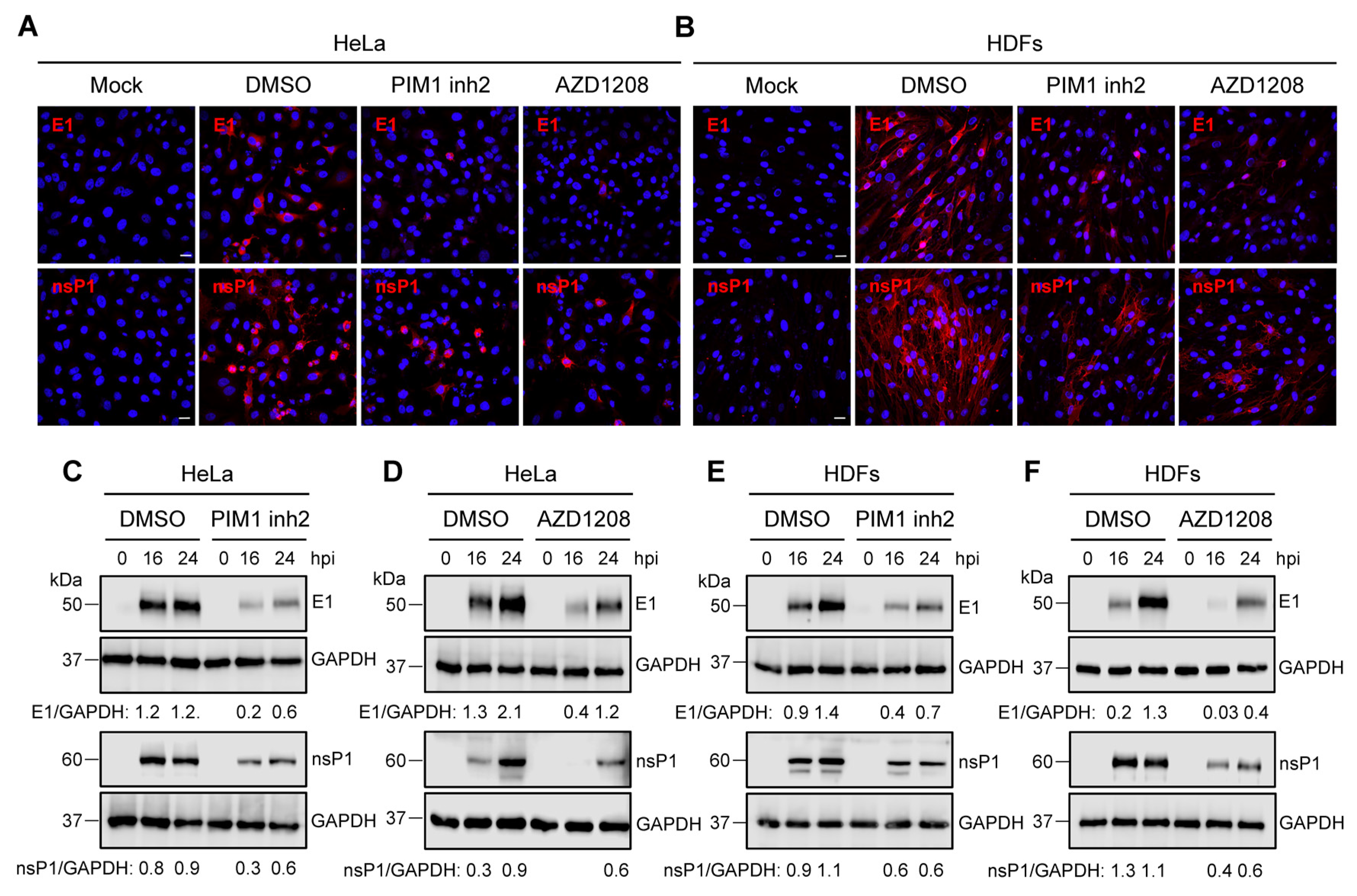
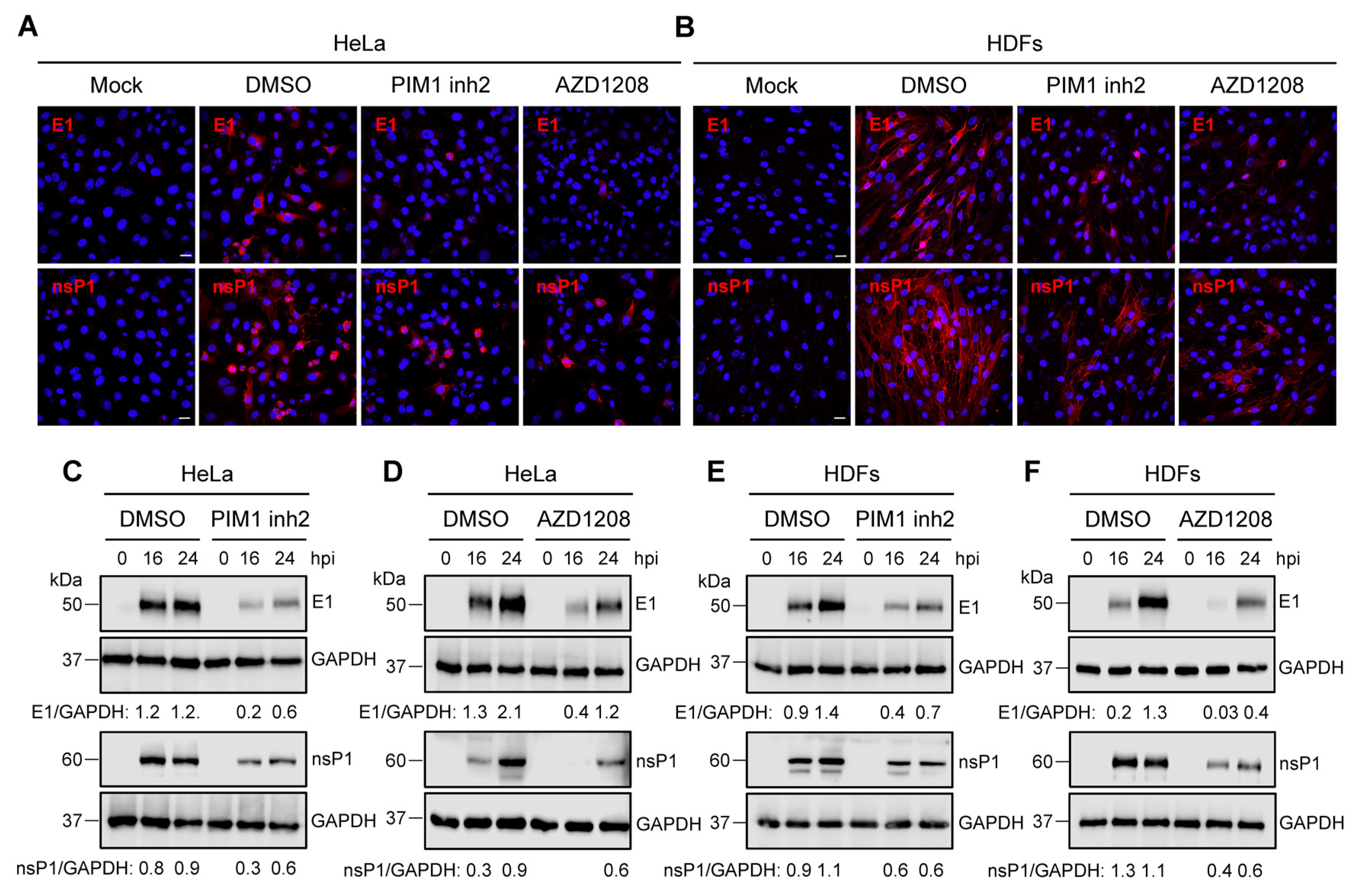
3.4. PIM Kinase Inhibitors Downmodulate the Expression of MAYV Strutural Protein E1 and Non-Structural Protein 1 (nsP1)
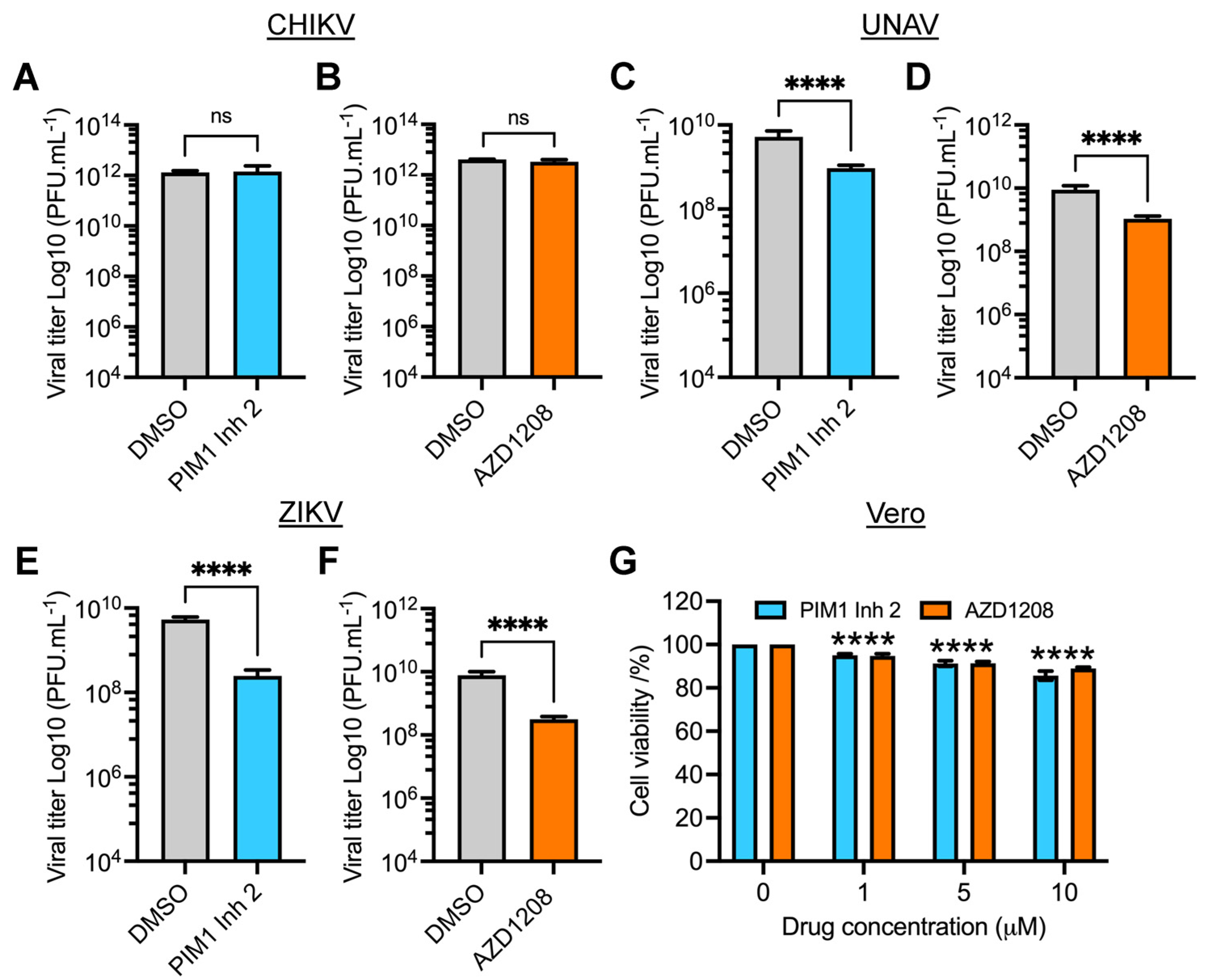
3.5. PIM Kinase Inhibitors Reduce Viral Replication of Una (UNAV) and Zika (ZIKV) Viruses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Acosta-Ampudia, Y.; Monsalve, D.M.; Rodriguez, Y.; Pacheco, Y.; Anaya, J.M.; Ramirez-Santana, C. Mayaro: An emerging viral threat? Emerg. Microbes Infect. 2018, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, M.L.; Figueiredo, L.T. Emerging alphaviruses in the Americas: Chikungunya and Mayaro. Rev. Soc. Bras. Med. Trop. 2014, 47, 677–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, L.I.; Vignuzzi, M. Arthritogenic Alphaviruses: A Worldwide Emerging Threat? Microorganisms 2019, 7, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaid, A.; Burt, F.J.; Liu, X.; Poo, Y.S.; Zandi, K.; Suhrbier, A.; Weaver, S.C.; Texeira, M.M.; Mahalingam, S. Arthritogenic alphaviruses: Epidemiological and clinical perspective on emerging arboviruses. Lancet Infect. Dis. 2021, 21, e123–e133. [Google Scholar] [CrossRef]
- Ganjian, N.; Riviere-Cinnamond, A. Mayaro virus in Latin America and the Caribbean. Rev. Panam. Salud Publica 2020, 44, e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzi, L.; Rodriguez-Morales, A.J.; Reusken, C.B.; Ribeiro, G.S.; LaBeaud, A.D.; Lourenco-de-Oliveira, R.; Brasil, P.; Lecuit, M.; Failloux, A.B.; Gallian, P.; et al. GloPID-R report on chikungunya, o’nyong-nyong and Mayaro virus, part 3: Epidemiological distribution of Mayaro virus. Antiviral. Res. 2019, 172, 104610. [Google Scholar] [CrossRef]
