Abstract
Persons who inject drugs (PWID) and their risk-related behaviors (e.g., unprotected sex and sharing needles/syringes/other injection equipment) have caused severe public health problems, especially in the rapid spread of HIV-1 and HCV. Here, we reconstructed the epidemic history of HIV-1 circulating recombinant form (CRF) 01_AE, CRF07_BC, and HCV subtype-6w among Taiwanese PWID. The timescales were estimated using phylogenetic and Bayesian coalescent analyses. The results revealed that CRF01_AE started to circulate in the Taiwanese PWID population in central Taiwan at 1992.5 (95% credible region: 1988.8–1995.9) and spread to other regions of Taiwan, while CRF07_BC was first identified in southern Taiwan at 2000.0 (95% CR: 1997.8–2002.2) and then spread northward to central-northern Taiwan. All HCV-6 strains were from Asia (that is, China, Myanmar, Taiwan, and Vietnam) and originated in 1928.1 (95% CR: 1890.2–1966.0). Furthermore, subtype-6w isolates from different regions of Taiwan appeared to share a common source that existed in the mid-1990s (95% CR: 1985.0–2001.8) or thereabouts. The routes of drug trafficking and the resulting high prevalence of HIV-1/HCV co-infections among PWID might have contributed to the virus transmission and promoted its spread worldwide. Long-term monitoring and policy implementation in at-risk populations would be useful for disease control.
1. Introduction
In 2021, 38.4 million people globally were estimated to have lived with HIV and the worst region is still sub-Saharan Africa [1]. A systematic review revealed that of the 16 million people who inject drugs (PWID), one-fourth lived in East and Southeast Asia [2]. PWID and their risk-related behaviors (e.g., unprotected sex and sharing syringes/heroin solutions/other injection equipment) have caused server public health problems, especially regarding the rapid spread of HIV-1 and HCV. According to the distribution data for the HIV-1 subtypes and the circulating recombinant forms (CRFs) in Asia, the HIV-1 subtype B’ and CRF01_AE are the main subtypes among PWID in Thailand [3,4,5], Myanmar [6], and Vietnam [7], while CRF07_BC is the predominant strain among PWID in China and Taiwan [8,9,10,11,12]. The strain CRF07_BC is derived from the Thai subtype B’ and Indian subtype C lineages [13]. Since 1997, CRF07_BC has been isolated from PWID in several provinces of China [8,9]. This strain is presumed to originate in Yunnan province [14,15] and has spread northwest to Xinjiang province and east to Guangxi province [13,16]. In this context, Yunnan plays an important role as an entry point for heroin trafficking into China [17] and is considered an epicenter of HIV/AIDS in China [13].
In Taiwan, most HIV-1 infected cases in the PWID were CRF07_BC [10,11,12]. Phylogenetic analyzes revealed that Taiwanese CRF07_BC strains clustered with CRF07_BC isolates from Xinjiang (97CN54, 97CN001, and 98CN009) and Guangxi (CNGL179) provinces of China [10,11,12]. Furthermore, it has been suggested that CRF07_BC initially circulated in southern Taiwan (Tainan) and then to other regions of Taiwan later [10,11]. Although predominate in PWID, CRF07_BC has also been spread to men who have sex with men (MSM) and heterosexuality [11]. Besides CRF07_BC, an outbreak of CRF01_AE infection was identified among PWID in Central-North Taiwan [10,11]. Evidence suggests that CRF01_AE was introduced into Taiwanese PWID through unprotected sex [11].
Both HIV-1 and HCV are considered blood-borne diseases, which are transmitted mainly through blood contact with infections. The prevalence of chronic hepatitis C in the Taiwanese population is approximately 2.1%, which was among the countries with the higher prevalence in Asia [18]. Paintsil et al. pointed out that HCV dried on inanimate surfaces can remain infectious for up to six weeks at normal room temperature [19]. Currently, the new HCV infection has shifted to PWID and other high-risk groups, including HIV-infected people and MSM who did not take effective safety measures. Traditional HCV treatment was a combination therapy (interferon injection plus oral ribavirin medicine), which successfully cured 70–80% of genotype 1 and >90% of genotype 2 patients [20,21,22,23,24]. With direct-acting antivirals (DAAs) that were launched in 2014, the successful cure rate increased to 95%.
During the explosive 2004–2006 outbreak of HIV-1 CRF07_BC among Taiwanese PWID, almost all of them (~99.3%) were HIV-1/HCV co-infections [10,11,25]. Previous studies on the geographical distribution of HCV showed that different dominant types existed throughout the world [26,27] and profound prevalence changes were observed in different genotypes of HCV over time [28,29]. Four predominant subtypes, namely 6a, 1a, 1b, and 3a, were identified in blood specimens from Taiwanese PWID infected with HCV. In particular, subtype-6w (1.4%) was detected at the same time in this subpopulation (see Table A1). Several studies have attempted to reconstruct the epidemic history of HIV-1 outbreaks among PWID in Asia [17,30,31]. In our previous study, we included HIV diagnosis when submitting virus nucleotide sequences to GenBank. To avoid errors in the calculation, we used the specimen collection date for all isolates to estimate the epidemic period. Furthermore, we focused only on PWID infected with HIV-1 CRF01_AE, CRF07_BC, and HCV-6. Since HCV subtype-6w is uncommon and no literature has yet described its transmission routes and studies in Taiwanese IDUs focused only on subtype-6w, we conducted an investigation of HCV to track its transmission routes of subtype-6w and estimate the time of emergence of genotype-6 among Asian PWID.
2. Materials and Methods
2.1. Subjects
The research procedures for the current study are shown in Figure A1. A total of 1427 PWID were recruited from Taipei City Hospital, Sindian Drug Abuse Treatment Center, Taipei Detention Center and Prison, Taoyuan Woman’s Prison (Northern Taiwan), Taichung Detention Center and Prison, Yunlin Second Prison, Nantou Detention Center (Central Taiwan), Tainan Detention Center, and Kaohsiung Prison (Southern Taiwan). Among the blood samples collected, 611 cases collected between 2004 and 2005 were shown to be infected with HIV-1 while 9 cases collected between 2005 and 2008 were HCV-6w. To track the routes of Taiwanese HIV-1 CRF01_AE, CRF07_BC, and HCV-6w transmission, we integrated sequences of Asian isolates available from the NCBI database (https://www.ncbi.nlm.nih.gov/nucleotide/, accessed on 8 July 2021) in our evolutionary analysis. All Taiwanese PWID were obtained by direct sequencing in our laboratory and those retrieved from GenBank were listed in Table A2.
Sociodemographic data and information on the types of illegal drugs used, history of drug abuse, risk factors associated with HIV-1 transmission, and years of the first HIV/HCV positive diagnosis were collected using a self-administered questionnaire. Peripheral blood samples were collected to allow analysis of virus genotype. Informed consent was obtained from all participants. Our research protocol was approved by the prisons and detentions administration system, as well as the Institutional Review Board of the National Yang-Ming University, Taiwan.
2.2. HIV-1 and HCV Subtyping
Viral RNA was extracted from plasma samples using the QIAamp Viral RNA mini kit (QIAGEN, Hilden, Germany). Random primer (Promega) was used in reverse transcription to generate cDNA for reverse transcriptase-polymerase chain reaction (RT-PCR). Anti-HCV antibodies from serum samples were detected using an enzyme immunoassay system (Murex 3nd, Abbott Laboratories, North Chicago, IL, USA). Specimens determined with anti-HCV antibodies or confirmed as HIV-1 positive were further analyzed. The genotypes/subtypes of HCV and HIV-1 infections were determined according to the methods described previously [10,25,32]. A set of primers, OF9-2 (forward) 5′-CGACATTACGCAGAAGTTGCCC-3′ and OR9 (reverse) 5′-AGTGTTGCTTAAGGCCTCCTGC-3′, were used to amplification of the HCV NS5B gene near full-length. Proviral nucleotide sequences were obtained by direct sequencing of PCR products using a DNA analyzer (ABI 3730, Applied Biosystems, Foster City, CA, USA).
2.3. Phylogenetic Analysis
Sequence alignment analysis with various reference strains from the Los Alamos HIV-1 database (https://www.hiv.lanl.gov/content/index, accessed on 28 May 2021) and the HCV database (https://hcv.lanl.gov/content/index, accessed on 8 July 2021) was performed using the BioEdit v7.2.6.1 program [33]. The MEGA X program [34] was used to find the best fit nucleotide substitution model and to construct phylogenetic trees using neighbor-joining (NJ) and maximum likelihood (ML) methods. For example, taking the HIV-1 env gene, the substitution model GTR + G was incorporated into the ML method, while TN93 + G (GTR and HKY models are not available here) was used to calculate the evolutionary distance for the NJ tree followed by bootstrap analysis with 1000 replicates [35]. Considering the best-fit models for the HCV NS5B gene, the substitution models for both the ML and NJ tree were K2 + G + I. At least four strains of all subtypes were used as reference sequences and isolates from Asian PWID were included for phylogenetic analysis (see Figure A2). Bootstrap values (≥70%) were used as an indicator of the significance of the clusters.