- Diagne, C.T.; Bengue, M.; Choumet, V.; Hamel, R.; Pompon, J.; Misse, D. Mayaro Virus Pathogenesis and Transmission Mechanisms. Pathogens 2020, 9, 738. [Google Scholar] [CrossRef] [PubMed]
- Dieme, C.; Ciota, A.T.; Kramer, L.D. Transmission potential of Mayaro virus by Aedes albopictus, and Anopheles quadrimaculatus from the USA. Parasit. Vect. 2020, 13, 613. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.N.; Carvalho, F.D.; De Mendonca, S.F.; Rocha, M.N.; Moreira, L.A. Vector competence of Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus mosquitoes for Mayaro virus. PLoS Negl. Trop. Dis. 2020, 14, e0007518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barroso, M.M.; Lima, C.S.; Silva-Neto, M.A.; Da Poian, A.T. Mayaro virus infection cycle relies on casein kinase 2 activity. Biochem. Biophys. Res. Commun. 2002, 296, 1334–1339. [Google Scholar] [CrossRef]
- Llamas-Gonzalez, Y.Y.; Campos, D.; Pascale, J.M.; Arbiza, J.; Gonzalez-Santamaria, J. A Functional Ubiquitin-Proteasome System is Required for Efficient Replication of New World Mayaro and Una Alphaviruses. Viruses 2019, 11, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauch, J.; Volinsky, N.; Romano, D.; Kolch, W. The secret life of kinases: Functions beyond catalysis. Cell Commun. Sign. 2011, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beziau, A.; Brand, D.; Piver, E. The Role of Phosphatidylinositol Phosphate Kinases during Viral Infection. Viruses 2020, 12, 1124. [Google Scholar] [CrossRef] [PubMed]