2.4. Nucleotide Sequence Accession Numbers
The HIV-1 env sequences (OM287868–OM287928) and the HCV NS5B sequences (OM287929–OM287937) were obtained from the current study and deposited in GenBank.
2.5. Bayesian Coalescent Inference
Evolutionary rates were obtained using the Bayesian Markov chain Monte Carlo (MCMC) approach implemented in BEAST v2.5.1 [36]. General time-reversible (GTR) [37,38,39] substitution models with gamma-distributed among-site rate variation involving six categories [40] were used to estimate evolutionary rates and construct tree topologies. Constantly sized, exponentially growing, and Bayesian skyline coalescent models were used for each case [41] and each MCMC chain was run for at least 10,000,000 states and sampled in every 1000 states. Posterior probability densities were calculated, and the convergence of the chains was verified using the Tracer v1.7.1 [42] with 10% of each chain discarded as burn-in.
2.6. Statistical Analysis
The Pearson χ2 test and Fisher’s exact test were performed in univariate analysis of demographic data. The difference between groups with a p-value < 0.05 was considered statistically significant. The p-values were two-tailed and unadjusted for multiple comparisons.
3. Results
3.1. Geographical Distribution of HIV-1 CRF01_AE, CRF07_BC, and HCV Subtype-6w among Taiwanese PWID
From 2004 to 2005, almost all Taiwanese HIV-1 positive PWID were infected with CRF07_BC. However, we found another small-scale outbreak strain that circulated in this population in central-north Taiwan. These were judged according to their time of crime and the place of sentences.
As shown in Figure A3, the distribution of Taiwanese PWID infected with HIV-1 CRF01_AE (n = 24), HIV-1 CRF07_BC (n = 982), and HCV subtype 6w (n = 9) during 2004–2008. The dates of HIV diagnosis and sample collection for most CRF01_AE infections were mainly in 2005 (Figure A2 and Table A2). The CRF07_BC sequences of Taiwanese PWID were grouped into several distinct phylogenetic clusters based on collection places [11] (details shown in Table A2). Based on the dates of HIV diagnosis, our data implied that CRF01_AE started to circulate in the Taiwanese PWID population in Central Taiwan and then spread to other regions of the island. In contrast, CRF07_BC first appeared in the south and moved northward to expand to central-north Taiwan (Figure A2 and Table 1).

Table 1.
Evolutionary characteristics of CRF01_AE and CRF07_BC.
The nine Taiwanese PWID infected with HCV-6w were identified when serving their prison sentences. Among them, five were HCV mono-infections and the other four cases were HIV/HCV coinfections. Six of the cases were from northern Taiwan (Taipei Detention Center, Sindian Drug Abuse Treatment Center and Taoyuan Woman’s Prison), two were from central Taiwan (Taichung Prison and Yunlin Second Prison), and one was from southern Taiwan (Kaohsiung Prison) (Figure A3).
3.2. An Estimated Timescale of the Spread of CRF01_AE and CRF07_BC among Taiwanese PWID
When estimating the time scale of the spread of HIV-1 CRF01_AE and CRF07_BC in Asia, we adopted the data set based on GTR + Γ6 constant model to pinpoint the time of the most recent common ancestor (tMRCA) of the HIV-1 strains circulating in this area. Compared to the results of the three models, similar conclusions could be reached (Table 1). After systematic analyses, we followed the likelihood of constant size, exponential growth, and the Bayesian skyline model (CRF01_AE: −15,559.7307, −15,548.8808, and −15,719.0827; CRF07_BC: −4939.2117, −4933.7994, and −4957.3370) and found that the exponential growth model was the best to present its transmission.
The estimated phylogeny using the env gene showed that all Taiwanese CRF01_AE and reference strains formed a single clade. However, there are two different risk groups in Taiwan [11], namely PWID and other sexual groups (e.g., homo, hetero, and bisexuals). The former population that contained sequences from Taipei Detention Center, Taipei Prison, Yunlin Second Prison, and Taichung Prison have a common point. Almost all samples were obtained in 2005 (Table S2). All CRF01_AE strains from Asia (i.e., China, Myanmar, Taiwan, and Vietnam) were dated 1979.0 (95% credible region, CR: 1973.1–1984.0). Concerned about drug addicts, CRF01_AE was introduced to China in 1986.5 (95% CR: 1980.0–1990.8) and then spread to Vietnam in 1988.4 (95% CR: 1984.7–1991.8), Taiwan in 1992, and Myanmar in 1994.6 (95% CR: 1990.2–1999.8). Additionally, CRF01_ AE was first introduced to other Taiwanese sexual groups in 1988.0 (95% CR: 1984.5–1991.1) and then spread to Central Taiwan in 1992.5 (95% CR: 1988.8–1995.9) and Northern Taiwan in 1994.1 (95% CR: 1989.5–1998.4) (Table 1 and Figure 1a). A comparison with our previous findings [11] suggested that CRF01_AE was introduced into Taiwanese PWID through unprotected sex and then caused a local epidemic among PWID through the exchange of injection equipment.
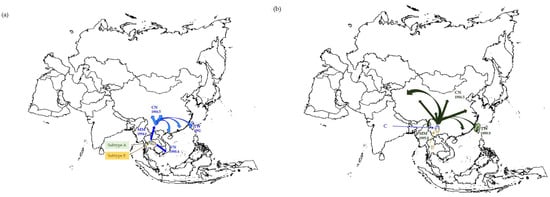
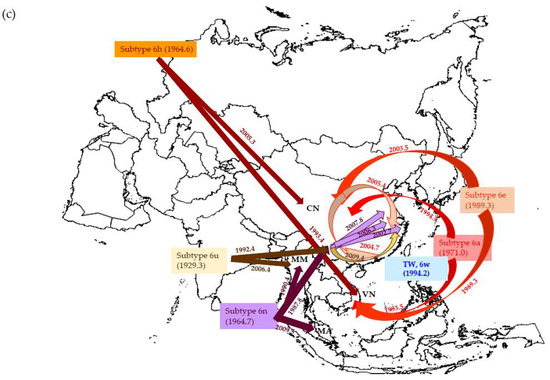
Figure 1.
The estimated timescale of the spread of (a) HIV-1 CRF01_AE, (b) CRF07_BC, and (c) HCV genotype-6 in Asian PWID population (CN: China, MM: Myanmar, MA: Malaysia, TW: Taiwan, and VN: Vietnam). In this particular figure, we adopted the data set based on GTR + Γ6/GTR + Γ6 + I constant model to show the tMRCAs of HIV-1 and HCV strains circulating in this area.
As summarized in Table 1, all CRF07_BC strains from Asia (i.e., China, Myanmar, and Taiwan) were rooted in 1987.9 (95% CR: 1981.0–1993.9). CRF07_ BC was introduced to China at 1987.9 and spread to Myanmar at 1995.0 (95% CR: 1987.6–2002.0) and to Taiwan at 1999.9 (95% CR: 1997.8–2001.9). Subsequently, this strain spread to other regions of Taiwan in 2001.3 (95% CR: 1998.6–2004.0), 2002.9 (95% CR: 2001.2–2004.2), and 2000.0 (95% CR: 1997.8–2002.2) in Northern, Central, and Southern Taiwan, respectively. The CRF07_BC strains from different regions of Taiwan seem to share a common source that existed in 2000 or thereabouts (95% CR: 1997.8–2001.9) and was part of the Southern Taiwan PWID. This suggests that southern Taiwan was the entry site for CRF07_ BC (Table 1 and Figure 1b).
3.3. An Estimated Timescale of the Spread of HCV Subtype-6w among Taiwanese PWID
Similarly to the estimation of the timescale of HIV-1 spread, we adopted the data set based on GTR + Γ6 + I constant model to pinpoint the tMRCAs of HCV-6 and found that the exponential growth model (likelihood in CS: −9744.679, EG: −9717.44, and BS: −9868.01) is the best way to present its transmission.
Phylogeny analysis using the NS5B gene showed that all Asian PWID and reference strains formed a single clade. As summarized in Table 2, all genotype-6 strains from Asia (i.e., China, Myanmar, Taiwan, and Vietnam) were rooted in 1928.1 (95% CR: 1890.2–1966.0). Subtypes 6a, -6n, and -6w had existed in the Taiwanese PWID population (Table A1). Taking subtype-6a for example, it was introduced into Vietnam at 1993.5 (95% CR: 1977.5–2001.3) and later into China at 1994.5 (95% CR: 1988.9–2000.9). Subtype-6n was initially introduced to China (1987.8, 1952.0–2005.0) and then spread to Myanmar (1990.4, 1954.7–2007.7). It is noteworthy that this strain was found to originate in Yunnan (1987.8, 1953.1–2007.0) and spread eastward to Suzhou, Zhenjiang, and Jiangsu in the early and mid-2000s. Furthermore, subtype-6w isolates from different regions in Taiwan seem to share a common source that existed in mid-1990 (95% CR: 1985.0–2001.8) or thereabouts (Table 2 and Figure 1c).