- Keck, F.; Ataey, P.; Amaya, M.; Bailey, C.; Narayanan, A. Phosphorylation of Single Stranded RNA Virus Proteins and Potential for Novel Therapeutic Strategies. Viruses 2015, 7, 5257–5273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keating, J.A.; Striker, R. Phosphorylation events during viral infections provide potential therapeutic targets. Rev. Med. Virol. 2012, 22, 166–181. [Google Scholar] [CrossRef] [PubMed]
- Sugasti-Salazar, M.; Llamas-Gonzalez, Y.Y.; Campos, D.; Gonzalez-Santamaria, J. Inhibition of p38 Mitogen-Activated Protein Kinase Impairs Mayaro Virus Replication in Human Dermal Fibroblasts and HeLa Cells. Viruses 2021, 13, 1156. [Google Scholar] [CrossRef] [PubMed]
- Santio, N.M.; Koskinen, P.J. PIM kinases: From survival factors to regulators of cell motility. Int. J. Biochem. Cell Biol. 2017, 93, 74–85. [Google Scholar] [CrossRef]
- Liu, Z.; Han, M.; Ding, K.; Fu, R. The role of Pim kinase in immunomodulation. Am. J. Cancer Res. 2020, 10, 4085–4097. [Google Scholar]
- Eerola, S.K.; Kohvakka, A.; Tammela, T.L.J.; Koskinen, P.J.; Latonen, L.; Visakorpi, T. Expression and ERG regulation of PIM kinases in prostate cancer. Cancer Med. 2021, 10, 3427–3436. [Google Scholar] [CrossRef]
- Wu, J.; Chu, E.; Kang, Y. PIM Kinases in Multiple Myeloma. Cancers 2021, 13, 4304. [Google Scholar] [CrossRef]
- Stafman, L.L.; Waldrop, M.G.; Williams, A.P.; Aye, J.M.; Stewart, J.E.; Mroczek-Musulman, E.; Yoon, K.J.; Whelan, K.; Beierle, E.A. The presence of PIM3 increases hepatoblastoma tumorigenesis and tumor initiating cell phenotype and is associated with decreased patient survival. J. Pediatr. Surg. 2019, 54, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, M.; Kundu, J.K.; Lee, M.H.; Liu, Z.Z. PIM Kinase as an Executional Target in