Table 2.
Evolutionary characteristics of HCV subtype 6w.
4. Discussion
Takebe et al. reported that CRF07_BC strains from different regions in China (including Xinjiang, Liaoning, and probably Guangdong and Sichuan) were likely to share a common ancestor that existed in Yunnan province around 1993 (95% CR: 1991.2–1995.2; gag) [13,31]. This suggests that CRF07_BC spreads almost simultaneously to various regions of China [13,31]. Furthermore, CRF07_BC also spread to Taiwan from the South around 1999.7 (95% CR: 1998.2–2001.1; env) and spread to the central-north part of Taiwan in 2002.1 (95% CR: 2001.3–2002.9; env) [10,13,30], resulting in a major HIV epidemic among PWID in Taiwan [10,11,12,13,31]. The dissemination routes of CRF07_BC in China and Taiwan were those reported in previous studies [11,13,30,31]. To compare the main differences between the use of the date of sample collection versus the date of HIV-1 diagnosis to estimate the time of the emergence of the CRF01_AE and CRF07_BC strains, and to consolidate the integrity of our data, we added more sequences from West Taiwan (e.g., Sindian, Taoyuan, Taichung, Yunlin, and Kaohsiung) in the analysis. As we all know that CRF07_BC circulated in southern Taiwan first, even adding the sequences from the most south area (i.e., Kaohsiung), the tMRCAs of CF07_BC among Taiwanese PWID were behind the estimates as previously reported [13,30,31]. The data obtained using the date of sample collection are more accurate than those using the date of diagnosis. This finding showed why it is necessary to use the correct date to estimate the time of emergence of HIV-1 subtypes or CRFs. Furthermore, our results revealed that the estimated introduction time of CRF01_AE in Taiwan PWID (1992 later) was earlier than that of CRF07_BC (1999.9), and because of the less aggressiveness of CRF01_AE [43], it only caused a local epidemic initially.
The estimated prevalence of HCV-6 in some regions of Southeast Asia, especially among patients with PWID and major thalassemia, is as high as 50% [44]. Injecting drug abuse is possibly responsible for the high frequency of this genotype in certain parts of Asia. HCV-6 has considerable genetic diversity with 23 subtypes (a–w). HCV-6 infected with HCV-6 respond better to interferon-based therapy compared to genotype 1, although the clinical characteristics and side effect profiles in patients are similar between HCV-6 and other genotypes [44]. Our study showed that HCV-6 was as common as genotype 1 (34.7% vs. 43.5%, Table S1) in the Taiwanese PWID population. According to a large-scale survey on the seroprevalence of HCV in Taiwan [45], the prevalence of injecting drug abuse and incomplete disinfection of medical utensils would cause a small-scale outbreak of HCV in local areas. Furthermore, residents have a higher prevalence of HCV when they were born in an earlier cohort [45]. As shown in Table A1, a cross-sectional study with 624 PWID recruited in Taiwan was conducted in 2007–2008. The overall prevalence of HIV and HCV infection was 44.1% (275/624) and 80.4% (502/624), respectively. The prevalence of HCV mono-infection and HIV/HCV co-infection was 36.4% (227/624) and 44.1% (275/624), respectively. The issues of HCV prevention include the following: to prevent healthy people from being contaminated with infected blood and to avoid reinfection with HCV in cured cases. For those who have been cured and non-infected, regular screening tests are encouraged. Through a series of analyses, our findings appear to support the hypothesis that HCV-6 originated in Southeast Asia (Table 2). HCV-6 is highly divergent from other genotypes and has distinct genetic differences from other strains, suggesting that there may be unclassified subtypes in Asia. Therefore, the accumulation of such genetic heterogeneity suggests that this genotype has circulated, adapted, and evolved in this area for a long period.
There are several limitations to this study. First, all Asian isolates (e.g., China, Malaysia, Myanmar, and Vietnam) were restricted and obtained from the NCBI website. Second, some Asian isolates were excluded from the evolutionary analysis because the sequences were too short or contained missing sequences. Despite the limitations, this study sheds light on the routes of drug trafficking and the resulting high prevalence of HIV-1/HCV coinfections among PWID that could have contributed to regional and global transmissions. In conclusion, for the first time, we report ‘the time of emergence of common HCV and HIV-1 strains among Taiwanese PWID’ and provide a comprehensive profile suggesting the initial circulation of CRF07_BC in southern Taiwan before spreading to other regions of Taiwan. Furthermore, the importance of using the date of sample collection versus the date of HIV-1 diagnosis was also highlighted when estimating the time of the emergence of the CRF01_AE and CRF07_BC strains. Long-term monitoring and implementation in the population at risk would be useful for disease control.
Author Contributions
Conceptualization, methodology, validation, formal analysis, and writing—original draft preparation, Y.-J.C.; investigation, K.-F.H.; resources, H.-C.T. and Y.-H.L.; data curation and writing—review and editing, J.C.H., K.-F.H. and H.-F.L.; supervision and project administration, H.-F.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from VGH-NYMU research (VGHUST108-G3-2-2) and was a continuation of the previous two projects (DOH95-DC-1109 and DOH97-DC-1202).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board of National Yang-Ming University (940008R, 10 November 2005; 970021, 16 May 2008) and Taipei City Hospital Research Ethics Committee (TCHIRB-10808010, 12 December 2018) for studies involving humans.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study. Also written informed consent has been obtained from the patient(s) to publish this paper.
Data Availability Statement
The data presented in this study are available in [Section 2.4 and Table A2]. Part of this paper was presented at the 8th Second Member Conference and Academic Symposium held in Taiwan in 2020.
Acknowledgments
We thank the PWID who participated in this study, the peer educators and social workers from the AIDS Prevention and Research Center who helped collect the questionnaires, and the staff of the Genomic Research Center of the National Yang Ming Chiao Tung University, as well as the health clinics of the various prisons and detention centers for their administrative support and technical assistance. The Sequencing Core Facility is supported by the National Research Program for Genomic Medicine (NRPGM) of the National Science Council. Most importantly, we would like to thank Wing-Wai Wong, who provided clinical samples but unfortunately passed away in October 2018, for his contribution to this study.
Conflicts of Interest
All authors of this article do not have commercial or other associations that might pose a conflict of interest. In addition, the funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A

Figure A1.
Study flowchart. A total of 1427 Taiwanese PWID blood samples were collected from hospitals, detention centers, and prisons during 2004–2008. Four hundred sequences were used in phylogenetic and evolutionary analyzes.

Figure A2.
Phylogenetic analysis of HIV-1 CRF01_AE, CRF07_BC, and HCV sequences among Asian PWID. (a) CRF01_AE, the NJ tree was constructed using MEGA under the TN93 + G model with 1000 bootstrap replicates. (b) CRF07_BC, TN93 + G and (c) HCV, K2 + G. The red point indicates the Taiwanese PWID sequences.

Figure A3.
Distribution of Taiwanese PWID infected with HIV-1 CRF01_AE (n = 24), HIV-1 CRF07_BC (n = 982), and HCV subtype 6w (n = 9) during 2004–2008. Each strain was labeled with a different color to denote patient characteristics, including collection place and year of HIV-1 diagnosis. The proportion of viral strains circulated into Northern-Central-Southern Taiwan per year, divided by total cases infected with the same strain, is shown in pie charts. Because Taiwanese PWID infected with HCV-6w were rare, we directly labeled their locations on the map.
Table A1.
Distribution of HCV genotypes/subtypes in the different groups among Taiwanese PWID.
Table A1.
Distribution of HCV genotypes/subtypes in the different groups among Taiwanese PWID.