Cancer. J. Cancer Prev. 2018, 23, 109–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, R.K.; Warfel, N.A. Targeting PIM Kinases to Overcome Therapeutic Resistance in Cancer. Mol. Cancer Ther. 2021, 20, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Cortes, J.; Tamura, K.; DeAngelo, D.J.; de Bono, J.; Lorente, D.; Minden, M.; Uy, G.L.; Kantarjian, H.; Chen, L.S.; Gandhi, V.; et al. Phase I studies of AZD1208, a proviral integration Moloney virus kinase inhibitor in solid and haematological cancers. Br. J. Cancer 2018, 118, 1425–1433. [Google Scholar] [CrossRef]
- Iida, S.; Sunami, K.; Minami, H.; Hatake, K.; Sekiguchi, R.; Natsume, K.; Ishikawa, N.; Rinne, M.; Taniwaki, M. A phase I, dose-escalation study of oral PIM447 in Japanese patients with relapsed and/or refractory multiple myeloma. Int. J. Hematol. 2021, 113, 797–806. [Google Scholar] [CrossRef]
- Rainio, E.M.; Ahlfors, H.; Carter, K.L.; Ruuska, M.; Matikainen, S.; Kieff, E.; Koskinen, P.J. Pim kinases are upregulated during Epstein-Barr virus infection and enhance EBNA2 activity. Virology 2005, 333, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Min, S.; Park, E.M.; Lim, Y.S.; Kang, S.; Suzuki, T.; Shin, E.C.; Hwang, S.B. Pim Kinase Interacts with Nonstructural 5A Protein and Regulates Hepatitis C Virus Entry. J. Virol. 2015, 89, 10073–10086. [Google Scholar] [CrossRef] [Green Version]
- de Vries, M.; Smithers, N.P.; Howarth, P.H.; Nawijn, M.C.; Davies, D.E. Inhibition of Pim1 kinase reduces viral replication in primary bronchial epithelial cells. Eur. Resp. J. 2015, 45, 1745–1748. [Google Scholar] [CrossRef] [Green Version]
- Pereiro, P.; Alvarez-Rodriguez, M.; Valenzuela-Munoz, V.; Gallardo-Escarate, C.; Figueras, A.; Novoa, B. RNA-Seq analysis reveals that spring viraemia of carp virus induces a broad spectrum of PIM kinases in zebrafish kidney that promote viral entry. Fish Shellfish Immunol. 2020, 99, 86–98. [Google Scholar] [CrossRef]
- Powers, A.M.; Aguilar, P.V.; Chandler, L.J.; Brault, A.C.; Meakins, T.A.; Watts, D.; Russell, K.L.; Olson, J.; Vasconcelos, P.F.; Da Rosa, A.T.; et al. Genetic relationships among Mayaro and Una viruses suggest distinct patterns of transmission. Am. J. Trop. Med. Hyg. 2006, 75, 461–469. [Google Scholar] [CrossRef]
- Causey, O.R.; Casals, J.; Shope, R.E.; Udomsakdi, S. Aura and Una, Two New Group a Arthropod-Borne Viruses. Am. J. Trop. Med. Hyg. 1963, 12, 777–781. [Google Scholar] [CrossRef] [PubMed]
- Carrera, J.P.; Diaz, Y.; Denis, B.; Barahona de Mosca, I.; Rodriguez, D.; Cedeno, I.; Arauz, D.; Gonzalez, P.; Cerezo, L.; Moreno, L.; et al. Unusual pattern of chikungunya virus epidemic in the Americas, the Panamanian experience. PLoS Negl. Trop. Dis. 2017, 11, e0005338. [Google Scholar] [CrossRef] [PubMed]
- Pierce, A.C.; Jacobs, M.; Stuver-Moody, C. Docking study yields four novel inhibitors of the protooncogene Pim-1 kinase. J. Med. Chem. 2008, 51, 1972–1975. [Google Scholar] [CrossRef] [PubMed]
- Dakin, L.A.; Block, M.H.; Chen, H.; Code, E.; Dowling, J.E.; Feng, X.; Ferguson, A.D.; Green, I.; Hird, A.W.; Howard, T.; et al. Discovery of novel benzylidene-1,3-thiazolidine-2,4-diones as potent and selective inhibitors of the PIM-1, PIM-2, and PIM-3 protein kinases. Bioorg. Med. Chem. Lett. 2012, 22, 4599–4604. [Google Scholar] [CrossRef]