| 2004–2006 HIV-1/HCV Co-Infection (n = 165) a | 2007–2008 HIV-1/HCV Co-Infection (n = 275) b | 2007–2008 HCV Mono-Infection (n = 227) b | p-Value | |
|---|---|---|---|---|
| Single infection | <0.001 | |||
| 1a | 21 (12.7) | 64 (23.3) | 53 (23.3) | |
| 1b | 48 (29.1) | 25 (9.1) | 30 (13.2) | |
| 2a | 6 (3.6) | 10 (3.6) | 6 (2.6) | |
| 2b | 6 (3.6) | 21 (7.6) | 5 (2.2) | |
| 3a | 11 (6.7) | 24 (8.7) | 25 (11.0) | |
| 3b | 1 (0.6) | 3 (1.1) | 3 (1.3) | |
| 6a | 43 (26.1) | 80 (29.1) | 53 (23.3) | |
| 6n | 4 (2.4) | 1 (0.4) | 3 (1.3) | |
| 6w | 1 (0.6) | 2 (0.7) | 5 (2.2) | |
| Subtotal_1 | 141 (85.5) | 230 (83.6) | 183 (80.6) | |
| Double infection | <0.001 | |||
| 1a/1b | 5 (3.0) | 5 (1.8) | 0 (0.0) | |
| 1a/2a | 0 (0.0) | 4 (1.5) | 1 (0.4) | |
| 1a/2b | 0 (0.0) | 18 (6.5) | 5 (2.2) | |
| 1a/3a | 0 (0.0) | 1 (0.4) | 2 (0.9) | |
| 1a/6a | 1 (0.6) | 0 (0.0) | 1 (0.4) | |
| 1b/2a | 1 (0.6) | 0 (0.0) | 0 (0.0) | |
| 1b/2b | 1 (0.6) | 0 (0.0) | 0 (0.0) | |
| 1b/3a | 3 (1.8) | 0 (0.0) | 0 (0.0) | |
| 1b/3b | 0 (0.0) | 1 (0.4) | 0 (0.0) | |
| 1b/6a | 6 (3.6) | 4 (1.5) | 6 (2.6) | |
| 1b/6w | 1 (0.6) | 0 (0.0) | 0 (0.0) | |
| 2a/2b | 0 (0.0) | 1 (0.4) | 1 (0.4) | |
| 2a/3a | 0 (0.0) | 0 (0.0) | 3 (1.3) | |
| 2a/6a | 0 (0.0) | 8 (2.9) | 20 (8.8) | |
| 2b/6a | 0 (0.0) | 2 (0.7) | 1 (0.4) | |
| 3a/6a | 3 (1.8) | 1 (0.4) | 2 (0.9) | |
| 6a/6n | 1 (0.6) | 0 (0.0) | 0 (0.0) | |
| Subtotal_2 | 22 (13.3) | 45 (16.4) | 42 (18.5) | |
| Triple infection | 0.261 | |||
| 1a/1b/3a | 1 (0.6) | 0 (0.0) | 0 (0.0) | |
| 1a/1b/6a | 1 (0.6) | 0 (0.0) | 0 (0.0) | |
| 1a/2a/3a | 0 (0.0) | 0 (0.0) | 1 (0.4) | |
| 1b/3a/6a | 0 (0.0) | 0 (0.0) | 1 (0.4) | |
| Subtotal_3 | 2 (1.2) | 0 (0.0) | 2 (0.9) |
a 180 blood samples randomly selected by the following strains (50 HIV-1 CRF07_BC-infected cases each from northern, central, and southern regions of Taiwan; 20 HIV-1 subtype B-infected cases; 10 HIV-1 CRF01_AE-infected cases) in years 2004–2006, among which 91.7% (165/180) were completed genotype identified by RT-PCR and phylogenetic analysis. b A total of 624 blood specimens were collected in the years 2007–2008; among them, 80.4% (502/624) were completed genotype identified by RT-PCR and phylogenetic analysis.
Table A2.
Location and collection date of Asian isolates from PWID infected with HIV-1 CRF01_AE, CRF07_BC, and HCV-6 and other related reference strains from GenBank.
Table A2.
Location and collection date of Asian isolates from PWID infected with HIV-1 CRF01_AE, CRF07_BC, and HCV-6 and other related reference strains from GenBank.
| Infections | Accession Numbers/Sample ID | Location | Collection Date |
|---|---|---|---|
| HIV-1 CRF01_AE (reference strains) | 90CF11697/F197340 | Central African Republic | 1990 |
| 90CF4071/AF197341 | Central African Republic | 1990 | |
| 90CR402/U51188 | Central African Republic | 1990 | |
| HIV-1 CRF01_AE a (Asian PWID) | HM215409 | Yunnan | 2007 |
| HM215410 | Yunnan | 2006 | |
| HM215418 | Yunnan | 2007 | |
| HM215419 | Yunnan | 2007 | |
| HM215422 | Yunnan | 2006 | |
| HM215427 | Yunnan | 2006 | |
| JN223054 | Myanmar | 2009 | |
| JX112823 | Guangdong | 8 Nov 2007 | |
| JX112824 | Guangdong | 23 Jul 2007 | |
| JX112825 | Guangdong | 23 Jul 2007 | |
| JX112826 | Guangdong | 4 Jul 2007 | |
| JX112828 | Guangdong | 11 Jul 2007 | |
| JX112831 | Guangxi | 22 May 2007 | |
| JX112837 | Guangxi | 22 Oct 2007 | |
| JX112839 | Guangxi | 8 Nov 2007 | |
| JX112840 | Guizhou | 19 Jul 2007 | |
| JX112842 | Guizhou | 6 Sep 2007 | |
| JX112857 | Sichuan | 3 Dec 2006 | |
| JX112861 | Yunnan | 4 Jun 2002 | |
| JX112863 | Yunnan | 4 Jun 2002 | |
| KP401977 | Viet Nam | 6 Jan 2012 | |
| KP401980 | Viet Nam | 9 Jan 2012 | |
| KP401983 | Viet Nam | 9 Jan 2012 | |
| KP401984 | Viet Nam | 9 Jan 2012 | |
| KP401987 | Viet Nam | 10 Jan 2012 | |
| KP401992 | Viet Nam | 11 Jan 2012 | |
| KP401995 | Viet Nam | 12 Jan 2012 | |
| KP401997 | Viet Nam | 11 Jan 2012 | |
| KP401998 | Viet Nam | 7 Feb 2012 | |
| KP402000 | Viet Nam | 8 Feb 2012 | |
| KP402001 | Viet Nam | 8 Feb 2012 | |
| KP402003 | Viet Nam | 8 Feb 2012 | |
| KP402004 | Viet Nam | 8 Feb 2012 | |
| KP402005 | Viet Nam | 9 Feb 2012 | |
| KP402009 | Viet Nam | 13 Feb 2012 | |
| KP402010 | Viet Nam | 13 Feb 2012 | |
| KP402014 | Viet Nam | 15 Feb 2012 | |
| KP402015 | Viet Nam | 16 Feb 2012 | |
| KP402017 | Viet Nam | 16 Feb 2012 | |
| KP402019 | Viet Nam | 20 Feb 2012 | |
| KP402020 | Viet Nam | 20 Feb 2012 | |
| KP402021 | Viet Nam | 20 Feb 2012 | |
| KP402022 | Viet Nam | 21 Feb 2012 | |
| KP402024 | Viet Nam | 21 Feb 2012 | |
| KP402026 | Viet Nam | 21 Feb 2012 | |
| KP402027 | Viet Nam | 22 Feb 2012 | |
| KP402030 | Viet Nam | 22 Feb 2012 | |
| HIV-1 CRF01_AE a (Asian PWID) | KP402031 | Viet Nam | 22 Feb 2012 |
| KP402037 | Viet Nam | 28 Feb 2012 | |
| KP402039 | Viet Nam | 9 Mar 2012 | |
| KP402040 | Viet Nam | 9 Mar 2012 | |
| KP402043 | Viet Nam | 14 Mar 2012 | |
| KP402044 | Viet Nam | 15 Mar 2012 | |
| KP402045 | Viet Nam | 28 Mar 2012 | |
| KP402046 | Viet Nam | 29 Mar 2012 | |
| KP402050 | Viet Nam | 16 Apr 2012 | |
| KP402051 | Viet Nam | 10 May 2012 | |
| KP402052 | Viet Nam | 15 May 2012 | |
| KP402056 | Viet Nam | 29 Feb 2012 | |
| KU820849 | Myanmar | 8 Nov 2014 | |
| Group 1 (Taiwanese PWID only) | D212 | Taipei Detention Center | 4 May 2005 |
| D261 | Taipei Prison | 18 May 2005 | |
| D291 | Yunlin Second Prison | 9 Jun 2005 | |
| D299 | Yunlin Second Prison | 9 Jun 2005 | |
| D301 | Yunlin Second Prison | 9 Jun 2005 | |
| D308 | Yunlin Second Prison | 9 Jun 2005 | |
| D336 | Yunlin Second Prison | 31 Aug 2005 | |
| D370 | Yunlin Second Prison | 31 Aug 2005 | |
| D376 | Yunlin Second Prison | 31 Aug 2005 | |
| D412 | Taipei Detention Center | 6 Oct 2005 | |
| D423 | Taipei Detention Center | 6 Oct 2005 | |
| D455 | Taipei Detention Center | 6 Oct 2005 | |
| D568 | Yunlin Second Prison | 22 Dec 2005 | |
| D580 | Yunlin Second Prison | 22 Dec 2005 | |
| D737 | Taichung Prison | 26 Mar 2008 | |
| Group 2 (Taiwanese other sexual-groups, e.g., homo-, hetero-, and bisexuals) | 4V370 | Taipei City Hospital | 7 Jan 2005 |
| 4V396 | Taipei City Hospital | 14 Feb 2005 | |
| 4V428 | Taipei City Hospital | 17 Mar 2005 | |
| 4V481 | Taipei City Hospital | 30 May 2005 | |
| 4V503 | Taipei City Hospital | 14 Jul 2005 | |
| 4V507 | Taipei City Hospital | 20 Jul 2005 | |
| 4V545 | Taipei City Hospital | 13 Sep 2005 | |
| 4V569 | Taipei City Hospital | 4 Oct 2005 | |
| 4V590 | Taipei City Hospital | 28 Oct 2005 | |
| 4V614 | Taipei City Hospital | 11 Nov 2005 | |
| 4V680 | Taipei City Hospital | 3 Jan 2006 | |
| 4V689 | Taipei City Hospital | 4 Jan 2006 | |
| 4V691 | Taipei City Hospital | 12 Jan2006 | |
| 4V714 | Taipei City Hospital | 24 Feb 2006 | |
| 4V724 | Taipei City Hospital | 20 Feb 2006 | |
| 4V740 | Taipei City Hospital | 7 Mar 2006 | |
| 4V744 | Taipei City Hospital | 9 Mar 2006 | |
| 4V795 | Taipei City Hospital | 9 May 2006 | |
| 4V872 | Taipei City Hospital | 21 Aug 2006 | |
| HIV-1 CRF07_BC (reference strains) | AF286226/97CN001 | China | 1997 |
| /C54A | China | 1997 | |
| /C54D | China | 1997 | |
| /C54 | China | 1997 | |
| /CN54 | China | 1997 | |
| AF286230/98CN009 | China | 1998 | |
| AF503396/CNGL179 | China | - | |
| HIV-1 CRF07_BC a (Asian PWID) | JN223069 | Myanmar | 2009 |
| JX392363 | Gansu | 2002 | |
| JX392364 | Ningxia | 2002 | |
| JX392365 | Ningxia | 2002 | |
| JX392366 | Ningxia | 2002 | |
| JX392367 | Ningxia | 2005 | |
| JX392368 | Qinghai | 2006 | |
| JX392369 | Qinghai | 2005 | |
| JX392370 | Sichuan | 1999 | |
| JX392371 | Sichuan | 1998 | |
| JX392372 | Sichuan | 1999 | |
| JX392373 | Sichuan | 2003 | |
| JX392374 | Sichuan | 2002 | |
| JX392375 | Yunnan | 1996 | |
| JX392376 | Yunnan | 1997 | |
| JX392377 | Yunnan | 2001 | |
| JX392378 | Sichuan | 2006 | |
| JX392379 | Sichuan | 2006 | |
| JX392380 | Sichuan | 2006 | |
| JX392381 | Sichuan | 2006 | |
| JX392382 | Sichuan | 2006 | |
| JX392384 | Xinjiang | 2006 | |
| KF250368 | Guangdong | 2007 | |
| KF250369 | Guangdong | 2007 | |
| KF250370 | Guangdong | 2007 | |
| KF250371 | Guangdong | 2007 | |
| KF250375 | Ningxia | 2007 | |
| KF250376 | Sichuan | 2006 | |
| KF250377 | Xinjiang | 2007 | |
| KF250378 | Yunnan | 1996 | |
| KF250379 | Yunnan | 1996 | |
| KF250380 | Yunnan | 1996 | |
| KU820832 | Myanmar | 28 Nov 2013 | |
| Group 1 (Taiwanese PWID only) | D74 | Tainan Detention Center | 30 Dec 2004 |
| D75 | Tainan Detention Center | 30 Dec 2004 | |
| D76 | Tainan Detention Center | 30 Dec 2004 | |
| D97 | Tainan Detention Center | 30 Dec 2004 | |
| D114 | Tainan Detention Center | 30 Dec 2004 | |
| D118 | Tainan Detention Center | 30 Dec 2004 | |
| D120 | Tainan Detention Center | 30 Dec 2004 | |
| Group 2 (Taiwanese PWID only) | 4V780 | Taipei City Hospital | 27 Apr 2006 |
| 4V784 | Taipei City Hospital | 24 Apr 2006 | |
| 4V793 | Taipei City Hospital | 8 May 2006 | |
| 4V807 | Taipei City Hospital | 17 May 2006 | |
| 4V844 | Taipei City Hospital | 11 Jul 2006 | |
| D126 | Tainan Detention Center | 30 Dec 2004 | |
| D338 | Yunlin Second Prison | 31 Aug 2005 | |
| D734 | Taichung Prison | 26 Mar 2008 | |
| D757 | Taichung Prison | 26 Mar 2008 | |
| D776 | Taichung Prison | 26 Mar 2008 | |
| Group 3 (Taiwanese PWID only) | 4V457 | Taipei City Hospital | 27 Apr 2005 |
| 4V467 | Taipei City Hospital | 9 May 2005 | |
| D733 | Taichung Prison | 26 Mar 2008 | |
| D735 | Taichung Prison | 26 Mar 2008 | |
| D736 | Taichung Prison | 26 Mar 2008 | |
| D771 | Taichung Prison | 26 Mar 2008 | |
| D775 | Taichung Prison | 26 Mar 2008 | |
| D780 | Taichung Prison | 26 Mar 2008 | |
| D839 | Taichung Prison | 26 Mar 2008 | |
| D845 | Taichung Prison | 26 Mar 2008 | |
| D850 | Kaohsiung Prison | 9 Apr 2008 | |
| D863 | Kaohsiung Prison | 9 Apr 2008 | |
| D902 | Kaohsiung Prison | 9 Apr 2008 | |
| D907 | Kaohsiung Prison | 9 Apr 2008 | |
| D947 | Kaohsiung Prison | 9 Apr 2008 | |
| D966 | Taoyuan Woman’s Prison | 6 May 2008 | |
| D970 | Taoyuan Woman’s Prison | 6 May 2008 | |
| D1007 | Taoyuan Woman’s Prison | 6 May 2008 | |
| HCV-6 (reference strains) | DQ278892/GZ52557 | China | 2002 |
| KC191671/10MYKJ032 | Malaysia | 2010 | |
| Subtype 6a (reference strains) | HQ912954/PR58 | China | 2008 |
| HQ912955/PR144 | China | 2008 | |
| KC844037/ZS221 | China | 2009 | |
| KC844038/ZS674 | China | 2011 | |
| AY859526/6a33 | Hong Kong | 2004 | |
| AY973865/cs6a-16 | Hong Kong | 2004 | |
| AY973866/cs6a-18 | Hong Kong | 2004 | |
| DQ480512/6a77 | Hong Kong | 2004 | |
| DQ480513/6a35 | Hong Kong | 2004 | |
| DQ480514/6a63 | Hong Kong | 2004 | |
| DQ480515/6a64 | Hong Kong | 2004 | |
| DQ480516/6a61 | Hong Kong | 2004 | |
| DQ480517/6a73 | Hong Kong | 2004 | |
| DQ480518/6a65 | Hong Kong | 2004 | |
| DQ480519/6a66 | Hong Kong | 2004 | |
| DQ480520/6a67 | Hong Kong | 2004 | |
| DQ480521/6a69 | Hong Kong | 2004 | |
| DQ480522/6a72 | Hong Kong | 2004 | |
| DQ480523/6a62 | Hong Kong | 2004 | |
| DQ480524/6a74 | Hong Kong | 2004 | |
| Y12083/EUHK2 | Hong Kong | 1997 | |
| EU246930/D9 | Viet Nam | - | |
| Subtype 6a (Asian PWID) | JX102891 | Viet Nam | 22 Aug 2008 |
| JX102895 | Viet Nam | 30 Aug 2008 | |
| JX102896 | Viet Nam | 30 Aug 2008 | |
| JX102897 | Viet Nam | 30 Aug 2008 | |
| JX102908 | Viet Nam | 31 Aug 2008 | |
| JX102957 | Viet Nam | 21 May 2008 | |
| JX102963 | Viet Nam | 21 Feb 2008 | |
| JX102965 | Viet Nam | 21 May 2008 | |
| JX102973 | Viet Nam | 21 May 2008 | |
| JX102975 | Viet Nam | 29 May 2008 | |
| JX103004 | Viet Nam | 5 Sep 2008 | |
| JX103010 | Viet Nam | 15 Oct 2008 | |
| JX103038 | Viet Nam | 28 Jun 2009 | |
| JX103047 | Viet Nam | 1 Jul 2009 | |
| JX103051 | Viet Nam | 3 Jul 2009 | |
| JX103104 | Viet Nam | 7 Sep 2009 | |
| JX103106 | Viet Nam | 7 Sep 2009 | |
| JX103117 | Viet Nam | 9 Sep 2009 | |
| JF721080 | Guangdong | 15 Nov 2010 | |
| JF721081 | Guangdong | 15 Nov 2010 | |
| JF721082 | Guangdong | 15 Nov 2010 | |
| JF721083 | Guangdong | 15 Nov 2010 | |
| JF721084 | Guangdong | 15 Nov 2010 | |
| JF721085 | Guangdong | 15 Nov 2010 | |
| JF721086 | Guangdong | 15 Nov 2010 | |
| Subtype 6a (Asian PWID) | JF721087 | Guangdong | 15 Nov 2010 |
| JF721088 | Guangdong | 15 Nov 2010 | |
| JF721089 | Guangdong | 15 Nov 2010 | |
| JF721090 | Guangdong | 15 Nov 2010 | |
| JF721091 | Guangdong | 15 Nov 2010 | |
| JF721092 | Guangdong | 15 Nov 2010 | |
| JF721093 | Guangdong | 15 Nov 2010 | |
| JF721094 | Guangdong | 15 Nov 2010 | |
| JF721095 | Guangdong | 15 Nov 2010 | |
| JF721096 | Guangdong | 15 Nov 2010 | |
| JF721097 | Guangdong | 15 Nov 2010 | |
| JF721098 | Guangdong | 15 Nov 2010 | |
| JF721099 | Guangdong | 15 Nov 2010 | |
| JF721100 | Guangdong | 15 Nov 2010 | |
| JF721101 | Guangdong | 15 Nov 2010 | |
| JF721102 | Guangdong | 15 Nov 2010 | |
| JF721103 | Guangdong | 15 Nov 2010 | |
| JF721104 | Guangdong | 15 Nov 2010 | |
| JF721105 | Guangdong | 15 Nov 2010 | |
| JF721106 | Guangdong | 15 Nov 2010 | |
| JF721107 | Guangdong | 15 Nov 2010 | |
| JF721108 | Guangdong | 15 Nov 2010 | |
| JF721109 | Guangdong | 15 Nov 2010 | |
| JF721110 | Guangdong | 15 Nov 2010 | |
| JF721111 | Guangdong | 15 Nov 2010 | |