- Pereira, T.N.; Virginio, F.; Souza, J.I.; Moreira, L.A. Emergent Arboviruses: A Review About Mayaro virus and Oropouche orthobunyavirus. Front. Trop. Dis. 2021, 2, 737436. [Google Scholar] [CrossRef]
- Zhou, F.; Wan, Q.; Chen, Y.; Chen, S.; He, M.L. PIM1 kinase facilitates Zika virus replication by suppressing host cells’ natural immunity. Sign. Transduct. Target Ther. 2021, 6, 207. [Google Scholar] [CrossRef]
- Keeton, E.K.; McEachern, K.; Dillman, K.S.; Palakurthi, S.; Cao, Y.; Grondine, M.R.; Kaur, S.; Wang, S.; Chen, Y.; Wu, A.; et al. AZD1208, a potent and selective pan-Pim kinase inhibitor, demonstrates efficacy in preclinical models of acute myeloid leukemia. Blood 2014, 123, 905–913. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugasti-Salazar, M.; Campos, D.; Valdés-Torres, P.; Galán-Jurado, P.E.; González-Santamaría, J. Targeting Host PIM Protein Kinases Reduces Mayaro Virus Replication. Viruses 2022, 14, 422. https://doi.org/10.3390/v14020422
Sugasti-Salazar M, Campos D, Valdés-Torres P, Galán-Jurado PE, González-Santamaría J. Targeting Host PIM Protein Kinases Reduces Mayaro Virus Replication. Viruses. 2022; 14(2):422. https://doi.org/10.3390/v14020422
Chicago/Turabian StyleSugasti-Salazar, Madelaine, Dalkiria Campos, Patricia Valdés-Torres, Paola Elaine Galán-Jurado, and José González-Santamaría. 2022. "Targeting Host PIM Protein Kinases Reduces Mayaro Virus Replication" Viruses 14, no. 2: 422. https://doi.org/10.3390/v14020422
APA StyleSugasti-Salazar, M., Campos, D., Valdés-Torres, P., Galán-Jurado, P. E., & González-Santamaría, J. (2022). Targeting Host PIM Protein Kinases Reduces Mayaro Virus Replication. Viruses, 14(2), 422. https://doi.org/10.3390/v14020422