| JF721112 | Guangdong | 15 Nov 2010 | |
| JF721113 | Guangdong | 15 Nov 2010 | |
| JF721114 | Guangdong | 15 Nov 2010 | |
| JF721115 | Guangdong | 15 Nov 2010 | |
| JF721116 | Guangdong | 15 Nov 2010 | |
| JF721117 | Guangdong | 15 Nov 2010 | |
| JF721118 | Guangdong | 15 Nov 2010 | |
| JF721119 | Guangdong | 15 Nov 2010 | |
| JF721120 | Guangdong | 15 Nov 2010 | |
| JF721121 | Guangdong | 15 Nov 2010 | |
| JF721122 | Guangdong | 15 Nov 2010 | |
| JF721123 | Guangdong | 15 Nov 2010 | |
| JF721124 | Guangdong | 15 Nov 2010 | |
| JF721125 | Guangdong | 15 Nov 2010 | |
| JF721126 | Guangdong | 15 Nov 2010 | |
| JF721127 | Guangdong | 15 Nov 2010 | |
| JF721128 | Guangdong | 15 Nov 2010 | |
| JF721129 | Guangdong | 15 Nov 2010 | |
| JF721130 | Guangdong | 15 Nov 2010 | |
| JF721131 | Guangdong | 15 Nov 2010 | |
| JF721132 | Guangdong | 15 Nov 2010 | |
| JF721133 | Guangdong | 15 Nov 2010 | |
| JF721134 | Guangdong | 15 Nov 2010 | |
| JF721135 | Guangdong | 15 Nov 2010 | |
| JF721136 | Guangdong | 15 Nov 2010 | |
| Subtype 6a (Asian PWID) | JF721137 | Guangdong | 15 Nov 2010 |
| JF721138 | Guangdong | 15 Nov 2010 | |
| JF721139 | Guangdong | 15 Nov 2010 | |
| JF721140 | Guangdong | 15 Nov 2010 | |
| JF721141 | Guangdong | 15 Nov 2010 | |
| JF721142 | Guangdong | 15 Nov 2010 | |
| JF721143 | Guangdong | 15 Nov 2010 | |
| JF721144 | Guangdong | 15 Nov 2010 | |
| JF721145 | Guangdong | 15 Nov 2010 | |
| JF721146 | Guangdong | 15 Nov 2010 | |
| JF721147 | Guangdong | 15 Nov 2010 | |
| JF721148 | Guangdong | 15 Nov 2010 | |
| JF721149 | Guangdong | 15 Nov 2010 | |
| JF721150 | Guangdong | 15 Nov 2010 | |
| JF721151 | Guangdong | 15 Nov 2010 | |
| JF721152 | Guangdong | 15 Nov 2010 | |
| JF721153 | Guangdong | 15 Nov 2010 | |
| JF721154 | Guangdong | 15 Nov 2010 | |
| JF721155 | Guangdong | 15 Nov 2010 | |
| JF721156 | Guangdong | 15 Nov 2010 | |
| JF721157 | Guangdong | 15 Nov 2010 | |
| Subtype 6e (reference strains) | DQ314805/GX004 | China | - |
| LC435023/N12-2804-Cam | Cambodia | 22 Aug 2012 | |
| LC435027/N16-2804-Cam | Cambodia | 3 Sep 2016 | |
| EU246931/D42 | Viet Nam | - | |
| EU246932/D88 | Viet Nam | - | |
| EU408326/537798 | USA | - | |
| Subtype 6e (Asian PWID) | AB523168 | Viet Nam | 2007 |
| AB523178 | Viet Nam | 2007 | |
| AB523179 | Viet Nam | 2007 | |
| AB523190 | Viet Nam | 2007 | |
| AB523248 | Viet Nam | 2007 | |
| AB523321 | Viet Nam | 2007 | |
| AB523326 | Viet Nam | 2007 | |
| AB523330 | Viet Nam | 2007 | |
| AB523339 | Viet Nam | 2007 | |
| AB523345 | Viet Nam | 2007 | |
| AB523346 | Viet Nam | 2007 | |
| HM009307 | Hong Kong | Jan 2006 | |
| HQ318922 | Zhenjiang | 2009 | |
| HQ318923 | Zhenjiang | 2009 | |
| HQ318924 | Zhenjiang | 2009 | |
| HQ318925 | Zhenjiang | 2009 | |
| HQ318926 | Zhenjiang | 2009 | |
| HQ318927 | Zhenjiang | 2009 | |
| JX102902 | Viet Nam | 30 Aug 2008 | |
| JX102969 | Viet Nam | 21 May 2008 | |
| JX102982 | Viet Nam | 29 May 2008 | |
| JX103102 | Viet Nam | 10 Jul 2009 | |
| JX103119 | Viet Nam | 10 Sep 2009 | |
| Subtype 6h (reference strains) | D84265/VN004 | Viet Nam | - |
| Subtype 6h (Asian PWID) | AB523188 | Viet Nam | 2007 |
| AB523257 | Viet Nam | 2007 | |
| AB523259 | Viet Nam | 2007 | |
| AB523289 | Viet Nam | 2007 | |
| AB523290 | Viet Nam | 2007 | |
| AB523312 | Viet Nam | 2007 | |
| AB523332 | Viet Nam | 2007 | |
| HM009308 | Hong Kong | 2006 | |
| JX102967 | Viet Nam | 21 May 2008 | |
| JX103044 | Viet Nam | 1 Jul 2009 | |
| Subtype 6n (reference strains) | DQ278894/KM42 | China | 2002 |
| DQ835768/D86/93 | Thailand | - | |
| EU246937/TH22 | Thailand | - | |
| EU246938/TH31 | Thailand | - | |
| Subtype 6n (Asian PWID) | HQ318920 | Zhenjiang | 2009 |
| HQ318921 | Zhenjiang | 2009 | |
| JQ303547 | Suzhou | 2011 | |
| JQ303548 | Suzhou | 2011 | |
| JQ303549 | Suzhou | 2011 | |
| JQ303550 | Suzhou | 2011 | |
| JQ303551 | Suzhou | 2010 | |
| JQ303552 | Suzhou | 2010 | |
| JQ303553 | Suzhou | 2011 | |
| JQ303554 | Suzhou | 2010 | |
| KC878938 | Jiangsu | 2011 | |
| KC878983 | Jiangsu | 2011 | |
| KM285079 | Yunnan | 2011 | |
| KM285080 | Yunnan | 2011 | |
| KM285081 | Yunnan | 2012 | |
| KM285082 | Yunnan | 2012 | |
| KM285083 | Yunnan | 2012 | |
| KM285084 | Yunnan | 2009 | |
| KM285085 | Yunnan | 2010 | |
| KM285086 | Yunnan | 2012 | |
| KM285087 | Yunnan | 2009 | |
| KM285088 | Yunnan | 2012 | |
| KM285089 | Yunnan | 2012 | |
| KM285090 | Yunnan | 2012 | |
| KM285091 | Yunnan | 2012 | |
| KM285092 | Yunnan | 2012 | |
| KM285093 | Yunnan | 2012 | |
| KM285094 | Yunnan | 2010 | |
| KM285095 | Yunnan | 2010 | |
| KM285096 | Yunnan | 2011 | |
| KM285097 | Yunnan | 2009 | |
| KM285098 | Yunnan | 2011 | |
| KR108496 | Malaysia | 2009 | |
| KR108497 | Malaysia | 2009 | |
| Subtype 6n (Asian PWID) | KT735662 | Yunnan | 2014 |
| KT735668 | Yunnan | 2014 | |
| KT735681 | Yunnan | 2014 | |
| KT735687 | Yunnan | 2014 | |
| KT735689 | Yunnan | 2014 | |
| KT735695 | Yunnan | 2014 | |
| KT735697 | Yunnan | 2014 | |
| KT735698 | Yunnan | 2014 | |
| KT735704 | Yunnan | 2014 | |
| KT735709 | Yunnan | 2014 | |
| KT735715 | Yunnan | 2014 | |
| KT735718 | Yunnan | 2014 | |
| KT735723 | Yunnan | 2014 | |
| KT735726 | Yunnan | 2014 | |
| KT735727 | Yunnan | 2014 | |
| KT735732 | Yunnan | 2014 | |
| KT735733 | Yunnan | 2014 | |
| KT735736 | Yunnan | 2014 | |
| KT735743 | Yunnan | 2014 | |
| KT735759 | Yunnan | 2014 | |
| KT735779 | Yunnan | 2014 | |
| KT735804 | Yunnan | 2014 | |
| KT735814 | Yunnan | 2014 | |
| KT735820 | Yunnan | 2014 | |
| KT735821 | Yunnan | 2014 | |
| KT735822 | Yunnan | 2014 | |
| KT735827 | Yunnan | 2014 | |
| KT735834 | Yunnan | 2014 | |
| KT735855 | Yunnan | 2014 | |
| KT735878 | Yunnan | 2014 | |
| MH458979 | Myanmar | 2014 | |
| MH458983 | Myanmar | 2014 | |
| MH458986 | Myanmar | 2014 | |
| MH458995 | Myanmar | 2014 | |
| Subtype 6r (reference strains) | EU408328/QC245 | Canada | - |
| LC435024/N12-2911-Cam | Cambodia | 22 Aug 2012 | |
| LC435028/N16-2911-Cam | Cambodia | 3 Sep 2016 | |
| Subtype 6v (reference strains) | EU158186/NK46 | China | 2004 |
| EU798760/KMN-02 | China | - | |
| EU798761/KM046 | China | - | |
| FJ435090/KM181 | China | - | |
| Subtype 6u (reference strains) | EU246940/D83 | Viet Nam | - |
| EU408330/DH012 | China | 2001 | |
| EU408331/DH014 | China | 2001 | |
| EU408332/DH028 | China | 2001 | |
| Subtype 6u (Asian PWID) | JQ303555 | Suzhou | 2011 |
| KM285099 | Yunnan | 2012 | |
| KM285100 | Yunnan | 2009 | |
| KM285101 | Yunnan | 2012 | |
| KM285102 | Yunnan | 2009 | |
| Subtype 6u (Asian PWID) | KM285103 | Yunnan | 2011 |
| KM285104 | Yunnan | 2009 | |
| KM285105 | Yunnan | 2011 | |
| KM285106 | Yunnan | 2011 | |
| KM285107 | Yunnan | 2011 | |
| KM285108 | Yunnan | 2011 | |
| KM285109 | Yunnan | 2009 | |
| KM285110 | Yunnan | 2012 | |
| KM285111 | Yunnan | 2011 | |
| KM285112 | Yunnan | 2011 | |
| KM285113 | Yunnan | 2010 | |
| KM285114 | Yunnan | 2009 | |
| KM285115 | Yunnan | 2012 | |
| KM285116 | Yunnan | 2012 | |
| KM285117 | Yunnan | 2011 | |
| KM285118 | Yunnan | 2009 | |
| KT735674 | Yunnan | 2014 | |
| KT735690 | Yunnan | 2014 | |
| KT735700 | Yunnan | 2014 | |
| KT735712 | Yunnan | 2014 | |
| KT735744 | Yunnan | 2014 | |
| KT735810 | Yunnan | 2014 | |
| MH458978 | Myanmar | 2014 | |
| MH458980 | Myanmar | 2014 | |
| MH458984 | Myanmar | 2014 | |
| MH458993 | Myanmar | 2014 | |
| Subtype 6w (reference strains) | EU643834/HCV-6-D140 | Taiwan | 2005 |
| EU643836/HCV-6-D370 | Taiwan | 2005 | |
| HCV subtype 6w (Taiwanese PWID only) | D140 | Taipei Detention Center | 20 Jan 2005 |
| D370 | Yunlin Second Prison | 31 Aug 2005 | |
| D778 | Taichung Prison | 26 Mar 2008 | |
| D866 | Kaohsiung Prison | 9 Apr 2008 | |
| ND141 | Taoyuan Woman’s Prison | 27 May 2008 | |
| ND153 | Taoyuan Woman’s Prison | 27 May 2008 | |
| ND187 | Taoyuan Woman’s Prison | 27 May 2008 | |
| ND414 | Sindian Drug Abuser Treatment Center | 18 Jun 2008 | |
| ND415 | Sindian Drug Abuser Treatment Center | 18 Jun 2008 |
Note: a The lengths of the env fragment in CRF01_AE and CRF07_BC were up to 537 and 552 bp, respectively. The length of the query shorter than the standard was labeled with a different color, such as ‘light gray, <500 bp’ and ‘dark gray, <300 bp’.
References
- UNAIDS. In Danger: UNAIDS Global AIDS Update 2022. 2022. Available online: https://www.unaids.org/en/resources/documents/2022/in-danger-global-aids-update (accessed on 16 September 2022).
- Mathers, B.M.; Degenhardt, L.; Phillips, B.; Wiessing, L.; Hickman, M.; Strathdee, S.A.; Wodak, A.; Panda, S.; Tyndall, M.; Toufik, A.; et al. Global epidemiology of injecting drug use and HIV among people who inject drugs: A systematic review. Lancet 2008, 372, 1733–1745. [Google Scholar] [CrossRef]
- Ou, C.Y.; Takebe, Y.; Weniger, B.G.; Luo, C.C.; Kalish, M.L.; Auwanit, W.; Yamazaki, S.; Gayle, H.D.; Young, N.L.; Schochetman, G. Independent introduction of two major HIV-1 genotypes into distinct high-risk populations in Thailand. Lancet 1993, 341, 1171–1174. [Google Scholar] [CrossRef]
- Kalish, M.L.; Baldwin, A.; Raktham, S.; Wasi, C.; Luo, C.C.; Schochetman, G.; Mastro, T.D.; Young, N.; Vanichseni, S.; Rubsamen-Waigmann, H.; et al. The evolving molecular epidemiology of HIV-1 envelope subtypes in injecting drug users in Bangkok, Thailand: Implications for HIV vaccine trials. AIDS 1995, 9, 851–857. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Zhang, C.; Duo, L.; Zhou, Y.H.; Yao, Z.H.; Liu, F.L.; Li, H.; Tu, Y.Q.; Zheng, Y.T. Extensive and complex HIV-1 recombination between B′, C and CRF01_AE among IDUs in south-east Asia. AIDS 2012, 26, 1121–1129. [Google Scholar] [CrossRef]
- Takebe, Y.; Motomura, K.; Tatsumi, M.; Lwin, H.H.; Zaw, M.; Kusagawa, S. High prevalence of diverse forms of HIV-1 intersubtype recombinants in Central Myanmar: Geographical hot spot of extensive recombination. AIDS 2003, 17, 2077–2087. [Google Scholar] [CrossRef]
- Kato, K.; Kusagawa, S.; Motomura, K.; Yang, R.; Shiino, T.; Nohtomi, K.; Sato, H.; Shibamura, K.; Nguyen, T.H.; Pham, K.C.; et al. Closely related HIV-1 CRF01_AE variant among injecting drug users in northern Vietnam: Evidence of HIV spread across the Vietnam-China border. AIDS Res. Hum. Retrovir. 2001, 17, 113–123. [Google Scholar] [CrossRef]
- Su, L.; Graf, M.; Zhang, Y.; von Briesen, H.; Xing, H.; Kostler, J.; Melzl, H.; Wolf, H.; Shao, Y.; Wagner, R. Characterization of a virtually full-length human immunodeficiency virus type 1 genome of a prevalent intersubtype (C/B’) recombinant strain in China. J. Virol. 2000, 74, 11367–11376. [Google Scholar] [CrossRef]
- Piyasirisilp, S.; McCutchan, F.E.; Carr, J.K.; Sanders-Buell, E.; Liu, W.; Chen, J.; Wagner, R.; Wolf, H.; Shao, Y.; Lai, S.; et al. A recent outbreak of human immunodeficiency virus type 1 infection in southern China was initiated by two highly homogeneous, geographically separated strains, circulating recombinant form AE and a novel BC recombinant. J. Virol. 2000, 74, 11286–11295. [Google Scholar] [CrossRef]
- Chen, Y.J.; Huang, Y.H.; Chuang, S.Y.; Kao, D.Y.; Lan, Y.C.; Yang, J.Y.; Chen, Y.M. Molecular epidemiology of HIV-1 subtype B, CRF01_AE, and CRF07_BC infection among injection drug users in Taiwan. J. Acquir. Immune Defic. Syndr. 2010, 53, 425–439. [Google Scholar] [CrossRef]
- Chen, Y.J.; Lee, C.M.; Chen, M.; Chuang, S.Y.; Liu, H.F.; Wong, W.W.; Lin, Y.H.; Tsai, H.C.; Wang, J.H.; Chen, Y.M. Molecular epidemiology of HIV-1 infection in Taiwan from 2005 to 2008: Further spread of CRF07_BC and emergence of CRF07_BC/subtype B dual infection. J. Acquir. Immune Defic. Syndr. 2012, 59, 438–446. [Google Scholar] [CrossRef]
- Lin, Y.T.; Lan, Y.C.; Chen, Y.J.; Huang, Y.H.; Lee, C.M.; Liu, T.T.; Wong, W.W.; Yang, J.Y.; Wang, C.T.; Chen, Y.M. Molecular epidemiology of HIV-1 infection and full-length genomic analysis of circulating recombinant form 07_BC strains from injection drug users in Taiwan. J. Infect. Dis. 2007, 195, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Takebe, Y.; Liao, H.; Hase, S.; Uenishi, R.; Li, Y.; Li, X.J.; Han, X.; Shang, H.; Kamarulzaman, A.; Yamamoto, N.; et al. Reconstructing the epidemic history of HIV-1 circulating recombinant forms CRF07_BC and CRF08_BC in East Asia: The relevance of genetic diversity and phylodynamics for vaccine strategies. Vaccine 2010, 28 (Suppl. 2), B39–B44. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Xia, X.; Kusagawa, S.; Zhang, C.; Ben, K.; Takebe, Y. On-going generation of multiple forms of HIV-1 intersubtype recombinants in the Yunnan Province of China. AIDS 2002, 16, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- McClutchan, F.E.; Carr, J.K.; Murphy, D.; Piyasirisilp, S.; Gao, F.; Hahn, B.; Yu, X.F.; Beyrer, C.; Birx, D.L. Precise mapping of recombination breakpoints suggests a common parent of two BC recombinant HIV type 1 strains circulating in China. AIDS Res. Hum. Retrovir. 2002, 18, 1135–1140. [Google Scholar] [CrossRef]
- Meng, Z.; Xin, R.; Zhong, P.; Zhang, C.; Abubakar, Y.F.; Li, J.; Liu, W.; Zhang, X.; Xu, J. A new migration map of HIV-1 CRF07_BC in China: Analysis of sequences from 12 provinces over a decade. PLoS ONE 2012, 7, e52373. [Google Scholar] [CrossRef]
- Beyrer, C.; Razak, M.H.; Lisam, K.; Chen, J.; Lui, W.; Yu, X.F. Overland heroin trafficking routes and HIV-1 spread in south and south-east Asia. AIDS 2000, 14, 75–83. [Google Scholar] [CrossRef]
- The Polaris Observatory HCV Collaborators. Global prevalence and genotype distribution of hepatitis C virus infection in 2015: A modelling study. Lancet Gastroenterol. Hepatol. 2017, 2, 161–176. [Google Scholar] [CrossRef]
- Paintsil, E.; Binka, M.; Patel, A.; Lindenbach, B.D.; Heimer, R. Hepatitis C virus maintains infectivity for weeks after drying on inanimate surfaces at room temperature: Implications for risks of transmission. J. Infect. Dis. 2014, 209, 1205–1211. [Google Scholar] [CrossRef]
- Chuang, W.L.; Yu, M.L.; Dai, C.Y.; Chang, W.Y. Treatment of chronic hepatitis C in southern Taiwan. Intervirology 2006, 49, 99–106. [Google Scholar] [CrossRef]
- Lee, S.D.; Yu, M.L.; Cheng, P.N.; Lai, M.Y.; Chao, Y.C.; Hwang, S.J.; Chang, W.Y.; Chang, T.T.; Hsieh, T.Y.; Liu, C.J.; et al. Comparison of a 6-month course peginterferon alpha-2b plus ribavirin and interferon alpha-2b plus ribavirin in treating Chinese patients with chronic hepatitis C in Taiwan. J. Viral Hepat. 2005, 12, 283–291. [Google Scholar] [CrossRef]
- Liu, C.H.; Liu, C.J.; Lin, C.L.; Liang, C.C.; Hsu, S.J.; Yang, S.S.; Hsu, C.S.; Tseng, T.C.; Wang, C.C.; Lai, M.Y.; et al. Pegylated interferon-alpha-2a plus ribavirin for treatment-naive Asian patients with hepatitis C virus genotype 1 infection: A multicenter, randomized controlled trial. Clin. Infect. Dis. 2008, 47, 1260–1269. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.L.; Dai, C.Y.; Huang, J.F.; Hou, N.J.; Lee, L.P.; Hsieh, M.Y.; Chiu, C.F.; Lin, Z.Y.; Chen, S.C.; Hsieh, M.Y.; et al. A randomised study of peginterferon and ribavirin for 16 versus 24 weeks in patients with genotype 2 chronic hepatitis C. Gut 2007, 56, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.L.; Dai, C.Y.; Huang, J.F.; Chiu, C.F.; Yang, Y.H.; Hou, N.J.; Lee, L.P.; Hsieh, M.Y.; Lin, Z.Y.; Chen, S.C.; et al. Rapid virological response and treatment duration for chronic hepatitis C genotype 1 patients: A randomized trial. Hepatology 2008, 47, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Lin, H.J.; Chen, Y.J.; Lee, C.M.; Wang, S.F.; Chang, K.Y.; Chen, T.L.; Liu, H.F.; Chen, Y.M. Molecular epidemiology of HCV genotypes among injection drug users in Taiwan: Full-length sequences of two new subtype 6w strains and a recombinant form_2b6w. J. Med. Virol. 2010, 82, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Humphreys, I.; Flaxman, A.; Brown, A.; Cooke, G.S.; Pybus, O.G.; Barnes, E. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology 2015, 61, 77–87. [Google Scholar] [CrossRef]
- Gower, E.; Estes, C.; Blach, S.; Razavi-Shearer, K.; Razavi, H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J. Hepatol. 2014, 61 (Suppl. S1), S45–S57. [Google Scholar] [CrossRef]
- Schroter, M.; Zollner, B.; Laufs, R.; Feucht, H.H. Changes in the prevalence of hepatitis C virus genotype among injection drug users: A highly dynamic process. J. Infect. Dis. 2004, 190, 1199–1200. [Google Scholar] [CrossRef]
- Sereno, S.; Perinelli, P.; Laghi, V. Changes in the prevalence of hepatitis C virus genotype among Italian injection drug users-relation to period of injection started. J. Clin. Virol. 2009, 45, 354–357. [Google Scholar] [CrossRef]
- Tee, K.K.; Pybus, O.G.; Liao, H.; Uenishi, R.; Hase, S.; Kamarulzaman, A.; Li, X.J.; Takebe, Y. Chronology of the HIV-1 CRF07_BC expansion in East Asia. AIDS 2008, 22, 156–158. [Google Scholar] [CrossRef]
- Tee, K.K.; Pybus, O.G.; Li, X.J.; Han, X.; Shang, H.; Kamarulzaman, A.; Takebe, Y. Temporal and spatial dynamics of human immunodeficiency virus type 1 circulating recombinant forms 08_BC and 07_BC in Asia. J. Virol. 2008, 82, 9206–9215. [Google Scholar] [CrossRef]
- Lee, Y.M.; Chen, Y.J.; Lee, C.M.; Kuo, L.H.; Wong, W.W.; Chen, Y.M. Detection of hepatitis C virus subtypes 6a, 6n, 6w and mixed infections using a modified multiplex real-time polymerase chain reaction protocol. J. Formos. Med. Assoc. 2011, 110, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. Available online: https://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=1383440 (accessed on 21 September 2022).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Using the quantitative genetic threshold model for inferences between and within species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1427–1434. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kuhnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Waddell, P.J.; Steel, M.A. General time-reversible distances with unequal rates across sites: Mixing gamma and inverse Gaussian distributions with invariant sites. Mol. Phylogenet. Evol. 1997, 8, 398–414. [Google Scholar] [CrossRef] [PubMed]
- Zwickl, D.; Holder, M. Model parameterization, prior distributions, and the general time-reversible model in Bayesian phylogenetics. Syst. Biol. 2004, 53, 877–888. [Google Scholar] [CrossRef]
- Tavaré, S. Some probabilistic and statistical problems in the analysis of DNA sequences. Lect. Math. Life Sci. 1986, 17, 57–86. [Google Scholar]
- Yang, Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: Approximate methods. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef]
- Drummond, A.J.; Nicholls, G.K.; Rodrigo, A.G.; Solomon, W. Estimating mutation parameters, population history and genealogy simultaneously from temporally spaced sequence data. Genetics 2002, 161, 1307–1320. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Tracer v1.7. 2018. Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 10 April 2018).
- Pant Pai, N.; Shivkumar, S.; Cajas, J.M. Does genetic diversity of HIV-1 non-B subtypes differentially impact disease progression in treatment-naive HIV-1-infected individuals? A systematic review of evidence: 1996–2010. J. Acquir. Immune Defic. Syndr. 2012, 59, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.T.; Abe, K.; Nguyen, M.H. Systematic review: Epidemiology of hepatitis C genotype 6 and its management. Aliment. Pharmacol. Ther. 2011, 34, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Yang, P.M.; Huang, G.T.; Lee, H.S.; Sung, J.L.; Sheu, J.C. Estimation of seroprevalence of hepatitis B virus and hepatitis C virus in Taiwan from a large-scale survey of free hepatitis screening participants. J. Formos. Med. Assoc. 2007, 106, 148–155. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).